Histone 2-Hydroxyisobutyryltransferase Encoded by Afngg1 Is Involved in Pathogenicity and Aflatoxin Biosynthesis in Aspergillus flavus


Abstract
:1. Introduction
2. Results
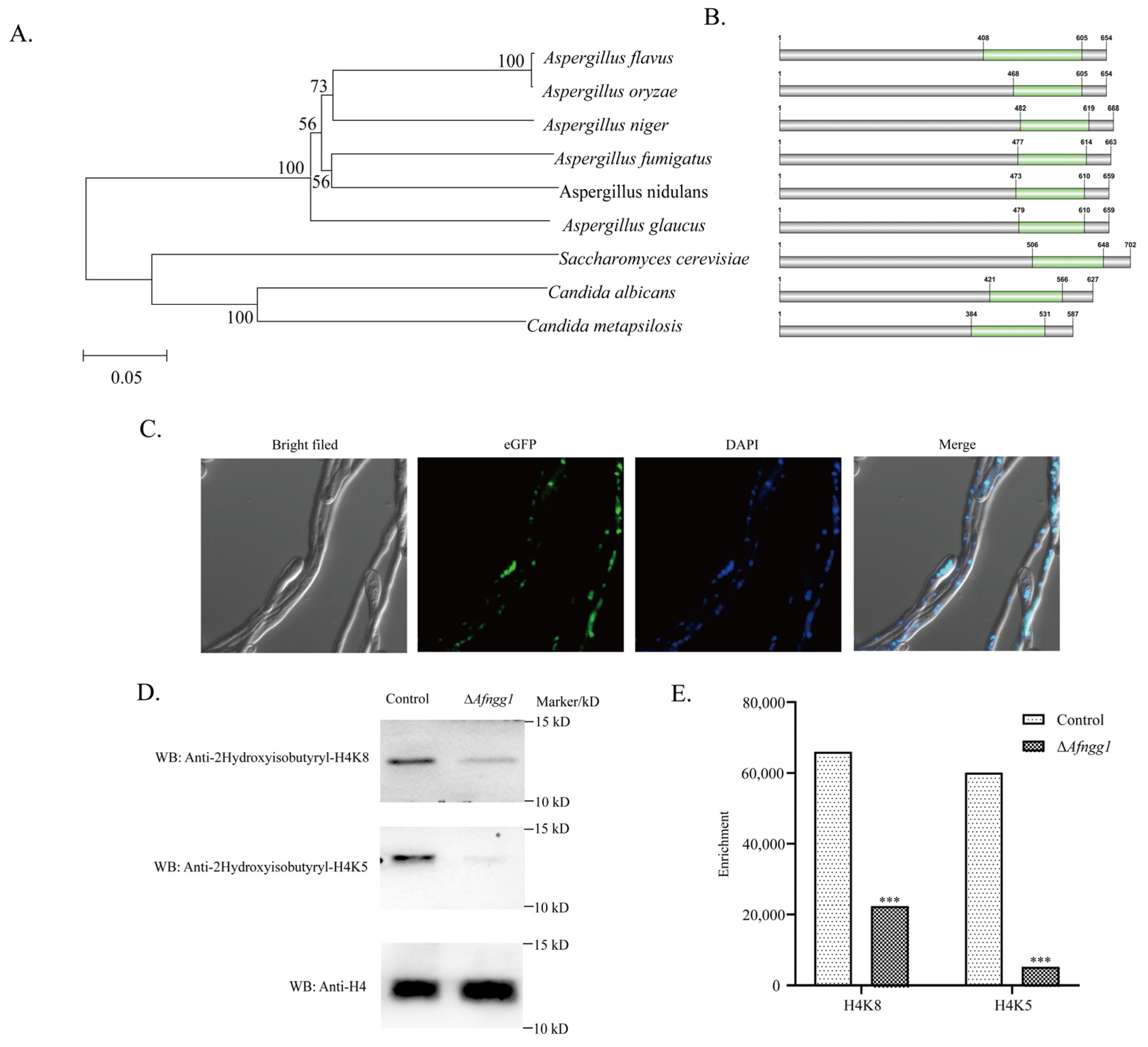
2.1. Characterisation of A. Flavus Histone 2-Hydroxyisobutyryltransferase Afngg1
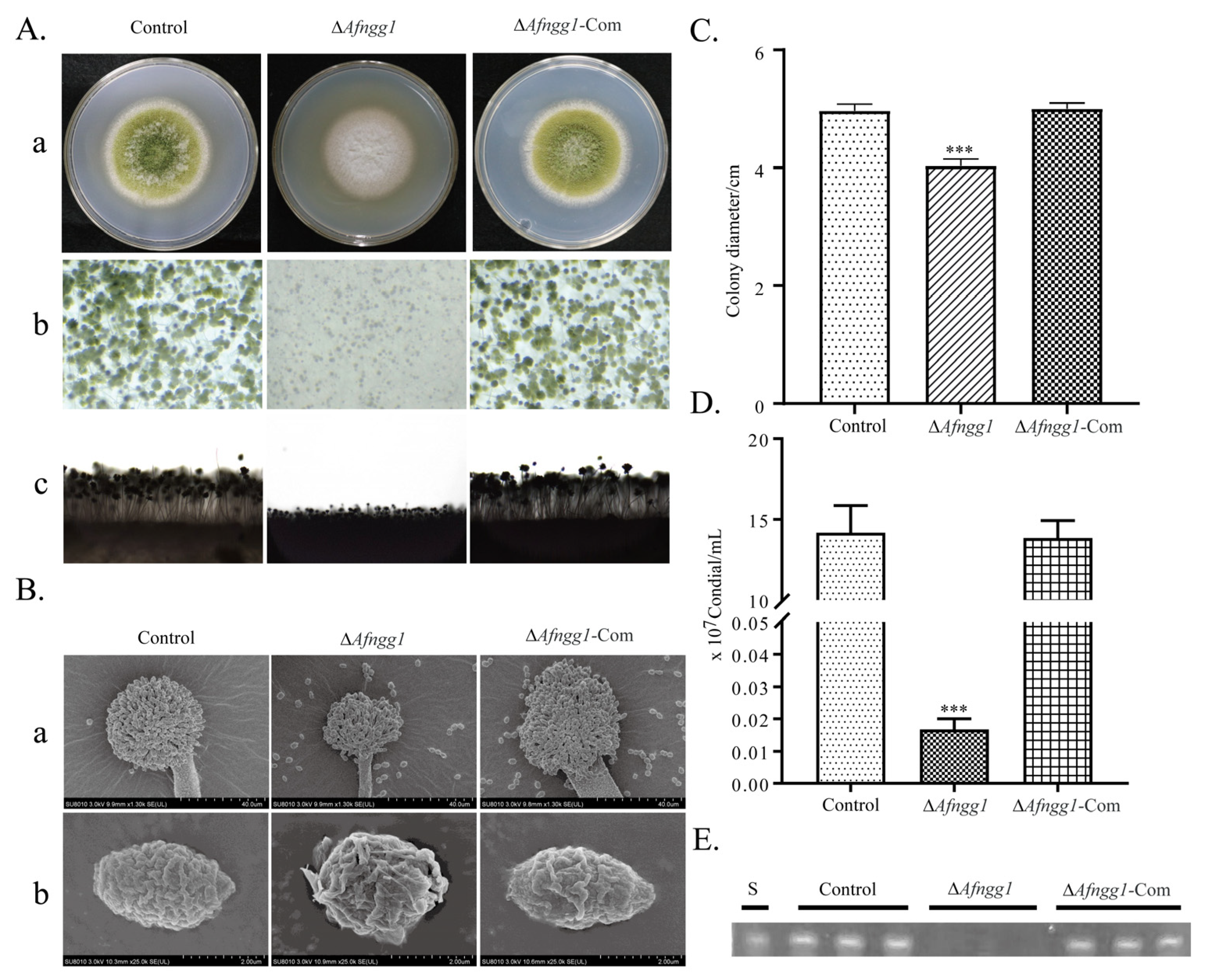
2.2. Afngg1 Is Involved in A. flavus Growth, Conidia Formation and Aflatoxin Biosynthesis
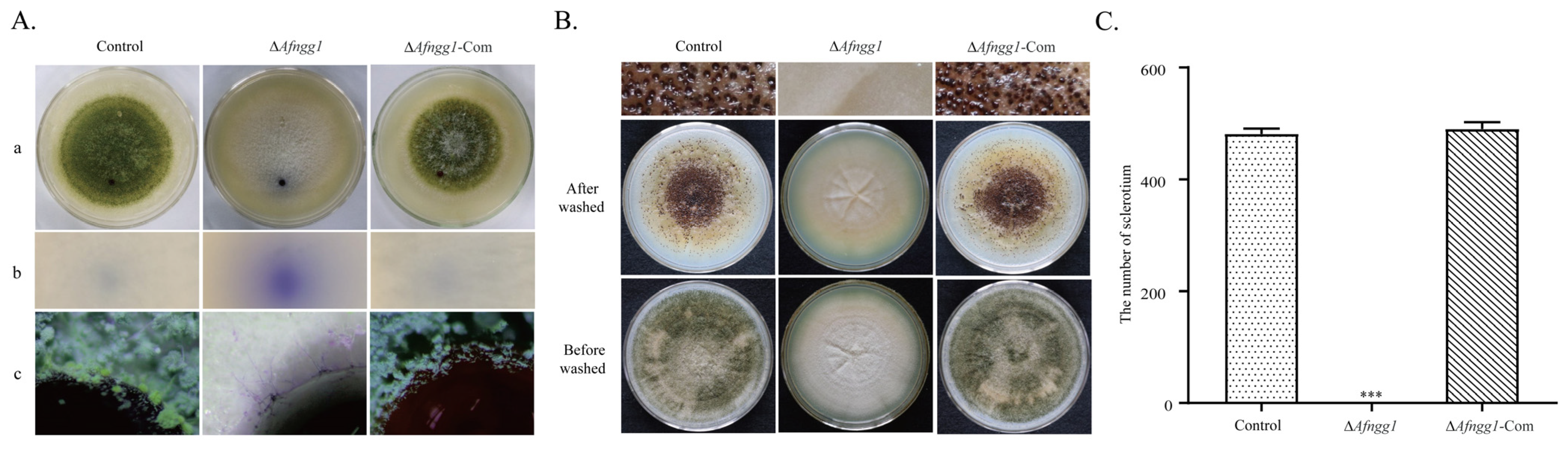
2.3. Effects of Afngg1 on Hydrophobicity and Sclerotia Formation of A. flavus Colonies
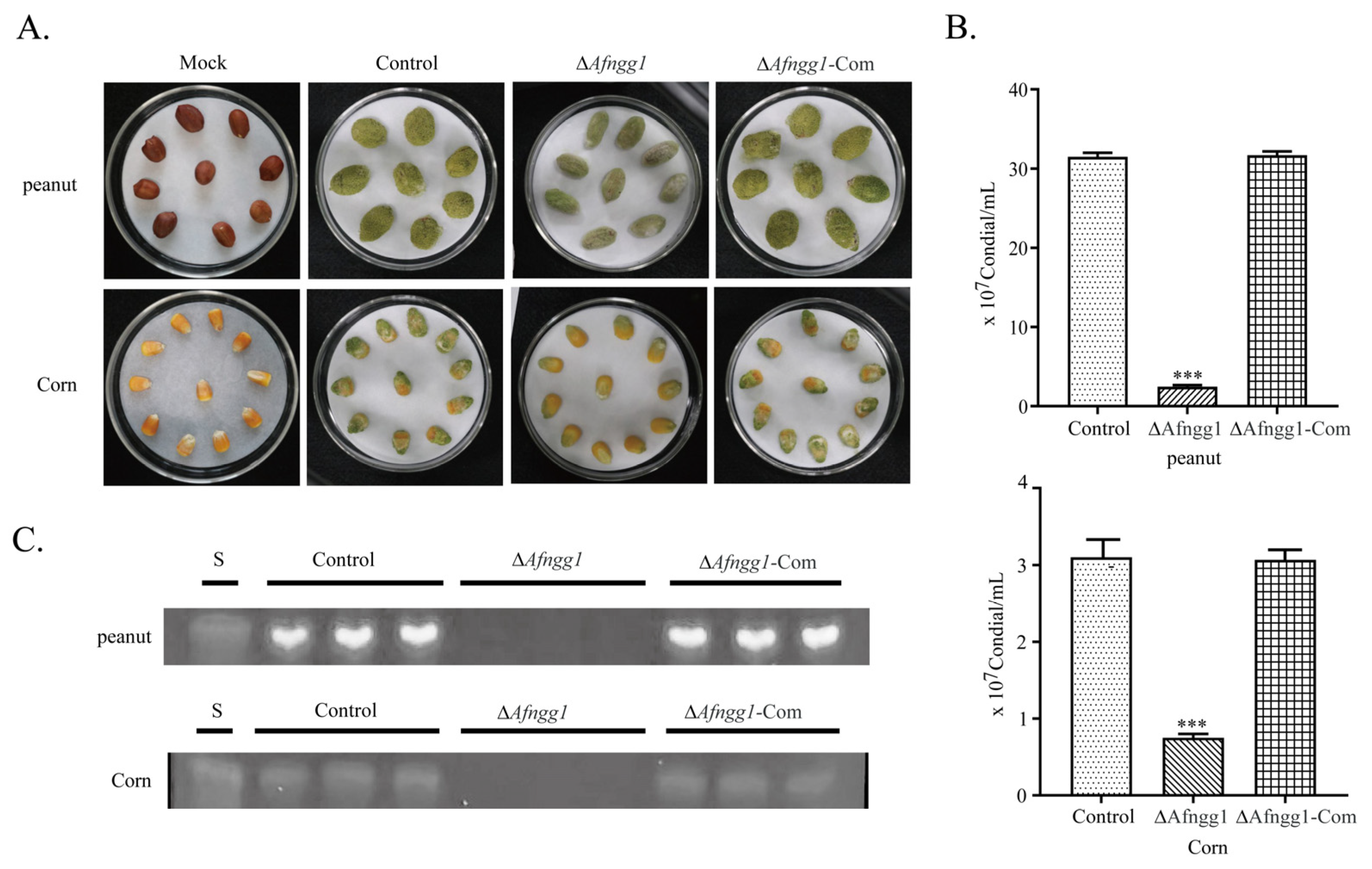
2.4. Afngg1 Influences the Infection Ability of A. flavus on Peanut and Corn
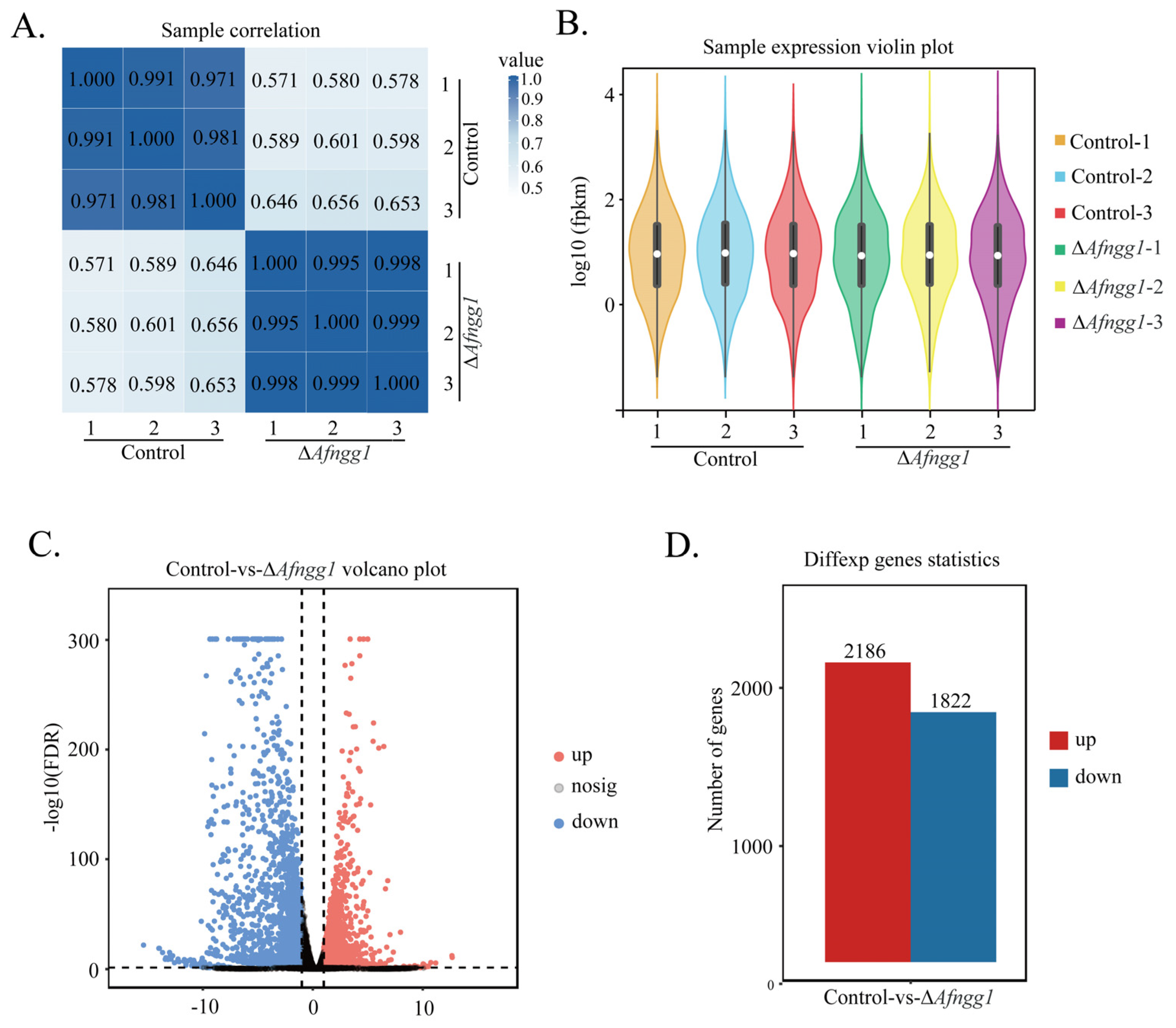
2.5. Transcriptome Overview
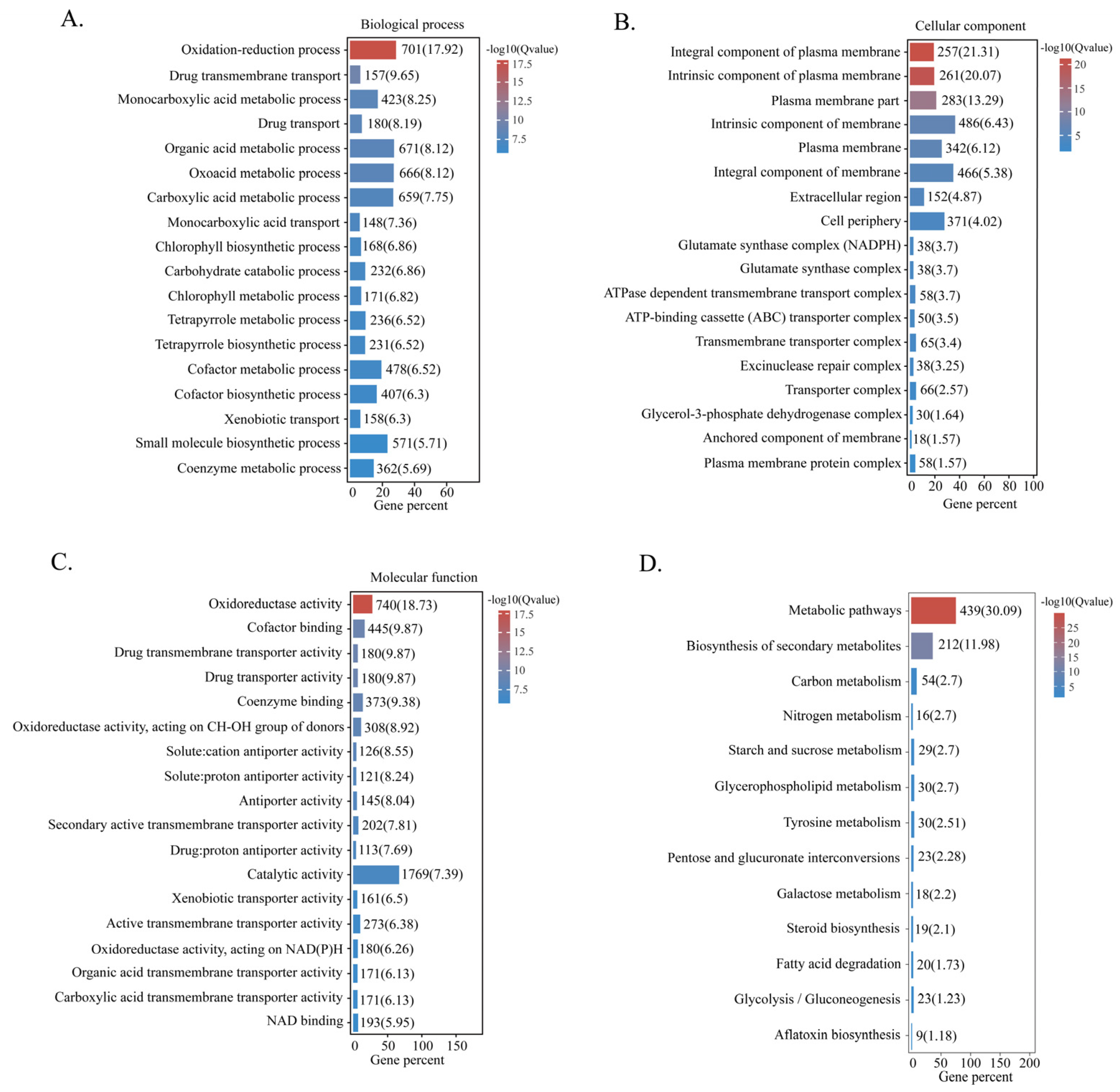
2.6. Categorisation of DEGs
3. Discussion
3.1. Afnng1 Correlates with the Level of Histone 2-Hydroxyisobutyrylation Modification
3.2. Afnng1 Is Involved in Altering Chromatin Remodelling
3.3. Afngg1 Affects Conidial Development and Sclerotium Formation
3.4. Afngg1 Is Required in the Formation of the Cell Wall
3.5. Afngg1 Plays a Key Role in Secondary Metabolism
3.6. Afngg1 Alters Oxidative Stress Homeostasis
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Sequence Resources and Phylogenetic Tree Analysis
4.3. Construction of ΔAfngg1, ΔAfngg1-Com and Afngg1-eGFP Strains
4.4. Subcellular Localisation
4.5. Protein Extraction and Western Blotting Analysis
4.6. Morphological and Physiological Analyses
4.7. Determination of AFB1 Production
4.8. Peanut and Corn Infection Assay
4.9. Transcriptome Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahmoud, M.A.; El-Samawaty, A.M.; Yassin, M.A.; Abd El-Aziz, A.R. Genetic diversity analysis of Aspergillus flavus isolates from plants and air by ISSR markers. Genet. Mol. Res. 2016, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Taniwaki, M.H.; Pitt, J.I.; Magan, N. Aspergillus species and mycotoxins: Occurrence and importance in major food commodities. Curr. Opin. Food Sci. 2018, 23, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Tumukunde, E.; Ma, G.; Li, D.; Yuan, J.; Qin, L.; Wang, S. Current research and prevention of aflatoxins in China. World Mycotoxin J. 2020, 13, 121–138. [Google Scholar] [CrossRef]
- Mitchell, N.J.; Bowers, E.; Hurburgh, C.; Wu, F. Potential economic losses to the US corn industry from aflatoxin contamination. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2016, 33, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Wu, F. Perspective: Time to face the fungal threat. Nature 2014, 516, 7. [Google Scholar] [CrossRef]
- Lan, H.; Sun, R.; Fan, K.; Yang, K.; Zhang, F.; Nie, X.Y.; Wang, X.; Zhuang, Z.; Wang, S. The Aspergillus flavus Histone Acetyltransferase AflGcnE Regulates Morphogenesis, Aflatoxin Biosynthesis, and Pathogenicity. Front. Microbiol. 2016, 7, 1324. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Wang, L.; Ma, X.; Yi, X.; Wang, B.; Liang, W. Proteome-Wide Analysis of Lysine 2-Hydroxyisobutyrylated Proteins in Fusarium oxysporum. Front. Microbiol. 2021, 12, 623735. [Google Scholar] [CrossRef]
- Dong, H.; Zhao, Y.; Bi, C.; Han, Y.; Zhang, J.; Bai, X.; Zhai, G.; Zhang, H.; Tian, S.; Hu, D.; et al. TmcA functions as a lysine 2-hydroxyisobutyryltransferase to regulate transcription. Nat. Chem. Biol. 2022, 18, 142–151. [Google Scholar] [CrossRef]
- Chen, X.; Li, P.; Liu, H.; Chen, X.; Huang, J.; Luo, C.; Li, G.; Hsiang, T.; Collinge, D.B.; Zheng, L. A novel transcription factor UvCGBP1 regulates development and virulence of rice false smut fungus Ustilaginoidea virens. Virulence 2021, 12, 1563–1579. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Liang, W.; Liu, M. Proteome-Wide Analysis of Lysine 2-Hydroxyisobutyrylation in the Phytopathogenic Fungus Botrytis cinerea. Front. Microbiol. 2020, 11, 585614. [Google Scholar] [CrossRef]
- Zheng, H.; Song, N.; Zhou, X.; Mei, H.; Li, D.; Li, X.; Liu, W. Proteome-Wide Analysis of Lysine 2-Hydroxyisobutyrylation in Candida albicans. mSystems 2021, 6, e01129-20. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Jiang, N.; Zhang, Y.; Wang, D.; Sang, X.; Feng, Y.; Chen, R.; Wang, X.; Yang, N.; Chen, Q. Global Lysine Crotonylation and 2-Hydroxyisobutyrylation in Phenotypically Different Toxoplasma gondii Parasites. Mol. Cell Proteom. 2019, 18, 2207–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Y.; Wang, J.; Yang, H.; Li, N.; Farzaneh, M.; Wei, S.; Zhai, H.; Zhang, S.; Hu, Y. Lysine 2-hydroxyisobutyrylation orchestrates cell development and aflatoxin biosynthesis in Aspergillus flavus. Environ. Microbiol. 2022, 24, 4356–4368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Cao, H.; Ma, Y.; Si, H.; Zang, J.; Bai, H.; Yu, L.; Pang, X.; Zhou, F.; Xing, J.; et al. Global analysis of lysine 2-hydroxyisobutyrylation during Fusarium graminearum infection in maize. Front. Plant Sci. 2022, 13, 1000039. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-D.; Fuchs, B.B.; Wang, Y.; Huang, X.-W.; Hu, D.-D.; Sun, Y.; Chai, D.; Jiang, Y.-Y.; Mylonakis, E. Histone acetyltransferase encoded by NGG1 is required for morphological conversion and virulence of Candida albicans. Future Microbiol. 2017, 12, 1497–1510. [Google Scholar] [CrossRef]
- Akiyama, D.Y.; Rocha, M.C.; Costa, J.H.; Teles, C.B.; da Silva Zuccoli, G.; Malavazi, I.; Fill, T.P. The Penicillium brasilianum Histone Deacetylase Clr3 Regulates Secondary Metabolite Production and Tolerance to Oxidative Stress. J. Fungi 2022, 8, 514. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Tang, S.; Ji, M.; Tang, Z.; Shimada, M.; Liu, X.; Qi, S.; Locasale, J.W.; Roeder, R.G.; Zhao, Y.; et al. p300-Mediated Lysine 2-Hydroxyisobutyrylation Regulates Glycolysis. Mol. Cell 2018, 70, 663–678.e6. [Google Scholar] [CrossRef]
- Lin, C.; Cao, X.; Qu, Z.; Zhang, S.; Naqvi, N.I.; Deng, Y.Z. The Histone Deacetylases MoRpd3 and MoHst4 Regulate Growth, Conidiation, and Pathogenicity in the Rice Blast Fungus Magnaporthe oryzae. mSphere 2021, 6, e0011821. [Google Scholar] [CrossRef]
- Creamer, K.M.; Job, G.; Shanker, S.; Neale, G.A.; Lin, Y.-C.; Bartholomew, B.; Partridge, J.F. The Mi-2 Homolog Mit1 Actively Positions Nucleosomes within Heterochromatin to Suppress Transcription. Mol. Cell Biol. 2014, 34, 2046–2061. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Xie, M.; Bai, N.; Yang, L.; Zhang, K.-Q.; Yang, J. The Autophagy-Related Gene Aolatg4 Regulates Hyphal Growth, Sporulation, Autophagosome Formation, and Pathogenicity in Arthrobotrys oligospora. Front. Microbiol. 2020, 11, 592524. [Google Scholar] [CrossRef]
- Cho, H.-J.; Son, S.-H.; Chen, W.; Son, Y.-E.; Lee, I.; Yu, J.-H.; Park, H.-S. Regulation of Conidiogenesis in Aspergillus flavus. Cells 2022, 11, 2796. [Google Scholar] [CrossRef] [PubMed]
- Mead, M.E.; Borowsky, A.T.; Joehnk, B.; Steenwyk, J.L.; Shen, X.-X.; Sil, A.; Rokas, A. Recurrent Loss of abaA, a Master Regulator of Asexual Development in Filamentous Fungi, Correlates with Changes in Genomic and Morphological Traits. Genome Biol. Evol. 2020, 12, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.Y.; Wu, J.; Miller, B.L. StuA is required for cell pattern formation in Aspergillus. Genes Dev. 1992, 6, 1770–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.-Y.; Mead, M.E.; Lee, M.-K.; Neuhaus, G.F.; Adpressa, D.A.; Martien, J.I.; Son, Y.-E.; Moon, H.; Amador-Noguez, D.; Han, K.-H.; et al. Transcriptomic, Protein-DNA Interaction, and Metabolomic Studies of VosA, VelB, and WetA in Aspergillus nidulans Asexual Spores. mBio 2021, 12, e03128-20. [Google Scholar] [CrossRef] [PubMed]
- Eom, T.-J.; Moon, H.; Yu, J.-H.; Park, H.-S. Characterization of the velvet regulators in Aspergillus flavus. J. Microbiol. 2018, 56, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Scharfenstein, L.L.; Mack, B.; Ehrlich, K.C. Deletion of the Aspergillus flavus Orthologue of A. nidulans fluG Reduces Conidiation and Promotes Production of Sclerotia but Does Not Abolish Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2021, 78, 7557–7563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.-K.; Scharfenstein, L.L.; Li, P.; Ehrlich, K.C. Aspergillus flavus VelB acts distinctly from VeA in conidiation and may coordinate with FluG to modulate sclerotial production. Fungal Genet. Biol. 2013, 58–59, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Scharfenstein, L.L.; Ehrlich, K.C.; Wei, Q.; Bhatnagar, D.; Ingber, B.F. Effects of laeA deletion on Aspergillus flavus conidial development and hydrophobicity may contribute to loss of aflatoxin production. Fungal Biol. 2012, 116, 298–307. [Google Scholar] [CrossRef]
- Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 2004, 150, 2029–2035. [Google Scholar] [CrossRef]
- Bernard, M.; Latgé, J.-P. Aspergillus fumigatus cell wall: Composition and biosynthesis. Med. Mycol. 2001, 39, 9–17. [Google Scholar] [CrossRef]
- Pan, S.; Tang, L.; Pan, X.; Qi, L.; Yang, J. A member of the glycoside hydrolase family 76 is involved in growth, conidiation, and virulence in rice blast fungus. Physiol. Mol. Plant Pathol. 2021, 113, 101587. [Google Scholar] [CrossRef]
- Sandini, S.; Stringaro, A.; Arancia, S.; Colone, M.; Mondello, F.; Murtas, S.; Girolamo, A.; Mastrangelo, N.; De Bernardis, F. The MP65 gene is required for cell wall integrity, adherence to epithelial cells and biofilm formation in Candida albicans. BMC Microbiol. 2011, 11, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhu, J.; Ying, S.-H.; Feng, M.-G. The GPI-anchored protein Ecm33 is vital for conidiation, cell wall integrity, and multi-stress tolerance of two filamentous entomopathogens but not for virulence. Appl. Microbiol. Biotechnol. 2014, 98, 5517–5529. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Yang, H.; Wang, J.; Wei, S.; Zhai, H.; Zhang, S.; Hu, Y. Afper1 contributes to cell development and aflatoxin biosynthesis in Aspergillus flavus. Int. J. Food Microbiol. 2022, 377, 109828. [Google Scholar] [CrossRef]
- Li, Q.Z.; He, Z.M. Advances in research of the structural gene characteristics of the aflatoxin biosynthetic gene cluster. J. Plant Sci. Phytopathol. 2015, 2, 068–082. [Google Scholar] [CrossRef] [Green Version]
- Crawford, J.M.; Korman, T.P.; Labonte, J.W.; Vagstad, A.L.; Hill, E.A.; Kamari-Bidkorpeh, O.; Tsai, S.-C.; Townsend, C.A. Structural basis for biosynthetic programming of fungal aromatic polyketide cyclization. Nature 2009, 461, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; He, Y.; Li, X.; Fasoyin, O.E.; Hu, Y.; Liu, Y.; Yuan, J.; Zhuang, Z.; Wang, S. Histone Methyltransferase aflrmtA gene is involved in the morphogenesis, mycotoxin biosynthesis, and pathogenicity of Aspergillus flavus. Toxicon 2017, 127, 112–121. [Google Scholar] [CrossRef]
- Cary, J.W.; Ehrlich, K.C.; Bland, J.M.; Montalbano, B.G. The Aflatoxin Biosynthesis Cluster Gene, aflX, Encodes an Oxidoreductase Involved in Conversion of Versicolorin A to Demethylsterigmatocystin. Appl. Environ. Microbiol. 2006, 72, 1096–1101. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Branà, M.T.; Haidukowski, M.; Gallo, A.; Zhang, Q.; Logrieco, A.F.; Li, P.; Zhao, S.; Altomare, C. Potential of Trichoderma spp. for Biocontrol of Aflatoxin-Producing Aspergillus flavus. Toxins 2022, 14, 86. [Google Scholar] [CrossRef]
- Jamali, M.; Karimipour, M.; Shams-Ghahfarokhi, M.; Amani, A.; Razzaghi-Abyaneh, M. Expression of aflatoxin genes aflO (omtB) and aflQ (ordA) differentiates levels of aflatoxin production by Aspergillus flavus strains from soils of pistachio orchards. Res. Microbiol. 2013, 164, 293–299. [Google Scholar] [CrossRef]
- Yabe, K.; Chihaya, N.; Hamamatsu, S.; Sakuno, E.; Hamasaki, T.; Nakajima, H.; Bennett, J.W. Enzymatic Conversion of Averufin to Hydroxyversicolorone and Elucidation of a Novel Metabolic Grid Involved in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2003, 69, 66–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.-K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered Pathway Genes in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Umemura, M.; Nagano, N.; Koike, H.; Kawano, J.; Ishii, T.; Miyamura, Y.; Kikuchi, M.; Tamano, K.; Yu, J.; Shin-Ya, K.; et al. Characterization of the biosynthetic gene cluster for the ribosomally synthesized cyclic peptide ustiloxin B in Aspergillus flavus. Fungal Genet. Biol. 2014, 68, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon-Chung, K.J.; Sugui, J.A. What do we know about the role of gliotoxin in the pathobiology of Aspergillus fumigatus? Med. Mycol. 2009, 47, S97–S103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.-N.; Toyotome, T.; Muraosa, Y.; Watanabe, A.; Wuren, T.; Bunsupa, S.; Aoyagi, K.; Yamazaki, M.; Takino, M.; Kamei, K. GliA in Aspergillus fumigatus is required for its tolerance to gliotoxin and affects the amount of extracellular and intracellular gliotoxin. Med. Mycol. 2014, 52, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, L.; Owens, R.A.; Dolan, S.K.; O’Keeffe, G.; Schrettl, M.; Kavanagh, K.; Jones, G.W.; Doyle, S. The Aspergillus fumigatus Protein GliK Protects against Oxidative Stress and Is Essential for Gliotoxin Biosynthesis. Eukaryot. Cell 2012, 11, 1226–1238. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nature reviews. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Langfelder, K.; Jahn, B.; Gehringer, H.; Schmidt, A.; Wanner, G.; Brakhage, A.A. Identification of a polyketide synthase gene (pksP) of Aspergillus fumigatus involved in conidial pigment biosynthesis and virulence. Med. Microbiol. Immunol. 1998, 187, 79–89. [Google Scholar] [CrossRef]
- Takano, Y.; Kubo, Y.; Shimizu, K.; Mise, K.; Okuno, T.; Furusawa, I. Structural analysis of PKS1, a polyketide synthase gene involved in melanin biosynthesis in Colletotrichum lagenarium. Mol. Gen. Genet. 1995, 249, 162–167. [Google Scholar] [CrossRef]
- Zhou, R.; Rasooly, R.; Linz, J.E. Isolation and analysis of fluP, a gene associated with hyphal growth and sporulation in Aspergillus parasiticus. Mol. Gen. Genet. 2000, 264, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yang, M.; Bai, Y.; Ge, F.; Wang, S. Antioxidant-related catalaseCTA1regulates development, aflatoxin biosynthesis, and virulence in pathogenic fungus Aspergillus flavus. Environ. Microbiol. 2020, 22, 2792–2810. [Google Scholar] [CrossRef] [PubMed]
- Domènech, A.; Ayté, J.; Antunes, F.; Hidalgo, E. Using in vivo oxidation status of one- and two-component redox relays to determine H2O2 levels linked to signaling and toxicity. BMC Biol. 2018, 16, 61. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, E.; Nayak, T.; Oakley, C.E.; Edgerton, H.; Xiong, Y.; Taheri-Talesh, N.; Osmani, S.A.; Oakley, B.R. Fusion PCR and gene targeting in Aspergillus nidulans. Nat. Protoc. 2006, 1, 3111–3120. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Yang, M.; Yue, Y.; Ge, F.; Li, Y.; Guo, X.; Zhang, J.; Zhang, F.; Nie, X.; Wang, S. Lysine Succinylation Contributes to Aflatoxin Production and Pathogenicity in Aspergillus flavus. Mol. Cell Proteom. 2018, 17, 457–471. [Google Scholar] [CrossRef] [Green Version]
- Mengjuan, Z.; Guanglan, L.; Xiaohua, P.; Weitao, S.; Can, T.; Xuan, C.; Yanling, Y.; Zhenhong, Z. The PHD transcription factor Cti6 is involved in the fungal colonization and aflatoxin B1 biological synthesis of Aspergillus flavus. IMA Fungus 2021, 12, 12. [Google Scholar] [CrossRef]
- Zhang, W.; Lv, Y.; Lv, A.; Wei, S.; Zhang, S.; Li, C.; Hu, Y. Sub3 inhibits Aspergillus flavus growth by disrupting mitochondrial energy metabolism, and has potential biocontrol during peanut storage. J. Sci. Food Agric. 2021, 101, 486–496. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, G.; Zhang, D.; Liu, Y.; Li, Y.; Lin, G.; Guo, Z.; Wang, S.; Zhuang, Z. The PHD Transcription Factor Rum1 Regulates Morphogenesis and Aflatoxin Biosynthesis in Aspergillus flavus. Toxins 2018, 10, 301. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.-K.; Zhang, Q.; Scharfenstein, L.; Mack, B.; Yoshimi, A.; Miyazawa, K.; Abe, K. Aspergillus flavus GPI-anchored protein-encoding ecm33 has a role in growth, development, aflatoxin biosynthesis, and maize infection. Appl. Microbiol. Biotechnol. 2018, 102, 5209–5220. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Category | Name | Log2(fc) | p-Value | Description |
|---|---|---|---|---|
| Chromatin remodelling | ||||
| AFLA_025120 | clr3 | 1.089 | 2.82 × 10−21 | histone deacetylase |
| MSTRG.3596 | RPD3 | 1.15 | 1.26 × 10−6 | histone deacetylase RPD3 |
| AFLA_055490 | epl1 | 1.51 | 5.53 × 10−44 | histone acetyltransferase complex component |
| AFLA_052300 | mit1 | 1.49 | 7.93 × 10−28 | chromatin remodelling complex subunit (Chd3) |
| Cell development | ||||
| AFLA_052030 | wetA | −2.32 | 1.96 × 10−120 | developmental regulatory protein WetA |
| AFLA_029620 | abaA | −3.17 | 1.99 × 10−171 | transcription factor abaA |
| AFLA_082850 | brlA | −5.27 | 0 | C2H2 type conidiation transcription factor |
| AFLA_046990 | stuA | −1.03 | 5.33 × 10−46 | APSES transcription factor StuA |
| AFLA_026900 | vosA | −2.28 | 1.98 × 10−75 | developmental regulator VosA |
| AFLA_039530 | fluG | −3.17 | 1.28 × 10−13 | FluG family protein |
| AFLA_044800 | con-6 | −8.10 | 7.93 × 10−60 | conidiation protein Con-6 |
| AFLA_098380 | rodA | −7.58 | 2.79 × 10−67 | RodA/RolA7 |
| AFLA_060780 | dewA | −6.81 | 8.80 × 10−9 | hydrophobin family protein |
| AFLA_023460 | ags1 | −2.80 | 6.03 × 10−201 | alpha-1,3-glucan synthase Ags1 |
| AFLA_134100 | ags2 | −2.28 | 4.34 × 10−119 | alpha-1,3-glucan synthase Ags2 |
| AFLA_052780 | MP65 | −3.87 | 1.26 × 10−141 | cell wall glucanase (Scw4) |
| AFLA_104680 | chiB1 | −1.64 | 1.57 × 10−22 | class V chitinase ChiB1 |
| AFLA_024280 | dcw1 | −5.49 | 0 | cell wall glycosyl hydrolase Dfg5 |
| AFLA_113120 | ecm33 | −1.02 | 6.90 × 10−76 | GPI-anchored cell wall organization protein |
| AFLA_138060 | erg4 | −2.95 | 1.32 × 10−108 | c-24(28) sterol reductase |
| AFLA_001030 | erg7 | −9.21 | 1.07 × 10−73 | lanosterol synthase |
| AFLA_028640 | erg5 | −5.79 | 2.08 × 10−143 | cytochrome P450 sterol C-22 desaturase |
| Secondary metabolism | ||||
| AFLA_139410 | aflC | −1.00 | 3.76 × 10−32 | aflC/pksA/pksL1/polyketide synthase |
| AFLA_139330 | aflH | −1.46 | 2.3 × 10−3 | aflH/short chain alcohol dehydrogenase |
| AFLA_139160 | aflX | −2.37 | 1.05 × 10−2 | aflX/ordB/monooxygenase/oxidase |
| AFLA_139320 | aflJ | −2.82 | 1.28 × 10−2 | aflJ/estA/esterase |
| AFLA_139170 | aflW | −3.82 | 6.16 × 10−6 | aflW/moxY/monooxygenase |
| AFLA_002920 | aflQ | −4.82 | 2.29 × 10−46 | flavonoid 3-hydroxylase |
| AFLA_024090 | aflT | 1.57 | 8.06 × 10−24 | efflux pump antibiotic resistance protein |
| AFLA_139310 | norA | −8.34 | 7 × 10−4 | aflE/norA/aad/adh-2/ NOR reductase |
| AFLA_094990 | ustYa | −7.92 | 2.80 × 10−21 | Ustiloxin B biosynthesis protein Ya |
| AFLA_094940 | ustO | −2.98 | 2.18 × 10−108 | Ustiloxin B biosynthesis protein O |
| AFLA_094960 | ustC | −4.14 | 1.27 × 10−48 | cytochrome P450 |
| AFLA_095010 | ustP | −4.59 | 1.18 × 10−84 | Ustiloxin B biosynthesis protein P |
| AFLA_095060 | ustQ | −4.75 | 1.78 × 10−35 | tyrosinase |
| AFLA_095040 | ustD | −6.18 | 9.10 × 10−2 | NRPS-like enzyme |
| AFLA_095050 | ustF2 | −7.29 | 5.02 × 10−20 | dimethylaniline monooxygenase |
| AFLA_064420 | gliK | −3.26 | 3.49 × 10−12 | gliotoxin biosynthesis protein GliK |
| AFLA_118990 | gliA | −4.91 | 1.47 × 10−127 | efflux pump antibiotic resistance protein |
| AFLA_064540 | gliC | −6.69 | 1.62 × 10−55 | cytochrome P450 oxidoreductase |
| AFLA_114820 | fluP | −1.44 | 1.78 × 10−50 | polyketide synthase |
| AFLA_006170 | pksP | −6.54 | 0 | polyketide synthetase PksP |
| AFLA_010000 | aurA | −2.23 | 2.02 × 10−84 | polyketide synthase |
| AFLA_054090 | pks1 | −2.22 | 7.5 × 10−3 | polyketide synthase |
| Oxidative stress | ||||
| AFLA_034380 | cta1 | −7.68 | 0 | catalase |
| AFLA_096210 | ctl-2 | −6.96 | 6.43 × 10−272 | catalase |
| AFLA_100250 | cat | −1.57 | 6.97 × 10−16 | catalase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Liang, L.; Wei, S.; Zhang, S.; Hu, Y.; Lv, Y. Histone 2-Hydroxyisobutyryltransferase Encoded by Afngg1 Is Involved in Pathogenicity and Aflatoxin Biosynthesis in Aspergillus flavus. Toxins 2023, 15, 7. https://doi.org/10.3390/toxins15010007
Wang J, Liang L, Wei S, Zhang S, Hu Y, Lv Y. Histone 2-Hydroxyisobutyryltransferase Encoded by Afngg1 Is Involved in Pathogenicity and Aflatoxin Biosynthesis in Aspergillus flavus. Toxins. 2023; 15(1):7. https://doi.org/10.3390/toxins15010007
Chicago/Turabian StyleWang, Jing, Liuke Liang, Shan Wei, Shuaibing Zhang, Yuansen Hu, and Yangyong Lv. 2023. "Histone 2-Hydroxyisobutyryltransferase Encoded by Afngg1 Is Involved in Pathogenicity and Aflatoxin Biosynthesis in Aspergillus flavus" Toxins 15, no. 1: 7. https://doi.org/10.3390/toxins15010007
APA StyleWang, J., Liang, L., Wei, S., Zhang, S., Hu, Y., & Lv, Y. (2023). Histone 2-Hydroxyisobutyryltransferase Encoded by Afngg1 Is Involved in Pathogenicity and Aflatoxin Biosynthesis in Aspergillus flavus. Toxins, 15(1), 7. https://doi.org/10.3390/toxins15010007