Preliminary Investigation of Biogenic Amines in Type I Sourdoughs Produced at Home and Bakery Level

 ,
,  ,
,  , , , , ,
, , , , ,  ,
,  and
and
Abstract
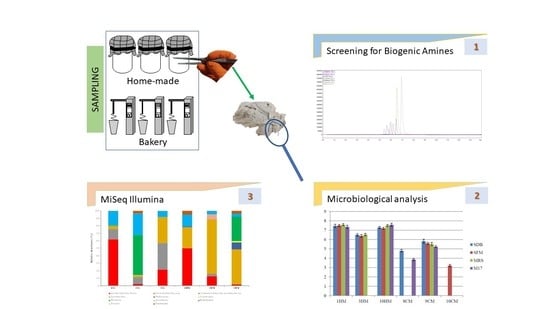
:
1. Introduction
2. Results and Discussion
2.1. Determination, Identification, and Quantification of Biogenic Amines in Sourdoughs
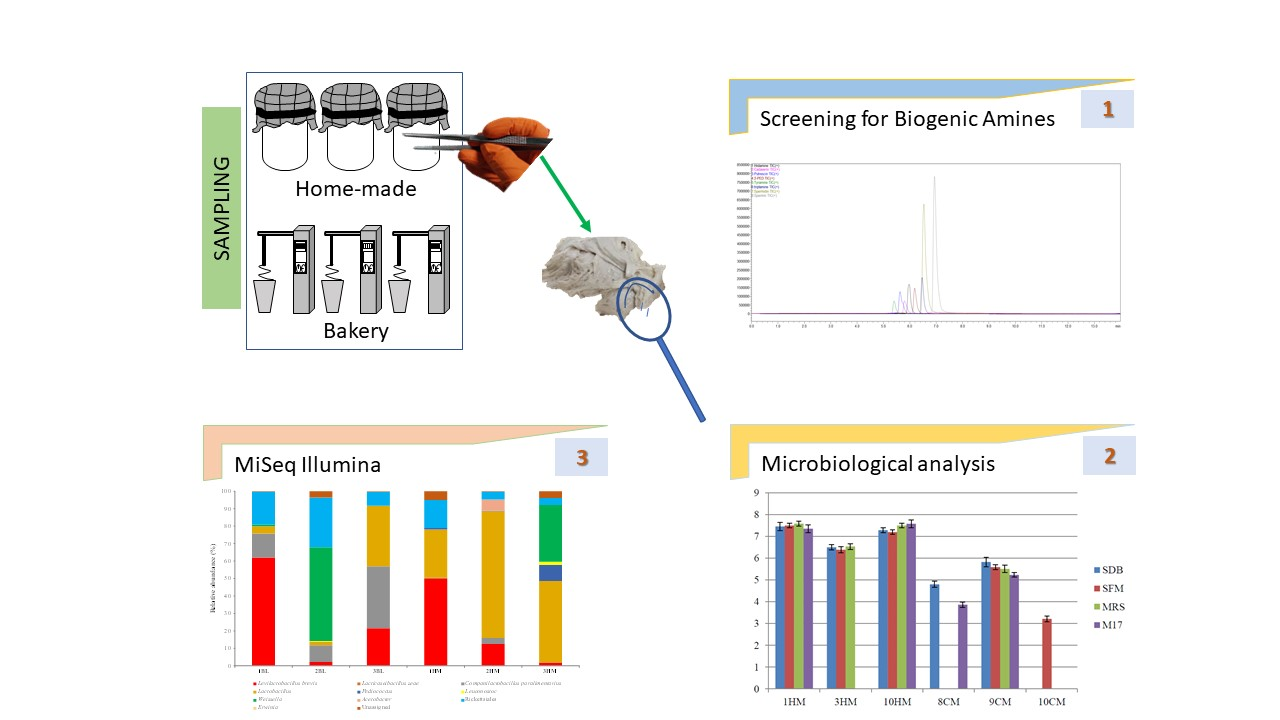
2.2. Taxonomic Distribution of Bacterial OTUs
2.3. Acidity of Sourdoughs
2.4. Levels of Viable LAB and Yeasts
2.5. Multivariate Analysis
3. Conclusions
4. Materials and Methods
4.1. Sourdough Collection
4.2. Screening for Biogenic Amines
4.3. UHPLC-MS/MS Evaluation of Sourdough Biogenic Amines
4.4. Culture-Independent Microbiological Analysis
4.4.1. DNA Extraction
4.4.2. Miseq Library Preparation and Illumina Sequencing
4.4.3. Illumina Data Analysis and Sequences Identification by QIIME2
4.5. Physicochemical Parameters and Microbiological Analysis
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silla-Santos, M.H. Biogenic amines: Their importance in food. Int. J. Food Microbiol. 1996, 29, 213–221. [Google Scholar] [CrossRef]
- Benkerroum, N. Biogenic amines in dairy products: Origin, incidence, and control means. Compr. Rev. Food Sci. Food Saf. 2016, 15, 801–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doeun, D.; Davaatseren, M.; Chung, M.S. Biogenic amines in foods. Food Sci. Biotechnol. 2017, 26, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Suzzi, G.; Torriani, S. Biogenic amines in foods. Front. Microbiol. 2015, 6, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadáková, E.; Křížek, P.; Pelikánová, T. Determination of biogenic amines in foods using ultra-performance liquid chromatography (UPLC). Food Chem. 2009, 116, 365–370. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific opinion on risk based control of biogenic amine formation in fermented foods. Efsa J. 2011, 9, 2393–2486. [Google Scholar] [CrossRef] [Green Version]
- Ladero, V.; Calles-Enríquez, M.; Fernández, M.; Alvarez, M. Toxicological effects of dietary biogenic amines. Curr. Nutr. Food Sci. 2010, 6, 145–156. [Google Scholar] [CrossRef]
- Kim, M.K.; Mah, J.H.; Hwang, H.J. Biogenic amine formation and bacterial contribution in fish, squid and shellfish. Food Chem. 2009, 116, 87–95. [Google Scholar] [CrossRef]
- Koral, S.; Tufan, B.; Ščavničar, A.; Kočar, D.; Pompe, M.; Köse, S. Investigation of the contents of biogenic amines and some food safety parameters of various commercially salted fish products. Food Control 2013, 32, 597–606. [Google Scholar] [CrossRef]
- Leuschner, R.G.; Hristova, A.; Robinson, T.; Hugas, M. The Rapid Alert System for Food and Feed (RASFF) database in support of risk analysis of biogenic amines in food. J. Food Compos. Anal. 2013, 29, 37–42. [Google Scholar] [CrossRef]
- Jørgensen, L.V.; Dalgaard, P.; Huss, H.H. Multiple compound quality index for cold-smoked salmon (Salmo salar) developed by multivariate regression of biogenic amines and pH. J. Agric. Food Chem. 2000, 48, 2448–2453. [Google Scholar] [CrossRef] [PubMed]
- Sims, G.G.; Farn, G.; York, R.K. Quality indices for canned skipjack tuna: Correlation of sensory attributes with chemical indices. J. Food Sci. 1992, 57, 1112–1115. [Google Scholar] [CrossRef]
- Kandasamy, S.; Kavitake, D.; Shetty, P.H. Lactic acid bacteria and yeasts as starter cultures for fermented foods and their role in commercialization of fermented foods. In Innovations in Technologies for Fermented Food and Beverage Industries; Panda, S.K., Shetty, P.H., Eds.; Springer: New York City, NY, USA, 2018; pp. 25–52. [Google Scholar]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- Karovičová, J.; Kohajdová, Z. Biogenic amines in food. Chem. Pap. 2005, 59, 70–79. [Google Scholar]
- Lonvaud-Funel, A. Biogenic amines in wines: Role of lactic acid bacteria. FEMS Microbiol. Lett. 2001, 199, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Brandt, M.J. Industrial production of sourdoughs for the baking branch–An overview. Int. J. Food Microbiol. 2019, 302, 3–7. [Google Scholar] [CrossRef]
- Del Rio, B.; Alvarez-Sieiro, P.; Redruello, B.; Martin, M.C.; Fernandez, M.; Ladero, V.; Alvarez, M.A. Lactobacillus rossiae strain isolated from sourdough produces putrescine from arginine. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Di Cagno, R.; De Angelis, M.; Gallo, G.; Settanni, L.; Berloco, M.G.; Siragusa, S.; Parente, E.; Corsetti, A.; Gobbetti, M. Genotypic and phenotypic diversity of Lactobacillus rossiae strains isolated from sourdough. J. Appl. Microbiol. 2007, 103, 821–835. [Google Scholar] [CrossRef]
- Scheirlinck, I.; Van der Meulen, R.; De Vuyst, L.; Vandamme, P.; Huys, G. Molecular source tracking of predominant lactic acid bacteria in traditional Belgian sourdoughs and their production environments. J. Appl. Microbiol. 2009, 106, 1081–1092. [Google Scholar] [CrossRef]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef] [Green Version]
- Valmorri, S.; Settanni, L.; Suzzi, G.; Gardini, F.; Vernocchi, P.; Corsetti, A. Application of a novel polyphasic approach to study the lactobacilli composition of sourdoughs from the Abruzzo region (Central Italy). Lett. Appl. Microbiol. 2006, 43, 343–349. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Comasio, A.; Kerrebroeck, S.V. Sourdough production: Fermentation strategies, microbial ecology, and use of non-flour ingredients. Crit. Rev. Food Sci. Nutr. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Kurt, Ş.; Zorba, Ö. The effects of ripening period, nitrite level and heat treatment on biogenic amine formation of “sucuk”—A Turkish dry fermented sausage. Meat Sci. 2009, 82, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Flick, G.J.; Oria, M.P.; Douglas, L. Potential hazards in cold-smoked fish: Biogenic amines. J. Food Sci. 2001, 66, S1088–S1099. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, 338, 1–26. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32005R2073 (accessed on 1 March 2022).
- Barros, D.; Barros, M.; Velho, M.V.; Pires, P.; Dias, R.; Pinto, R. Cadaverine and putrescine contents in traditional Portuguese pork sausages linked to the addition of starter cultures. Safe Pork 2019, 13, 199–201. [Google Scholar]
- Yeh, C.Y.; Lin, S.J.; Hwang, D.F. Biogenic amines, histamine and label of dressed fried fish meat products in Taiwan. Food Control 2006, 17, 423–428. [Google Scholar] [CrossRef]
- Nout, M.J.R. Food technologies: Fermentation. In Encyclopedia of Food Safety; Holt, S., Phadnis, R., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 3, pp. 168–177. [Google Scholar]
- Guarcello, R.; Diviccaro, A.; Barbera, M.; Giancippoli, E.; Settanni, L.; Minervini, F.; Moschetti, G.; Gobbetti, M. A survey of the main technology, biochemical and microbiological features influencing the concentration of biogenic amines of twenty Apulian and Sicilian (Southern Italy) cheeses. Int. Dairy J. 2015, 43, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Latorre-Moratalla, M.L.; Comas-Basté, O.; Bover-Cid, S.; Vidal-Carou, M.C. Tyramine and histamine risk assessment related to consumption of dry fermented sausages by the Spanish population. Food Chem. Toxicol. 2017, 99, 78–85. [Google Scholar] [CrossRef]
- Gillman, P.K. A reassessment of the safety profile of monoamine oxidase inhibitors: Elucidating tired old tyramine myths. J. Neural Trans. 2018, 125, 1707–1717. [Google Scholar] [CrossRef]
- del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernandez, M.; Martìn, M.C.; Alvarez, M.A. The biogenic amines putrescine and cadaverine show in vitro cytotoxicity at concentrations that can be found in foods. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernandez, M.; Martìn, M.C.; Alvarez, M.A. Spermine and spermidine are cytotoxic towards intestinal cell cultures, but are they a health hazard at concentrations found in foods? Food Chem. 2018, 269, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Wu, J.; An, F.; Yue, X.; Tao, D.; Wu, R.; Lee, Y. An integrated metagenomic/metaproteomic investigation of microbiota in dajiang-meju, a traditional fermented soybean product in Northeast China. Food Res. Int. 2019, 115, 414–424. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, Y.; Xiang, F.; Hou, Q.; Zhang, Z. Bacterial Diversity in Pickled Cowpea (Vigna unguiculata [Linn.] Walp) as Determined by Illumina MiSeq Sequencing and Culture-Dependent Methods. Curr. Microbiol. 2021, 78, 1286–1297. [Google Scholar] [CrossRef]
- Logares, R.; Audic, S.; Bass, D.; Bittner, L.; Boutte, C.; Christen, R.; Claverie, J.M.; Decelle, J.; Dolan, J.R.; Dunthorn, M.; et al. Patterns of rare and abundant marine microbial eukaryotes. Curr. Biol. 2014, 24, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Corsetti, A.; Settanni, L. Lactobacilli in sourdough fermentation: A review. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Settanni, L.; van Sinderen, D.; Rossi, J.; Corsetti, A. Rapid differentiation and in situ detection of 16 sourdough Lactobacillus species by multiplex PCR. Appl. Environ. Microbiol. 2005, 71, 3049–3059. [Google Scholar] [CrossRef] [Green Version]
- Settanni, L.; Valmorri, S.; Van Sinderen, D.; Suzzi, G.; Paparella, A.; Corsetti, A. Combination of multiplex PCR and PCR-DGGE for monitoring common sourdough-associated Lactobacillus species. Appl. Environ. Microbiol. 2006, 72, 3793–3796. [Google Scholar] [CrossRef] [Green Version]
- Ventimiglia, G.; Alfonzo, A.; Galluzzo, P.; Corona, O.; Francesca, N.; Caracappa, S.; Moschetti, G.; Settanni, L. Codominance of Lactobacillus plantarum and obligate heterofermentative lactic acid bacteria during sourdough fermentation. Food Microbiol. 2015, 51, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, W.; Sun, Z.; Bao, Q.; Wang, F.; Yu, J.; Chen, W.; Zhang, H. Diversity of lactic acid bacteria and yeasts in traditional sourdoughs collected from western region in Inner Mongolia of China. Food Control 2011, 22, 767–774. [Google Scholar] [CrossRef]
- Alfonzo, A.; Miceli, C.; Nasca, A.; Franciosi, E.; Ventimiglia, G.; Di Gerlando, R.; Tuohi, K.; Francesca, N.; Moschetti, G.; Settanni, L. Monitoring of wheat lactic acid bacteria from the field until the first step of dough fermentation. Food Microbiol. 2017, 62, 256–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaglio, R.; Cirlincione, F.; Di Miceli, G.; Franciosi, E.; Di Gerlando, R.; Francesca, N.; Settanni, L.; Moschetti, G. Microbial dynamics in durum wheat kernels during aging. Int. J. Food Microbiol. 2020, 324, 108631. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Ventimiglia, G.; Alfonzo, A.; Corona, O.; Miceli, A.; Moschetti, G. An integrated technological approach to the selection of lactic acid bacteria of flour origin for sourdough production. Food Res. Int. 2013, 54, 1569–1578. [Google Scholar] [CrossRef] [Green Version]
- Alfonzo, A.; Urso, V.; Corona, O.; Francesca, N.; Amato, G.; Settanni, L.; Di Miceli, G. Development of a method for the direct fermentation of semolina by selected sourdough lactic acid bacteria. Int. J. Food Microbiol. 2016, 239, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Corona, O.; Alfonzo, A.; Ventimiglia, G.; Nasca, A.; Francesca, N.; Martorana, A.; Moschetti, G.; Settanni, L. Industrial application of selected lactic acid bacteria isolated from local semolinas for typical sourdough bread production. Food Microbiol. 2016, 59, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Crespo, L.; Gaglio, R.; Martínez, F.G.; Moreno Martin, G.; Franciosi, E.; Madrid-Albarrán, Y.; Settanni, L.; Mozzi, F.; Pescuma, M. Bioaccumulation of selenium-by fruit origin lactic acid bacteria in tropical fermented fruit juices. LWT-Food Sci. Technol. 2021, 151, 112103. [Google Scholar] [CrossRef]
- Yu, X.J.; Walker, D.H. The order Rickettsiales. Prokaryotes 2006, 5, 493–528. [Google Scholar]
- Ripari, V.; Gänzle, M.G.; Berardi, E. Evolution of sourdough microbiota in spontaneous sourdoughs started with different plant materials. Int. J. Food Microbiol. 2016, 232, 35–42. [Google Scholar] [CrossRef]
- Scheirlinck, I.; Van der Meulen, R.; Van Schoor, A.; Vancanneyt, M.; De Vuyst, L.; Vandamme, P.; Huys, G. Taxonomic structure and stability of the bacterial community in Belgian sourdough ecosystems as assessed by culture and population fingerprinting. Appl. Environ. Microbiol. 2008, 74, 2414–2423. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, S.A.; Seitter, M.; Singer, U.; Brandt, M.J.; Hertel, C. Adaptability of lactic acid bacteria and yeasts to sourdoughs prepared from cereals, pseudocereals and cassava and use of competitive strains as starters. Int. J. Food Microbiol. 2009, 130, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Sagar, A.; Thomas, G.; Rai, S.; Mishra, R.K.; Ramteke, P.W. Enhancement of growth and yield parameters of wheat variety AAI-W6 by an organic farm isolate of plant growth promoting Erwinia Species (KP226572). Int. J. Agric. Environ. Biotechnol. 2018, 11, 159–171. [Google Scholar]
- Boreczek, J.; Litwinek, D.; Żylińska-Urban, J.; Izak, D.; Buksa, K.; Gawor, J.; Gromadka, R.; Bardowski, J.K.; Kowalczyk, M. Bacterial community dynamics in spontaneous sourdoughs made from wheat, spelt, and rye wholemeal flour. Microbiol. Open 2020, 9, e1009. [Google Scholar] [CrossRef] [PubMed]
- Chavan, R.S.; Chavan, S.R. Sourdough technology—A traditional way for wholesome foods: A review. Compr. Rev. Food Sci. Food Saf. 2011, 10, 169–182. [Google Scholar] [CrossRef]
- Lau, S.W.; Chong, A.Q.; Chin, N.L.; Talib, R.A.; Basha, R.K. Sourdough Microbiome Comparison and Benefits. Microorganisms 2021, 9, 1355. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G.; Ehmann, M.; Hammes, W.P. Modeling of growth of Lactobacillus sanfranciscensis and Candida milleri in response to process parameters of sourdough fermentation. Appl. Environ. Microbiol. 1998, 64, 2616–2623. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Lopez, M.L.; Prieto, M.; Otero, A. The physiological attributes of Gram-negative bacteria associated with spoilage of meat and meat products. In Microbiology of Meat and Poultry; Davies, A.R., Board, R.J., Board, R.G., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1998; pp. 1–34. [Google Scholar]
- Vogel, R.F.; Ehrmann, M.A.; Gänzle, M.G. Development and potential of starter lactobacilli resulting from exploration of the sourdough ecosystem. Antonie Van Leeuwenhoek 2002, 81, 631–638. [Google Scholar] [CrossRef]
- Guarcello, R.; De Angelis, M.; Settanni, L.; Formiglio, S.; Gaglio, R.; Minervini, F.; Moschetti, G.; Gobbetti, M. Selection of amine-oxidizing dairy lactic acid bacteria and identification of the enzyme and gene involved in the decrease of biogenic amines. Appl. Environ. Microbiol. 2016, 82, 6870–6880. [Google Scholar] [CrossRef] [Green Version]
- Fidanza, F.; Alberti, A.; Fruttini, D. The Nicotera diet: The reference Italian Mediterranean diet. World Rev. Nutr. Diet. 2005, 95, 115–121. [Google Scholar]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The application of a plant biostimulant based on seaweed and yeast extract improved tomato fruit development and quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claesson, M.J.; Wang, Q.; O’Sullivan, O.; Greene-Diniz, R.; Cole, J.R.; Ross, R.P.; O’Toole, P.W. Comparison of two next-generation sequencing technologies for resolving highly complex microbiota composition using tandem variable 16S rRNA gene regions. Nucleic Acids Res. 2010, 38, e200. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Kline, L.; Sugihara, T.F. Microorganisms of the San Francisco sour dough bread process: II. Isolation and characterization of undescribed bacterial species responsible for the souring activity. Appl. Microbiol. 1971, 21, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Vogel, R.F.; Bocker, G.; Stolz, P.; Ehrmann, M.; Fanta, D.; Ludwig, W.; Pot, B.; Kersters, K.; Schleifer, K.H.; Hammes, W.P. Identification of lactobacilli from sourdough and description of Lactobacillus pontis sp. nov. Int. J. Syst. Evol. Microbiol. 1994, 44, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Martorana, A.; Alfonzo, A.; Settanni, L.; Corona, O.; La Croce, F.; Caruso, T.; Moschetti, G.; Francesca, N. An innovative method to produce green table olives based on “pied de cuve” technology. Food Microbiol. 2015, 50, 126–140. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Sample | Biogenic Amine Content (HE) ppm |
|---|---|
| 1BL | 62.647 ± 5.407 |
| 2BL | 41.176 ± 2.912 |
| 3BL | 43.600 ± 1.664 |
| 4BL | <LOD |
| 5BL | <LOD |
| 6BL | <LOD |
| 7BL | <LOD |
| 8BL | <LOD |
| 9BL | <LOD |
| 10BL | <LOD |
| 1HM | 251.764 ± 23.292 |
| 2HM | 95.882 ± 2.496 |
| 3HM | 91.390 ± 8.943 |
| 4HM | <LOD |
| 5HM | <LOD |
| 6HM | <LOD |
| 7HM | <LOD |
| 8HM | <LOD |
| 9HM | <LOD |
| 10HM | <LOD |
| Biogenic Amine | Samples | Statistical Significance 1 | |||||
|---|---|---|---|---|---|---|---|
| 1BL | 2BL | 3BL | 1HM | 2HM | 3HM | ||
| Histamine | <LOD b | <LOD b | <LOD b | 6.733 ± 0.201 a | <LOD b | <LOD b | *** |
| Putrescine | 8.831 ± 0.524 c | 14.063 ± 0.584 b | 9.001 ± 0.649 c | 37.364 ± 1.239 a | 15.981 ± 0.923 b | 11.702 ± 0.634 c | *** |
| Cadaverine | <LODa | <LOD a | <LOD a | <LOD a | <LODa | <LODa | N.S. |
| 2-Phenylethylamine | <LOD b | <LOD b | <LOD b | 0.299 ± 0.012 a | <LOD b | <LOD b | *** |
| Tyramine | 0.178 ± 0.004 c | 0.307 ± 0.004 c | 0.354 ± 0.021 c | 112.583 ± 6.377 a | 4.634 ± 0.398 c | 47.874 ± 3.203 b | *** |
| Triptamine | <LOD d | <LOD d | <LOD d | 0.258 ± 0.002 c | 24.234 ± 0.318 a | 0.711 ± 0.038 b | *** |
| Spermidine | 10.205 ± 0.056 b | 6.815 ± 0.084 c | 9.553 ± 0.366 bc | 10.294 ± 0.046 b | 20.557 ± 2.649 a | 6.587 ± 0.383 c | *** |
| Spermine | 6.323 ± 0.09 c | 2.666 ± 0.114 f | 5.108 ± 0.055 d | 8.879 ± 0.142 b | 14.287 ± 0.132 a | 3.471 ± 0.064 e | *** |
| Sample | pH | TTA |
|---|---|---|
| 1BL | 4.74 ± 0.01 a | 8.80 ± 0.30 d |
| 2BL | 4.83 ± 0.01 a | 3.92 ± 0.40 e |
| 3BL | 4.20 ± 0.01 b | 9.65 ± 0.55 d |
| 1HM | 3.87± 0.01 c | 15.80 ± 0.30 b |
| 2HM | 3.78 ± 0.09 cd | 27.40 ± 0.40 a |
| 3HM | 3.69 ± 0.11 d | 14.05 ± 0.65 c |
| Statistical significance | *** | *** |
| Sample | Microbial Loads | |||||||
|---|---|---|---|---|---|---|---|---|
| PCA | mMRS | M17 | SDB | SFM | VRBGA | PAB | YPD | |
| 1BL | 5.77 ± 0.19 c | 4.86 ± 0.13 c | <2 c | 5.80 ± 0.15 c | <2 e | <1 a | <2 a | <2 c |
| 2BL | 6.86 ± 0.17 b | 6.51 ± 0.18 b | 6.23 ± 0.10 b | 6.83 ± 0.21 b | 6.59 ± 0.11 c | <1 a | <2 a | 5.33 ± 0.14 ab |
| 3BL | 5.06 ± 0.17 d | 4.29 ± 0.17 d | <2 c | 4.09 ± 0.11 d | 4.21 ± 0.13 d | <1 a | <2 a | <2 c |
| 1HM | 7.54 ± 0.16 a | 7.58 ± 0.12 a | 7.35 ± 0.18 a | 7.46 ± 0.19 a | 7.50 ± 0.10 a | <1 a | <2 a | 5.09 ± 0.13 b |
| 2HM | 6.69 ± 0.17 b | 6.53 ± 0.13 b | <2 c | 6.50 ± 0.13 b | 6.39 ± 0.15 c | <1 a | <2 a | 5.49 ± 0.19 a |
| 3HM | 7.76 ± 0.09 a | 7.50 ± 0.11 a | 7.57 ± 0.18 a | 7.28 ± 0.12 a | 7.20 ± 0.10 b | <1 a | <2 a | <2 c |
| Statistical significance | *** | *** | *** | *** | *** | N.S. | N.S. | *** |
| Sample | City (Province) 1 | Final Product | Age 2 (Years) | Type of Flour | DY | DY Factors 3 | Fermentation Temperature | Fermentation Duration (h) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Sourdough Inoculum (g) | Flour/ Semolina (g) | Water (mL) | ||||||||
| 1BL | Palermo (PA) | Bread | 1 | Tender flour | 275 | 100 | 100 | 75 | 22 °C | 15 |
| 2BL | Piana degli Albanesi (PA) | Bread | 2 | Durum semolina | 140 | 100 | 500 | 100 | ambient | 24 |
| 3BL | Piana degli Albanesi (PA) | Bread | 3 | Durum semolina | 160 | 100 | 1000 | 500 | 20 °C | 48 |
| 4BL | Cinisi (PA) | Pastry | 1 | Tender flour | 246 | 100 | 100 | 46 | 28 | 4 |
| 5BL | Palermo | Pastry | 70 | Tender flour | 225 | 100 | 120 | 50 | 28 | 3.5 |
| 6BL | Carini (PA) | Pizza | 1 | Tender flour | 400 | 100 | 50 | 50 | ambient | 12 |
| 7BL | Castronovo (PA) | Bread | 6 | Durum semolina | 250 | 100 | 100 | 50 | ambient | 8 |
| 8BL | Piana degli Albanesi (PA) | Bread | 4 | Durum semolina | 400 | 100 | 50 | 50 | 22 | 6 |
| 9BL | Castelbuono (PA) | Pizza | 50 | Tender flour | 300 | 100 | 100 | 100 | ambient | 24 |
| 10BL | Piana degli Albanesi (PA) | Bread | 60 | Durum semolina | 185 | 100 | 1000 | 750 | 22 | 4 |
| 1HM | Catania (CT) | Bread | 2 | Flour 50%/semolina 50% | 300 | 100 | 60 | 20 | ambient | 24 |
| 2HM | Alcamo (TP) | Focaccia pizza | >100 | Tender flour | 350 | 100 | 50 | 25 | ambient | 10 |
| 3HM | Rocca di Caprileone (ME) | Pizza | >100 | Flour 40%/semolina 60% | 230 | 100 | 135 | 75 | ambient | 12 |
| 4HM | Piana degli Albanesi (PA) | Bread | 1 | Tender flour | 350 | 100 | 50 | 25 | ambient | 24 |
| 5HM | Modica (RG) | Bread | 3 | Durum semolina | 260 | 100 | 100 | 60 | 20 | 12 |
| 6HM | Villabate (PA) | Bread | 2 | Durum semolina | 250 | 100 | 100 | 50 | ambient | 10 |
| 7HM | Sciacca (AG) | Bread | 40 | Durum semolina | 250 | 100 | 100 | 50 | ambient | 12 |
| 8HM | Sciacca (AG) | Pizza | 60 | Durum semolina | 250 | 100 | 100 | 50 | ambient | 18 |
| 9HM | Camporeale (PA) | Bread | 30 | Wholemeal flour | 400 | 100 | 50 | 50 | 8 | 120 |
| 10HM | Naso (ME) | Pastry | >40 | Durum semolina | 225 | 100 | 360 | 350 | ambient | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, G.; Cirlincione, F.; Gaglio, R.; Franciosi, E.; Francesca, N.; Moschetti, G.; Asteggiano, A.; Medana, C.; Gentile, C.; Settanni, L. Preliminary Investigation of Biogenic Amines in Type I Sourdoughs Produced at Home and Bakery Level. Toxins 2022, 14, 293. https://doi.org/10.3390/toxins14050293
Mannino G, Cirlincione F, Gaglio R, Franciosi E, Francesca N, Moschetti G, Asteggiano A, Medana C, Gentile C, Settanni L. Preliminary Investigation of Biogenic Amines in Type I Sourdoughs Produced at Home and Bakery Level. Toxins. 2022; 14(5):293. https://doi.org/10.3390/toxins14050293
Chicago/Turabian StyleMannino, Giuseppe, Fortunato Cirlincione, Raimondo Gaglio, Elena Franciosi, Nicola Francesca, Giancarlo Moschetti, Alberto Asteggiano, Claudio Medana, Carla Gentile, and Luca Settanni. 2022. "Preliminary Investigation of Biogenic Amines in Type I Sourdoughs Produced at Home and Bakery Level" Toxins 14, no. 5: 293. https://doi.org/10.3390/toxins14050293
APA StyleMannino, G., Cirlincione, F., Gaglio, R., Franciosi, E., Francesca, N., Moschetti, G., Asteggiano, A., Medana, C., Gentile, C., & Settanni, L. (2022). Preliminary Investigation of Biogenic Amines in Type I Sourdoughs Produced at Home and Bakery Level. Toxins, 14(5), 293. https://doi.org/10.3390/toxins14050293