Mixtures of Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Whole-Plant Corn Silages and Total Mixed Rations of Dairy Farms in Central and Northern Mexico

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Main Dietary Ingredients
2.2. Occurrence and Concentrations of the Detected Metabolites
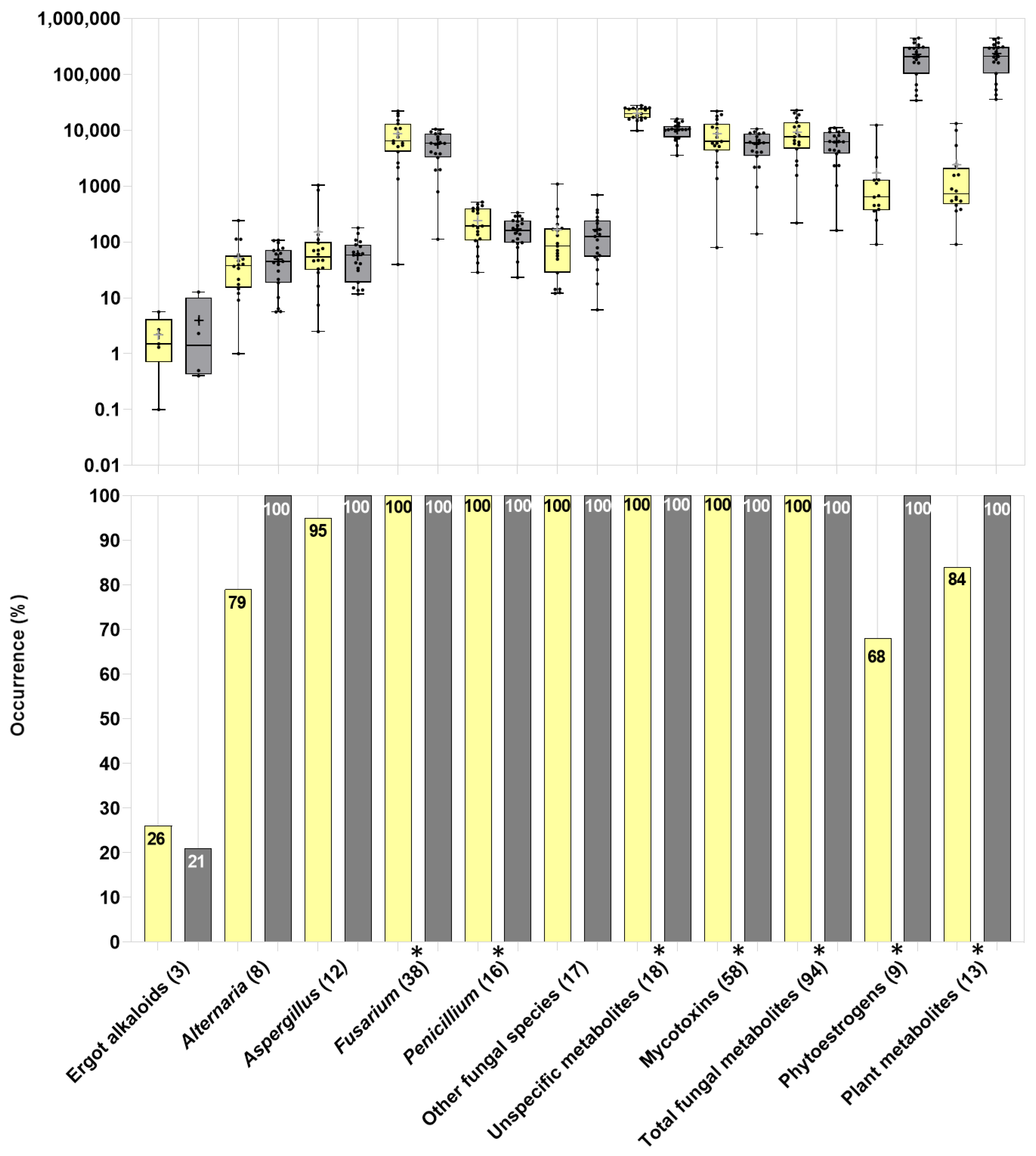
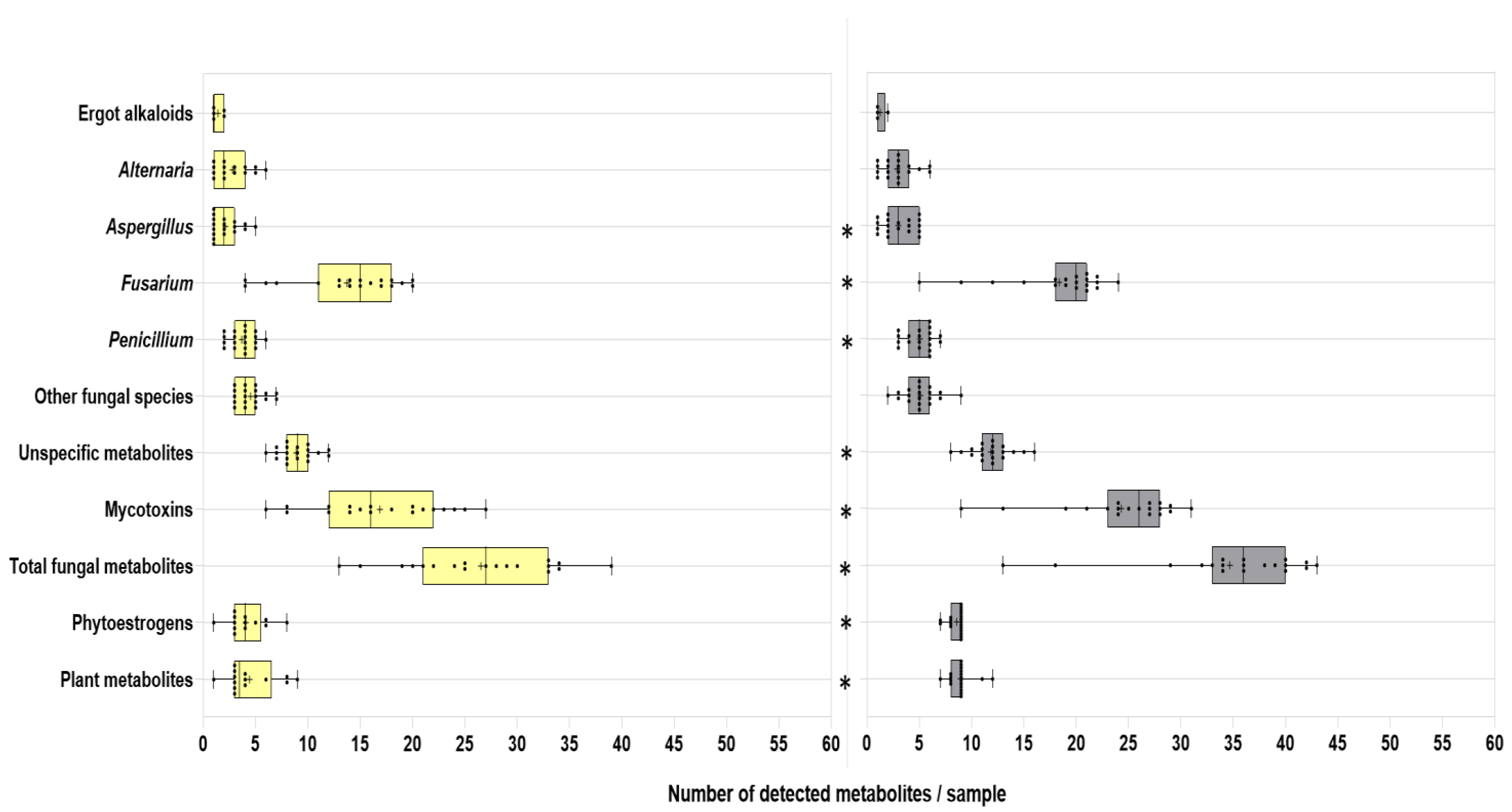
2.2.1. General Overview
2.2.2. Mycotoxins and Other Fungal Secondary Metabolites

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Metabolites | Metabolite | Positive Samples 1 (%) | Whole-Plant Corn Silages (n = 19) | Positive Samples 1 (%) | Total Mixed Rations (n = 19) | Wilcoxon Matched-Pairs Test | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Concentration (µg/kg DM) 2 | Concentration (µg/kg DM) 2 | p-Value * | ||||||||
| Average ± SD | Median | Range | Average ± SD | Median | Range | |||||
| Ergot alkaloids | Festuclavine + | 5 | – | – | 2.41 | 0 | – | – | – | >0.9999 |
| Dihydroergosine + | 26 | 1.35 ± 1.17 | 1.29 | 0.13–3.2 | 21 | 0.83 ± 0.97 | 0.44 | 0.18–2.28 | 0.0625 | |
| Chanoclavine + | 5 | – | – | 2.04 | 5 | – | – | 12.5 | >0.9999 | |
| Alternaria spp. | Altenuisol + | 32 | 2.5 ± 0 | 2.5 | 2.5–2.5 | 37 | 3.14 ± 1.7 | 2.5 | 2.5–6.99 | 0.7656 |
| Alternariol + | 5 | – | – | 5.5 | 11 | 9.77 ± 6.04 | 9.77 | 5.5–14 | 0.75 | |
| Alternariolmethylether + | 47 | 9.89 ± 7.59 | 5.5 | 5.5–27.4 | 42 | 6.39 ± 2.51 | 5.5 | 5.5–12.6 | 0.25 | |
| Altersetin + | 26 | 6.76 ± 4.5 | 5.16 | 1.25–12.7 | 42 | 15.7 ± 9.86 | 12.3 | 4.18–34.3 | 0.0488 | |
| Infectopyron | 21 | 97 ± 64 | 94 | 23.9–176 | 16 | 34.2 ± 3.38 | 36.1 | 30.3–36.2 | 0.1875 | |
| Macrosporin + | 16 | 3.75 ± 0 | 3.75 | 3.75–3.75 | 11 | 3.75 ± 0 | 3.75 | 3.75–3.75 | >0.9999 | |
| Tentoxin + | 42 | 7.71 ± 4.86 | 6.38 | 3.1–16 | 79 | 6.91 ± 2.87 | 6.41 | 2.48–11.3 | 0.0932 | |
| Tenuazonic acid + | 32 | 40.2 ± 10.4 | 37.5 | 30.1–60.4 | 53 | 49.4 ± 16.8 | 41.8 | 30.3–82.8 | 0.064 | |
| Aspergillus spp. | Averufin + | 42 | 3.6 ± 1.9 | 3.0 | 3.0–8.4 | 26 | 2.95 ± 0 | 2.95 | 2.95–2.95 | 0.125 |
| Deoxygerfelin | 0 | – | – | – | 11 | 2.41 ± 1.33 | 2.41 | 1.47–3.35 | 0.5 | |
| Flavoglaucin + | 11 | 2.8 ± 0.97 | 2.8 | 2.11–3.49 | 100 | 40.7 ± 29.6 | 41.6 | 3.63–111 | <0.0001 | |
| Fumigaclavine C + | 5 | – | – | 47.2 | 0 | – | – | – | >0.9999 | |
| Fumiquinazolin D + | 0 | – | – | – | 11 | 11.8 ± 5.34 | 11.8 | 8.01–15.6 | 0.5 | |
| Kojic acid + | 11 | 877 ± 130 | 877 | 785–69 | 5 | – | – | 145 | 0.5 | |
| Kotanin A | 11 | 2.5 ± 0 | 2.5 | 2.5–2.5 | 5 | – | – | 2.50 | >0.9999 | |
| Methylsulochrin | 5 | – | – | 4.5 | 11 | 4.5 ± 0 | 4.5 | 4.5–4.5 | >0.9999 | |
| Phenopyrrozin | 84 | 56.1 ± 28.1 | 53.1 | 16.2–132 | 79 | 12.4 ± 5.14 | 10.7 | 7.16–24.1 | <0.0001 | |
| seco-Sterigmatocystin + | 16 | 2.72 ± 1.8 | 3.58 | 0.65–3.91 | 42 | 0.9 ± 0.46 | 0.65 | 0.65–1.71 | >0.9999 | |
| Sterigmatocystin + | 0 | – | – | 11 | 2.65 ± 0 | 2.65 | 2.65–2.65 | 0.5 | ||
| Versicolorin C | 16 | 6.05 ± 3.98 | 3.75 | 3.75–10.6 | 0 | – | – | – | 0.25 | |
| Fusarium spp. | 15-Acetyldeoxynivalenol + | 11 | 142 ± 46.7 | 142 | 109–175 | 0 | – | – | – | 0.5 |
| 15-Hydroxyculmorin + | 32 | 2090 ± 1510 | 1580 | 464–4410 | 26 | 1270 ± 195 | 1280 | 993–1510 | 0.1563 | |
| Acuminatum B + | 32 | 151 ± 89.6 | 142 | 58.3–290 | 26 | 52.2 ± 21.8 | 55.8 | 27.6–83.2 | 0.1094 | |
| Antibiotic Y | 5 | – | – | 9.5 | 5 | – | – | 9.5 | >0.9999 | |
| Apicidin + | 16 | 7.14 ± 2.32 | 7.23 | 4.78–9.41 | 5 | – | – | 9.04 | 0.5 | |
| Aurofusarin + | 68 | 168 ± 386 | 48.8 | 3–1420 | 84 | 83.4 ± 67.3 | 67.1 | 11.4–224 | 0.2247 | |
| Beauvericin + | 100 | 57.8 ± 74.7 | 32.3 | 5.46–330 | 100 | 33.1 ± 24.3 | 29.1 | 3.84–84.2 | 0.2266 | |
| Beauvericin A + | 89 | 0.96 ± 1.55 | 0.45 | 0.45–6.87 | 84 | 0.55 ± 0.27 | 0.45 | 0.45–1.42 | 0.0204 | |
| Bikaverin + | 95 | 224 ± 253 | 99.4 | 15.3–879 | 100 | 115 ± 95.6 | 94.7 | 18.1–308 | 0.0204 | |
| Chrysogin + | 0 | – | – | – | 5 | – | – | 8.03 | >0.9999 | |
| Culmorin + | 58 | 865 ± 695 | 634 | 150–2090 | 58 | 505 ± 427 | 402 | 150–1420 | 0.0234 | |
| Deoxyfusapyron | 11 | 22 ± 12 | 22 | 13.5–30.5 | 16 | 591 ± 603 | 521 | 26.2–1230 | 0.375 | |
| Deoxynivalenol + | 53 | 1500 ± 1080 | 1370 | 323–3350 | 84 | 615 ± 491 | 376 | 78–1670 | 0.1928 | |
| DON-3-glucoside + | 26 | 74 ± 95.5 | 19.5 | 19.5–240 | 37 | 60.3 ± 23.5 | 65 | 19.5–86.6 | 0.3984 | |
| Enniatin A + | 11 | 1.02 ± 1.15 | 1.02 | 0.2–1.83 | 37 | 0.45 ± 0.37 | 0.2 | 0.2–1.19 | 0.3438 | |
| Enniatin A1 + | 11 | 0.4 ± 0 | 0.4 | 0.4–0.4 | 79 | 1.03 ± 0.82 | 0.4 | 0.4–2.6 | 0.0002 | |
| Enniatin B + | 0 | – | – | – | 68 | 4.63 ± 5.62 | 1.4 | 1.4–18.8 | 0.0002 | |
| Enniatin B1 + | 11 | 1.45 ± 0 | 1.45 | 1.45–1.45 | 89 | 3.99 ± 3.78 | 1.45 | 1.45–12.2 | <0.0001 | |
| Enniatin B2 + | 0 | – | – | – | 16 | 0.29 ± 0.05 | 0.29 | 0.24–0.34 | 0.25 | |
| Epiequisetin + | 16 | 3.37 ± 1.6 | 3.78 | 1.6–4.72 | 5 | – | – | 1.6 | 0.375 | |
| Equisetin + | 32 | 6.3 ± 6.27 | 3.05 | 1.6–14.7 | 42 | 5.19 ± 2.46 | 4.17 | 2.36–9.58 | 0.6836 | |
| Fumonisin A1 precursor + | 16 | 63.2 ± 40.5 | 61.2 | 23.7–105 | 58 | 14.3 ± 14.1 | 9.16 | 3.55–48.9 | 0.3096 | |
| Fumonisin A2 + | 11 | 43 ± 3.91 | 43 | 40.2–45.8 | 5 | – | – | 18 | 0.5 | |
| Fumonisin B1 + | 47 | 723 ± 1050 | 124 | 26.5–2700 | 84 | 218 ± 244 | 126 | 26.5–1010 | 0.2288 | |
| Fumonisin B2 + | 42 | 301 ± 371 | 72.6 | 18–987 | 68 | 103 ± 100 | 61.5 | 18–395 | 0.7722 | |
| Fumonisin B3 + | 16 | 276 ± 145 | 297 | 121–409 | 32 | 57.1 ± 41.8 | 40 | 26.5–131 | >0.9999 | |
| Fumonisin B4 + | 16 | 78.3 ± 70.5 | 61 | 18–156 | 32 | 26.1 ± 20 | 18 | 18–66.9 | 0.5625 | |
| Fusarium spp. | Fungerin | 0 | – | – | – | 5 | – | – | 26.5 | >0.9999 |
| Fusaproliferin + | 37 | 403 ± 628 | 166 | 61.5–1820 | 58 | 280 ± 252 | 226 | 60.8–989 | 0.3054 | |
| Fusapyron + | 5 | – | – | 1.5 | 5 | – | – | 5.46–5.46 | >0.9999 | |
| Fusaric acid + | 89 | 1210 ± 840 | 1130 | 260–3220 | 74 | 562 ± 235 | 503 | 298–1190 | <0.0001 | |
| Hydrolysed Fumonisin B1 + | 16 | 37 ± 49.1 | 10 | 7.29–93.7 | 5 | – | – | 30.4 | 0.75 | |
| Moniliformin + | 89 | 88.9 ± 76.9 | 48 | 9–263 | 100 | 101 ± 67 | 78.8 | 27.6–247 | 0.1956 | |
| Nivalenol + | 42 | 269 ± 184 | 209 | 103–614 | 68 | 872 ± 853 | 385 | 88.5–2600 | 0.0061 | |
| Sambutoxin + | 37 | 0.37 ± 0.19 | 0.3 | 0.3–0.79 | 5 | – | – | 0.3 | 0.0625 | |
| Siccanol + | 89 | 4620 ± 3530 | 3960 | 525–12,350 | 95 | 2510 ± 1650 | 2370 | 409–6130 | 0.0028 | |
| W493 | 79 | 171 ± 190 | 80.7 | 3.55–694 | 74 | 86.6 ± 65.4 | 101 | 3.55–190 | 0.0256 | |
| Zearalenone + | 68 | 58.7 ± 79.4 | 21.5 | 4.6–278 | 100 | 38.7 ± 57.2 | 17.8 | 4.6–246 | 0.9297 | |
| Total enniatins | 47 | 5.96 ± 7.24 | 1.60 | 0.60–19 | 89 | 11.2 ± 9.8 | 7.11 | 1.85–37 | 0.0144 | |
| Total fumonisins | 47 | 1150 ± 1570 | 203 | 26.5–4410 | 89 | 325 ± 396 | 155 | 3.6–1670 | 0.3867 | |
| Total Type B trichothecenes | 53 | 2000 ± 1230 | 1790 | 323–4230 | 89 | 1940 ± 1760 | 1156 | 78.0–5510 | 0.0505 | |
| Penicillium spp. | 7-Hydroxypestalotin | 53 | 17.3 ± 9.99 | 14.9 | 7.3–41.9 | 47 | 9.74 ± 4.87 | 9.74 | 2.6–16.7 | 0.0186 |
| Asterric acid | 5 | – | – | 12.5 | 5 | – | – | 12.5 | N/A | |
| Bilaid A | 100 | 20.3 ± 22.9 | 11.4 | 5.78–87.6 | 95 | 8.53 ± 7.04 | 6.77 | 3.49–27.3 | <0.0001 | |
| Citreoviridin + | 0 | – | – | – | 21 | 42.9 ± 12.2 | 41.3 | 31.1–58 | 0.125 | |
| Citrinin + | 0 | – | – | – | 5 | – | – | 77.9 | >0.9999 | |
| Cycloaspeptide A | 0 | – | – | – | 5 | – | – | 13.4 | >0.9999 | |
| Cyclopenin | 5 | – | – | 2.85 | 0 | – | – | – | >0.9999 | |
| Mycophenolic acid + | 11 | 90.2 ± 118 | 90.2 | 7–173 | 42 | 32 ± 42.9 | 11.4 | 7–127 | 0.1094 | |
| Mycophenolic acid IV + | 5 | – | – | 2.53 | 0 | – | – | – | >0.9999 | |
| NP 1243 | 5 | – | – | 34.1 | 0 | – | – | – | >0.9999 | |
| Oxaline | 16 | 68.9 ± 61.2 | 81.2 | 2.55–123 | 16 | 20.1 ± 14.9 | 12.9 | 10–37.2 | 0.5 | |
| Pestalotin | 53 | 29.2 ± 13.5 | 28.7 | 8.61–59.2 | 58 | 12.4 ± 6.93 | 11.2 | 3.3–24.5 | 0.0282 | |
| PF 1163A | 5 | – | – | 3.32 | 5 | – | – | 0.75 | >0.9999 | |
| Questiomycin | 5 | – | – | 1.5 | 89 | 8.71 ± 7.72 | 8.6 | 0.6–23 | <0.0001 | |
| Questiomycin Derivate | 95 | 184 ± 111 | 164 | 34.9–407 | 95 | 118 ± 64.2 | 106 | 18.1–238 | 0.0002 | |
| Quinolactacin A | 11 | 1.2 ± 0 | 1.2 | 1.2–1.2 | 21 | 1.2 ± 0 | 1.2 | 1.2–1.2 | 0.5 | |
| Other fungi | Ascochlorin | 21 | 11.9 ± 10.2 | 8.43 | 3.75–26.9 | 21 | 6.24 ± 4.99 | 3.75 | 3.75–13.7 | 0.625 |
| Ascofuranone | 21 | 2.26 ± 1.82 | 1.35 | 1.35–4.98 | 5 | – | – | 1.35 | 0.3125 | |
| Bassianolide | 37 | 3.17 ± 1.25 | 2.7 | 2.7–6 | 32 | 2.7 ± 0 | 2.7 | 2.7–2.7 | 0.5 | |
| Beauveriolide I_III | 26 | 1.5 ± 0 | 1.5 | 1.5–1.5 | 16 | 4.22 ± 3.01 | 3.71 | 1.5–7.45 | 0.6563 | |
| Cercosporin | 58 | 40.8 ± 25.6 | 36.5 | 13.2–87.9 | 79 | 72.2 ± 79.7 | 42.5 | 15.1–325 | 0.0479 | |
| Cytochalasin J | 0 | – | – | – | 11 | 136 ± 26.5 | 136 | 117–155 | 0.5 | |
| Destruxin B + | 0 | – | – | – | 21 | 1.25 ± 0.68 | 1.1 | 0.7–2.09 | 0.125 | |
| Ilicicolin A | 5 | – | – | 6.23 | 37 | 1.83 ± 0.61 | 1.6 | 1.6–3.21 | 0.2813 | |
| Ilicicolin B | 79 | 18.9 ± 20.7 | 4.45 | 4.45–69.4 | 89 | 14.1 ± 9.96 | 13 | 4.45–28.9 | 0.6848 | |
| Ilicicolin E | 5 | – | – | 1.7 | 11 | 1.7 ± 0 | 1.7 | 1.7–1.7 | >0.9999 | |
| Monocerin | 89 | 115 ± 237 | 37.4 | 2.1–990 | 74 | 85.9 ± 133 | 37.2 | 2.1–502 | 0.0024 | |
| Mycousnine | 0 | – | – | – | 11 | 0.75 ± 0 | 0.75 | 0.75–0.75 | 0.5 | |
| Myriocin + | 16 | 67.9 ± 52.1 | 48.1 | 28.6–127 | 32 | 44.6 ± 26.2 | 41.1 | 15.7–92.6 | 0.5625 | |
| Phomalone | 5 | – | – | 6.14 | 0 | – | – | – | >0.9999 | |
| Sporidesmolide II | 84 | 7.9 ± 13.2 | 2.92 | 0.75–44.7 | 74 | 4.4 ± 5.07 | 2.54 | 0.75–17.2 | 0.0643 | |
| Sporidesmolide III | 5 | 0.75 | 0.75 | 0.75 | 0 | – | – | – | >0.9999 | |
| Unspecific metabolites | 3-Nitropropionic acid | 21 | 63 ± 60.9 | 43 | 18.5–147 | 21 | 18.5 ± 0 | 18.5 | 18.5–18.5 | 0.5 |
| Asperglaucide | 5 | – | – | 5.99 | 100 | 27.3 ± 33.6 | 10.8 | 2.05–142 | <0.0001 | |
| Asperphenamate | 5 | – | – | 4.89 | 79 | 5.98 ± 7.37 | 3.35 | 1.93–31.4 | <0.0001 | |
| Brevianamid F | 89 | 171 ± 77.8 | 166 | 61–408 | 89 | 116 ± 40.6 | 112 | 49.2–228 | 0.0021 | |
| Chrysophanol | 47 | 226 ± 111 | 231 | 62.5–367 | 32 | 176 ± 65.1 | 205 | 62.5–226 | 0.0195 | |
| Citreorosein | 53 | 24.1 ± 12.4 | 19.1 | 14.7–54.4 | 37 | 19.1 ± 6.67 | 15.8 | 12.5–30.1 | 0.123 | |
| Cyclo(L-Pro-L-Tyr) | 100 | 4680 ± 2300 | 4570 | 926–8970 | 100 | 2180 ± 1110 | 1890 | 589–5360 | 0.0006 | |
| Cyclo(L-Pro-L-Val) | 100 | 14,760 ± 3820 | 13,450 | 6890–2200 | 100 | 7080 ± 2300 | 6790 | 2160–11,570 | <0.0001 | |
| Emodin | 95 | 9.62 ± 5.31 | 9.22 | 3.5–23.1 | 95 | 46.9 ± 102 | 8.49 | 3.5–422 | 0.2312 | |
| Fellutanine A | 95 | 128 ± 51.8 | 127 | 48.8–260 | 89 | 94.3 ± 38.7 | 86.7 | 34.8–199 | 0.0053 | |
| Iso-Rhodoptilometrin | 58 | 1.58 ± 0.59 | 1.4 | 1.4–3.35 | 53 | 1.4 ± 0 | 1.4 | 1.4–1.4 | 0.5 | |
| N-Benzoyl-Phenylalanine | 0 | – | – | – | 21 | 12.2 ± 2.47 | 12.1 | 9.56–15 | 0.125 | |
| Neoechinulin A | 0 | – | – | – | 100 | 133 ± 78.4 | 102 | 29.6–304 | <0.0001 | |
| Norlichexanthone | 5 | – | – | 1.9 | 47 | 1.9 ± 0 | 1.9 | 1.9 | 0.0078 | |
| Rugulusovine | 100 | 355 ± 153 | 373 | 137–681 | 100 | 204 ± 93.8 | 197 | 53.5–407 | <0.0001 | |
| Skyrin | 68 | 2.06 ± 1.07 | 1.85 | 0.55–3.96 | 89 | 4.42 ± 6.13 | 2.48 | 0.55–27 | 0.0097 | |
| Ternatin | 5 | – | – | 6.32 | 0 | – | – | – | >0.9999 | |
| Tryptophol | 42 | 258 ± 126 | 170 | 170–456 | 32 | 963 ± 817 | 642 | 170–2100 | 0.5508 | |
| Phytoestrogens | Biochanin | 5 | – | – | 147 | 79 | 36.3 ± 13 | 34.5 | 20.2–61.6 | 0.0081 |
| Coumestrol | 26 | 56 ± 104 | 8 | 8–241 | 89 | 157 ± 126 | 109 | 45.5–479 | 0.0011 | |
| Daidzein | 37 | 263 ± 351 | 89 | 89–1020 | 100 | 12,700 ± 6710 | 10,710 | 3820–27,620 | <0.0001 | |
| Daidzin | 68 | 428 ± 719 | 191 | 91–2730 | 100 | 63,690 ± 40,170 | 65,640 | 9350–125,770 | <0.0001 | |
| Genistein | 58 | 153 ± 272 | 47 | 47–947 | 100 | 11,760 ± 6170 | 11,190 | 3990–26,530 | <0.0001 | |
| Genistin | 63 | 1000 ± 1850 | 362 | 110–6700 | 100 | 118,150 ± 75,850 | 113,270 | 157,180–249,320 | <0.0001 | |
| Glycitein | 5 | – | – | 324 | 89 | 4790 ± 1840 | 4450 | 2220–8220 | <0.0001 | |
| Glycitin | 11 | 364 ± 292 | 364 | 158–570 | 100 | 13,340 ± 7920 | 12,070 | 1080–27,390 | <0.0001 | |
| Ononin | 5 | – | – | 46 | 100 | 176 ± 28 | 153.3 | 46–512 | <0.0001 | |
| Other plant metabolites | Abscisic acid | 42 | 1610 ± 2860 | 574 | 273–8670 | 100 | 1660 ± 636 | 1620 | 411–3270 | 0.0012 |
| Anisodamine | 16 | 514 ± 373 | 470 | 164–907 | 16 | 137.2 ± 101 | 141 | 34.5–236 | 0.375 | |
| Atropine | 16 | 318 ± 85 | 360 | 219–374 | 11 | 69.1 ± 22.4 | 69.1 | 53.3–84.9 | 0.25 | |
| Hyoscine | 16 | 427 ± 391 | 473 | 15–794 | 11 | 215.7 ± 93.1 | 216 | 150–282 | 0.375 | |
| Bacterial | Nonactin | 16 | 1 ± 0 | 1 | 1–1 | 26 | 1.3 ± 1.2 | 0.8 | 0.6–3.3 | 0.3906 |
2.2.3. Plant Secondary Metabolites (Phytoestrogens and Others)
2.2.4. Unspecific (Multi-Kingdom) and Bacterial Metabolites
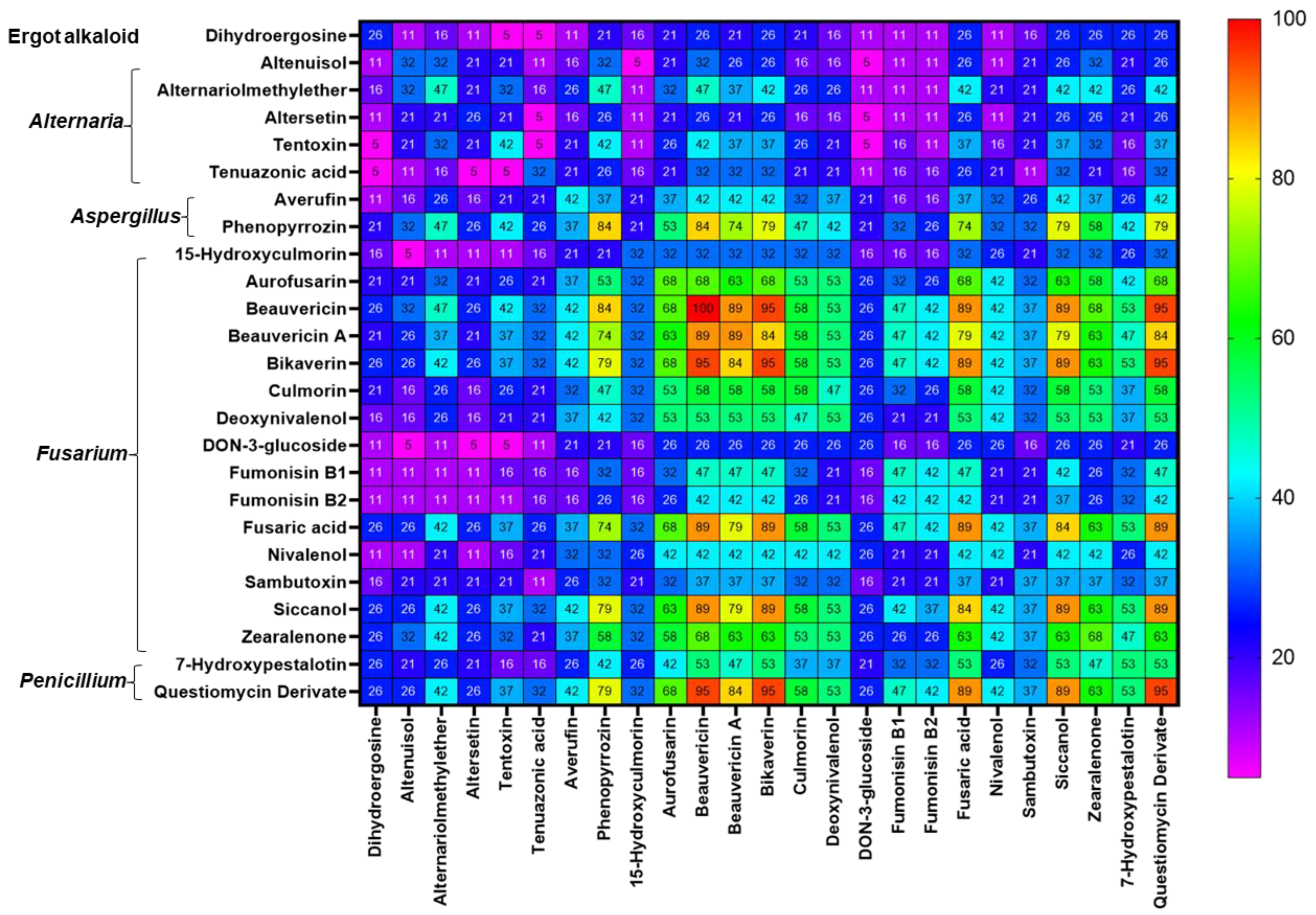
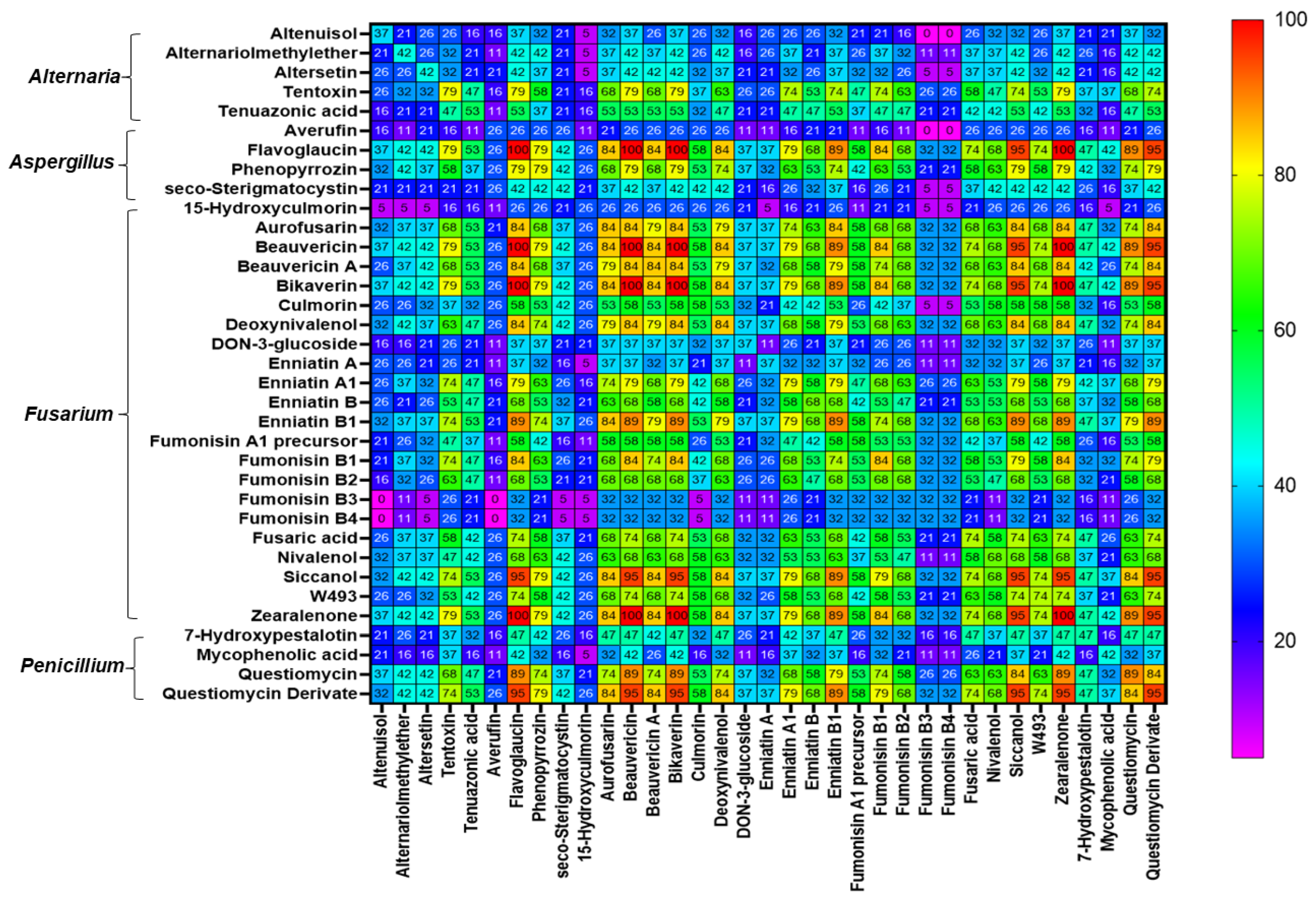
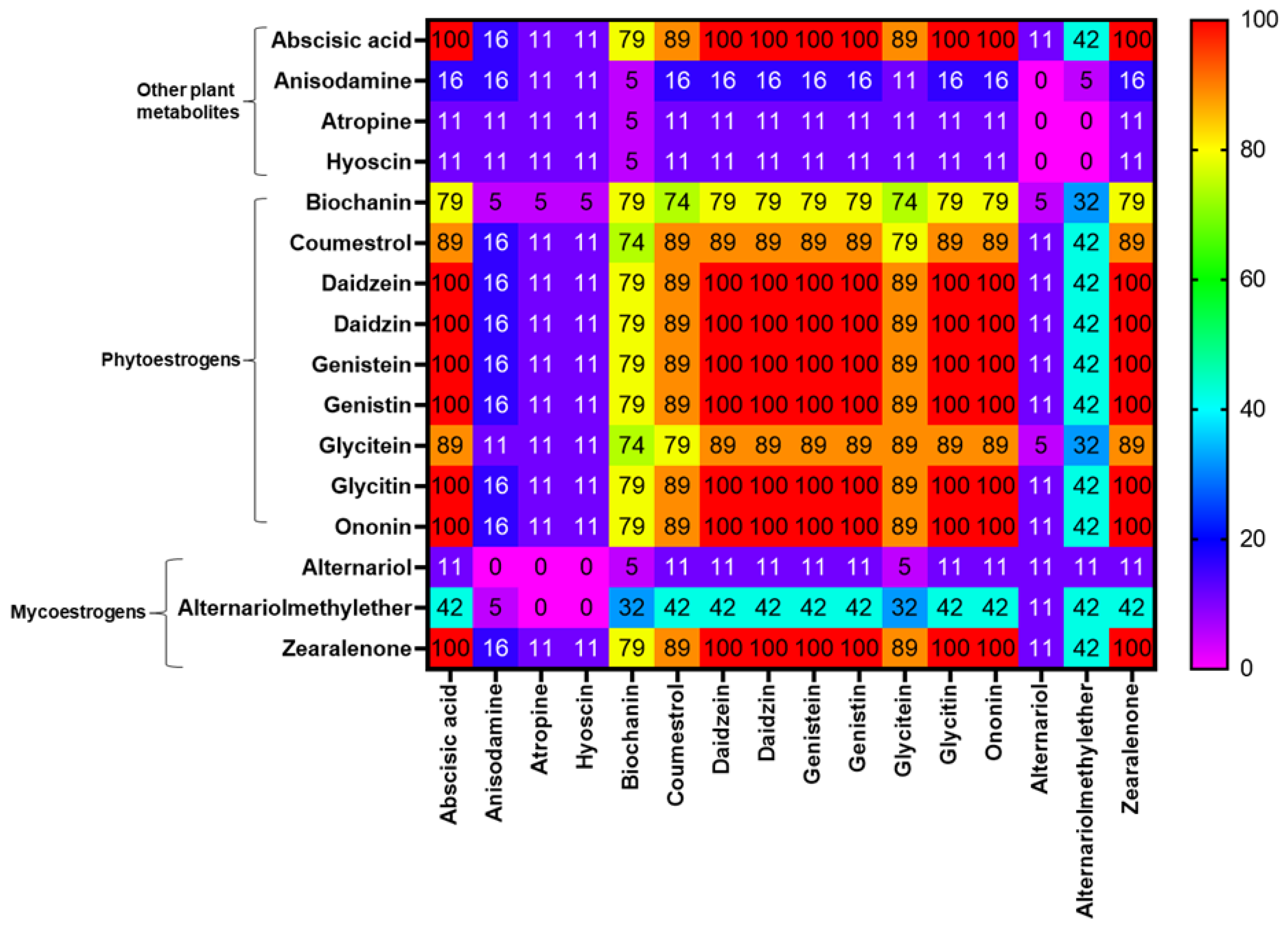
2.3. Co-occurrence of Mycotoxins and Phytoestrogens
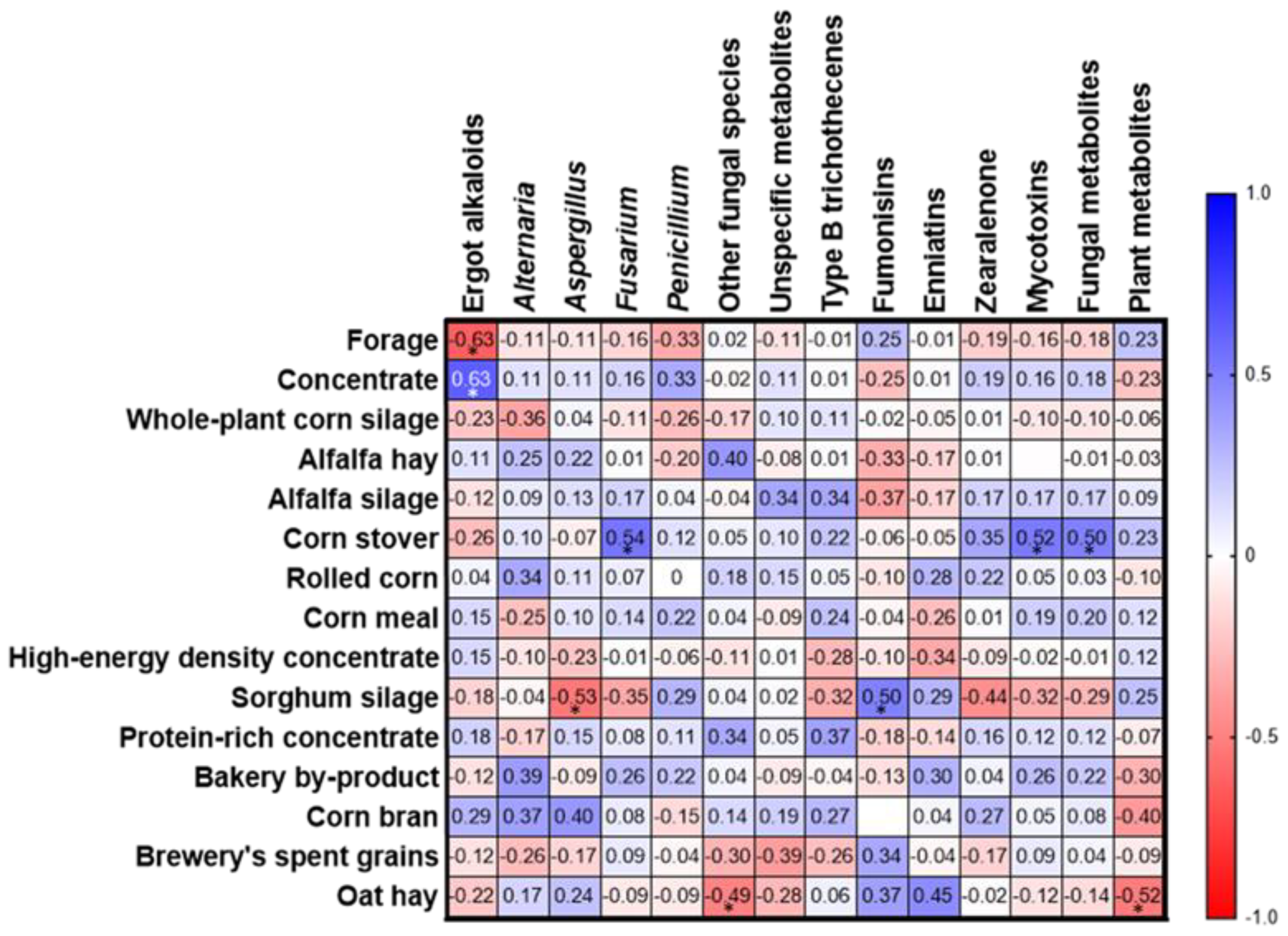
2.4. Relationship between Concentrations of Mycotoxin/Metabolite Groups and the Dietary Ingredients
3. Discussion
4. Conclusions
5. Materials and Methods
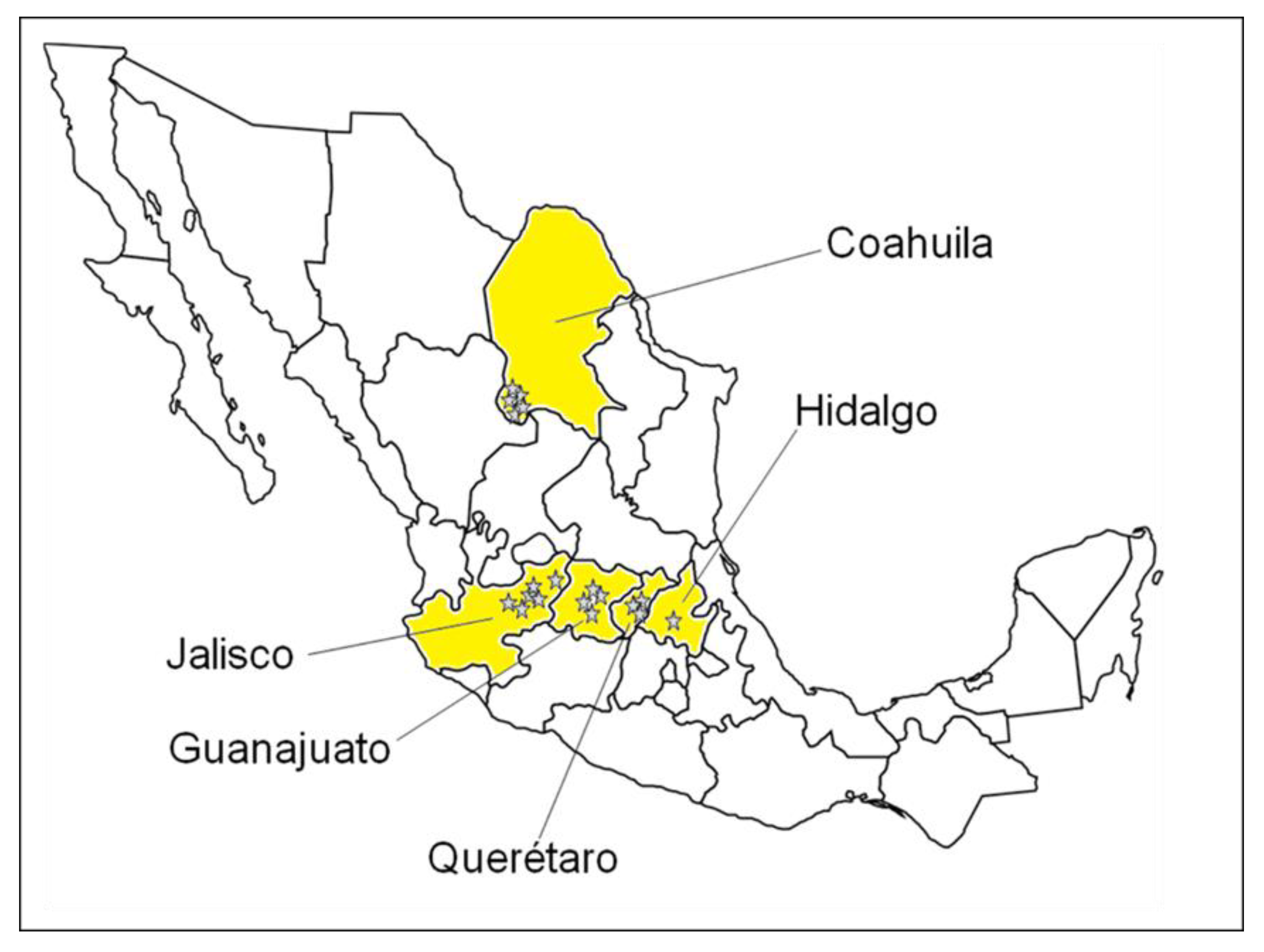
5.1. Sampling and Sample Preparation
5.2. Multi-Mycotoxin Analysis (LC-ESI–MS/MS)
5.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Dairy Market Review: Emerging Trends and Outlook, December 2021. Rome. 2021. Available online: https://www.fao.org/3/cb7982en/cb7982en.pdf (accessed on 25 January 2023).
- Penagos-Tabares, F.; Khiaosa-ard, R.; Schmidt, M.; Bartl, E.-M.; Kehrer, J.; Nagl, V.; Faas, J.; Sulyok, M.; Krska, R.; Zebeli, Q. Cocktails of Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Diets of Dairy Cows in Austria: Inferences from Diet Composition and Geo-Climatic Factors. Toxins 2022, 14, 493. [Google Scholar] [CrossRef] [PubMed]
- FAO; IDF; IFCN. World Mapping of Animal Feeding Systems in the Dairy Sector; FAO: Rome, Italy; IDF: Brussels, Belgium; IFCN: Rome, Italy, 2014; pp. 1–36. [Google Scholar]
- Battilani, P.; Palumbo, R.; Giorni, P.; Dall’Asta, C.; Dellafiora, L.; Gkrillas, A.; Toscano, P.; Crisci, A.; Brera, C.; De Santis, B. Mycotoxin mixtures in food and feed: Holistic, innovative, flexible risk assessment modelling approach: MYCHIF. EFSA Support. Publ. 2020, 17, 1757E. [Google Scholar] [CrossRef]
- Penagos-Tabares, F.; Khiaosa-Ard, R.; Nagl, V.; Faas, J.; Jenkins, T.; Sulyok, M.; Zebeli, Q. Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Austrian Pastures: Occurrences, Contamination Levels, and Implications of Geo-climatic Factors. Toxins 2021, 13, 460. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Asia Pac. J. Clin. Nutr. 2007, 16, 95–101. [Google Scholar] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, L.; Gong, G.; Zhang, L.; Shi, L.; Dai, J.; Han, Y.; Wu, Y.; Khalil, M.M.; Sun, L. Invited review: Remediation strategies for mycotoxin control in feed. J. Anim. Sci. Biotechnol. 2022, 13, 19. [Google Scholar] [CrossRef]
- Cinar, A.; Onbaşı, E. Mycotoxins: The hidden danger in foods. In Mycotoxins and Food Safety; IntechOpen: London, UK, 2020. [Google Scholar]
- Reyes-Velázquez, W.P.; Espinoza, V.H.I.; Rojo, F.; Jiménez-Plasencia, C.; Palacios, E.d.L.; Hernández-Góbora, J.; Ramírez-Álvarez, A. Occurrence of fungi and mycotoxins in corn silage, Jalisco State, Mexico. Rev. Iberoam. Micol. 2008, 25, 182–185. [Google Scholar]
- Álvarez-Días, F.; Torres-Parga, B.; Valdivia-Flores, A.G.; Quezada-Tristán, T.; Alejos-De La Fuente, J.I.; Sosa-Ramírez, J.; Rangel-Muñoz, E.J. Aspergillus flavus and Total Aflatoxins Occurrence in Dairy Feed and Aflatoxin M1 in Bovine Milk in Aguascalientes, Mexico. Toxins 2022, 14, 292. [Google Scholar] [CrossRef]
- Ortiz, C.M.F.; Portilla, L.B.H.; Medrano, J.V. Contaminación con micotoxinas en alimento balanceado y granos de uso pecuario en México en el año 2003. Rev. Mex. Cienc. Pecu. 2006, 44, 247–256. [Google Scholar]
- Betancourt, P.; Denise, S. Microbiota and mycotoxins in trilinear hybrid maize produced in natural environments at central region in Mexico. Adv. Microbiol. 2016, 6, 671–676. [Google Scholar] [CrossRef]
- Betancourt, S.D.P.; Ronquillo, J.C.C.; Manzano, E.P. Contaminación de Fusariotoxinas: Zearalenona, Fumonisinas y Deoxinivalenol en maíz (Zea mays L.), procedente de Puebla y de la Ciudad de México. Soc. Rural. Producción Medio Ambiente. 2020, 20, 16. [Google Scholar]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, L.; Xu, Z.; Liu, X.; Chen, L.; Dai, J.; Karrow, N.A.; Sun, L. Occurrence of Aflatoxin B1, Deoxynivalenol and Zearalenone in Feeds in China during 2018–2020. J. Anim. Sci. Biotechnol. 2021, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Commission, E. Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed. Luxemb. Off. J. Eur. Union. 2002, 140, 10–22. [Google Scholar]
- Commission, E. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (2006/576/EC). Off J Eur Union. 2006, 229, 7–9. [Google Scholar]
- Schingoethe, D.J. A 100-Year Review: Total mixed ration feeding of dairy cows. J. Dairy Sci. 2017, 100, 10143–10150. [Google Scholar] [CrossRef]
- Bueno, A.V.I.; Lazzari, G.; Jobim, C.C.; Daniel, J.L.P. Ensiling total mixed ration for ruminants: A review. Agronomy 2020, 10, 879. [Google Scholar] [CrossRef]
- Awapak, D.; Petchkongkaew, A.; Sulyok, M.; Krska, R. Co-occurrence and toxicological relevance of secondary metabolites in dairy cow feed from Thailand. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2021, 38, 1013–1027. [Google Scholar] [CrossRef]
- Rodriguez-Blanco, M.; Marin, S.; Sanchis, V.; Ramos, A.J. Fusarium mycotoxins in total mixed rations for dairy cows. Mycotoxin Res. 2020, 36, 277–286. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Toivonen, M.I. World Silage: A Survey of Forage Conservation around the World; Chalcombe Publications: Lincoln, UK, 2003. [Google Scholar]
- Alonso, V.A.; Pereyra, C.M.; Keller, L.A.M.; Dalcero, A.M.; Rosa, C.A.R.; Chiacchiera, S.M.; Cavaglieri, L.R. Fungi and mycotoxins in silage: An overview. J. Appl. Mirobiol. 2013, 115, 637–643. [Google Scholar] [CrossRef]
- Signorini, M.L.; Gaggiotti, M.; Molineri, A.; Chiericatti, C.A.; de Basilico, M.L.Z.; Basilico, J.C.; Pisani, M. Exposure assessment of mycotoxins in cow’s milk in Argentina. Food Chem. Toxicol. 2012, 50, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of mycotoxins in maize, grass and wheat silage for dairy cattle in the Netherlands. Food. Addit. Contam. Part B Surveill. 2008, 1, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Fink-Gremmels, J. Mycotoxin syndrome in dairy cattle: Characterisation and intervention results. World Mycotoxin J. 2014, 7, 357–366. [Google Scholar] [CrossRef]
- Botha, C.; Naude, T.; Moroe, M.; Rottinghaus, G. Gangrenous ergotism in cattle grazing fescue (Festuca elatior L.) in South Africa: Clinical communication. J. S. Afr. Vet. Assoc. 2004, 75, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Supriya, C.; Reddy, P.S. Prenatal exposure to aflatoxin B1: Developmental, behavioral, and reproductive alterations in male rats. Sci. Nat. 2015, 102, 26. [Google Scholar] [CrossRef]
- Gao, X.; Xiao, Z.; Li, C.; Zhang, J.; Zhu, L.; Sun, L.; Zhang, N.; Khalil, M.M.; Rajput, S.A.; Qi, D. Prenatal exposure to zearalenone disrupts reproductive potential and development via hormone-related genes in male rats. Food Chem. Toxicol. 2018, 16, 11–19. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Rudyk, H.; Wojtysiak, D.; Donaldson, J.; Muszyński, S.; Arciszewski, M.B.; Lisova, N.; Brezvyn, O.; Puzio, I.; Abramowicz, B.; et al. Basal Blood Morphology, Serum Biochemistry, and the Liver and Muscle Structure of Weaned Wistar Rats Prenatally Exposed to Fumonisins. Animals 2022, 12, 2353. [Google Scholar] [CrossRef]
- Kras, K.; Rudyk, H.; Muszyński, S.; Tomaszewska, E.; Dobrowolski, P.; Kushnir, V.; Muzyka, V.; Brezvyn, O.; Arciszewski, M.B.; Kotsyumbas, I. Morphology and Chemical Coding of Rat Duodenal Enteric Neurons following Prenatal Exposure to Fumonisins. Animals 2022, 12, 1055. [Google Scholar] [CrossRef]
- Smith, M.-C.; Madec, S.; Coton, E.; Hymery, N. Natural co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef]
- Wolawek-Potocka, I.; Bah, M.M.; Korzekwa, A.; Piskula, M.K.; Wiczkowski, W.; Depta, A.; Skarzynski, D.J. Soybean-derived phytoestrogens regulate prostaglandin secretion in endometrium during cattle estrous cycle and early pregnancy. Exp. Biol. Med. 2005, 230, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.F.M. Fertility of herbivores consuming phytoestrogen-containing Medicago and Trifolium species. Agriculture 2016, 6, 35. [Google Scholar] [CrossRef]
- Wocławek-Potocka, I.; Korzekwa, A.; Skarżyński, D. Can phytoestrogens pose a danger in the reproduction of cows? Med. Weter. 2008, 64, 515–519. [Google Scholar]
- Wocławek-Potocka, I.; Mannelli, C.; Boruszewska, D.; Kowalczyk-Zieba, I.; Waśniewski, T.; Skarżyński, D.J. Diverse effects of phytoestrogens on the reproductive performance: Cow as a model. Int. J. Endocrinol. 2013, 2013, 650984. [Google Scholar] [CrossRef]
- Penagos-Tabares, F.; Khiaosa-Ard, R.; Schmidt, M.; Pacífico, C.; Faas, J.; Jenkins, T.; Nagl, V.; Sulyok, M.; Labuda, R.; Zebeli, Q. Fungal species and mycotoxins in mouldy spots of grass and maize silages in Austria. Mycotoxin Res. 2022, 38, 117–136. [Google Scholar] [CrossRef]
- Penagos-Tabares, F.; Sulyok, M.; Nagl, V.; Faas, J.; Krska, R.; Khiaosa-ard, R.; Zebeli, Q. Mixtures of mycotoxins, phytoestrogens, and pesticides co-occurring in wet brewery’s spent grains (BSG) intended for dairy cattle feeding in Austria. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2022, 39, 1855–1877. [Google Scholar] [CrossRef]
- Kemboi, D.C.; Ochieng, P.E.; Antonissen, G.; Croubels, S.; Scippo, M.-L.; Okoth, S.; Kangethe, E.K.; Faas, J.; Doupovec, B.; Lindahl, J.F. Multi-Mycotoxin Occurrence in Dairy Cattle and Poultry Feeds and Feed Ingredients from Machakos Town, Kenya. Toxins 2020, 12, 762. [Google Scholar] [CrossRef]
- Nichea, M.J.; Cendoya, E.; Zachetti, V.G.L.; Chiacchiera, S.M.; Sulyok, M.; Krska, R.; Torres, A.M.; Chulze, S.N.; Ramirez, M.L. Mycotoxin profile of Fusarium armeniacum isolated from natural grasses intended for cattle feed. World Mycotoxin J. 2015, 8, 451–457. [Google Scholar] [CrossRef]
- Reisinger, N.; Schurer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin Occurrence in Maize Silage-A Neglected Risk for Bovine Gut Health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef]
- De la Federación, D.O. Norma Oficial Mexicana NOM-188-SSA1–2002, Productos y Servicios: Control de Aflatoxinas en Cereales para Consumo Humano y Animal, Especificaciones Sanitarias. D. Of. Fed. 2002. Available online: https://www.dof.gob.mx/nota_detalle.php?codigo=719385&fecha=15/10/2002 (accessed on 25 January 2023).
- De la Federación, D.O. Norma Oficial Mexicana NOM-247-SSA1-2008, Productos y servicios. Cereales y sus Productos. Cereales, h.d.c., Sémolas o Semolinas. Alimentos a Base de: Cereales, Semillas Comestibles, de Harinas, Sémolas o Semolinas o sus Mezclas. Productos de Panificación. Disposiciones y Especificaciones Sanitarias y Nutrimentales. Métodos de Prueba. D. Of. Fed. 2008. Available online: https://dof.gob.mx/nota_detalle.php?codigo=5100356&fecha=27/07/2009#gsc.tab=0 (accessed on 25 January 2023).
- Molina-Pintor, I.; Ruíz-Arias, M.; Guerrero-Flores, M.; Rojas-García, A.; Barrón-Vivanco, B.S.; Medina-Díaz, I.M.; Bernal-Hernández, Y.; Ortega-Cervantes, L.; Rodríguez-Cervantes, C.; Ramos, A. Preliminary survey of the occurrence of mycotoxins in cereals and estimated exposure in a northwestern region of Mexico. Int. J. Environ. Health Res. 2022, 32, 2271–2285. [Google Scholar] [CrossRef]
- US Food and Drug Administration (US FDA). Guidance for Industry and FDA: Advisory Levels for Deoxynivalenol (DON) in Finished Wheat Products for Human Consumption and Grains and Grain by-Products Used for Animal Feed. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-and-fda-advisory-levels-deoxynivalenol-don-finished-wheat-products-human#:~:text=The%20advisory%20levels%20for%20DON%20are%20as%20follows%3A,germ%2C%20that%20may%20potentially%20be%20consumed%20by%20humans. (accessed on 25 January 2023).
- US Food and Drug Administration (US FDA). Guidance for Industry: Action Levels for Poisonous or Deleterious Substances in Human Food and Animal Feed. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-action-levels-poisonous-or-deleterious-substances-human-food-and-animal-feed#afla (accessed on 25 January 2023).
- García, S.; Heredia, N. Mycotoxins in Mexico: Epidemiology, management, and control strategies. Mycopathologia 2006, 162, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Thapa, A.; Horgan, K.A.; White, B.; Walls, D. Deoxynivalenol and Zearalenone—Synergistic or Antagonistic Agri-Food Chain Co-Contaminants? Toxins 2021, 13, 561. [Google Scholar] [CrossRef] [PubMed]
- Szabó-Fodor, J.; Szabó, A.; Kócsó, D.; Marosi, K.; Bóta, B.; Kachlek, M.; Mézes, M.; Balogh, K.; Kövér, G.; Nagy, I. Interaction between the three frequently co-occurring Fusarium mycotoxins in rats. J. Anim. Physiol. Anim. Nutr. 2019, 103, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Kolf-Clauw, M.; Gauthier, T.; Abrami, R.; Abiola, F.A.; Oswald, I.P.; Puel, O. New insights into mycotoxin mixtures: The toxicity of low doses of Type B trichothecenes on intestinal epithelial cells is synergistic. Toxicol. Appl. Pharmacol. 2013, 272, 191–198. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Puel, O.; Pinton, P.; Cossalter, A.-M.; Chou, T.-C.; Oswald, I.P. Co-exposure to low doses of the food contaminants deoxynivalenol and nivalenol has a synergistic inflammatory effect on intestinal explants. Arch. Toxicol. 2017, 91, 2677–2687. [Google Scholar] [CrossRef]
- McKay, Z.C.; Averkieva, O.; Rajauria, G.; Pierce, K.M. The effect of feedborne Fusarium mycotoxins on dry matter intake, milk production and blood metabolites of early lactation dairy cows. Anim. Feed Sci. Technol. 2019, 253, 39–44. [Google Scholar] [CrossRef]
- Shimshoni, J.A.; Cuneah, O.; Sulyok, M.; Krska, R.; Galon, N.; Sharir, B.; Shlosberg, A. Mycotoxins in corn and wheat silage in Israel. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2013, 30, 1614–1625. [Google Scholar] [CrossRef]
- Hernandez-Valdivia, E.; Valdivia-Flores, A.; Cruz-Vazquez, C.; Martinez-Saldaña, M.; Quezada-Tristan, T.; Rangel-Muñoz, E.; Ortiz-Martinez, R.; Medina-Esparza, L.; Jaramillo-Juarez, F. Diagnosis of Subclinical Aflatoxicosis by Biochemical Changes in Dairy Cows under Field Conditions. Pak. Vet. J. 2021, 41, 33–38. [Google Scholar] [CrossRef]
- Jiménez-Pérez, C.; Alatorre-Santamaría, S.; Tello-Solís, S.; Gómez-Ruiz, L.; Rodríguez-Serrano, G.; García-Garibay, M.; Cruz-Guerrero, A. Analysis of aflatoxin M1 contamination in milk and cheese produced in Mexico: A review. World Mycotoxin J. 2021, 14, 269–285. [Google Scholar] [CrossRef]
- Betancourt, S.D.P.; Carranza, B.V.; Manzano, E.P. Estimation of mycotoxin multiple contamination in Mexican hybrid seed maize by HPLC-MS/MS. Agric. Sci. 2015, 6, 1089. [Google Scholar] [CrossRef]
- Carvajal, M.; Bolaños, A.; Rojo, F.; Mendez, I. Aflatoxin M1 in pasteurized and ultrapasteurized milk with different fat content in Mexico. J. Food Prot. 2003, 66, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Ehrlich, K.C.; Bland, J.M.; Montalbano, B.G. The aflatoxin biosynthesis cluster gene, aflX, encodes an oxidoreductase involved in conversion of versicolorin A to demethylsterigmatocystin. Appl. Environ. Microbiol. 2006, 72, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, D.; Lin, M.; Yao, R. Conversion of sterigmatocystin to aflatoxin B1 by Aspergillus parasiticus. Biochem. Biophys. Res. Commun. 1973, 52, 992–997. [Google Scholar] [CrossRef]
- McCormick, S.P.; Bhatnagar, D.; Lee, L.S. Averufanin is an aflatoxin B1 precursor between averantin and averufin in the biosynthetic pathway. Appl. Environ. Microbiol. 1987, 53, 14–16. [Google Scholar] [CrossRef]
- Lin, M.; Hsieh, D. Averufin in the biosynthesis of aflatoxin B1. J. Am. Chem. Soc. 1973, 95, 1668–1669. [Google Scholar] [CrossRef]
- Muaz, K.; Manzoor, S.; Akhtar, S.; Riaz, M.; Amir, M.; Akram, K.; Ismail, A. Aflatoxin Biosynthesis. In Aflatoxins in Food: A Recent Perspective; Hakeem, K.R., Oliveira, C.A.F., Ismail, A., Eds.; Springer: Cham, Switzerland, 2022; pp. 19–40. [Google Scholar]
- EFSA. Scientific Opinion on the risk for public and animal health related to the presence of sterigmatocystin in food and feed. EFSA J. 2013, 11, 3254. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging mycotoxins: Beyond traditionally determined food contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- Chuang, W.Y.; Hsieh, Y.C.; Lee, T.-T. The effects of fungal feed additives in animals: A review. Animals 2020, 10, 805. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Storm, I.; Rasmussen, R.R.; Rasmussen, P.H. Occurrence of Pre- and Post-Harvest Mycotoxins and Other Secondary Metabolites in Danish Maize Silage. Toxins 2014, 6, 2256–2269. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Giffel, M.C.T. Occurrence of Mycotoxins in Feedstuffs of Dairy Cows and Estimation of Total Dietary Intakes. J. Dairy Sci. 2008, 91, 4261–4271. [Google Scholar] [CrossRef] [PubMed]
- Parrish, F.; Wiley, B.; Simmons, E.; Long Jr, L. Production of aflatoxins and kojic acid by species of Aspergillus and Penicillium. Appl. Microbiol. 1966, 14, 139. [Google Scholar] [CrossRef] [PubMed]
- Morton, H.E.; Kocholaty, W.; Junowicz-Kocholaty, R.; Kelner, A. Toxicity and antibiotic activity of kojic acid produced by Aspergillus luteo-virescens. J. Bacteriol. 1945, 50, 579–584. [Google Scholar] [CrossRef]
- Kotani, T.; Ichimoto, I.; Tatsumi, C.; Fujita, T. Bacteriostatic activities and metal chelation of kojic acid analogs. Agric. Biol. Chem. 1976, 40, 765–770. [Google Scholar]
- Bashir, F.; Sultana, K.; Khalid, M.; Rabia, H. Kojic Acid: A Comprehensive Review. Asian J. Allied Health Sci. 2021, 6, 13–21. [Google Scholar] [CrossRef]
- Geisen, R.; Schmidt-Heydt, M.; Touhami, N.; Himmelsbach, A. New aspects of ochratoxin A and citrinin biosynthesis in Penicillium. Curr. Opin. Food Sci. 2018, 23, 23–31. [Google Scholar] [CrossRef]
- Timonin, M.; Rouatt, J. Production of citrinin by Aspergillus sp. of the Candidus group. Can. J. Public Health 1944, 35, 80–88. [Google Scholar]
- Hayakawa, S.; Minato, H.; Katagiri, K. The ilicicolins, antibiotics from Cylindrocladium ilicicola. J. Antibiot. 1971, 24, 653–654. [Google Scholar] [CrossRef]
- Jouda, J.-B.; Tamokou, J.-d.-D.; Mbazoa, C.D.; Douala-Meli, C.; Sarkar, P.; Bag, P.K.; Wandji, J. Antibacterial and cytotoxic cytochalasins from the endophytic fungus Phomopsis sp. harbored in Garcinia kola (Heckel) nut. BMC Complement. Altern. Med. 2016, 16, 462. [Google Scholar] [CrossRef]
- Aldridge, D.; Armstrong, J.; Speake, R.; Turner, W. The cytochalasins, a new class of biologically active mould metabolites. Chem. Commun. 1967, 1, 26–27. [Google Scholar] [CrossRef]
- Lynch, F.; Geoghegan, M. Antibiotic activity of a fungal perylene-quinone and some of its derivatives. Trans. Br. Mycol. Soc. 1979, 72, 31–37. [Google Scholar] [CrossRef]
- Singh, K.; Kumari, A. Mycotoxins Co-occurrence Poisoning. In Mycotoxins and Mycotoxicoses; Springer: Singapore, 2022; pp. 129–136. [Google Scholar]
- Coop, I.E. Depression of lambing performance from mating on lucerne. Proc. N. Z. Soc. Anim. Prod. 1977, 37, 149–151. [Google Scholar]
- Mostrom, M.; Evans, T.J. Phytoestrogens. In Veterinary Toxicology: Basic and Clinical Principles, 3rd ed.; Gupta, R.C., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 817–833. [Google Scholar]
- Mostrom, M.; Evans, T.J. Phytoestrogens. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press—Medical: Amsterdam, The Netherlands, 2011; pp. 707–722. [Google Scholar]
- Grgic, D.; Varga, E.; Novak, B.; Müller, A.; Marko, D. Isoflavones in Animals: Metabolism and Effects in Livestock and Occurrence in Feed. Toxins 2021, 13, 836. [Google Scholar] [CrossRef] [PubMed]
- Vejdovszky, K.; Schmidt, V.; Warth, B.; Marko, D. Combinatory estrogenic effects between the isoflavone genistein and the mycotoxins zearalenone and alternariol in vitro. Mol. Nutr. Food Res. 2017, 61, 1600526. [Google Scholar] [CrossRef]
- Hessenberger, S.; Botzi, K.; Degrassi, C.; Kovalsky, P.; Schwab, C.; Schatzmayr, D.; Schatzmayr, G.; Fink-Gremmels, J. Interactions between plant-derived oestrogenic substances and the mycoestrogen zearalenone in a bioassay with MCF-7 cells. Pol. J. Vet. Sci. 2017, 20, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P.K.; Pradhan, S.P.; Kumar, P.S. Isolation, elucidation, and structure–activity relationships of phytoalkaloids from Solanaceae. Stud. Nat. Prod. Chem. 2022, 72, 371–389. [Google Scholar]
- Kohnen-Johannsen, K.L.; Kayser, O. Tropane Alkaloids: Chemistry, pharmacology, biosynthesis and production. Molecules 2019, 24, 796. [Google Scholar] [CrossRef]
- Romera-Torres, A.; Romero-González, R.; Vidal, J.L.M.; Frenich, A.G. Study of the occurrence of tropane alkaloids in animal feed using LC-HRMS. Anal. Methods 2018, 10, 3340–3346. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on Tropane Alkaloids in food and feed. EFSA J. 2013, 11, 1–113. [Google Scholar]
- Ruan, Z.; Wang, X.; Liu, Y.; Liao, W. Corn. In Integrated Processing Technologies for Food and Agricultural By-Products; Pan, Z., Zhang, R., Zicari, S., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 59–72. [Google Scholar]
- Karls, C.; Schaefer, M.; Schaefer, D. PSXI-8 Corn stover feedlot diets with elevated mycotoxins and binder effect on cattle performance. J. Anim. Sci. 2020, 98, 394. [Google Scholar] [CrossRef]
- Reyes-Velázquez, W.P.; Anguiano-Sevilla, C.N.; Anguiano-Estrella, R.; Rojo, F.G. Association of acute equine leukoencephalomalacia (ELEM) with fumonisins concentrations in corn stover in an outbreak in the state of Jalisco, Mexico. Austral J. Vet. Sci. 2018, 50, 111–113. [Google Scholar] [CrossRef]
- Coufal-Majewski, S.; Stanford, K.; McAllister, T.; Blakley, B.; McKinnon, J.; Chaves, A.V.; Wang, Y. Impacts of cereal ergot in food animal production. Front. Vet. Sci. 2016, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Treviño, A.; Dávila-Aviña, J.E.; Sánchez, E.; Heredia, N.; García, S. Occurrence of mycotoxins in alfalfa (Medicago sativa L.), sorghum [Sorghum bicolor (L.) Moench], and grass (Cenchrus ciliaris L.) retailed in the state of Nuevo León, México. Agrociencia 2016, 50, 825–836. [Google Scholar]
- Del Palacio, A.; Mionetto, A.; Bettucci, L.; Pan, D. Evolution of fungal population and mycotoxins in sorghum silage. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 1864–1872. [Google Scholar] [CrossRef]
- da Silva, J.B.; Pozzi, C.R.; Mallozzi, M.A.; Ortega, E.M.; Corrêa, B. Mycoflora and occurrence of aflatoxin B1 and fumonisin B1 during storage of Brazilian sorghum. J. Agric. Food Chem. 2000, 48, 4352–4356. [Google Scholar] [CrossRef]
- McElhinney, C.; Danaher, M.; Grant, J.; Elliott, C.T.; Okiely, P. Variation associated with sampling bale or pit silage for mycotoxins and conventional chemical characteristics. World Mycotoxin J. 2016, 9, 331–342. [Google Scholar] [CrossRef]
- Steiner, D.; Sulyok, M.; Malachová, A.; Mueller, A.; Krska, R. Realizing the simultaneous liquid chromatography-tandem mass spectrometry based quantification of >1200 biotoxins, pesticides and veterinary drugs in complex feed. J. Chormatogr. A. 2020, 1629, 461502. [Google Scholar] [CrossRef]
- Hinkle, D.E.; Wiersma, W.; Jurs, S.G. Correlation. In Applied Statistics for the Behavioral Sciences; Houghton Mifflin College Division: Boston, MA, USA, 2003; pp. 485–528. [Google Scholar]







| Dietary Ingredient | Frequency of Inclusion (n = 19) (%) | Dietary Content (% of DM Basis) | ||
|---|---|---|---|---|
| Average ± SD | Median | Range | ||
| Whole-plant corn silage | 100 | 38.9 ± 6.62 | 40.0 | 27.5–53.0 |
| Alfalfa hay | 79 | 9.1 ± 4.36 | 9.0 | 3.5–16.5 |
| High-energy density concentrate | 74 | 45.5 ± 8.85 | 48.5 | 28.3–60.0 |
| Protein-rich concentrate | 26 | 22.6 ± 3.58 | 25.0 | 17.0–25.0 |
| Corn stover | 21 | 3.5 ± 2.58 | 2.9 | 1.0–7.0 |
| Rolled corn | 21 | 25.5 ± 6.26 | 27.9 | 16.3–30.0 |
| Corn meal | 16 | 24.2 ± 9.80 | 23.0 | 15.0–34.5 |
| Oat hay | 16 | 3.3 ± 0.76 | 3.5 | 2.5–4.0 |
| Sorghum silage | 11 | 13.0 ± 2.83 | 13.0 | 11.0–15.0 |
| Corn bran | 11 | 13.8 ± 0.35 | 13.8 | 13.5–14.0 |
| Alfalfa silage | 5 | – | – | 5.0 |
| Bakery byproduct | 5 | – | – | 7.0 |
| Brewery spent grain | 5 | – | – | 9.5 |
| Forage | 100 | 49.9 ± 4.42 | 50.0 | 40.0–60.0 |
| Concentrate | 100 | 50.1 ± 4.42 | 50.0 | 40.0–60.0 |
| Concentration (µg/kg DM) 1 | Group of Metabolites | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ergot Alkaloids | Alternaria | Aspergillus | Fusarium | Penicillium | Other Fungal Species | Mycotoxins | Total Fungal Metabolites | Unspecific Metabolites | Phytoestrogens | Plant Metabolites | ||
| Whole-plant corn silages (n = 19) | Average | 2.24 | 56.4 | 154 | 8700 | 243 | 164 | 8710 | 9300 | 20,320 | 1740 | 2450 |
| ±SD | 2.09 | 60.4 | 292 | 6410 | 163 | 247 | 6340 | 6620 | 4950 | 3290 | 3820 | |
| Median | 1.5 | 39.1 | 53.8 | 6460 | 193 | 84.8 | 6330 | 7670 | 19,920 | 638 | 726 | |
| Minimum | 0.1 | 9.10 | 2.5 | 39.6 | 28.7 | 12 | 79.5 | 219 | 9860 | 90.5 | 90.5 | |
| Maximum | 5.6 | 242 | 1040 | 22,110 | 520 | 1090 | 21,960 | 22,800 | 27,790 | 12,350 | 13,190 | |
| Total mixed rations (n = 19) | Average | 3.98 | 48.7 | 61.7 | 5550 | 171 | 169 | 5590 | 6000 | 10,240 | 224,260 | 225,960 |
| ±SD | 5.88 | 33.8 | 46.5 | 3160 | 89.1 | 167 | 3000 | 3260 | 3200 | 129,040 | 129,040 | |
| Median | 1.4 | 44.5 | 58.4 | 5840 | 161.1 | 125.4 | 5970 | 6190 | 10,300 | 209,740 | 211,540 | |
| Minimum | 0.4 | 5.6 | 11.7 | 112 | 23.2 | 6.1 | 139 | 161 | 3540 | 34,270 | 35,910 | |
| Maximum | 12.7 | 108 | 179 | 10,510 | 337 | 697 | 10,570 | 11,070 | 15,980 | 448,670 | 449,400 | |
| Wilcoxon matched-pairs test | p-value * | 0.625 | 0.418 | 0.7086 | 0.0005 | 0.0108 | 0.984 | 0.0006 | 0.0004 | <0.0001 | <0.0001 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penagos-Tabares, F.; Sulyok, M.; Artavia, J.-I.; Flores-Quiroz, S.-I.; Garzón-Pérez, C.; Castillo-Lopez, E.; Zavala, L.; Orozco, J.-D.; Faas, J.; Krska, R.; et al. Mixtures of Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Whole-Plant Corn Silages and Total Mixed Rations of Dairy Farms in Central and Northern Mexico. Toxins 2023, 15, 153. https://doi.org/10.3390/toxins15020153
Penagos-Tabares F, Sulyok M, Artavia J-I, Flores-Quiroz S-I, Garzón-Pérez C, Castillo-Lopez E, Zavala L, Orozco J-D, Faas J, Krska R, et al. Mixtures of Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Whole-Plant Corn Silages and Total Mixed Rations of Dairy Farms in Central and Northern Mexico. Toxins. 2023; 15(2):153. https://doi.org/10.3390/toxins15020153
Chicago/Turabian StylePenagos-Tabares, Felipe, Michael Sulyok, Juan-Ignacio Artavia, Samanta-Irais Flores-Quiroz, César Garzón-Pérez, Ezequías Castillo-Lopez, Luis Zavala, Juan-David Orozco, Johannes Faas, Rudolf Krska, and et al. 2023. "Mixtures of Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Whole-Plant Corn Silages and Total Mixed Rations of Dairy Farms in Central and Northern Mexico" Toxins 15, no. 2: 153. https://doi.org/10.3390/toxins15020153
APA StylePenagos-Tabares, F., Sulyok, M., Artavia, J. -I., Flores-Quiroz, S. -I., Garzón-Pérez, C., Castillo-Lopez, E., Zavala, L., Orozco, J. -D., Faas, J., Krska, R., & Zebeli, Q. (2023). Mixtures of Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Whole-Plant Corn Silages and Total Mixed Rations of Dairy Farms in Central and Northern Mexico. Toxins, 15(2), 153. https://doi.org/10.3390/toxins15020153