The Effect of Mushroom Culture Filtrates on the Inhibition of Mycotoxins Produced by Aspergillus flavus and Aspergillus carbonarius

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
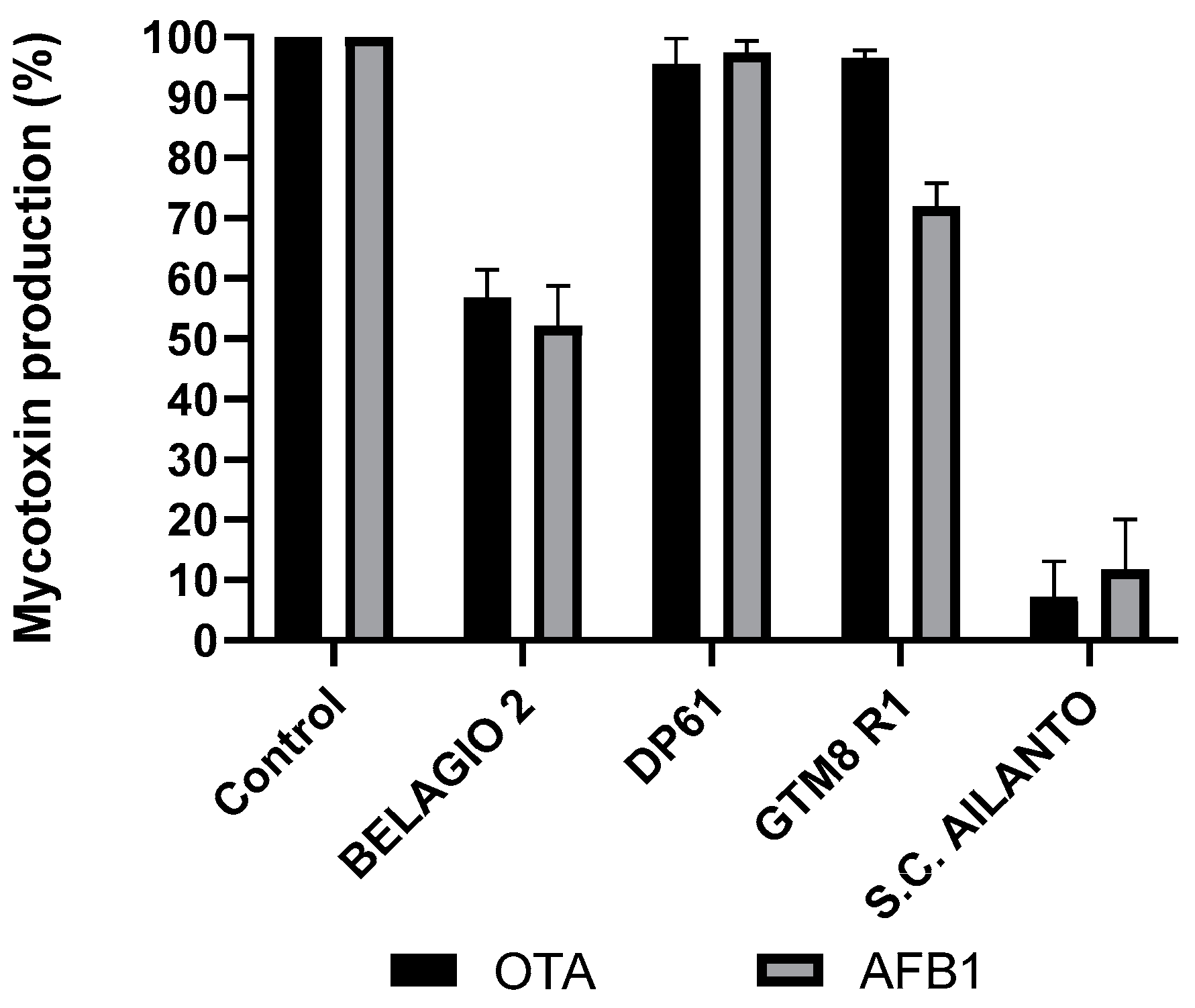
2.1. Wide Screening Mushrooms Assay
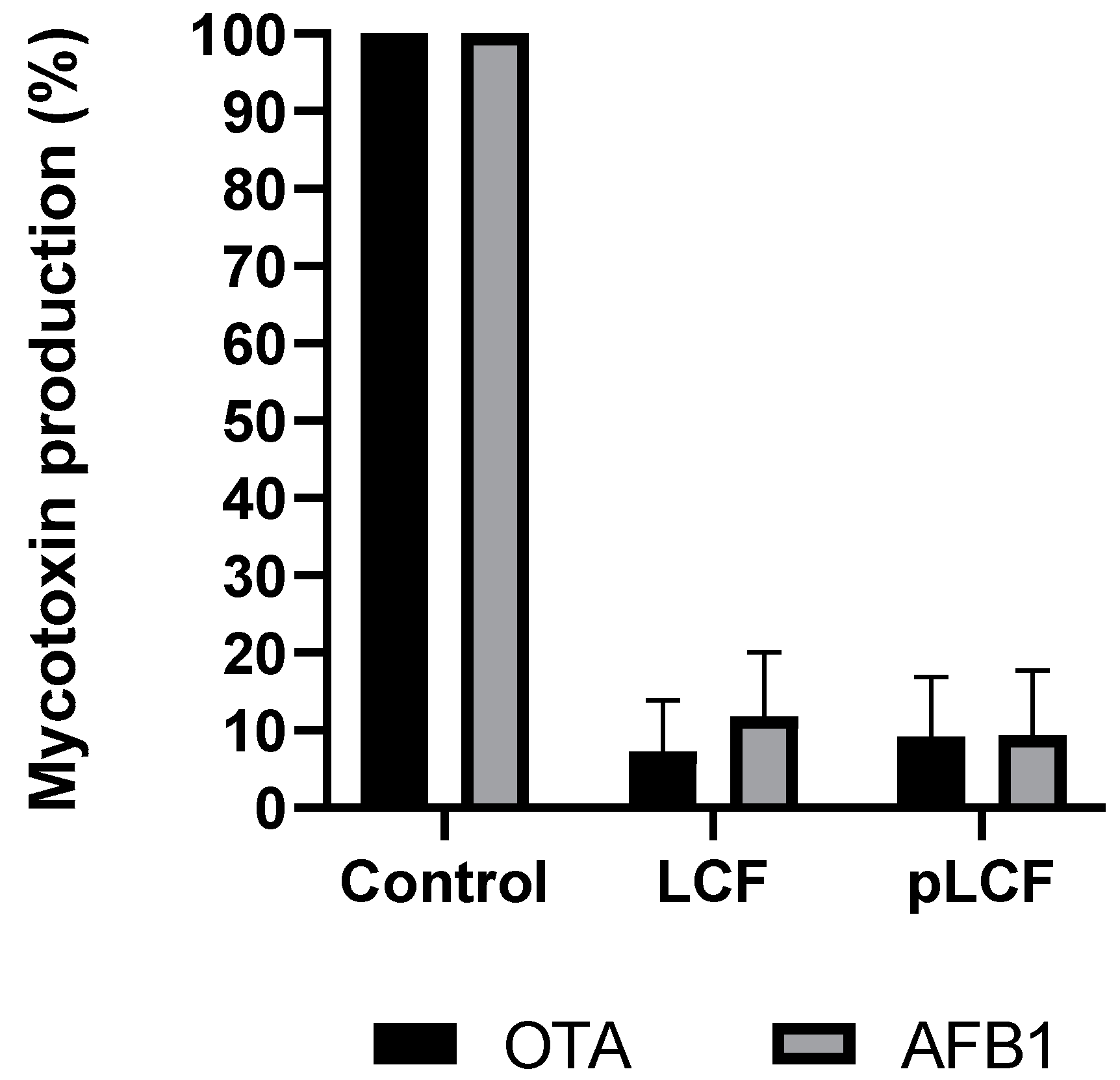
2.2. Influence of the LCF of S. commune on AFB1 and OTA Present in the Media
2.3. Assay of the Influence of LCF and Partially Purified pLCF of S. commune S. C. Alianto
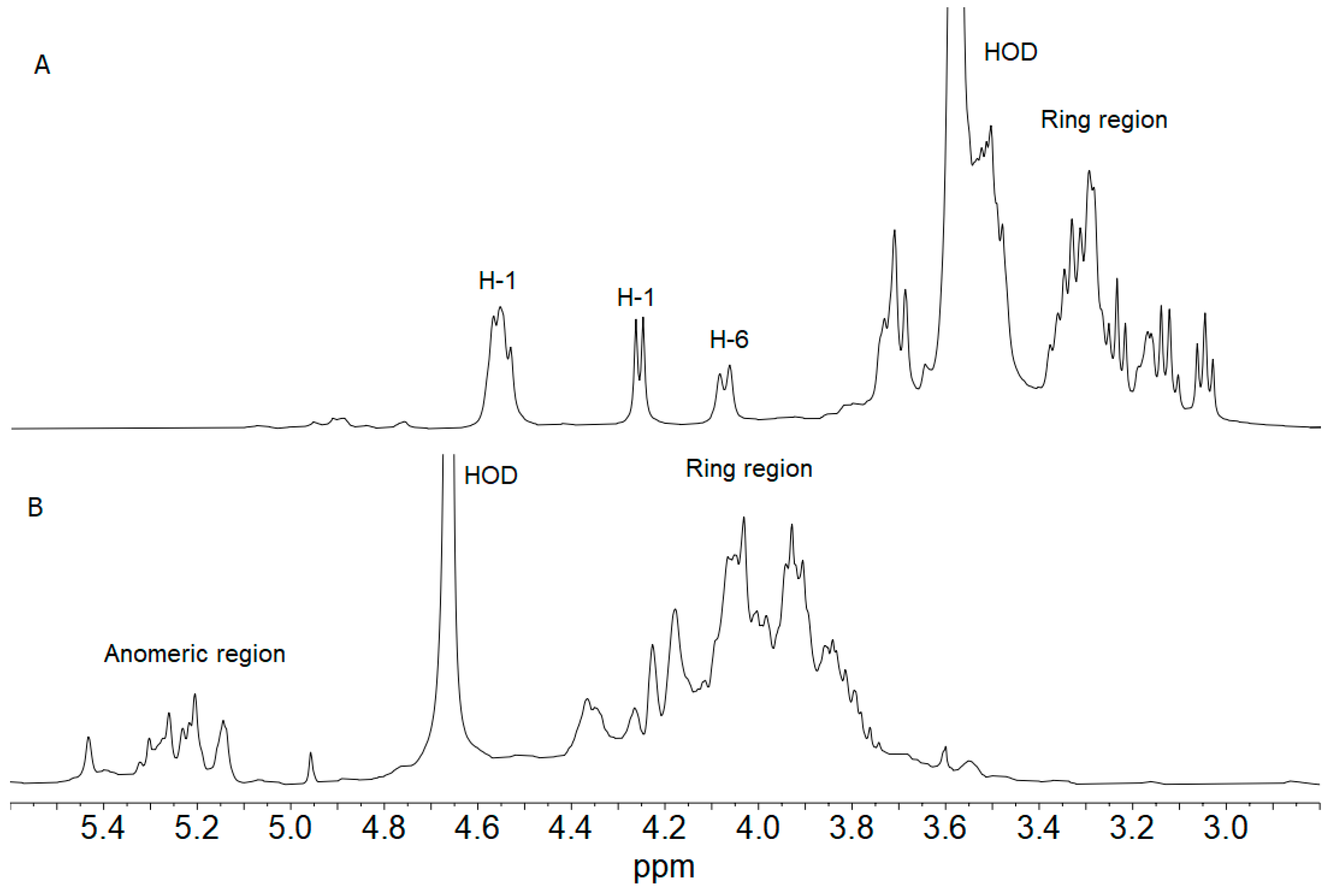
2.4. Composition Analysis of pLCF and sLCF Fractions
2.5. 1H NMR Spectroscopy of pLCF and sLCF Fractions
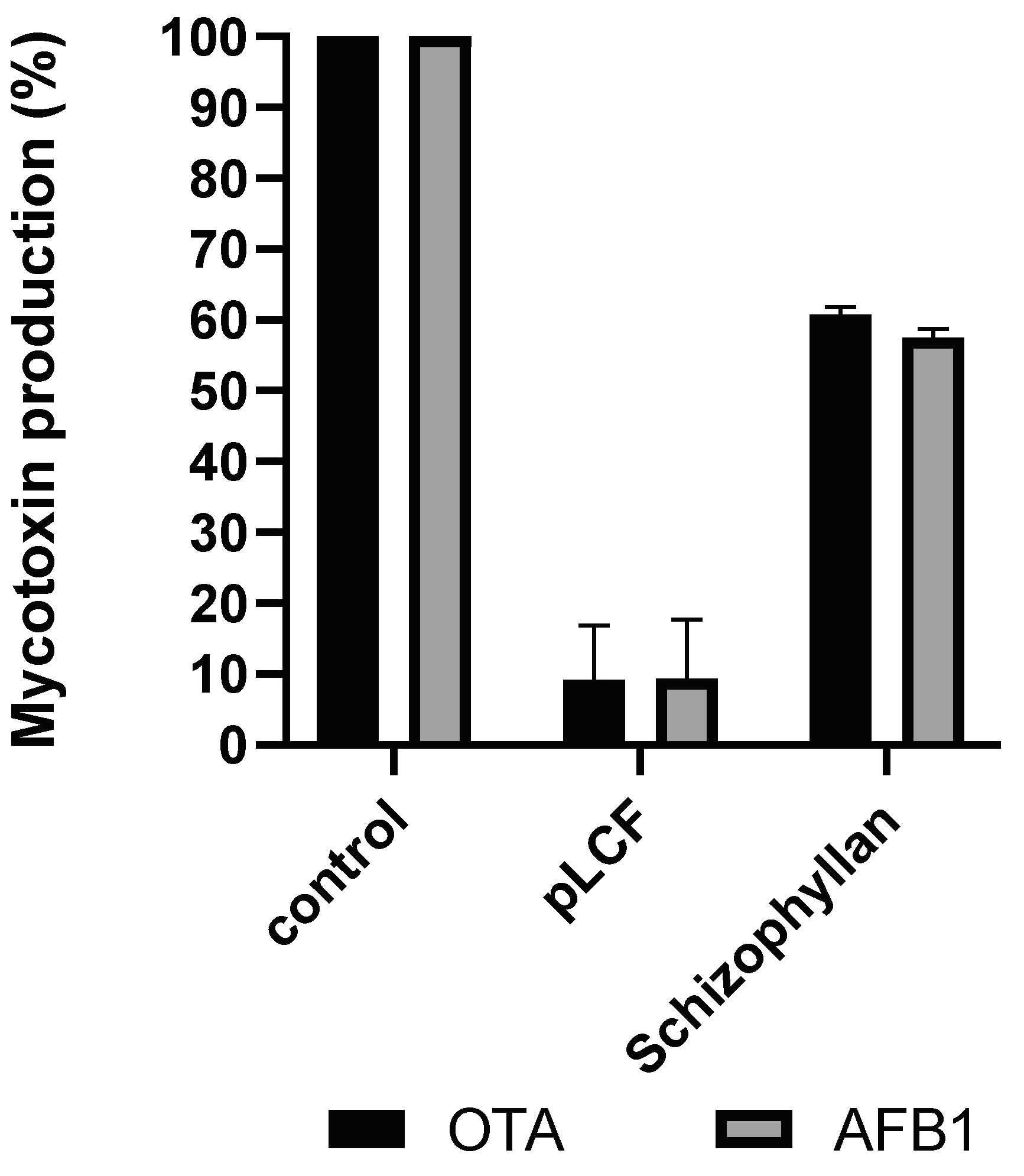
2.6. Assay of the Influence of Partially Purified pLCF of S. commune S. C. Alianto and Commercial Schizophylan (SCH) on AFB1 and OTA Biosynthesis
2.7. Mechanism of Action of pLCF
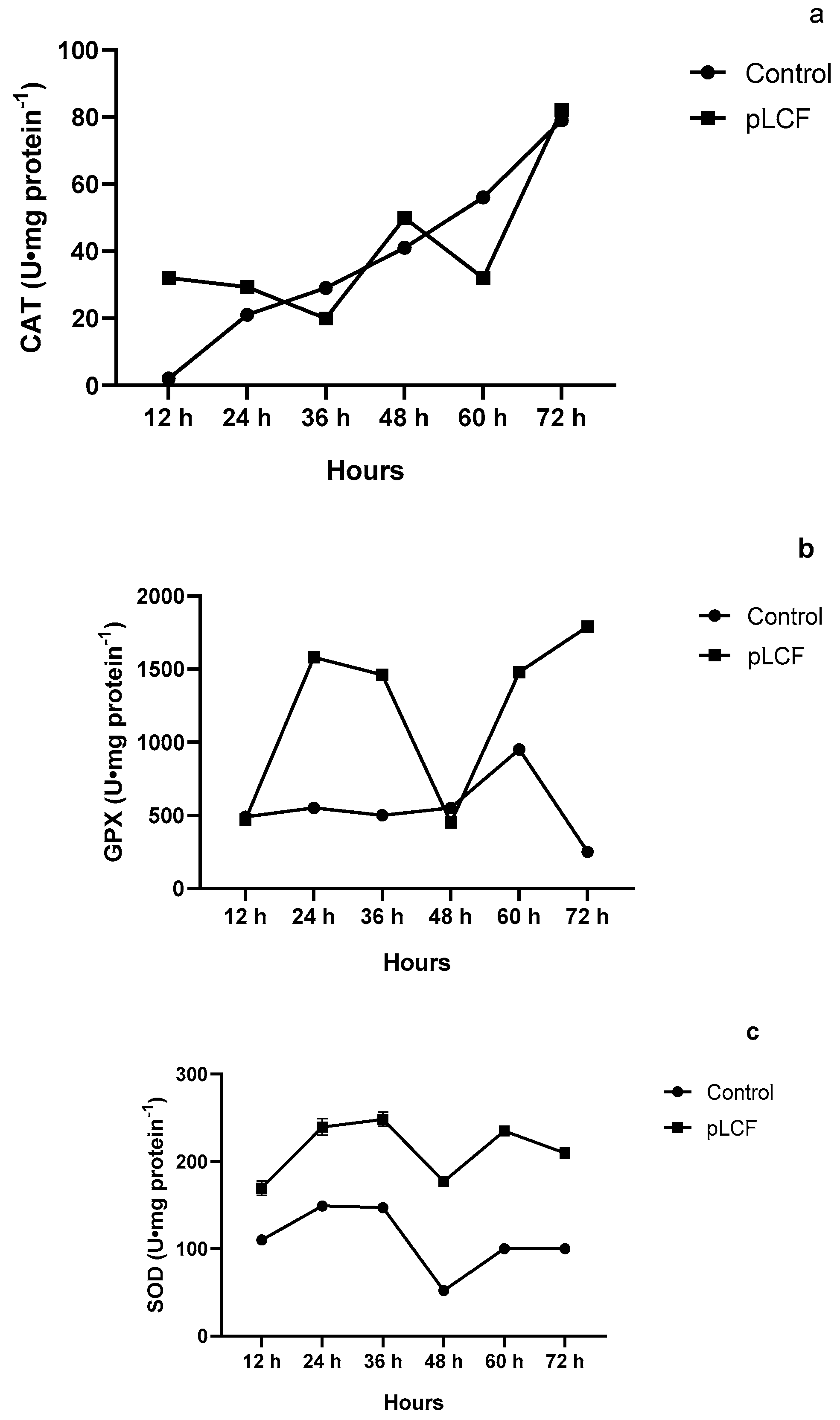
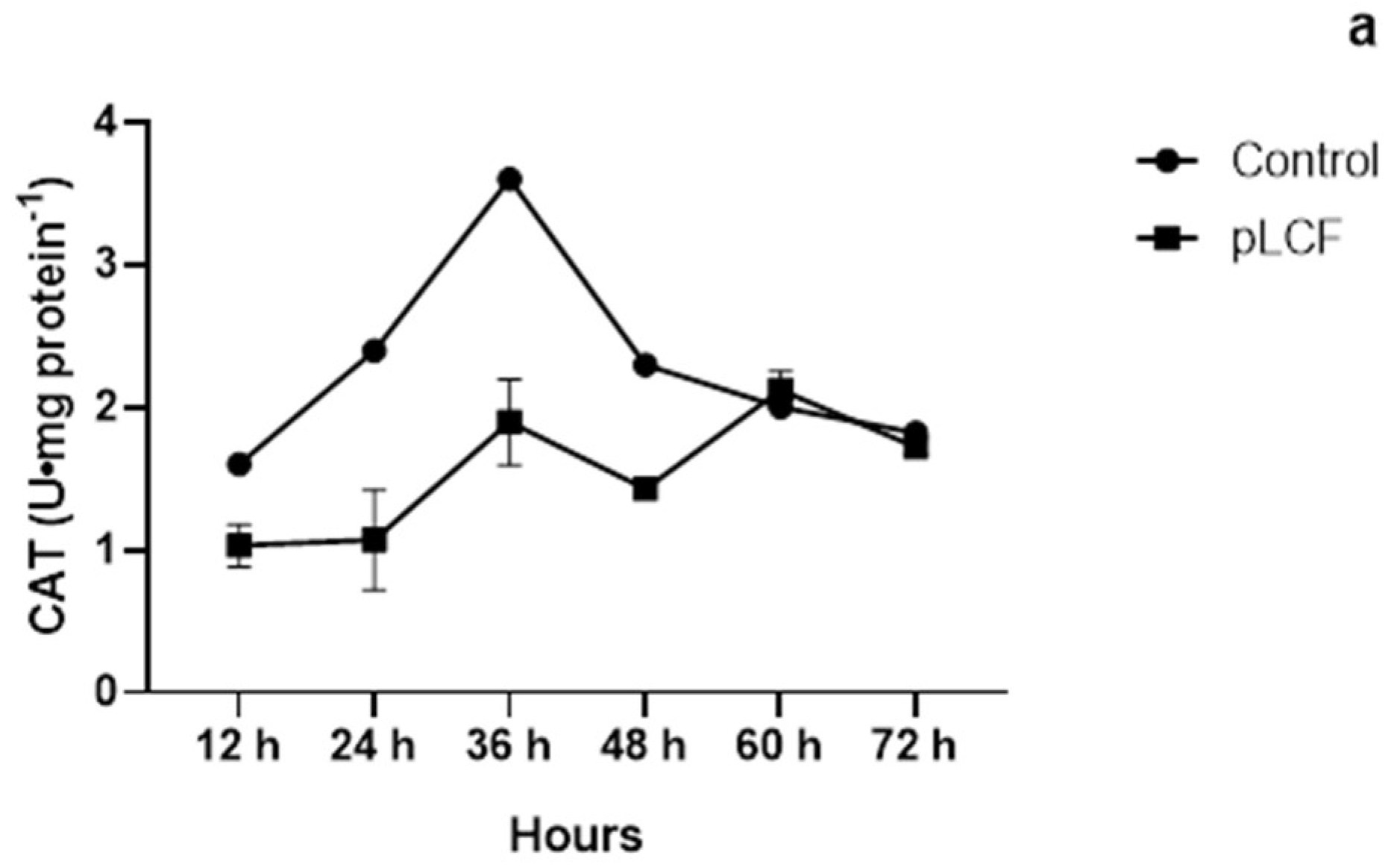
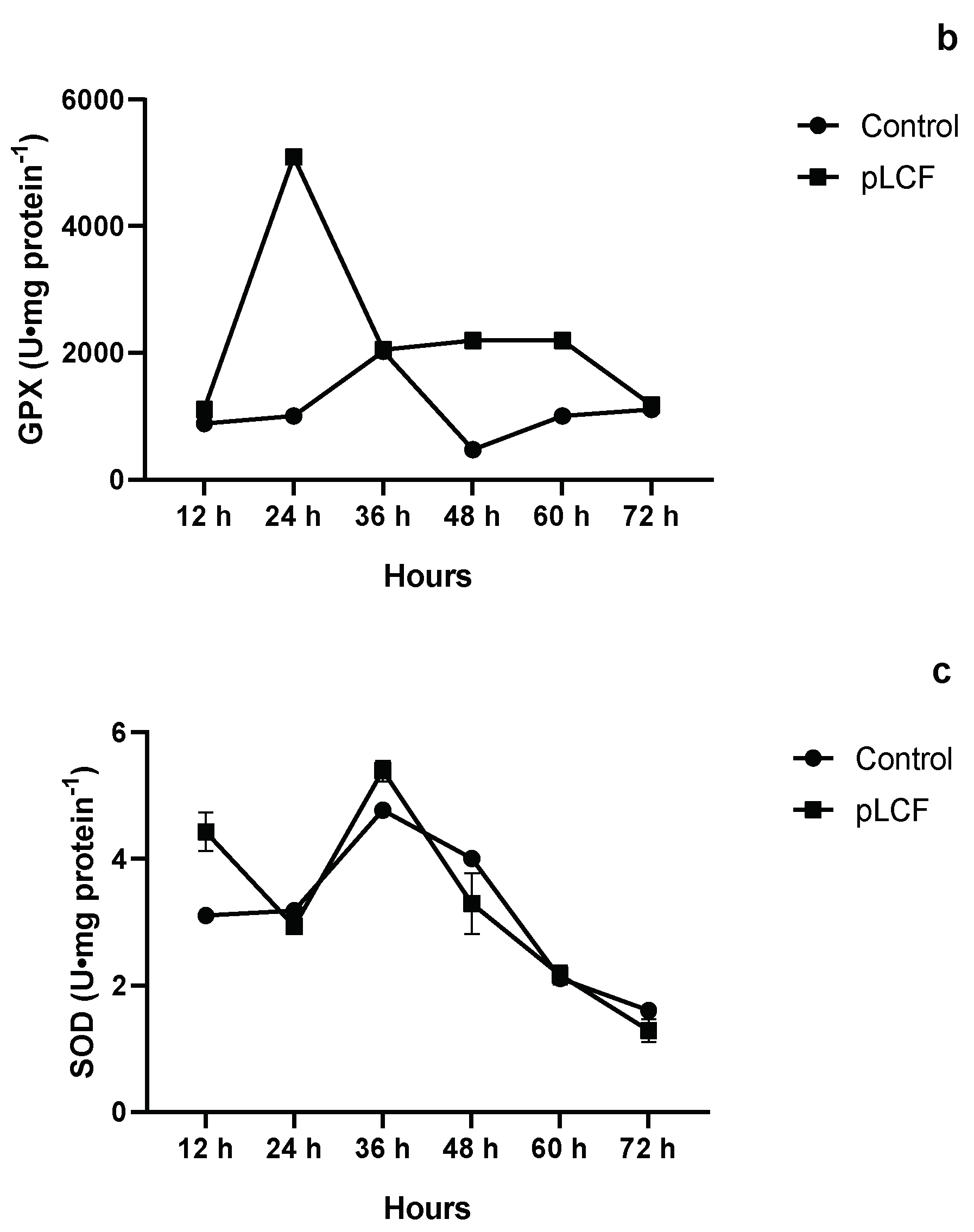
2.7.1. The Activity of Antioxidant Enzymes CAT (a), GPX (b), and SOD (c), during the Time Course in the Mycelium of A. flavus
2.7.2. The Activity of Antioxidant Enzymes CAT (a), GPX (b), and SOD (c), during the Time Course in the Mycelium of A. carbonarius
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strains and Mushrooms Used in this Study
5.2. Production of Mushroom Unrefined Lyophilized Filtrates (LCFs)
5.3. Yield
5.4. In Vitro Screening of Mushroom Polysaccharides on Inhibition of Mycotoxin Synthesis by Aspergillus flavus and Aspergillus carbonarius
5.5. Mycotoxins Extraction and Quantification
5.5.1. Ochratoxin A
5.5.2. Aflatoxin B1
5.6. Influence of LCF on Mycotoxins Already Present in the Culture
5.7. Mycotoxin Inhibition Assay with S. commune Unrefined Extract (LCF)
5.8. Analysis of Monosaccharides in pLCF and sLCF Fractions
5.9. Purification of S. commune Unrefined Polysaccharide (LCF)
5.10. Assays of the Different Variants of S. commune Polysaccharide (pLCF and sLCF) on Mycotoxin Inhibition
5.10.1. Assay of the Semi purified Lyophilized (pLCF) Polysaccharide Fraction from S. commune on Aflatoxin B1 and Ochratoxin A Biosynthesis
5.10.2. Assay of the Fraction of the Polysaccharide from S. commune (sLCF) on Ochratoxin A and Aflatoxin B1 Biosynthesis
5.11. Assay of the Comparison between Commercial Schizophyllan and Semi-Purified Extract (pLCF) of S. Commune on Inhibition of Mycotoxin Synthesis
5.12. Ultraviolet Visible Spectrophotometry
5.13. 1H NMR Spectroscopy
5.14. Analysis of Antioxidant Enzyme Activities in A. flavus and A. carbonarius Treated and Nontreated with Lyophilized Semipurified Filtrates (pLCF) of S. commune
5.15. Quantification of the Proteins
5.16. Statistics
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciegler, A.; Bennett, J.W. Mycotoxins and Mycotoxicoses. BioScience 1980, 30, 512–515. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.; Patial, V. Food mycotoxins: Dietary interventions implicated in the prevention of mycotoxicosis. ACS Food Sci. Technol. 2021, 1, 1717–1739. [Google Scholar] [CrossRef]
- Pereira, C.S.; Cunha, S.C.; Fernandes, J.O. Prevalent mycotoxins in animal feed: Occurrence and analytical methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef] [PubMed]
- Speijers, G.J.A.; Speijers, M.H.M. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- IARC. Monographs on the Evaluation of Carcinogenic Risks to Humans: Chemical Agents and Related Occupations. A Review of Human Carcinogens; IARC: Lyon, France, 2012; pp. 224–248. [Google Scholar]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; et al. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, A.; ul Abidin, Z. Mycotoxicosis–diagnosis, prevention and control: Past practices and future perspectives. Toxin Rev. 2018, 39, 1–16. [Google Scholar] [CrossRef]
- Navale, V.; Vamkudoth, K.R.; Ajmera, S.; Dhuri, V. Aspergillus derived mycotoxins in food and the environment: Prevalence, detection, and toxicity. Toxicol. Rep. 2021, 8, 1008–1030. [Google Scholar] [CrossRef] [PubMed]
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Asia Pac. J. Clin. Nutr. 2007, 16, 95–101. [Google Scholar] [CrossRef] [PubMed]
- European Union. COMMISSION REGULATION (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, L364, 5–24. [Google Scholar]
- Van Egmond, H.P.; Schothorst, R.C.; Jonker, M.A. Regulations relating to mycotoxins in food. Anal. Bioanal. Chem. Res. 2007, 389, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, E.M. Managing the risk of mycotoxins in modern feed production. J. Anim. Feed Sci. Technol. 2007, 133, 149–166. [Google Scholar] [CrossRef]
- Loncar, J.; Bellich, B.; Parroni, A.; Reverberi, M.; Rizzo, R.; Zjalić, S.; Cescutti, P. Oligosaccharides derived from Tramesan: Their structure and activity on mycotoxin inhibition in Aspergillus flavus and Aspergillus carbonarius. Biomolecules 2021, 11, 243. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.; Quintieri, L.; Cimmarusti, M.; Monaci, L.; Haidukowski, M.; Logrieco, A.; Sanjust, E.; et al. Aflatoxin B1 and M1 Degradation by Lac2 from Pleurotus pulmonarius and Redox Mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of Oral Exposure to Aflatoxin B1-Induced Renal Dysfunction, Oxidative Stress, and Cell Apoptosis in Mice Kidney by Curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ahmed, M.F.E.; Sangare, L.; Zhao, Y.; Selvaraj, J.N.; Xing, F.; Wang, Y.; Yang, H.; Liu, Y. Novel aflatoxin-degrading enzyme from Bacillus shackletonii L7. Toxins 2017, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.S. Aflatoxin detoxification by manganese peroxidase purified from Pleurotus ostreatus. Braz. J. Microbiol. 2014, 45, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Li, R.; Ng, T.B.; Lai, Y.; Yang, J.; Ye, X. A new laccase of Lac 2 from the white rot fungus Cerrena unicolor 6884 and Lac 2-mediated degradation of aflatoxin B1. Toxins 2020, 12, 476. [Google Scholar] [CrossRef] [PubMed]
- Zaccaria, M.; Dawson, W.; Russel Kish, D.; Reverberi, M.; Bonaccorsi di Patti, M.C.; Domin, M.; Cristiglio, V.; Chan, B.; Dellafiora, L.; Gabel, F.; et al. Experimental–theoretical study of laccase as a detoxifier of aflatoxins. Sci. Rep. 2023, 13, 860. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Fabbri, A.A.; Zjalic, S.; Ricelli, A.; Punelli, F.; Fanelli, C. Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl. Microbiol. Biotechnol. 2005, 69, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Ricelli, A.; Zjalic, S.; Fabbri, A.A.; Fanelli, C. Natural functions of mycotoxins and control of their biosynthesis in fungi. Appl. Microbiol. Biotechnol. 2010, 87, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Zjalic, S.; Reverberi, M.; Ricelli, A.; Granito, M.V.; Fanelli, C.; Fabbri, A.A. Trametes versicolor: A possible tool for aflatoxin control. Int. J. Food Microbiol. 2006, 107, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Scarpari, M.; Parroni, A.; Zaccaria, M.; Fattorini, L.; Bello, C.; Fabbri, A.A.; Bianchi, G.; Scala, V.; Zjalic, S.; Fanelli, C. Aflatoxin control in maize by Trametes versicolor. Toxins 2014, 6, 3426–3437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricelli, A.; Fabbri, A.A.; Trionfetti-Nisini, P.; Reverberi, M.; Zjalic, S.; Fanelli, C. Inhibiting effect of different edible and medicinal mushrooms on the growth of two ochratoxigenic microfungi. Int. J. Med. Mushrooms 2002, 4, 8. [Google Scholar] [CrossRef]
- Tada, R.; Harada, T.; Nagi-Miura, N.; Adachi, Y.; Nakajima, M.; Yadomae, T.; Ohno, N. NMR characterization of the structure of a β-(1→3)-d-glucan isolate from cultured fruit bodies of Sparassis crispa. Carbohydr. Res. 2007, 342, 2611–2618. [Google Scholar] [CrossRef] [PubMed]
- Rau, U. Glucans secreted by fungi. Turk. Electron. J. Biotechnol. 2004, 2, 30–36. [Google Scholar]
- Zhang, G.Q.; Chen, Q.J.; Sun, J.; Wang, H.X.; Han, C.H. Purification and characterization of a novel acid phosphatase from the split gill mushroom Schizophyllum commune. J. Basic Microbiol. 2013, 53, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Slameňová, D.; Lábaj, J.; Križková, L.; Kogan, G.; Šandula, J.; Bresgen, N.; Eckl, P. Protective effects of fungal (1→3)-β-D-glucan derivatives against oxidative DNA lesions in V79 hamster lung cells. Cancer Lett. 2003, 198, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Punelli, F.; Camera, E.; Fabbri, C.; Picardo, M.; Fanelli, C.; Fabbri, A.A. Modulation of antioxidant defense in Aspergillus parasiticus is involved in aflatoxin biosynthesis: A role for the Ap yapA gene. Eukaryot. Cell 2008, 7, 988–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Mohawed, S.M.; Bhatnagar, D.; Cleveland, T.E. Substrate induced lipase gene expression and aflatoxin production in Aspergillus parasiticus and Aspergillus flavus. J. Appl. Microbiol. 2003, 95, 1334–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barberis, C.; Astoreca, A.; Fernandez-Juri, G.; Chulze, S.; Dalcero, A.; Magnoli, C. Use of propyl paraben to control growth and ochratoxin A production by Aspergillus section Nigri species on peanut meal extract agar. Int. J. Food Microbiol. 2009, 136, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Franz, G. Polysaccharides in pharmacy: Current applications and future concepts. Planta Med. 1989, 55, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T.; Tabata, K.; Itoh, W.; Yanaki, T. Molecular weight dependence of the antitumor activity of schizophyllan. J. Agric. Food Chem. 1986, 50, 231–232. [Google Scholar] [CrossRef]
- Kikumoto, S. Polysaccharide produced by Schizophyllum commune, I. Formation and some properties of an extracellular polysaccharide. Nippon. Nogeikagaku Kaishi 1970, 44, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Kidd, P.M. The use of mushroom glucans and proteoglycans in cancer treatment. Altern. Med. Rev. 2000, 5, 4–27. [Google Scholar] [PubMed]
- Reverberi, M.; Zjalic, S.; Punelli, F.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Apyap1 affects aflatoxin biosynthesis during Aspergillus parasiticus growth in maize seeds. Food Addit. Contam. 2007, 24, 1070–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahoney, N.; Molyneux, R.J. Phytochemical inhibition of aflatoxigenicity in Aspergillus flavus by constituents of walnut (Juglans regia). J. Agric. Food Chem. 2004, 52, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Heredia, N.; García, S. Inhibition of growth and mycotoxin production of Aspergillus flavus and Aspergillus parasiticus by extracts of Agave species. Int. J. Food Microbiol. 2005, 98, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Abuajah, C.I.; Ogbonna, A.C.; Osuji, C.M. Functional components and medicinal properties of food: A review. J. Food Sci. Technol. 2015, 52, 2522–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parroni, A.; Bellabarba, A.; Beccaccioli, M.; Scarpari, M.; Reverberi, M.; Infantino, A. Use of the secreted proteome of Trametes versicolor for controlling the cereal pathogen Fusarium langsethiae. Int. J. Mol. Sci. 2019, 20, 4167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakehi, K.; Honda, S. Silyl ethers of carbohydrates. In Analysis of Carbohydrates by GLC and MS, 1st ed.; Biermann, C.J., McGinnis, G.D., Eds.; CRC Press: Boca Raton, FL, USA, 2021; Volume 1, pp. 43–85. [Google Scholar]
- Fanelli, C.; Tasca, V.; Ricelli, A.; Reverberi, M.; Zjalic, S.; Finotti, E.; Fabbri, A.A. Inhibiting effect of medicinal mushroom Lentinus edodes (Berk.) Sing. (Agaricomycetideae) on aflatoxin production by Aspergillus parasiticus Speare. Int. J. Med. Mushrooms 2000, 2, 4. [Google Scholar] [CrossRef]
- Amezqueta, S.; Schorr-Galindo, S.; Murillo-Arbizu, M.; Gonzalez-Peñas, E.; De Cerain, A.L.; Guiraud, J.P. OTA-producing fungi in foodstuffs: A review. Food Control 2012, 26, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Kruger, N.J. The Bradford Method for protein quantitation. In The Protein Protocols Handbook; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2019. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Genera | Species | Inhibition of OTA(%) ± SD | T-Value (p) | Inhibition of AF (%) ± SD | T-Value (p) | Yield g • L−1 |
|---|---|---|---|---|---|---|---|
| CF 16 | Agrocybe | aegerita | 6.64 ± 1.55 | 134.41 (<0.001) | 70.18 ± 7.61 | 8.76 (0.001) | 7.21 ± 0.50 |
| CF 249 | Agrocybe | aegerita | 31.48 ± 3.75 | 40.82 (<0.001) | 61.73 ± 7.38 | 11.60 (<0.001) | 6.82 ± 0.52 |
| CF 250 | Agrocybe | aegerita | 23.17 ± 3.33 | 51.66 (<0.001) | 58.48 ± 8.24 | 11.27 (<0.001) | 7.22 ± 0.48 |
| CF 278 | Armillaria | mellea | 4.35 ± 2.60 | 82.24 (<0.001) | 25.60 ± 2.58 | 64.56 (<0.001) | 8.12 ± 0.46 |
| CF 253 | Fomes | fomentarius | 37.06 ± 5.55 | 25.34 (<0.001) | 19.91 ± 3.04 | 58.88 (<0.001) | 9.32 ± 0.53 |
| CF 255 | Fomes | fomentarius | 35.68 ± 4.61 | 31.17 (<0.001) | 57.00 ± 7.92 | 12.14 (<0.001) | 9.29 ± 0.50 |
| CF 262 | Fomes | fomentarius | 7.24 ± 5.45 | 38.09 (<0.001) | 45.70 ± 5.21 | 23.33 (<0.001) | 9.45 ± 0.49 |
| P.COLLEGNO 2A | Ganoderma | adspersum | 25.30 ± 3.33 | 50.21 (<0.001) | 3.45 ± 0.95 | 227.61 (<0.001) | 9.11 ± 0.52 |
| COLETTA 12 | Ganoderma | adspersum | 4.97 ± 3.69 | 57.62 (<0.001) | 5.78 ± 3.57 | 59.06 (<0.001) | 9.51 ± 0.60 |
| ROBUR (69.2) | Ganoderma | adspersum | 8.20 ± 6.20 | 33.11 (<0.001) | 69.94 ± 7.59 | 8.86 (0.001) | 9.59 ± 049 |
| Meisino 9 | Ganoderma | adspersum | 8.00 ± 4.95 | 41.57 (<0.001) | 53.01 ± 5.41 | 19.42 (<0.001) | 7.62 ± 0.51 |
| Tiglio | Ganoderma | adspersum | 25.36 ± 3.12 | 53.56 (<0.001) | 26.28 ± 3.48 | 47.33 (<0.001) | 9.54 ± 0.54 |
| Cep 10 | Ganoderma | adspersum | 4.14 ± 1.61 | 133.24 (<0.001) | 68.32 ± 9.20 | 7.70 (0.002) | 8.18 ± 0.57 |
| Diano 2 | Ganoderma | adspersum | 27.10 ± 3.54 | 46.11 (<0.001) | 54.20 ± 7.29 | 14.05 (<0.001) | 9.89 ± 0.54 |
| CF 17 | Ganoderma | lucidum | 4.60 ± 2.69 | 79.20 (<0.001) | 5.11 ± 4.85 | 43.79 (<0.001) | 9.56± 0.57 |
| CF 223 | Ganoderma | lucidum | 7.31 ± 4.37 | 47.48 (<0.001) | 20.02 ± 2.21 | 80.94 (<0.001) | 9.12 ± 0.49 |
| CF 264 | Ganoderma | lucidum | 28.82 ± 2.93 | 54.24 (<0.001) | 54.70 ± 5.40 | 18.75 (<0.001) | 9.53 ± 0.51 |
| DP 1 | Ganoderma | resinaceum | 20.23 ± 2.64 | 67.55 (<0.001) | 21.58 ± 2.94 | 59.58 (<0.001) | 9.68 ± 0.52 |
| PLATANO ROMA | Ganoderma | resinaceum | 33.80 ± 3.76 | 39.33 (<0.001) | 5.18 ± 4.79 | 44.23 (<0.001) | 9.72 ± 0.48 |
| P.VERDI 920 | Ganoderma | resinaceum | 41.27 ± 5.80 | 22.65 (<0.001) | 25.28 ± 3.54 | 47.20 (<0.001) | 7.88 ± 0.43 |
| 780.2 | Ganoderma | resinaceum | 42.06 ± 5.77 | 22.45 (<0.001) | 25.10 ± 2.79 | 60.08 (<0.001) | 9.82 ± 0.51 |
| DP 63 | Ganoderma | resinaceum | 43.34 ± 5.14 | 24.67 (<0.001) | 36.70 ± 5.28 | 26.81 (<0.001) | 9.79 ± 0.46 |
| ASPROMONTE 52 | Ganoderma | resinaceum | 53.36 ± 8.07 | 12.93 (<0.001) | 31.29 ± 3.18 | 48.38 (<0.001) | 8.89 ± 0.41 |
| DP 23 | Ganoderma | resinaceum | 6.32 ± 4.92 | 42.62 (<0.001) | 4.87 ± 5.80 | 36.65 (<0.001) | 9.68 ± 0.50 |
| C.TRENTO 43 B | Ganoderma | resinaceum | 30.53 ± 4.21 | 36.86 (<0.001) | 2.45 ± 1.10 | 198.08 (<0.001) | 10.14 ± 0.47 |
| P.VERDI 4A | Ganoderma | resinaceum | 5.18 ± 3.74 | 56.64 (<0.001) | 3.60 ± 1.56 | 138.07 (<0.001) | 9.99 ± 0.52 |
| G.R. 853 | Ganoderma | resinaceum | 6.43 ± 3.24 | 64.65 (<0.001) | 28.20 ± 3.00 | 53.53 (<0.001) | 9.53 ± 0.49 |
| CF 258 | Ganoderma | resinaceum | 4.17 ± 1.09 | 196.13 (<0.001) | 42.71 ± 6.38 | 20.09 (<0.001) | 7.78 ± 0.51 |
| CF 6 | Grifola | frondosa | 10.09 ± 3.28 | 61.24 (<0.001) | 6.25 ± 4.14 | 50.67 (<0.001) | 10.25 ± 0.41 |
| CF 19 | Grifola | frondosa | 7.47 ± 3.48 | 59.44 (<0.001) | 24.95 ± 3.13 | 53.68 (<0.001) | 9.94 ± 0.51 |
| CF 225 | Heterobasidion | annosum | 5.84 ± 3.09 | 68.06 (<0.001) | 23.00 ± 3.00 | 57.33 (<0.001) | 7.34 ± 0.49 |
| CF 224 | Phellinus | pfefferi | 5.76 ± 4.28 | 49.27 (<0.001) | 5.91 ± 4.53 | 46.40 (<0.001) | 6.85 ± 0.59 |
| CF 25 | Pleurotus | eryngii | 26.56 ± 3.23 | 50.85 (<0.001) | 6.27 ± 4.16 | 50.40 (<0.001) | 6.83 ± 0.50 |
| CF 38 | Pleurotus | eryngii | 69.92 ± 9.66 | 6.96 (0.002) | 23.38 ± 3.21 | 53.33 (<0.001) | 7.48 ± 0.55 |
| CF 28 | Polyporus | sulphureus | 24.92 ± 2.50 | 67.26 (<0.001) | 2.53 ± 2.18 | 100.12 (<0.001) | 9.58 ± 0.72 |
| CF 280 | Trametes | hirsuta | 7.22 ± 5.65 | 36.69 (<0.001) | 20.16 ± 2.08 | 86.00 (<0.001) | 7.92 ± 0.54 |
| T.V. 117 | Trametes | versicolor | 85.75 ± 11.76 | 2.71 (0.054) | 89.00 ± 9.00 | 2.73 (0.052) | 6.68 ± 0.50 |
| BELAGIO 2 | Schizophyllum | commune | 43.20 ± 5.23 | 24.31 (<0.001) | 47.91 ± 6.72 | 17.34 (<0.001) | 9.81 ± 0.48 |
| DP61 | Schizophyllum | commune | 4.56 ± 4.86 | 43.87 (<0.001) | 2.59 ± 1.96 | 111.31 (<0.001) | 7.92 ± 0.51 |
| GTM8 R1 | Schizophyllum | commune | 3.46 ± 1.53 | 141.44 (<0.001) | 28.12 ± 3.95 | 40.68 (<0.001) | 8.76 ± 0.56 |
| CF 18 | Stropharia | eugosoannulata | 2.76 ± 0.44 | 490.75 (<0.001) | 35.34 ± 4.90 | 29.51 (<0.001) | 9.92 ± 0.49 |
| S.C. AILANTO | Schizophyllum | commune | 92.76 ± 6.61 | 2.45 (0.070) | 88.20 ± 8.21 | 3.22 (0.032) | 8.59 ± 0.50 |
| AFB1 | OTA | |||
|---|---|---|---|---|
| Time (Days) | 0 | 3 | 0 | 3 |
| CONT | 100 | 97.03 ± 2.87 | 100 | 96.14 ± 3.75 |
| LCF | 100 | 95.94 ± 3.85 | 100 | 96.48 ± 3.01 |
| Code | Genus | Species |
|---|---|---|
| CF 16 | Agrocybe | Aegerita |
| CF 249 | Agrocybe | Aegerita |
| CF 250 | Agrocybe | Aegerita |
| CF 278 | Armillaria | Mellea |
| CF 253 | Fomes | fomentarius |
| CF 255 | Fomes | fomentarius |
| CF 262 | Fomes | fomentarius |
| P.COLLEGNO 2A | Ganoderma | adspersum |
| COLLETTA 12 | Ganoderma | adspersum |
| ROBUR (69.2) | Ganoderma | adspersum |
| Meisino 9 | Ganoderma | adspersum |
| Tiglio | Ganoderma | adspersum |
| Cep.10 | Ganoderma | adspersum |
| Diano 2 | Ganoderma | adspersum |
| CF 17 | Ganoderma | lucidum |
| CF 223 | Ganoderma | lucidum |
| CF 264 | Ganoderma | lucidum |
| DP1 | Ganoderma | resinaceum |
| PLATANO ROMA | Ganoderma | resinaceum |
| P.VERDI 920 | Ganoderma | resinaceum |
| 780.2 | Ganoderma | resinaceum |
| DP63 | Ganoderma | resinaceum |
| ASPROMONTE 52 | Ganoderma | resinaceum |
| DP23 | Ganoderma | resinaceum |
| C.TRENTO 34 B | Ganoderma | resinaceum |
| P. VERDI 4A | Ganoderma | resinaceum |
| G.R. 853 | Ganoderma | resinaceum |
| CF 258 | Ganoderma | resinaceum |
| CF 6 | Grifola | frondosa |
| CF 19 | Grifola | frondosa |
| CF 225 | Heterobasidion | annosum |
| CF 224 | Phellinus | pfefferi |
| CF 25 | Pleurotus | eryngii |
| CF 38 | Pleurotus | eryngii |
| CF 28 | Polyporus | sulphureus |
| BELLAGIO 2 | Schizophyllum | commune |
| DP61 | Schizophyllum | commune |
| GTM8 R1 | Schizophyllum | commune |
| S.C. AILANTO | Schizophyllum | commune |
| CF 18 | Stropharia | rugosoannulata |
| CF 280 | Trametes | hirsuta |
| T.V. 117 | Trametes | versicolor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loncar, J.; Bellich, B.; Cescutti, P.; Motola, A.; Beccaccioli, M.; Zjalic, S.; Reverberi, M. The Effect of Mushroom Culture Filtrates on the Inhibition of Mycotoxins Produced by Aspergillus flavus and Aspergillus carbonarius. Toxins 2023, 15, 177. https://doi.org/10.3390/toxins15030177
Loncar J, Bellich B, Cescutti P, Motola A, Beccaccioli M, Zjalic S, Reverberi M. The Effect of Mushroom Culture Filtrates on the Inhibition of Mycotoxins Produced by Aspergillus flavus and Aspergillus carbonarius. Toxins. 2023; 15(3):177. https://doi.org/10.3390/toxins15030177
Chicago/Turabian StyleLoncar, Jelena, Barbara Bellich, Paola Cescutti, Alice Motola, Marzia Beccaccioli, Slaven Zjalic, and Massimo Reverberi. 2023. "The Effect of Mushroom Culture Filtrates on the Inhibition of Mycotoxins Produced by Aspergillus flavus and Aspergillus carbonarius" Toxins 15, no. 3: 177. https://doi.org/10.3390/toxins15030177
APA StyleLoncar, J., Bellich, B., Cescutti, P., Motola, A., Beccaccioli, M., Zjalic, S., & Reverberi, M. (2023). The Effect of Mushroom Culture Filtrates on the Inhibition of Mycotoxins Produced by Aspergillus flavus and Aspergillus carbonarius. Toxins, 15(3), 177. https://doi.org/10.3390/toxins15030177