Zearalenone Promotes Uterine Hypertrophy through AMPK/mTOR Mediated Autophagy

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
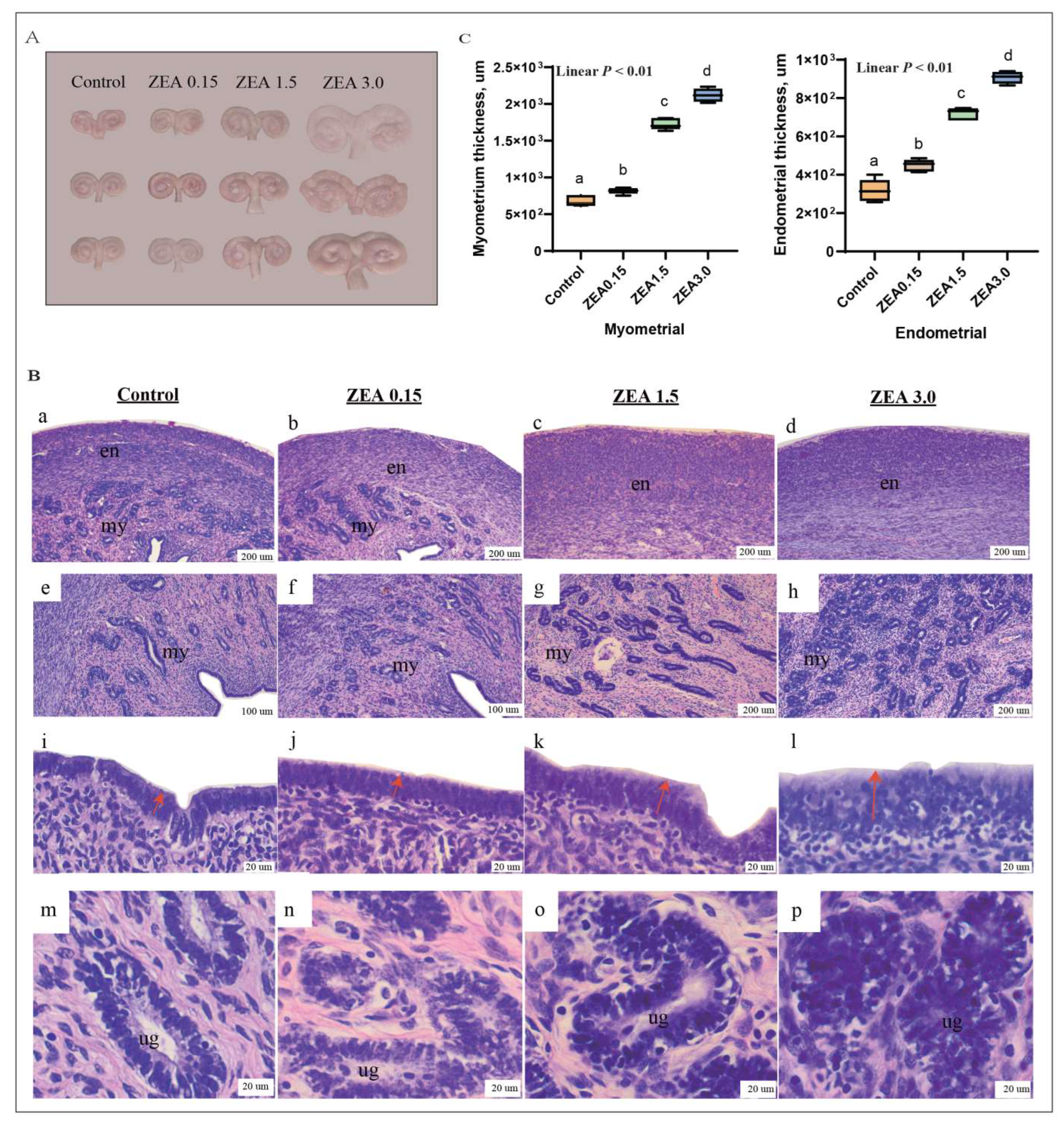
2.1. ZEN-Induced Uterine Hypertrophy in Weaned Piglets
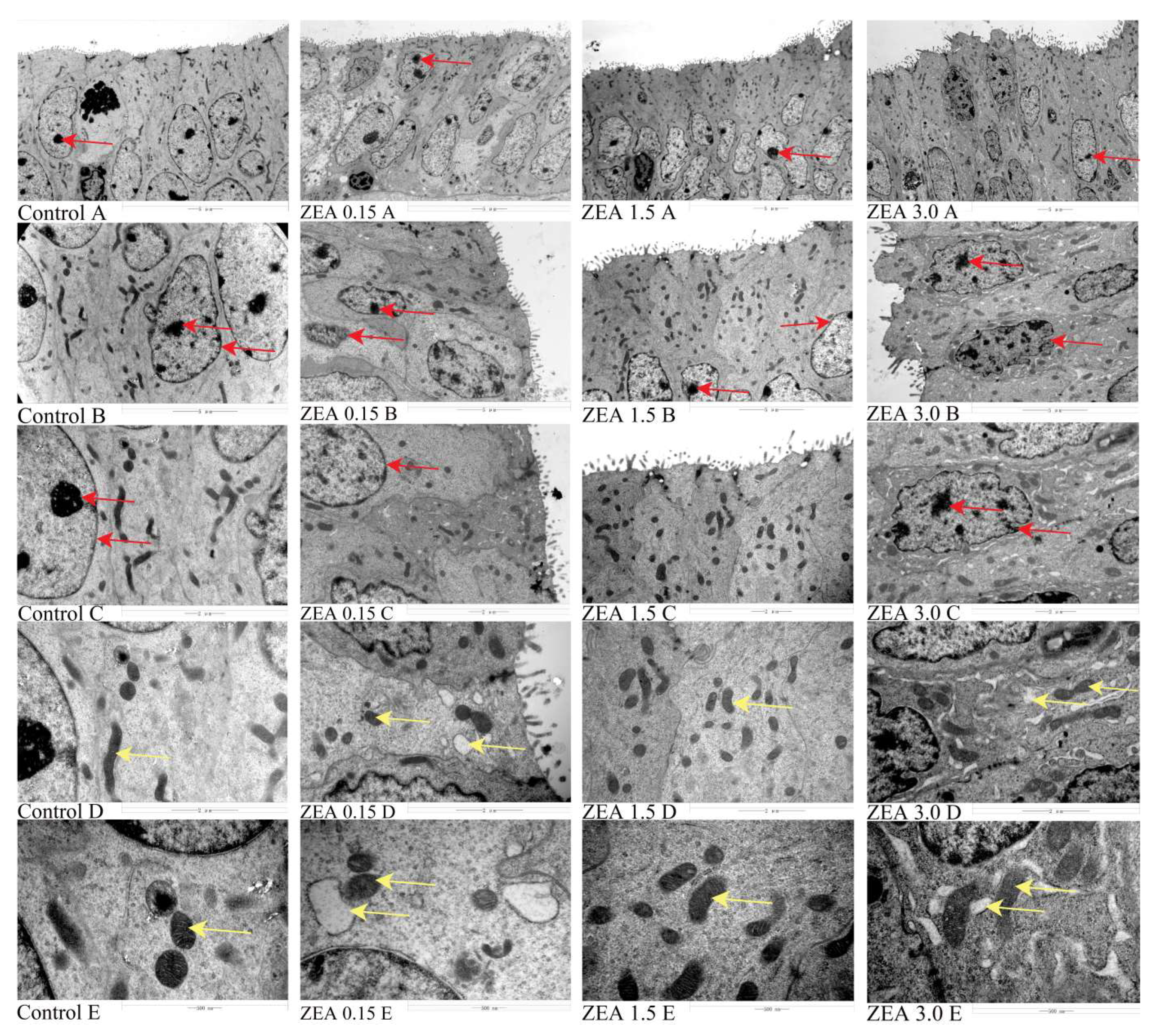
2.2. ZEN-Induced Endometrial Epithelial Cells to Be Active
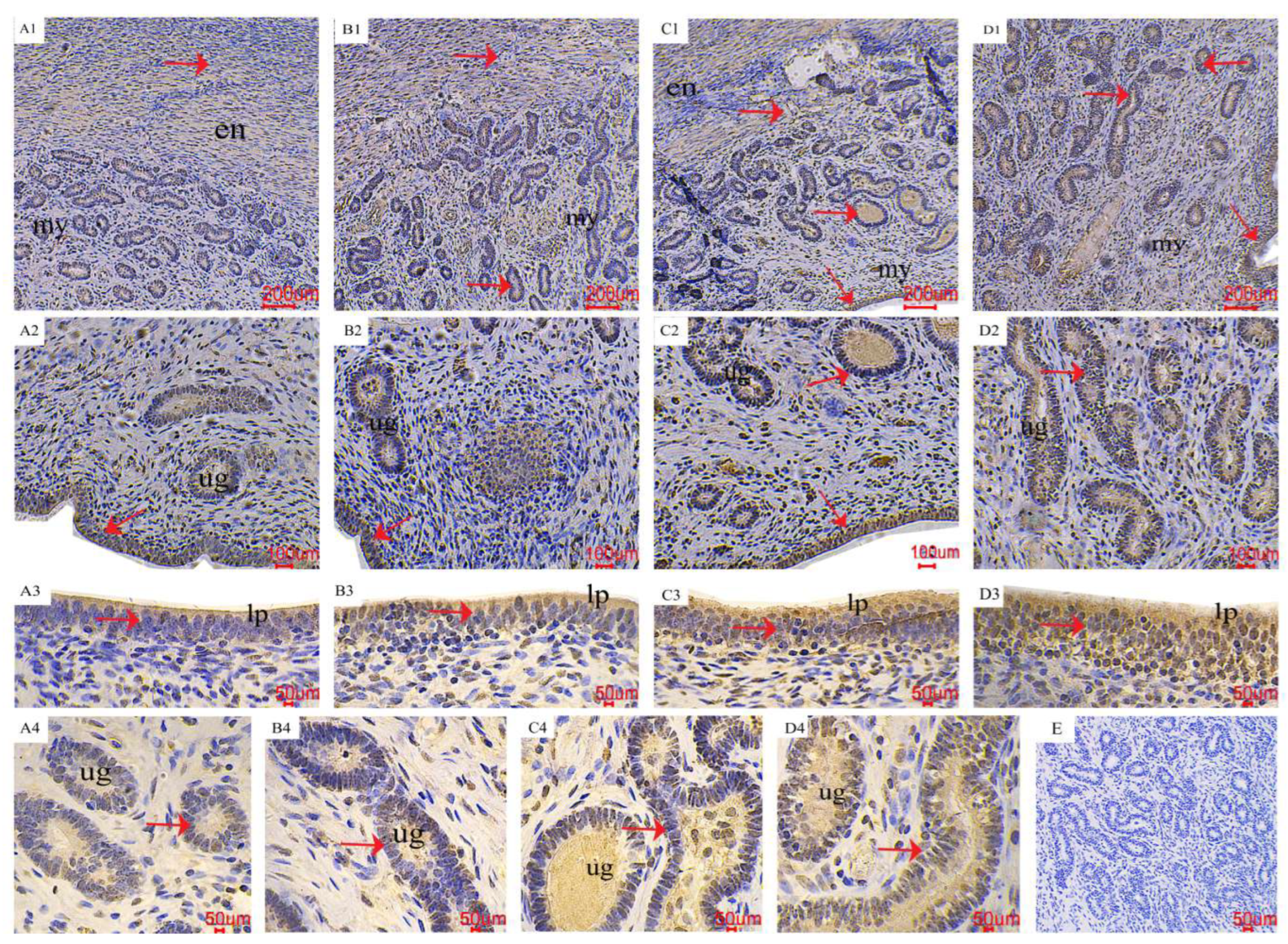
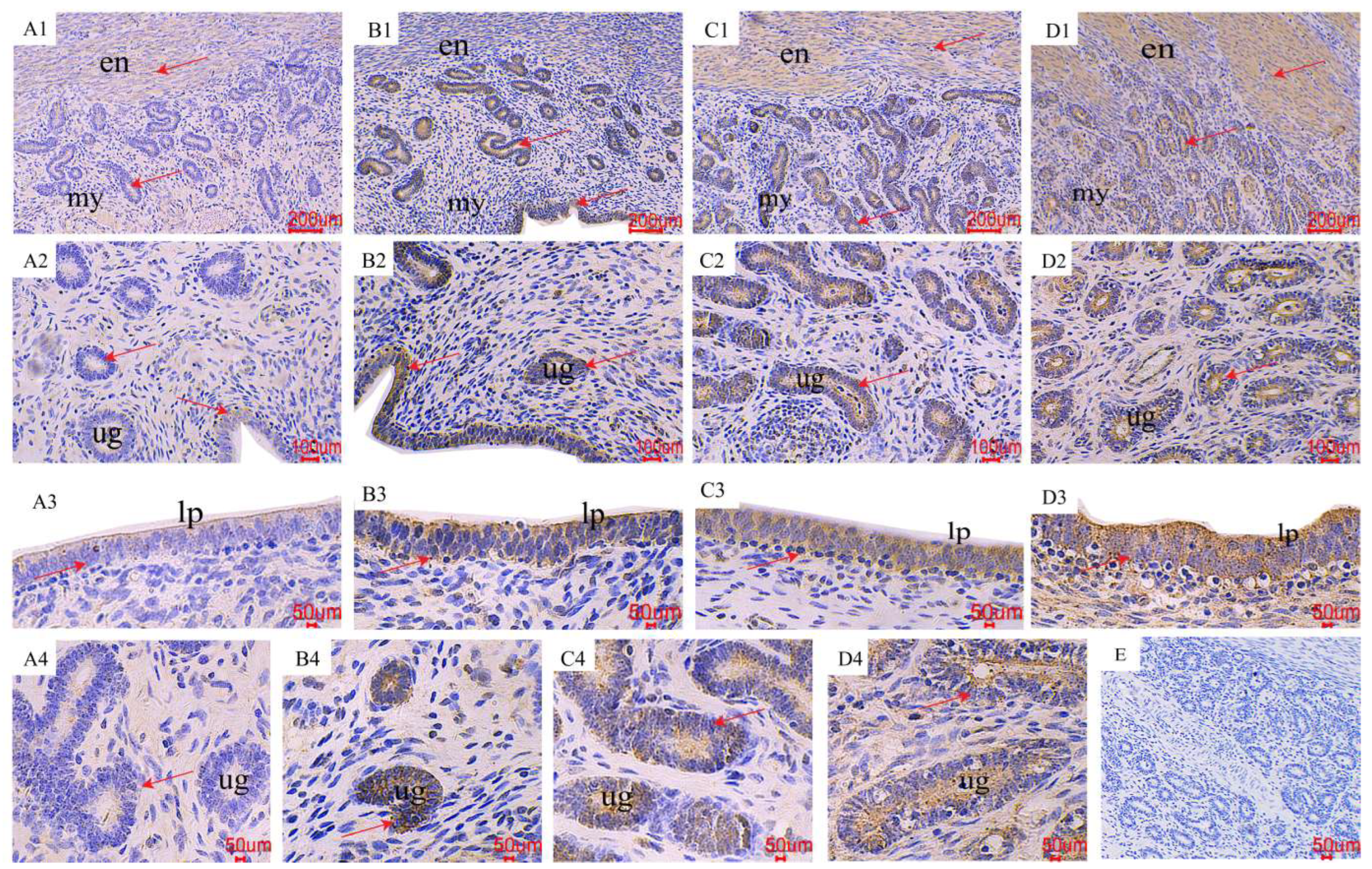
2.3. The Distribution of AMPK/mTOR Pathway-Related Genes in the Uterus
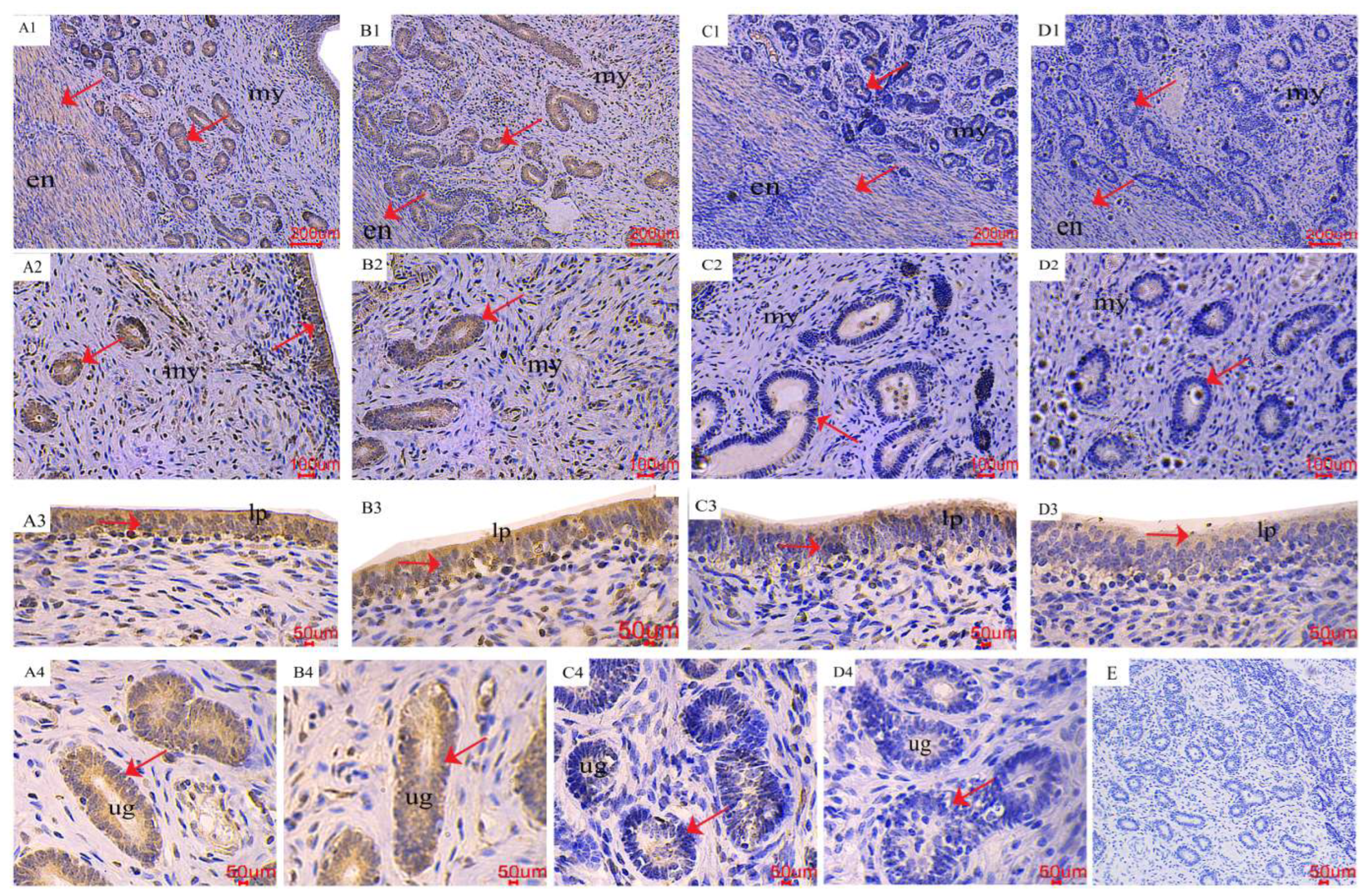
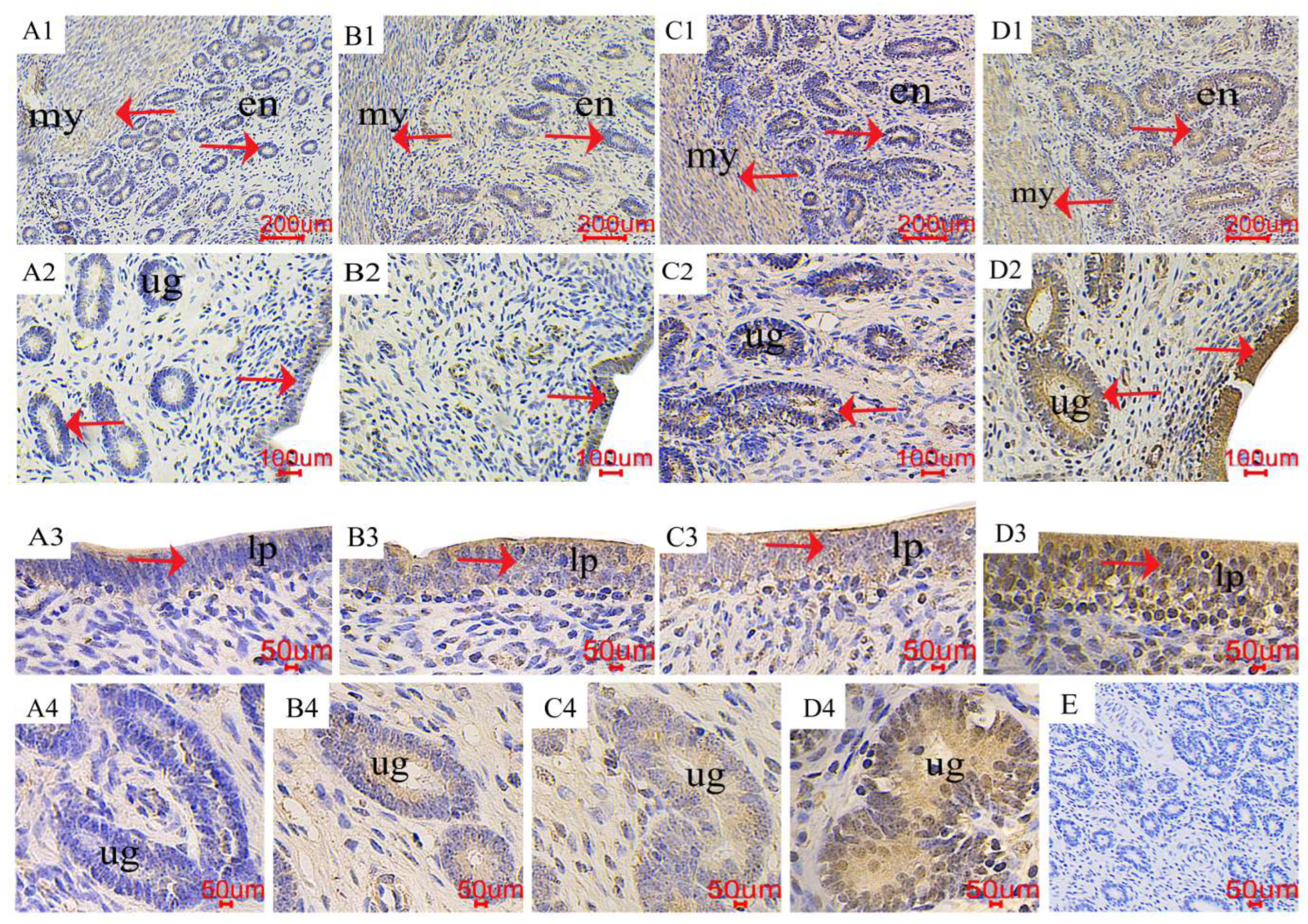
2.4. The Distribution of LC3 and Beclin1 in the Uterus
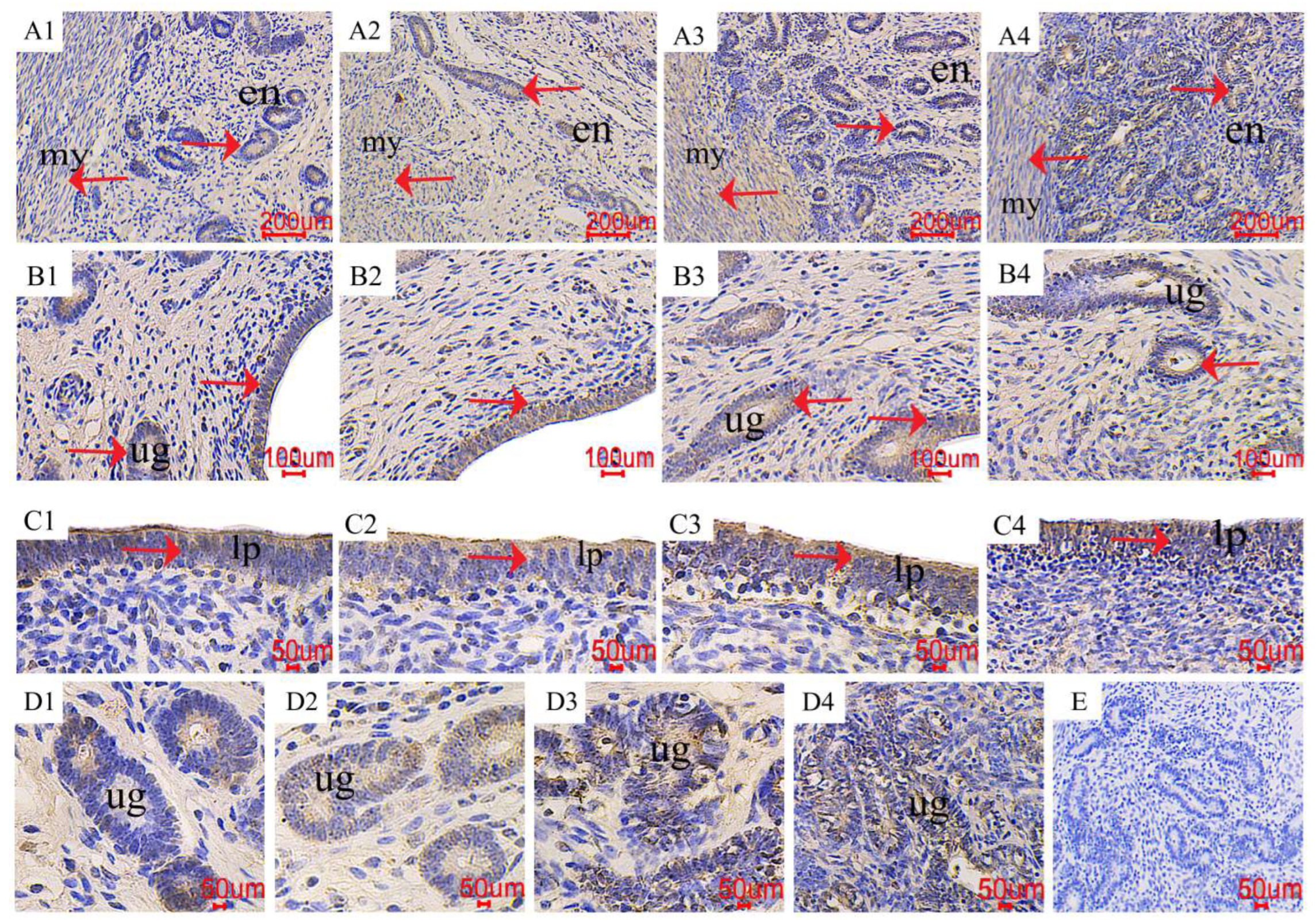
2.5. The Distribution of PCNA in the Uterus
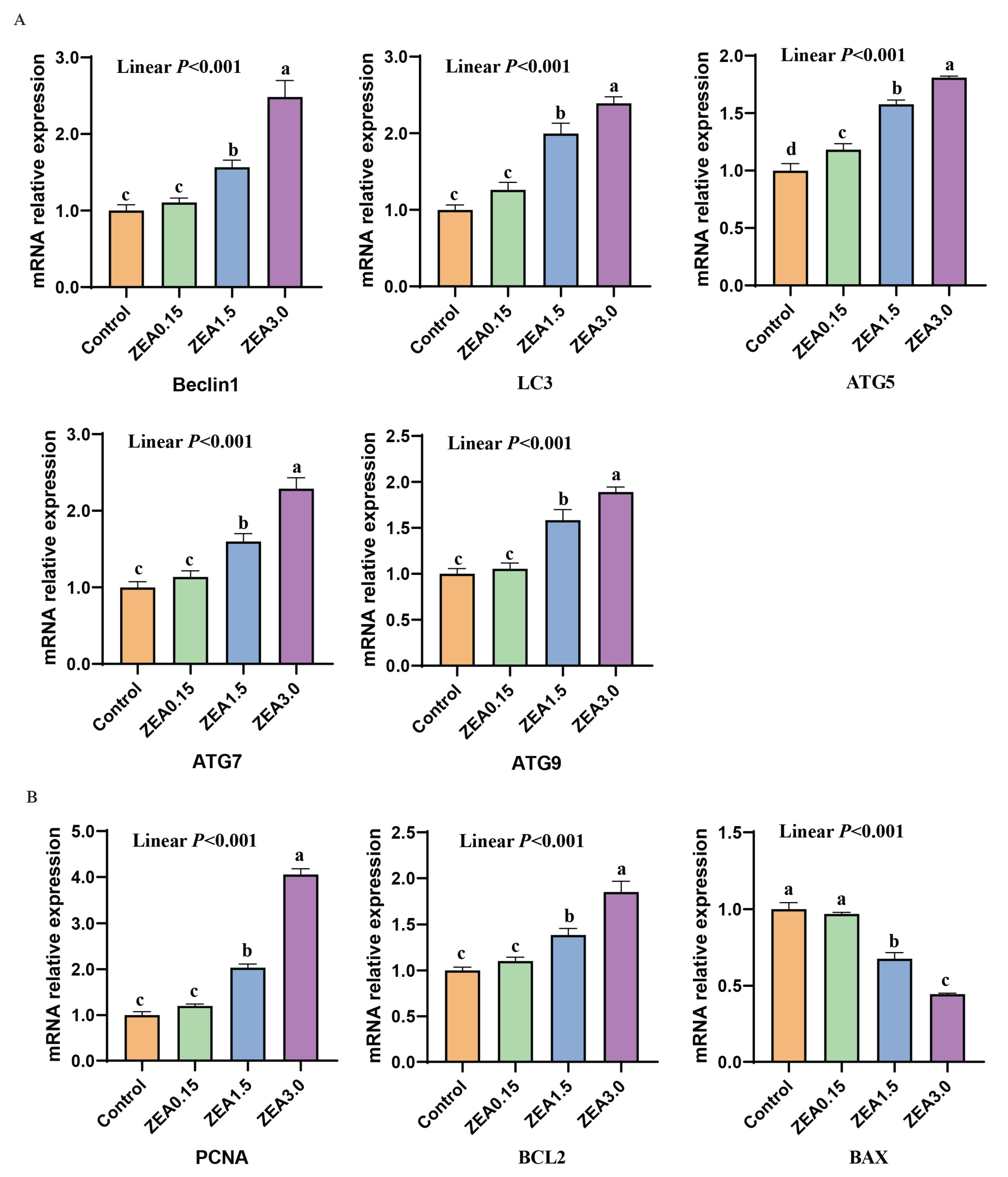
2.6. The Expression of Genes Involved in Autophagy, Proliferation, and Apoptosis
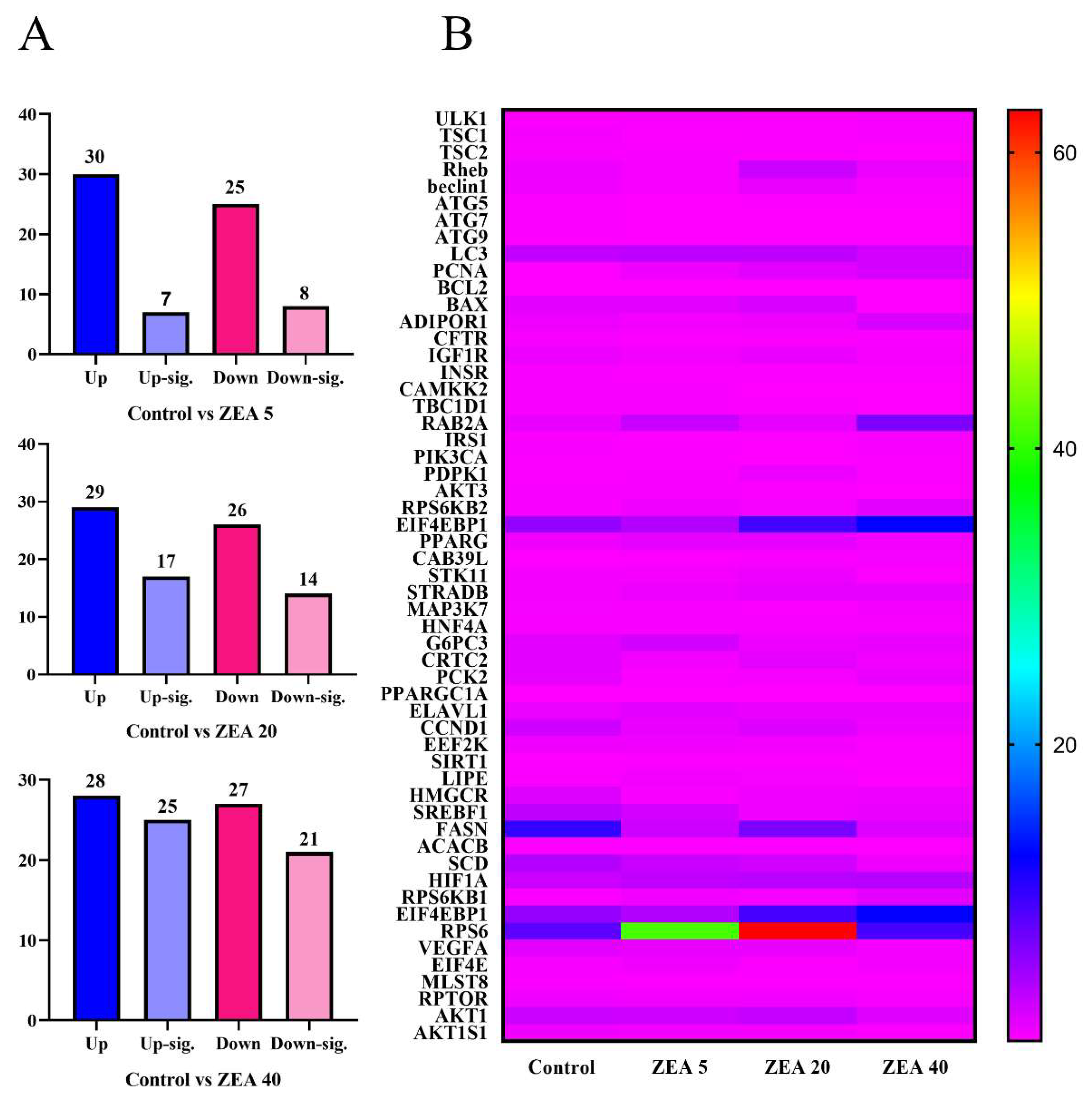
2.7. Transcriptome Analysis of Endometrial Epithelial Cells
2.8. Validation of Relative mRNA and Protein Expression of Candidate Genes
3. Discussion
3.1. ZEN-Induced Uterine Hypertrophy and Endometrial Epithelial Cells to Be Active
3.2. The Pathway of AMPK/mTOR
3.3. Expression of Autophagy-Related Genes
3.4. Expression of Proliferation and Apoptosis-Related Genes
4. Conclusions
5. Material and Methods
5.1. Preparation of ZEN Supplemented Diet
5.2. Animals, Experimental Design, and Management
5.3. Histological Examination
5.4. The Observation of Transmission Electron Microscopy
5.5. Immunohistochemistry
5.6. Total RNA Extraction, cDNA Preparation, and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
5.7. Western Blot Procedure
5.8. Determination of Endometrial Epithelial Cell Activity
5.9. Transcriptome Analysis of Endometrial Epithelial Cells
5.10. Verification of Relative Protein Expression of Genes
5.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Malir, F.; Pickova, D.; Toman, J.; Grosse, Y.; Ostry, V. Hazard characterisation for significant mycotoxins in food. Mycotoxin Res. 2023, 39, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Huangfu, B.; Xu, T.; Xu, W.; Asakiya, C.; Huang, K.; He, X. Research Progress of Safety of Zearalenone: A Review. Toxins 2022, 14, 386. [Google Scholar] [CrossRef]
- Jeyakumar JM, J.; Zhang, M.; Thiruvengadam, M. Determination of mycotoxins by HPLC, LC-ESI-MS/MS, and MALDI-TOF MS in Fusarium species-infected sugarcane. Microb. Pathog. 2018, 123, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Jiang, X.; Sun, J.; Li, X.; Li, X.; Jiao, R.; Peng, Z.; Li, Y.; Bai, W. Toxic effects of zearalenone on gametogenesis and embryonic development: A molecular point of review. Food Chem. Toxicol. 2018, 119, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Zhao, F.; Ye, X.Q. Effects of mycoestrogens on female reproduction. Reprod. Dev. Med. 2018, 2, 52–58. [Google Scholar] [CrossRef]
- Li, L.; Zhang, T.T.; Ren, X.H.; Li, B.X.; Wang, S.Q. Male reproductive toxicity of zearalenone-meta-analysis with mechanism review. Ecotox. Environ. Safe 2021, 221, 112457. [Google Scholar] [CrossRef]
- Lu, Q.; Luo, J.Y.; Ruan, H.N.; Wang, C.J.; Yang, M.H. Structure-toxicity relationships, toxicity mechanisms and health risk assessment of food-borne modified deoxynivalenol and zearalenone: A comprehensive review. Sci. Total Environ. 2022, 806, 151192. [Google Scholar] [CrossRef]
- Zheng, Y.L. The Molecular Mechanisms of ZEA Induced-Cell Apoptosis and Destroyed Cytoskeleton in TM4 Sertoli Cells. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2019. [Google Scholar]
- Chen, Y.; Xing, M.; Chen, T.; Tian, S.; Li, B. Effects and mechanisms of plant bioactive compounds in preventing fungal spoilage and mycotoxin contamination in postharvest fruits: A review. Food Chem. 2023, 415, 135787. [Google Scholar] [CrossRef]
- Guo, Y.; Huo, X.L.; Zhao, Q.; Ma, J.; Zhang, C.; Ji, L. Protective effects of Bacillus subtilis ANSB060, Bacillus subtilis ANSB01G, and Devosia sp. ANSB714-based mycotoxin biodegradation agent on mice fed with naturally moldy diets. Probiotics Antimicrob. Proteins 2020, 12, 994–1001. [Google Scholar] [CrossRef]
- Gatica, D.; Lahiri, V.; Klionsky, D.J. Cargo recognition and degradation by selective autophagy. Nat. Cell Biol. 2018, 20, 233–242. [Google Scholar] [CrossRef]
- Ren, Z.; Guo, C.; Yu, S.; Zhu, L.; Wang, Y.; Hu, H.; Deng, J. Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. Int. J. Mol. Sci. 2019, 20, 2777. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, X.Q. Regulation of mTORC1 by Small GTPases in Response to Nutrients. J. Nutr. 2020, 150, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Hale, C.M.; Cheng, Q.; Ortuno, D.; Huang, M.; Nojima, D.; Kassner, P.D.; Wang, S.; Ollmann, M.M.; Carlisle, H.J. Identification of modulators of autophagic flux in an image-based high content siRNA screen. Autophagy 2016, 12, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Bento, C.F.; Deretic, V. Therapeutic targeting of autophagy in neurodegenerative and infectious diseases. J. Exp. Med. 2015, 212, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Lin, P.F.; Wang, N.; Yang, D.Q.; Wen, X.; Zhou, D.; Wang, A.H.; Jin, Y.P. Herp depletion inhibits zearalenone-induced cell death in RAW 264.7 macrophages. Toxicol. Vitr. 2016, 32, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.M.; Wrobel, L.; Rubinsztein, D.C. Post-translational modifications of Beclin 1 provide multiple strategies for autophagy regulation. Cell Death Differ. 2019, 26, 617–629. [Google Scholar] [CrossRef]
- Kaur, S.; Changotra, H. The beclin 1 interactome: Modification and roles in the pathology of autophagy-related disorders. Biochimie 2020, 175, 34–49. [Google Scholar] [CrossRef]
- Bates, J.H.T.; Herrmann, J.; Casey, D.T.; Béla, S. An agent-based model of tissue maintenance and self-repair. Am. J. Physiol. Cell Physiol. 2023, 324, 941–950. [Google Scholar] [CrossRef]
- Kowalski, S.; Karska, J.; Łapińska, Z.; Hetnał, B.; Saczko, J.; Kulbacka, J. An overview of programmed cell death: Apoptosis and pyroptosis-Mechanisms, differences, and significance in organism physiology and pathophysiology. J. Cell. Biochem. 2023, 124, 765–784. [Google Scholar] [CrossRef]
- Yefimova, M.G.; Ravel, C.; Rolland, A.D.; Bourmeyster, N.; Jégou, B. MERTK-Mediated LC3-Associated Phagocytosis (LAP) of Apoptotic Substrates in Blood-Separated Tissues: Retina, Testis, Ovarian Follicles. Cells 2021, 10, 1443. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Abdellah, Z.; Jose, M.S.; Juan, C.M.; Jordi, M. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the Immune Response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef]
- Lemke, S.L.; Mayura, K.; Reeves, W.R. Investigation of Organophilic Montmorillonite Clay Inclusion in Zearalenone-Contaminated Diets Using the Mouse Uterine Weight Bioassay. J. Toxicol. Environ. Health 2001, 62, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Ahmetovic, N.; Mackic, S. Zearalenone Effect on Uterine Weight of Rats. Agric. Conspec. Sci. 2012, 77, 137–141. [Google Scholar]
- Zhou, M.; Yang, L.; Chen, Y.; Sun, T.; Wang, N.; Chen, X.; Yang, Z.; Ge, J.; Jiang, S. Comparative study of stress response, growth and development of uteri in post-weaning gilts challenged with zearalenone and estradiol benzoate. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1885–1894. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xiao, Z.H.; Liu, M.; Zhang, N.Y.; Khalil, M.M.; Gu, C.Q.; Qi, D.S.; Sun, L.H. Dietary Silymarin Supplementation Alleviates Zearalenone-Induced Hepatotoxicity and Reproductive Toxicity in Rats. J. Nutr. 2018, 148, 1209–1216. [Google Scholar] [CrossRef]
- Wu, L.; Duan, Q.; Gao, D.; Wang, Y.; Xue, S.; Li, W.; Lei, M. Zearalenone Blocks Autophagy Flow and Induces Cell Apoptosis during Embryo Implantation in Gilts. Toxicol. Sci. 2020, 175, 126–139. [Google Scholar] [CrossRef]
- Wang, B.J.; Feng, N.N.; Zheng, W.L.; Xue, H.M.; Da, J.S.; Zou, H.; Gu, J.H.; Yuan, Y.; Liu, X.Z.; Liu, Z.P.; et al. Study on the Role of ERK Signal in Zearalenone Induced Autophagy of Testicular Sertoli Cells. Chin. J. Vet. Sci. 2017, 47, 798–804. [Google Scholar]
- Zheng, W.; Wang, B.; Xi, L.; Wang, T.; Zou, H.; Gu, J.H.; Yuan, Y.; Liu, X.Z.; Bai, J.F.; Bian, J.C.; et al. Zearalenone Promotes Cell Proliferation or Causes Cell Death? Toxins 2018, 10, 184. [Google Scholar] [CrossRef]
- Yoon, J.E.; Lee, K.Y.; Seok, J.S.; Cheng, W.N.; Kwon, H.C.; Jeong, C.H.; Han, S.G. Zearalenone Induces Endoplasmic Reticulum Stress and Modulates the Expression of Phase I/II Enzymes in Human Liver Cells. Toxins 2019, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Song, T.T. Mechanism of Zearalenone Induced Uterine Hypertrophy in Post-Weaning Piglets. Ph.D. Thesis, Shandong Agricultural University, Taian, China, 2020. [Google Scholar]
- Mei, R.; Lou, P.; You, G.; Jiang, T.; Yu, X.; Guo, L. 17β-Estradiol Induces Mitophagy Upregulation to Protect Chondrocytes via the SIRT1-Mediated AMPK/mTOR Signaling Pathway. Front. Endocrinol. 2021, 11, 615250. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Wang, B.; Cai, P.; Zheng, W.L.; Zou, H.; Gu, J.G.; Yuan, Y.; Liu, X.Z.; Liu, Z.P.; Bian, J.C. ZEA-induced autophagy in TM4 cells was mediated by the release of Ca2+ activates CaMKKβ-AMPK signaling pathway in the endoplasmic reticulum. Toxicol. Lett. 2020, 323, 1–9. [Google Scholar] [CrossRef]
- Song, G.; Fang, J.; Shang, C.; Li, Y.; Zhu, Y.; Xiu, Z.; Sun, L.; Jin, N.; Li, X. Ad-apoptin inhibits glycolysis, migration and invasion in lung cancer cells targeting AMPK/mTOR signaling pathway. Exp. Cell Res. 2021, 409, 112926. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Xin, W.; Lin, P.; Chen, H.; Wang, A.; Jin, Y.P. Herp depletion inhibits zearalenone-induced apoptosis through autophagy activation in mouse ovarian granulosa cells. Toxicol. Lett. 2019, 301, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Feng, Z.; Feng, N.; Zou, H.; Gu, J.; Liu, X.; Liu, Z.; Yuan, Y.; Bian, J. Activated AMPK promoted the decrease of lactate production in rat Sertoli cells exposed to Zearalenone. Ecotoxicol. Environ. Saf. 2021, 220, 112367. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Sun, D.J.; Chen, Y.; Ouyang, L. Targeting mTOR for fighting diseases: A revisited review of mTOR inhibitors. Eur. J. Med. Chem. 2020, 199, 112391. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Zhao, S.; Yang, Z.; Yang, W.; Huang, L.; Liu, F.; Liu, M.; Ge, J.; Wang, Y.; Jiang, S. Illicium verum and Eucommia ulmoides leaf extracts promote nutrient availability and antioxidant capacity in piglets by upregulating duodenal and jejunal Nrf2/TNF-α. J. Anim. Physiol. Anim. Nutr. 2021, 105, 916–926. [Google Scholar] [CrossRef]
- Nurdinov, N.; Çınar, V.; Güler, A.; Yilmaz, S.G.; Kökçü, N.D.; Hamurcu, Z. LC3 and Beclin-1 as Markers of Autophagic Activity in Breast Cancer. Erciyes Med. J./Erciyes Tip Derg. 2021, 43, 333–336. [Google Scholar] [CrossRef]
- Lin, X.L.; Xiao, W.J.; Xiao, L.L.; Liu, M.H. Molecular mechanisms of autophagy in cardiac ischemia/reperfusion injury. Mol. Med. Rep. 2018, 18, 675–683. [Google Scholar] [CrossRef]
- Hanada, T.; Noda, N.N.; Satomi, Y.; Ichimura, Y.; Fujioka, Y.; Takao, T.; Inagaki, F.; Ohsumi, Y. The Atg12-Atg5 Conjugate Has a Novel E3-like Activity for Protein Lipidation in Autophagy. J. Biol. Chem. 2007, 282, 37298–37302. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Wang, L.L.; Deng, R.; Tang, J.; Shen, Y.; Guo, J.F.; Wang, Y.; Xia, L.P.; Feng, G.K.; Liu, Q.Q.; et al. The pivotal role of c-Jun NH2-terminal kinase-mediated Beclin 1 expression during anticancer agents-induced autophagy in cancer cells. Oncogene 2009, 28, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Ying, Y.; Ma, F.; Zou, C.; Wang, Y. Zearalenone disrupts the placental function of rats: A possible mechanism causing intrauterine growth restriction. Food Chem. Toxicol. 2020, 145, 111698. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Miao, Y.; Ding, C.; Fan, W.T.; Liu, S.H.; Lv, Y.N.; Gao, X.A.; Boevre, M.D.; Yang, L.P.; Okoth, S.; et al. Activation of the p38/MAPK pathway regulates autophagy in response to the CYPOR-dependent oxidative stress induced by zearalenone in porcine intestinal epithelial cells. Food Chem. Toxicol. 2019, 131, 110527. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Bio. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Kikuchi, S.; Hishiki, A.; Shao, Y.; Heath, R.; Evison, B.J.; Fujii, N.A. Small molecule inhibitor of monoubiquitinated Proliferating Cell Nuclear Antigen (PCNA) inhibits repair of interstrand DNA cross-link, enhances DNA double strand break, and sensitizes cancer cells to cisplatin. J. Biol. Chem. 2014, 289, 7109–7120. [Google Scholar] [CrossRef] [PubMed]
- Molaeeghaleh, N.; Tork, S.; Abdi, S.; Movassaghi, S. Evaluating the effects of dfferent concentrations of human follicular fluid on growth, development, and PCNA gene expression of mouse ovarian follicles. Cells Tissues Organs 2020, 209, 75–82. [Google Scholar] [CrossRef]
- Hong, X.; Mao, Y.H.; Yang, C.Q.; Liu, Z.J.; Li, M.; Du, D.L. Contamination of Zearalenone from China in 2019 by a Visual and Digitized Immunochromatographic Assay. Toxins 2020, 12, 521. [Google Scholar] [CrossRef]
- Catz, S.D.; Johnson, J.L. BCL-2 in prostate cancer: A minireview. Apoptosis 2003, 8, 29–37. [Google Scholar] [CrossRef]
- Knudson, C.M.; Ksk, T.; Tourtellotte, W.G.; Brown GA, J.; Korsmeyer, S.J. Bax-Deficient Mice with Lymphoid Hyperplasia and Male Germ Cell Death. Science 1995, 270, 96–99. [Google Scholar] [CrossRef]
- Dieckman, L. Something’s gotta give: How PCNA alters its structure in response to mutations and the implications on cellular processes. Prog. Biophys. Mol. Biol. 2021, 163, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Moldoveanu, T.; Czabotar, P.E. BAX, BAK, and BOK: A Coming of Age for the BCL-2 Family Effector Proteins. Cold Spring Harb. Perspect. Biol. 2020, 2, a036319. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Deng, Y.; Yuan, L.; Jing, W.; Yuan, Z.; Yi, J.; Zhang, M.; Guo, C.Z.; Wen, L.X.; Li, R.F.; et al. Gynostemma pentaphyllum protects mouse male germ cells against apoptosis caused by zearalenone via bax and bcl-2 regulation. Toxicol. Mech. Method 2010, 20, 153–158. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Liao, W.; Dong, J.; Chen, X.; Huang, L.; Yang, W.; Jiang, S. Zearalenone Promotes Uterine Hypertrophy through AMPK/mTOR Mediated Autophagy. Toxins 2024, 16, 73. https://doi.org/10.3390/toxins16020073
Yang L, Liao W, Dong J, Chen X, Huang L, Yang W, Jiang S. Zearalenone Promotes Uterine Hypertrophy through AMPK/mTOR Mediated Autophagy. Toxins. 2024; 16(2):73. https://doi.org/10.3390/toxins16020073
Chicago/Turabian StyleYang, Lijie, Wenshuang Liao, Jiuyuan Dong, Xiangjin Chen, Libo Huang, Weiren Yang, and Shuzhen Jiang. 2024. "Zearalenone Promotes Uterine Hypertrophy through AMPK/mTOR Mediated Autophagy" Toxins 16, no. 2: 73. https://doi.org/10.3390/toxins16020073
APA StyleYang, L., Liao, W., Dong, J., Chen, X., Huang, L., Yang, W., & Jiang, S. (2024). Zearalenone Promotes Uterine Hypertrophy through AMPK/mTOR Mediated Autophagy. Toxins, 16(2), 73. https://doi.org/10.3390/toxins16020073