Microbial Diversity Impacts Non-Protein Amino Acid Production in Cyanobacterial Bloom Cultures Collected from Lake Winnipeg

 ,
, 
 , and
, and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Establishment and Semi-Purification of Cyanobacterial Cultures from Lake Winnipeg
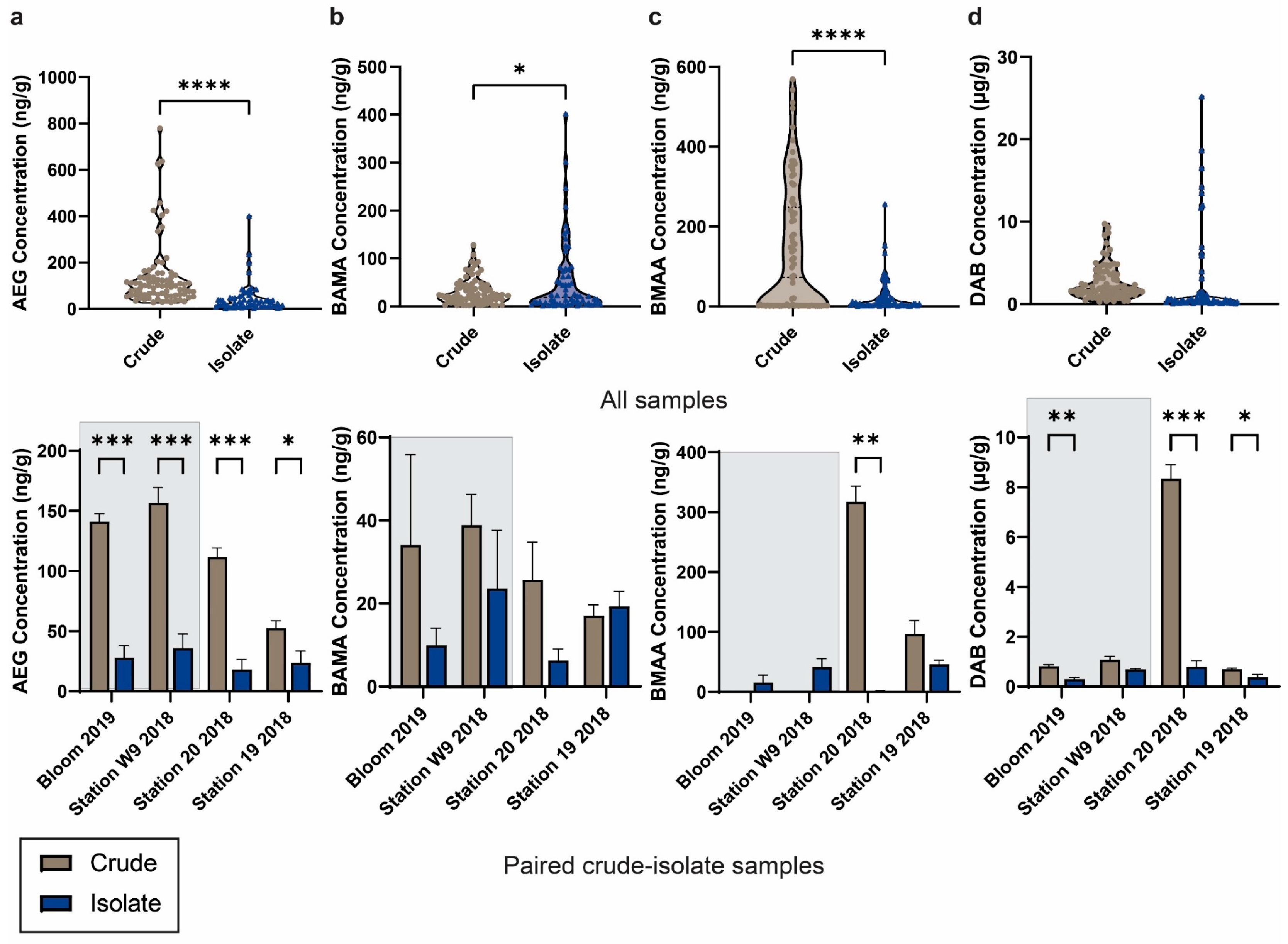
2.2. Distribution of NPAAs in Crude and Isolate Cyanobacterial Cultures
2.2.1. Concentration Profiles of NPAAs across Lake Winnipeg
2.2.2. Occurrence of NPAAs in Paired Crude–Isolate Cyanobacterial Cultures
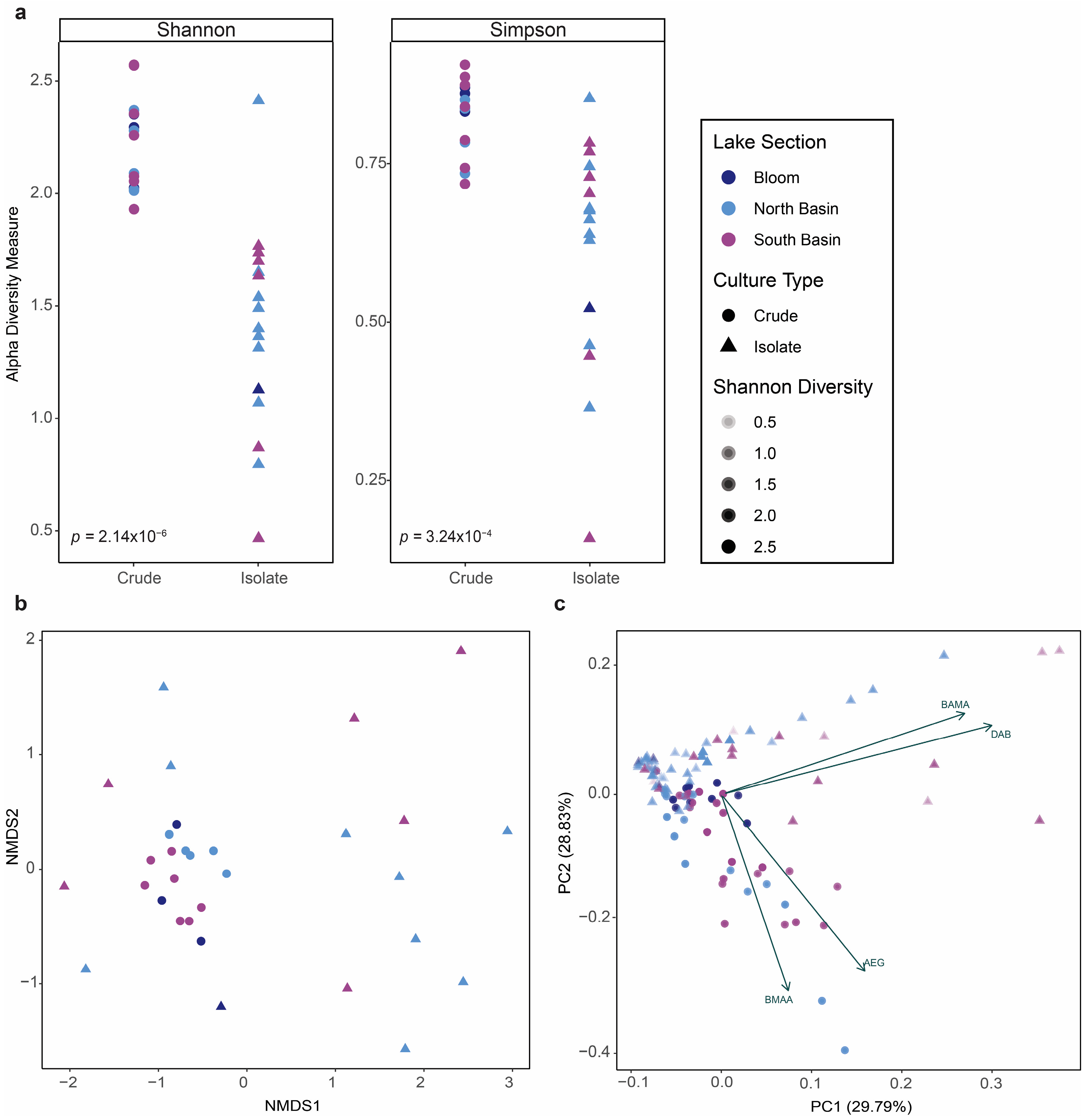
2.3. Analysis of NPAA Profiles and Alpha Diversity in Cyanobacterial Cultures
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Cyanobacterial Culture Establishment
4.1.1. Field Sample Collection
4.1.2. Crude Cyanobacterial Culture Establishment
4.1.3. Isolate Cyanobacterial Culture Establishment
4.2. Detection and Quantification of Non-Protein Amino Acids
4.2.1. Sample Preparation
4.2.2. Derivatization Reaction and Quantification of NPAAs
4.3. Taxonomic Analysis of Cyanobacterial Cultures and Data Visualization
4.3.1. Microscopic Images and Visual Identification of Cyanobacterial Orders
4.3.2. Next-Generation Sequencing (NGS) of Cyanobacterial Cultures
DNA Extraction
16S rRNA Sequencing
DADA2 Taxonomic Identification
4.3.3. Statistical Analysis and Visualization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bashir, F.; Bashir, A.; Bouaïcha, N.; Chen, L.; Codd, G.A.; Neilan, B.; Xu, W.L.; Ziko, L.; Rajput, V.D.; Minkina, T.; et al. Cyanotoxins, Biosynthetic Gene Clusters, and Factors Modulating Cyanotoxin Biosynthesis. World J. Microbiol. Biotechnol. 2023, 39, 241. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W.; Boyer, G.L. Health Impacts from Cyanobacteria Harmful Algae Blooms: Implications for the North American Great Lakes. Harmful Algae 2016, 54, 194–212. [Google Scholar] [CrossRef] [PubMed]
- Brunskill, G.J.; Elliott, S.E.M.; Campbell, P. Morphometry, Hydrology, and Watershed Data Pertinent to the Limnology of Lake Winnipeg; Department of Fisheries and Oceans: Winnipeg, MB, Canada, 1980. [Google Scholar]
- Scott, K.J.; Mccullough, G.; Stainton, M.; Ayles, B.; Hann, B. Lake Winnipeg-State of the Science What Is the Scientific Basis for Understanding and Protecting Lake Winnipeg? Lake Winnipeg Research Consortium Workshop Report: Winnipeg, MB, Canada, 2011. [Google Scholar]
- Wassenaar, L.I.; Rao, Y.R. Lake Winnipeg: The Forgotten Great Lake. J. Great Lakes Res. 2012, 38, 1–5. [Google Scholar] [CrossRef]
- McCullough, G.K.; Page, S.J.; Hesslein, R.H.; Stainton, M.P.; Kling, H.J.; Salki, A.G.; Barber, D.G. Hydrological Forcing of a Recent Trophic Surge in Lake Winnipeg. J. Great Lakes Res. 2012, 38, 95–105. [Google Scholar] [CrossRef]
- Bhateria, R.; Jain, D. Water Quality Assessment of Lake Water: A Review. Sustain. Water Resour. Manag. 2016, 2, 161–173. [Google Scholar] [CrossRef]
- Cottingham, K.L.; Ewing, H.A.; Greer, M.L.; Carey, C.C.; Weathers, K.C. Cyanobacteria as Biological Drivers of Lake Nitrogen and Phosphorus Cycling. Ecosphere 2015, 6, 1–19. [Google Scholar] [CrossRef]
- Spears, B.M.; Carvalho, L.; Perkins, R.; Paterson, D.M. Effects of Light on Sediment Nutrient Flux and Water Column Nutrient Stoichiometry in a Shallow Lake. Water Res. 2008, 42, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Zastepa, A.; Pick, F.R.; Blais, J.M. Fate and Persistence of Particulate and Dissolved Microcystin-LA from Microcystis Blooms. Hum. Ecol. Risk Assess. 2014, 20, 1670–1686. [Google Scholar] [CrossRef]
- Pick, F.R. Blooming Algae: A Canadian Perspective on the Rise of Toxic Cyanobacteria. Can. J. Fish. Aquat. Sci. 2016, 73, 1149–1158. [Google Scholar] [CrossRef]
- Cirés, S.; Ballot, A. A Review of the Phylogeny, Ecology and Toxin Production of Bloom-Forming Aphanizomenon Spp. and Related Species within the Nostocales (Cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Scott, K.J. Lake Winnipeg Research Consortium Inc. 2016/2017 Annual Report, Science and Education Programs; Lake Winnipeg Research Consortium: Winnipeg, MB, Canada, 2017. [Google Scholar]
- Zhang, Y.; Whalen, J.K. Production of the Neurotoxin Beta-N-Methylamino-l-Alanine May Be Triggered by Agricultural Nutrients: An Emerging Public Health Issue. Water Res. 2020, 170, 115335. [Google Scholar] [CrossRef] [PubMed]
- McKindles, K.M.; Zimba, P.V.; Chiu, A.S.; Watson, S.B.; Gutierrez, D.B.; Westrick, J.; Kling, H.; Davis, T.W. A Multiplex Analysis of Potentially Toxic Cyanobacteria in Lake Winnipeg during the 2013 Bloom Season. Toxins 2019, 11, 587. [Google Scholar] [CrossRef]
- Bishop, S.L.; Kerkovius, J.K.; Menard, F.; Murch, S.J. N-β-Methylamino-L-Alanine and Its Naturally Occurring Isomers in Cyanobacterial Blooms in Lake Winnipeg. Neurotox. Res. 2018, 33, 133–142. [Google Scholar] [CrossRef]
- Whiting, M.G. Toxicity of Cycads. Econ. Bot. 1963, 17, 270–302. [Google Scholar] [CrossRef]
- Vega, A.; Bell, E.A. α-Amino-β-Methylaminopropionic Acid, a New Amino Acid from Seeds of Cycas Circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Spencer, P.S.; Nunn, P.B.; Hugon, J.; Ludolph, A.C.; Stephen, M.; Roy, D.N.; Robertson, R.C.; Spencer, P.S.; Nunn, P.B.; Hugon, J.; et al. Guam Amyotrophic Lateral Sclerosis--Parkinsonism--Dementia Linked to a Plant Excitant Neurotoxin. Science 1987, 237, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A Mechanism for Slow Release of Biomagnified Cyanobacterial Neurotoxins and Neurodegenerative Disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse Taxa of Cyanobacteria Produce Beta-N-Methylamino-L-Alanine, a Neurotoxic Amino Acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef]
- Cox, P.A.; Sacks, O.W. Cycad Neurotoxins, Consumption of Flying Foxes, and ALS-PDC Disease in Guam. Neurology 2002, 58, 956–959. [Google Scholar] [CrossRef]
- Yan, B.Y.; Liu, Z.Q.; Huang, R.; Xu, Y.P.; Liu, D.M.; Wang, W.; Zhao, Z.W.; Cui, F.Y.; Shi, W.X. Impact Factors on the Production of Beta-Methylamino-L-Alanine (BMAA) by Cyanobacteria. Chemosphere 2020, 243, 125355. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Banack, S.A.; Bishop, S.L.; Metcalf, J.S.; Murch, S.J.; Davis, D.A.; Stommel, E.W.; Karlsson, O.; Brittebo, E.B.; Chatziefthimiou, A.D.; et al. Is Exposure to BMAA a Risk Factor for Neurodegenerative Diseases? A Response to a Critical Review of the BMAA Hypothesis. Neurotox. Res. 2021, 39, 81–106. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Wang, S.; Zhang, J.Y.; Lin, G.M.; Gan, N.; Song, L.; Zeng, X.; Zhang, C.C. Investigation on Cyanobacterial Production of the Proposed Neurotoxin β-N-Methylamino-L-Alanine (BMAA). Water Biol. Secur. 2023, 2, 100208. [Google Scholar] [CrossRef]
- Bishop, S.L.; Murch, S.J. A Systematic Review of Analytical Methods for the Detection and Quantification of β-: N -Methylamino-l-Alanine (BMAA). Analyst 2020, 145, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Glover, W.B.; Mash, D.C.; Murch, S.J. The Natural Non-Protein Amino Acid N-β-Methylamino-L-Alanine (BMAA) Is Incorporated into Protein during Synthesis. Amino Acids 2014, 46, 2553–2559. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The Non-Protein Amino Acid BMAA Is Misincorporated into Human Proteins in Place of L-Serine Causing Protein Misfolding and Aggregation. PLoS ONE 2013, 8, e75376. [Google Scholar] [CrossRef] [PubMed]
- Han, N.C.; Bullwinkle, T.J.; Loeb, K.F.; Faull, K.F.; Mohler, K.; Rinehart, J.; Ibba, M. The Mechanism of β-N-Methylamino-L-Alanine Inhibition of TRNA Aminoacylation and Its Impact on Misincorporation. J. Biol. Chem. 2020, 295, 1402–1410. [Google Scholar] [CrossRef]
- Korn, A.; Höfling, C.; Zeitschel, U.; Krueger, M.; Roßner, S.; Huster, D. Incorporation of the Nonproteinogenic Amino Acid β-Methylamino-Alanine Affects Amyloid β Fibril Properties and Toxicity. ACS Chem. Neurosci. 2020, 11, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Rush, T.; Zapata, J.; Lobner, D. β-N-Methylamino-l-Alanine Induces Oxidative Stress and Glutamate Release through Action on System Xc-. Exp. Neurol. 2009, 217, 429–433. [Google Scholar] [CrossRef]
- Albano, R.; Lobner, D. Transport of BMAA into Neurons and Astrocytes by System Xc-. Neurotox. Res. 2018, 33, 1–5. [Google Scholar] [CrossRef]
- Schneider, T.; Simpson, C.; Desai, P.; Tucker, M.; Lobner, D. Neurotoxicity of Isomers of the Environmental Toxin L-BMAA. Toxicon 2020, 184, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Lobner, D.; Piana, P.M.T.; Salous, A.K.; Peoples, R.W. β-N-Methylamino-l-Alanine Enhances Neurotoxicity through Multiple Mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Burton, B.; Collins, K.; Brooks, J.; Marx, K.; Renner, A.; Wilcox, K.; Moore, E.; Osowski, K.; Riley, J.; Rowe, J.; et al. The Biotoxin BMAA Promotes Dysfunction via Distinct Mechanisms in Neuroblastoma and Glioblastoma Cells. PLoS ONE 2023, 18, e0278793. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Stallrich, J.; Bereman, M.S. Mixture Designs to Investigate Adverse Effects upon Co-Exposure to Environmental Cyanotoxins. Toxicology 2019, 421, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Bell, E.A.; Nunn, P.B. The Preparation of L- and d-α-Amino-β-Methylaminopropionic Acids and the Identification of the Compound Isolated from Cycas Circinalis as the l-Isomer. Phytochemistry 1968, 7, 1885–1887. [Google Scholar] [CrossRef]
- Davis, D.A.; Cox, P.A.; Banack, S.A.; Lecusay, P.D.; Garamszegi, S.P.; Hagan, M.J.; Powell, J.T.; Metcalf, J.S.; Palmour, R.M.; Beierschmitt, A.; et al. L-Serine Reduces Spinal Cord Pathology in a Vervet Model of Preclinical ALS/MND. J. Neuropathol. Exp. Neurol. 2020, 79, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary Exposure to an Environmental Toxin Triggers Neurofibrillary Tangles and Amyloid Deposits in the Brain. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152397. [Google Scholar] [CrossRef]
- Nunn, P.B.; Codd, G.A. Metabolic Solutions to the Biosynthesis of Some Diaminomonocarboxylic Acids in Nature: Formation in Cyanobacteria of the Neurotoxins 3-N-Methyl-2,3-Diaminopropanoic Acid (BMAA) and 2,4-Diaminobutanoic Acid (2,4-DAB). Phytochemistry 2017, 144, 253–270. [Google Scholar] [CrossRef]
- Bishop, S.L.; Tymm, F.J.M.; Perry, K.; Kerkovius, J.K.; Menard, F.; Brady, A.; Slater, G.; Lim, D.S.S.; Metcalf, J.S.; Banack, S.A.; et al. Early-Earth Nonprotein Amino Acid Metabolites in Modern Cyanobacterial Microbialites. Environ. Chem. Lett. 2020, 18, 467–473. [Google Scholar] [CrossRef]
- Reveillon, D.; Sechet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by Diatoms (Phaeodactylum tricornutum, Chaetoceros Sp., Chaetoceros calcitrans and, Thalassiosira pseudonana) and Bacteria Isolated from a Diatom Culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef]
- Mantas, M.J.Q.; Nunn, P.B.; Codd, G.A.; Barker, D. Genomic Insights into the Biosynthesis and Physiology of the Cyanobacterial Neurotoxin 3-N-Methyl-2,3-Diaminopropanoic Acid (BMAA). Phytochemistry 2022, 200, 113198. [Google Scholar] [CrossRef] [PubMed]
- Pokorny, L.; Hausmann, B.; Pjevac, P.; Schagerl, M. How to Verify Non-Presence—The Challenge of Axenic Algae Cultivation. Cells 2022, 11, 2594. [Google Scholar] [CrossRef] [PubMed]
- Glover, W.B.; Baker, T.C.; Murch, S.J.; Bnown, P.N. Determination of Beta-N-Methylamino-L-Alanine, N-(2-Aminoethyl)Glycine, and 2,4-Diaminobutyric Acid in Food Products Containing Cyanobacteria by Ultra-Performance Liquid Chromatography and Tandem Mass Spectrometry: Single-Laboratory Validation. J. AOAC Int. 2015, 98, 1559–1565. [Google Scholar] [CrossRef]
- Koksharova, O.A.; Safronova, N.A. Non-Proteinogenic Amino Acid β-N-Methylamino-L-Alanine (BMAA): Bioactivity and Ecological Significance. Toxins 2022, 14, 539. [Google Scholar] [CrossRef]
- Lage, S.; Strom, L.; Godhe, A.; Rydberg, S. Kinetics of Beta-N-Methylamino-L-Alanine (BMAA) and 2, 4-Diaminobutyric Acid (DAB) Production by Diatoms: The Effect of Nitrogen. Eur. J. Phycol. 2019, 54, 115–125. [Google Scholar] [CrossRef]
- Kuzyk, S.B.; Ma, X.; Yurkov, V. Seasonal Dynamics of Lake Winnipeg’s Microbial Communities Reveal Aerobic Anoxygenic Phototrophic Populations Coincide with Sunlight Availability. Microorganisms 2022, 10, 1690. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Tanasupawat, S.; Nakajima, K.; Kanamori, H.; Ishizaki, S.; Kodama, K.; Okamoto-Kainuma, A.; Koizumi, Y.; Yamamoto, Y.; Yamasato, K. Alkalibacterium thalassium Sp. Nov., Alkalibacterium pelagium Sp. Nov., Alkalibacterium putridalgicola Sp. Nov. and Alkalibacterium kapii Sp. Nov., Slightly Halophilic and Alkaliphilic Marine Lactic Acid Bacteria Isolated from Marine Organisms and Salted Foods Collected in Japan and Thailand. Int. J. Syst. Evol. Microbiol. 2009, 59, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Koerner, R.J.; Goodfellow, M.; Jones, A.L. The Genus Dietzia: A New Home for Some Known and Emerging Opportunist Pathogens. FEMS Immunol. Med. Microbiol. 2009, 55, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Li, A.F.; Tian, Z.J.; Li, J.; Yu, R.C.; Banack, S.A.; Wang, Z.Y. Detection of the Neurotoxin BMAA within Cyanobacteria Isolated from Freshwater in China. Toxicon 2010, 55, 947–953. [Google Scholar] [CrossRef]
- Esterhuizen, M.; Downing, T.G. Beta-N-Methylamino-L-Alanine (BMAA) in Novel South African Cyanobacterial Isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [Google Scholar] [CrossRef]
- Błaszczyk, A.; Siedlecka-Kroplewska, K.; Woźniak, M.; Mazur-Marzec, H. Presence of SS-N-Methylamino-L-Alanine in Cyanobacteria and Aquatic Organisms from Waters of Northern Poland; BMAA Toxicity Studies. Toxicon 2021, 194, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Gross, M. The Mysteries of the Diatoms. Curr. Biol. 2012, 22, R581–R585. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Burian, A.; Rasmussen, U.; Costa, P.R.; Annadotter, H.; Godhe, A.; Rydberg, S. BMAA Extraction of Cyanobacteria Samples: Which Method to Choose? Environ. Sci. Pollut. Res. 2016, 23, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A Novel Source for the Neurotoxin BMAA in Aquatic Environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef] [PubMed]
- Violi, J.P.; Facey, J.A.; Mitrovic, S.M.; Colville, A.; Rodgers, K.J. Production of β-Methylamino-L-Alanine (BMAA) and Its Isomers by Freshwater Diatoms. Toxins 2019, 11, 512. [Google Scholar] [CrossRef] [PubMed]
- Li, A.F.; Yan, Y.J.; Qiu, J.B.; Yan, G.W.; Zhao, P.; Li, M.; Ji, Y.; Wang, G.X.; Meng, F.P.; Li, Y.; et al. Putative Biosynthesis Mechanism of the Neurotoxin Beta-N-Methylamino-L-Alanine in Marine Diatoms Based on a Transcriptomics Approach. J. Hazard. Mater. 2023, 441, 129953. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.A.; Metcalf, J.S.; Cox, P.A.; Downing, T.G. Nitrogen Starvation of Cyanobacteria Results in the Production of Beta-N-Methylamino-L-Alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Downing, S.; Downing, T.G. Improved Sensitivity Using Liquid Chromatography Mass Spectrometry (LC-MS) for Detection of Propyl Chloroformate Derivatised Beta-N-Methylamino-L-Alanine (BMAA) in Cyanobacteria. Water SA 2011, 37, 133–138. [Google Scholar] [CrossRef]
- Peters, S.J.; Rodgers, K.J.; Mitrovic, S.M.; Bishop, D.P. The Changes in Cyanobacterial Concentration of β-Methylamino-L-Alanine during a Bloom Event. Molecules 2022, 27, 7382. [Google Scholar] [CrossRef]
- Berntzon, L.; Erasmie, S.; Celepli, N.; Eriksson, J.; Rasmussen, U.; Bergman, B. BMAA Inhibits Nitrogen Fixation in the Cyanobacterium Nostoc Sp. PCC 7120. Mar. Drugs 2013, 11, 3091–3108. [Google Scholar] [CrossRef]
- Popova, A.A.; Rasmussen, U.; Semashko, T.A.; Govorun, V.M.; Koksharova, O.A. Stress Effects of Cyanotoxin Beta-Methylamino-L-Alanine (BMAA) on Cyanobacterial Heterocyst Formation and Functionality. Environ. Microbiol. Rep. 2018, 10, 369–377. [Google Scholar] [CrossRef]
- Popova, A.A.; Semashko, T.A.; Kostina, N.V.; Rasmussen, U.; Govorun, V.M.; Koksharova, O.A. The Cyanotoxin BMAA Induces Heterocyst Specific Gene Expression in Anabaena Sp. PCC 7120 under Repressive Conditions. Toxins 2018, 10, 478. [Google Scholar] [CrossRef]
- Jiao, Y.Y.; Chen, Q.K.; Chen, X.; Wang, X.; Liao, X.W.; Jiang, L.J.; Wu, J.; Yang, L.Y. Occurrence and Transfer of a Cyanobacterial Neurotoxin Beta-Methylamino-L-Alanine within the Aquatic Food Webs of Gonghu Bay (Lake Taihu, China) to Evaluate the Potential Human Health Risk. Sci. Total Environ. 2014, 468, 457–463. [Google Scholar] [CrossRef]
- Pip, E.; Munford, K.E.; Bowman, L. Seasonal Nearshore Occurrence of the Neurotoxin β-N-Methylamino-L-Alanine (BMAA) in Lake Winnipeg, Canada. Environ. Pollut. 2016, 5, 110. [Google Scholar] [CrossRef]
- Gutiérrez-García, K.; Whitaker, M.R.L.; Bustos-Díaz, E.D.; Salzman, S.; Ramos-Aboites, H.E.; Reitz, Z.L.; Pierce, N.E.; Cibrián-Jaramillo, A.; Barona-Gómez, F. Gut Microbiomes of Cycad-Feeding Insects Tolerant to β-Methylamino-L-Alanine (BMAA) Are Rich in Siderophore Biosynthesis. ISME Commun. 2023, 3, 122. [Google Scholar] [CrossRef]
- Kost, C.; Patil, K.R.; Friedman, J.; Garcia, S.L.; Ralser, M. Metabolic Exchanges Are Ubiquitous in Natural Microbial Communities. Nat. Microbiol. 2023, 8, 2244–2252. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.L.; Drikic, M.; Wacker, S.; Chen, Y.Y.; Kozyrskyj, A.L.; Lewis, I.A. Moving beyond Descriptive Studies: Harnessing Metabolomics to Elucidate the Molecular Mechanisms Underpinning Host-Microbiome Phenotypes. Mucosal Immunol. 2022, 15, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Glover, W.B.; Murch, S.J. Analysis of B-Methylamino-Alanine in Environmental and Food Samples. In Phytochemicals-Biosynthesis, Function and Application; Jetter, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 149–174. [Google Scholar]
- EPA Definition and Procedure for the Determination of the Method Detection Limit, Revision 2; United States Environmental Protection Agency Office of Science and Technology: Washington, DC, USA, 2016.
- Komárek, J.; Katovský, J.; Mare, J.; Johansen, J.R. Taxonomic Classification of Cyanoprokaryotes (Cyanobacterial Genera) 2014, Using a Polyphasic Approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Nienaber, M.A.; Steinitz-Kannan, M. Identification and Impact; University Press of Kentucky: Lexington, KY, USA, 2018; ISBN 9780813175591. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Lefler, F.W.; Berthold, D.E.; Dail Laughinghouse IV, H. CyanoSeq: A Database of Cyanobacterial 16S RRNA Sequences with Curated Taxonomy. J. Phycol. 2023, 59, 470–480. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, S.; Ooms, J. Classes for “GeoJSON”. Available online: https://github.com/ropensci/geojson, https://docs.ropensci.org/geojson/ (accessed on 18 April 2023).
- Agriculture and Agri Food Canada. Lake Winnipeg Outline; Canadian Watershed Information Network, V2; University of Manitoba Centre for Earth Observation Science: Winnipeg, Canada, 2017. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Basel, Switzerland, 2016. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bishop, S.L.; Solonenka, J.T.; Giebelhaus, R.T.; Bakker, D.T.R.; Li, I.T.S.; Murch, S.J. Microbial Diversity Impacts Non-Protein Amino Acid Production in Cyanobacterial Bloom Cultures Collected from Lake Winnipeg. Toxins 2024, 16, 169. https://doi.org/10.3390/toxins16040169
Bishop SL, Solonenka JT, Giebelhaus RT, Bakker DTR, Li ITS, Murch SJ. Microbial Diversity Impacts Non-Protein Amino Acid Production in Cyanobacterial Bloom Cultures Collected from Lake Winnipeg. Toxins. 2024; 16(4):169. https://doi.org/10.3390/toxins16040169
Chicago/Turabian StyleBishop, Stephanie L., Julia T. Solonenka, Ryland T. Giebelhaus, David T. R. Bakker, Isaac T. S. Li, and Susan J. Murch. 2024. "Microbial Diversity Impacts Non-Protein Amino Acid Production in Cyanobacterial Bloom Cultures Collected from Lake Winnipeg" Toxins 16, no. 4: 169. https://doi.org/10.3390/toxins16040169
APA StyleBishop, S. L., Solonenka, J. T., Giebelhaus, R. T., Bakker, D. T. R., Li, I. T. S., & Murch, S. J. (2024). Microbial Diversity Impacts Non-Protein Amino Acid Production in Cyanobacterial Bloom Cultures Collected from Lake Winnipeg. Toxins, 16(4), 169. https://doi.org/10.3390/toxins16040169