The Role of Lymphostatin/EHEC Factor for Adherence-1 in the Pathogenesis of Gram Negative Infection

{kind=link}
Abstract
:1. Introduction
2. Molecular Studies
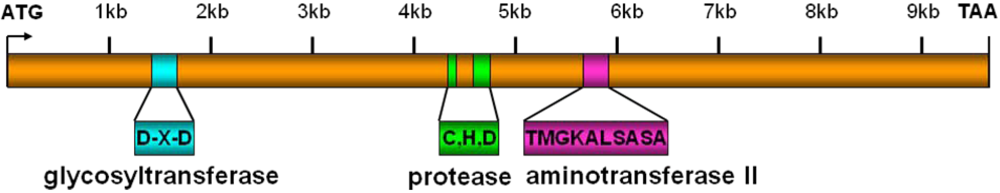
3. Functions of Lymphostatin/EFA-1

4. Conclusion and Future Outlook
References
- Farthing, M.J. Diarrhoea: A significant worldwide problem. Int. J. Antimicrob. Agents 2000, 14, 65–69. [Google Scholar] [CrossRef]
- Clarke, S.C.; Haigh, R.D.; Freestone, P.P.; Williams, P.H. Virulence of enteropathogenic Escherichia coli, a global pathogen. Clin. Microbiol. Rev. 2003, 16, 365–378. [Google Scholar] [CrossRef]
- Welinder-Olsson, C.; Kaijser, B. Enterohemorrhagic Escherichia coli (EHEC). Scand. J. Infect. Dis. 2005, 37, 405–416. [Google Scholar] [CrossRef]
- Noel, J.M.; Boedeker, E.C. Enterohemorrhagic Escherichia coli: a family of emerging pathogens. Dig. Dis. 1997, 15, 67–91. [Google Scholar] [CrossRef]
- Tobe, T.; Hayashi, T.; Han, C.G.; Schoolnik, G.K.; Ohtsubo, E.; Sasakawa, C. Complete DNA sequence and structural analysis of the enteropathogenic Escherichia coli adherence factor plasmid. Infect. Immun. 1999, 67, 5455–5462. [Google Scholar]
- McDaniel, T.K.; Jarvis, K.G.; Donnenberg, M.S.; Kaper, J.B. A genetic locus of enterocyte effacement conserved among diverse enterobacterial pathogens. Proc. Natl. Acad. Sci. USA 1995, 92, 1664–1668. [Google Scholar]
- Hicks, S.; Frankel, G.; Kaper, J.B.; Dougan, G.; Phillips, A.D. Role of intimin and bundle-forming pili in enteropathogenic Escherichia coli adhesion to pediatric intestinal tissue in vitro. Infect. Immun. 1998, 66, 1570–1578. [Google Scholar]
- Chong, Y.; Fitzhenry, R.; Heuschkel, R.; Torrente, F.; Frankel, G.; Phillips, A.D. Human intestinal tissue tropism in Escherichia coli O157: H7-initial colonization of terminal ileum and Peyer's patches and minimal colonic adhesion ex vivo. Microbiology 2007, 153, 794–802. [Google Scholar]
- Mead, P.S.; Griffin, P.M. Escherichia coli O157:H7. Lancet 1998, 352, 1207–1212. [Google Scholar]
- Klapproth, J.M.; Scaletsky, I.C.; McNamara, B.P.; Lai, L.C.; Malstrom, C.; James, S.P.; Donnenberg, M.S. A large toxin from pathogenic Escherichia coli strains that inhibits lymphocyte activation. Infect. Immun. 2000, 68, 2148–2155. [Google Scholar]
- Ogura, Y.; Ooka, T.; Iguchi, A.; Toh, H.; Asadulghani, M.; Oshima, K.; Kodama, T.; Abe, H.; Nakayama, K.; Kurokawa, K.; Tobe, T.; Hattori, M.; Hayashi, T. Comparative genomics reveal the mechanism of the parallel evolution of O157 and non-O157 enterohemorrhagic Escherichia coli. Proc. Natl. Acad. Sci. USA 2009, 106, 17939–17944. [Google Scholar]
- Afset, J.E.; Anderssen, E.; Bruant, G.; Harel, J.; Wieler, L.; Bergh, K. Phylogenetic backgrounds and virulence profiles of atypical enteropathogenic Escherichia coli strains from a case-control study using multilocus sequence typing and DNA microarray analysis. J. Clin. Microbiol. 2008, 46, 2280–2290. [Google Scholar]
- Morabito, S.; Tozzoli, R.; Oswald, E.; Caprioli, A. A mosaic pathogenicity island made up of the locus of enterocyte effacement and a pathogenicity island of Escherichia coli O157:H7 is frequently present in attaching and effacing E. coli. Infect. Immun. 2003, 71, 3343–3348. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Sonntag, A.K.; Schmidt, M.A.; Karch, H. Presence of virulence and fitness gene modules of enterohemorrhagic Escherichia coli in atypical enteropathogenic Escherichia coli O26. Microbes Infect. 2007, 9, 891–897. [Google Scholar] [CrossRef]
- Janka, A.; Bielaszewska, M.; Dobrindt, U.; Karch, H. Identification and distribution of the enterohemorrhagic Escherichia coli factor for adherence (efa1) gene in sorbitol-fermenting Escherichia coli O157: H. Int. J. Med. Microbiol. 2002, 292, 207–214. [Google Scholar]
- Nicholls, L.; Grant, T.H.; Robins-Browne, R.M. Identification of a novel genetic locus that is required for in vitro adhesion of a clinical isolate of enterohaemorrhagic Escherichia coli to epithelial cells. Mol. Microbiol. 2000, 35, 275–288. [Google Scholar]
- Szalo, I.M.; Goffaux, F.; Pirson, V.; Pierard, D.; Ball, H.; Mainil, J. Presence in bovine enteropathogenic (EPEC) and enterohaemorrhagic (EHEC) Escherichia coli of genes encoding for putative adhesins of human EHEC strains. Res. Microbiol. 2002, 153, 653–658. [Google Scholar]
- Badea, L.; Doughty, S.; Nicholls, L.; Sloan, J.; Robins-Browne, R.M.; Hartland, E.L. Contribution of Efa1/LifA to the adherence of enteropathogenic Escherichia coli to epithelial cells. Microb. Pathog. 2003, 34, 205–215. [Google Scholar] [CrossRef]
- Tauschek, M.; Strugnell, R.A.; Robins-Browne, R.M. Characterization and evidence of mobilization of the LEE pathogenicity island of rabbit-specific strains of enteropathogenic Escherichia coli. Mol. Microbiol. 2002, 44, 1533–1550. [Google Scholar] [CrossRef]
- Zhu, C.; Agin, T.S.; Elliott, S.J.; Johnson, L.A.; Thate, T.E.; Kaper, J.B.; Boedeker, E.C. Complete nucleotide sequence and analysis of the locus of enterocyte Effacement from rabbit diarrheagenic Escherichia coli RDEC-1. Infect. Immun. 2001, 69, 2107–2115. [Google Scholar]
- Klapproth, J.M.; Sasaki, M.; Sherman, M.; Babbin, B.; Donnenberg, M.S.; Fernandes, P.J.; Scaletsky, I.C.; Kalman, D.; Nusrat, A.; Williams, I.R. Citrobacter rodentium lifA/efa1 is essential for colonic colonization and crypt cell hyperplasia in vivo. Infect Immun 2005, 73, 1441–1451. [Google Scholar]
- Tatsuno, I.; Horie, M.; Abe, H.; Miki, T.; Makino, K.; Shinagawa, H.; Taguchi, H.; Kamiya, S.; Hayashi, T.; Sasakawa, C. toxB gene on pO157 of enterohemorrhagic Escherichia coli O157:H7 is required for full epithelial cell adherence phenotype. Infect. Immun. 2001, 69, 6660–6669. [Google Scholar]
- Perna, N.T.; Plunkett, G., 3rd; Burland, V.; Mau, B.; Glasner, J.D.; Rose, D.J.; Mayhew, G.F.; Evans, P.S.; Gregor, J.; Kirkpatrick, H.A.; Posfai, G.; Hackett, J.; Klink, S.; Boutin, A.; Shao, Y.; Miller, L.; Grotbeck, E.J.; Davis, N.W.; Lim, A.; Dimalanta, E.T.; Potamousis, K.D.; Apodaca, J.; Anantharaman, T.S.; Lin, J.; Yen, G.; Schwartz, D.C.; Welch, R.A.; Blattner, F.R. Genome sequence of enterohaemorrhagic Escherichia coli O157:H7. Nature 2001, 409, 529–533. [Google Scholar] [PubMed]
- Xie, G.; Bonner, C.A.; Jensen, R.A. Dynamic diversity of the tryptophan pathway in chlamydiae: Reductive evolution and a novel operon for tryptophan recapture. Genome Biol. 2002, 3, research0051. [Google Scholar]
- Belland, R.J.; Scidmore, M.A.; Crane, D.D.; Hogan, D.M.; Whitmire, W.; McClarty, G.; Caldwell, H.D. Chlamydia trachomatis cytotoxicity associated with complete and partial cytotoxin genes. Proc. Natl. Acad. Sci. USA 2001, 98, 13984–13989. [Google Scholar]
- Afset, J.E.; Bruant, G.; Brousseau, R.; Harel, J.; Anderssen, E.; Bevanger, L.; Bergh, K. Identification of virulence genes linked with diarrhea due to atypical enteropathogenic Escherichia coli by DNA microarray analysis and PCR. J. Clin. Microbiol. 2006, 44, 3703–3711. [Google Scholar]
- Vieira, M.A.; Salvador, F.A.; Silva, R.M.; Irino, K.; Vaz, T.M.; Rockstroh, A.C.; Guth, B.E.; Gomes, T.A. Prevalence and characteristics of the O122 pathogenicity island in typical and atypical enteropathogenic Escherichia coli strains. J. Clin. Microbiol. 2010, 48, 1452–1455. [Google Scholar]
- Jores, J.; Wagner, S.; Rumer, L.; Eichberg, J.; Laturnus, C.; Kirsch, P.; Schierack, P.; Tschape, H.; Wieler, L.H. Description of a 111-kb pathogenicity island (PAI) encoding various virulence features in the enterohemorrhagic E. coli (EHEC) strain RW1374 (O103:H2) and detection of a similar PAI in other EHEC strains of serotype 0103:H2. Int. J. Med. Microbiol. 2005, 294, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Wickham, M.E.; Lupp, C.; Mascarenhas, M.; Vazquez, A.; Coombes, B.K.; Brown, N.F.; Coburn, B.A.; Deng, W.; Puente, J.L.; Karmali, M.A.; Finlay, B.B. Bacterial genetic determinants of non-O157 STEC outbreaks and hemolytic-uremic syndrome after infection. J. Infect. Dis. 2006, 194, 819–827. [Google Scholar]
- Karmali, M.A.; Mascarenhas, M.; Shen, S.; Ziebell, K.; Johnson, S.; Reid-Smith, R.; Isaac-Renton, J.; Clark, C.; Rahn, K.; Kaper, J.B. Association of genomic O island 122 of Escherichia coli EDL 933 with verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 2003, 41, 4930–4940. [Google Scholar]
- Nadler, C.; Baruch, K.; Kobi, S.; Mills, E.; Haviv, G.; Farago, M.; Alkalay, I.; Bartfeld, S.; Meyer, T.F.; Ben-Neriah, Y.; Rosenshine, I. The type III secretion effector NleE inhibits NF-kappaB activation. PLoS Pathog. 2010, 6, e1000743. [Google Scholar]
- Aktories, K.; Barbieri, J.T. Bacterial cytotoxins: Targeting eukaryotic switches. Nat. Rev. Microbiol. 2005, 3, 397–410. [Google Scholar] [CrossRef]
- Shao, F.; Merritt, P.M.; Bao, Z.; Innes, R.W.; Dixon, J.E. A Yersinia effector and a Pseudomonas avirulence protein define a family of cysteine proteases functioning in bacterial pathogenesis. Cell 2002, 109, 575–588. [Google Scholar]
- Klapproth, J.M.; Donnenberg, M.S.; Abraham, J.M.; Mobley, H.L.; James, S.P. Products of enteropathogenic Escherichia coli inhibit lymphocyte activation and lymphokine production. Infect. Immun. 1995, 63, 2248–2254. [Google Scholar]
- Klapproth, J.M.; Donnenberg, M.S.; Abraham, J.M.; James, S.P. Products of enteropathogenic E. coli inhibit lymphokine production by gastrointestinal lymphocytes. Am. J. Physiol. 1996, 271, G841–G848. [Google Scholar] [PubMed]
- Malstrom, C.; James, S. Inhibition of murine splenic and mucosal lymphocyte function by enteric bacterial products. Infect. Immun. 1998, 66, 3120–3127. [Google Scholar]
- Abu-Median, A.B.; van Diemen, P.M.; Dziva, F.; Vlisidou, I.; Wallis, T.S.; Stevens, M.P. Functional analysis of lymphostatin homologues in enterohaemorrhagic Escherichia coli. FEMS Microbiol. Lett. 2006, 258, 43–49. [Google Scholar]
- Beatty, W.L.; Belanger, T.A.; Desai, A.A.; Morrison, R.P.; Byrne, G.I. Tryptophan depletion as a mechanism of gamma interferon-mediated chlamydial persistence. Infect. Immun. 1994, 62, 3705–3711. [Google Scholar]
- Babbin, B.A.; Sasaki, M.; Gerner-Schmidt, K.W.; Nusrat, A.; Klapproth, J.M. The bacterial virulence factor lymphostatin compromises intestinal epithelial barrier function by modulating rho GTPases. Am. J. Pathol. 2009, 174, 1347–1357. [Google Scholar]
- Stevens, M.P.; van Diemen, P.M.; Frankel, G.; Phillips, A.D.; Wallis, T.S. Efa1 influences colonization of the bovine intestine by shiga toxin-producing Escherichia coli serotypes O5 and O111. Infect. Immun. 2002, 70, 5158–5166. [Google Scholar]
- Dziva, F.; van Diemen, P.M.; Stevens, M.P.; Smith, A.J.; Wallis, T.S. Identification of Escherichia coli O157:H7 genes influencing colonization of the bovine gastrointestinal tract using signature-tagged mutagenesis. Microbiology 2004, 150, 3631–3645. [Google Scholar]
- Stevens, M.P.; Roe, A.J.; Vlisidou, I.; van Diemen, P.M.; La Ragione, R.M.; Best, A.; Woodward, M.J.; Gally, D.L.; Wallis, T.S. Mutation of toxB and a truncated version of the efa-1 gene in Escherichia coli O157:H7 influences the expression and secretion of locus of enterocyte effacement-encoded proteins but not intestinal colonization in calves or sheep. Infect. Immun. 2004, 72, 5402–5411. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Klapproth, J.-M.A. The Role of Lymphostatin/EHEC Factor for Adherence-1 in the Pathogenesis of Gram Negative Infection. Toxins 2010, 2, 954-962. https://doi.org/10.3390/toxins2050954
Klapproth J-MA. The Role of Lymphostatin/EHEC Factor for Adherence-1 in the Pathogenesis of Gram Negative Infection. Toxins. 2010; 2(5):954-962. https://doi.org/10.3390/toxins2050954
Chicago/Turabian StyleKlapproth, Jan-Michael A. 2010. "The Role of Lymphostatin/EHEC Factor for Adherence-1 in the Pathogenesis of Gram Negative Infection" Toxins 2, no. 5: 954-962. https://doi.org/10.3390/toxins2050954
APA StyleKlapproth, J. -M. A. (2010). The Role of Lymphostatin/EHEC Factor for Adherence-1 in the Pathogenesis of Gram Negative Infection. Toxins, 2(5), 954-962. https://doi.org/10.3390/toxins2050954