The Effects of the Toxic Cyanobacterium Limnothrix (Strain AC0243) on Bufo marinus Larvae

Abstract
:1. Introduction
1.1. Aim of the Experiments
1.2. Animal Ethics Committee Approval
2. Results and Discussion
2.1. Water Quality Testing
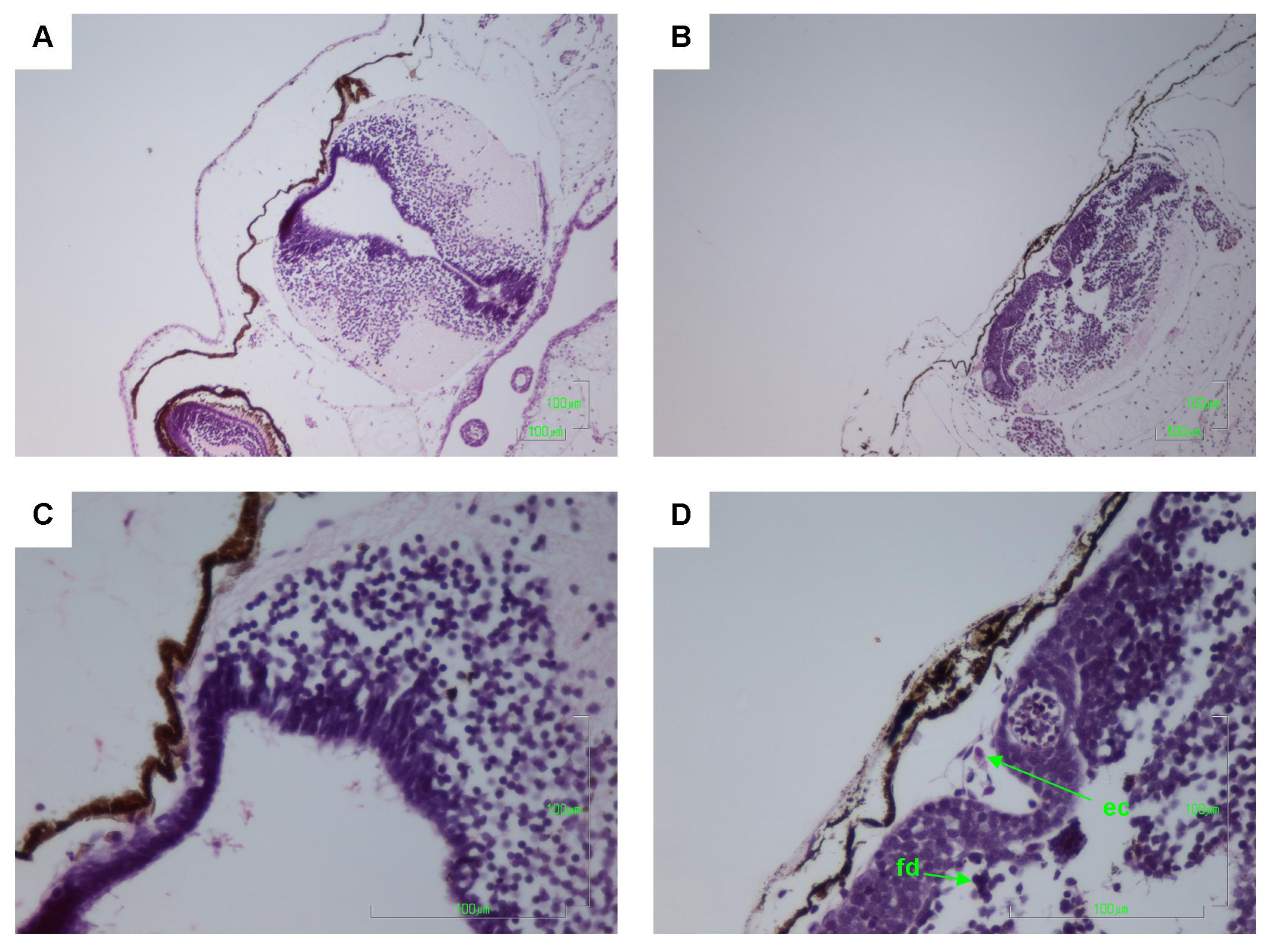
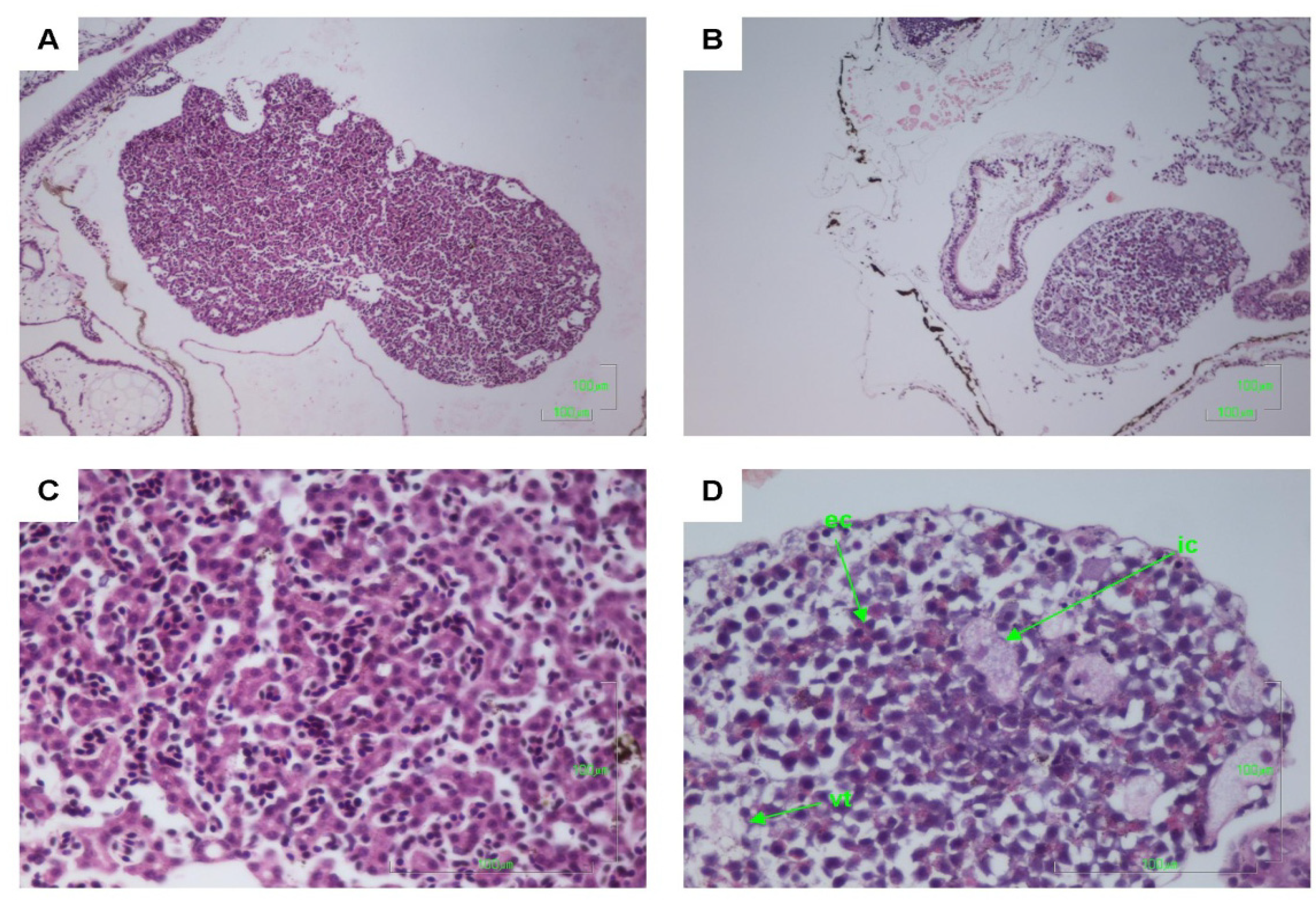
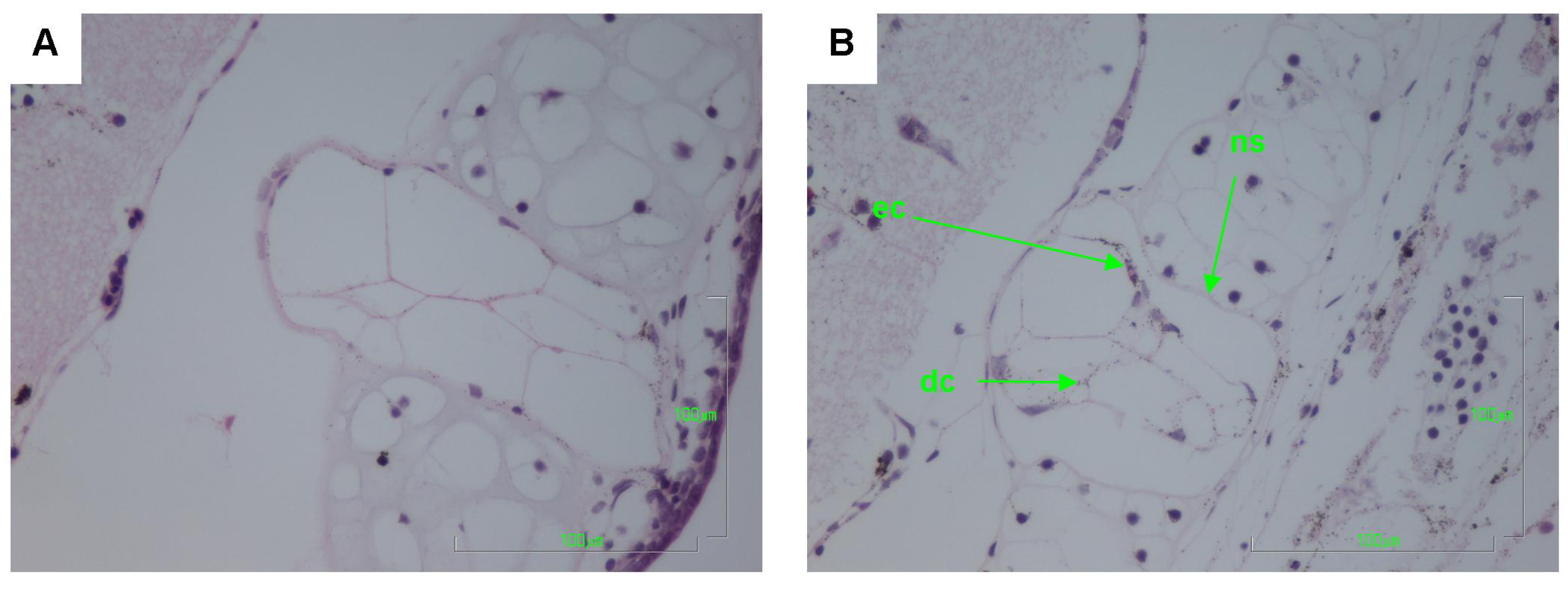
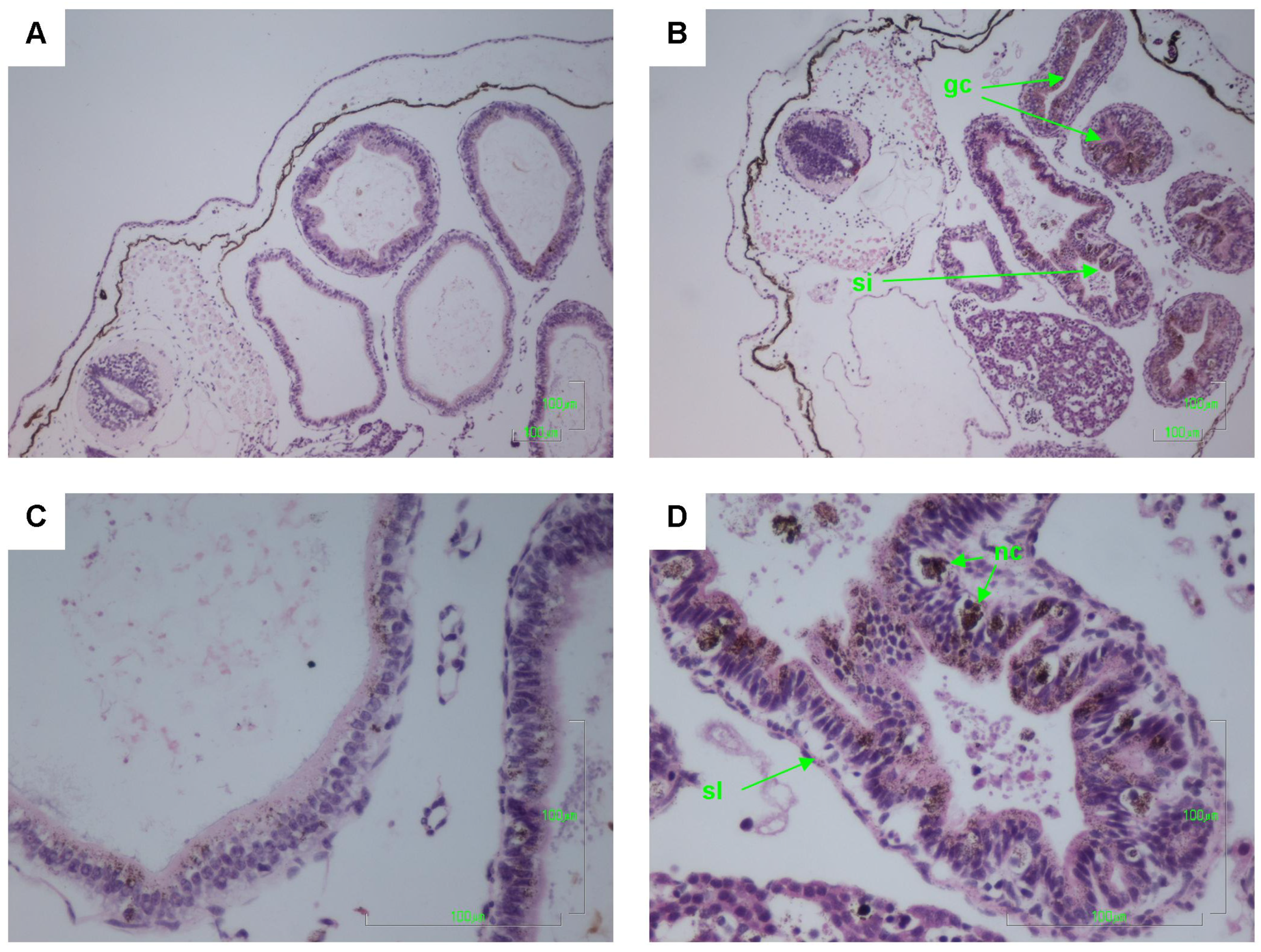
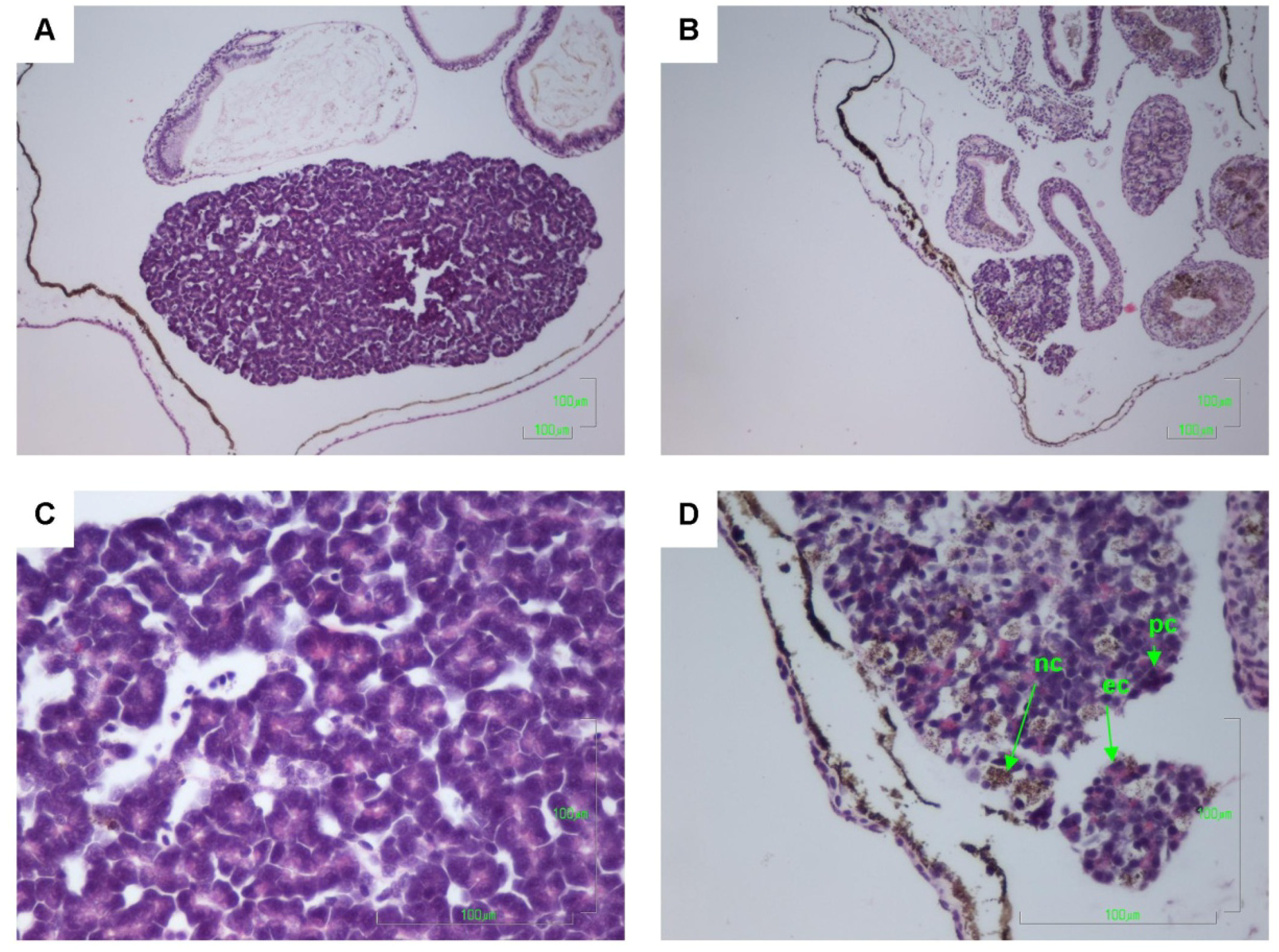
2.2. Histology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Variable | Mean | Std. deviation |
|---|---|---|---|
| Control | Dissolved oxygen (% saturation) | 33.23 | 8.27 |
| pH | 7.51 | 0.22 | |
| Water temperature (°C) | 19.75 | 0.77 | |
| Ammonia concentration (ppm) | <2 | N/A | |
| 100,000 cells mL−1 | Dissolved oxygen (% saturation) | 35.39 | 6.66 |
| pH | 7.89 | 0.27 | |
| Water temperature (°C) | 19.98 | 0.67 | |
| Ammonia concentration (ppm) | <2 | N/A |
| Body region | Pyknotic cells | Eosinophilic cells | Necrotic cells | Karyorrhexis | Inflammatory cells | Tissue vacuolation |
|---|---|---|---|---|---|---|
| Eyes | + | |||||
| Brain | + | + | + | + | + | + |
| Ventricle | + | |||||
| Gill filaments | + | + | ||||
| Kidneys | + | |||||
| Lung bud | + | |||||
| Liver | + | + | + | |||
| Pancreas | + | + | + | |||
| Gastrointestinal tract | + | |||||
| Notochord |





3. Experimental Section
3.1. Limnothrix (Strain AC0243) Culture Collection and Preparation
3.2. Larvae Collection and Test Preparation
3.3. Water Quality Testing
3.4. Euthanasia of Animals
3.5. Histology
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bernard, C.; Froscio, S.; Campbell, R.; Monis, P.; Humpage, A.; Fabbro, L. Novel toxic effects associated with a tropical Limnothrix /Geitlerinema-like cyanobacterium. Environ. Toxicol. 2011, 26, 260–270. [Google Scholar] [CrossRef]
- Humpage, A.; Falconer, I.; Bernard, C.; Froscio, S.; Fabbro, L. Toxicity of the cyanobacterium Limnothrix AC0243 to male balb/C mice. Water Res. 2012, 46, 1576–1583. [Google Scholar] [CrossRef]
- Aráoz, R.; Molgó, J.; Marsac, N.T.D. Neurotoxic cyanobacterial toxins. Toxicon 2010, 56, 813–828. [Google Scholar] [CrossRef]
- Carmichael, W. Health effects of toxin-producing cyanobacteria: “The cyanoHABs”. Hum. Ecol. Risk Assess. 2001, 7, 1393–1407. [Google Scholar] [CrossRef]
- Karlsson, O.; Lindquist, N.G.; Brittebo, E.B.; Roman, E. Selective brain uptake and behavioral effects of the cyanobacterial toxin BMAA (β-N-methylamino-l-alanine) following neonatal administration to rodents. Toxicol. Sci. 2009, 109, 286–295. [Google Scholar] [CrossRef]
- Watkins, S.; Reich, A.; Fleming, L.; Hammond, R. Neurotoxic shellfish poisoning. Mar. Drugs 2008, 6, 431–455. [Google Scholar] [CrossRef]
- World Health Organisation (WHO). Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; St Edmundsbury Press: Suffolk, UK, 1999.
- Chiu, A.S.; Gehringer, M.M.; Braidy, N.; Guillemin, G.J.; Welch, J.H.; Neilan, B.A. Excitotoxic potential of the cyanotoxin β-methyl-amino-l-alanine (BMAA) in primary human neurons. Toxicon 2012, 60, 1159–1165. [Google Scholar] [CrossRef]
- Okle, O.; Rath, L.; Galizia, C.G.; Dietrich, D.R. The cyanobacterial neurotoxin beta-N-methylamino-l-alanine (BMAA) induces neuronal and behavioral changes in honeybees. Toxicol. Appl. Pharmacol. 2013, 270, 9–15. [Google Scholar] [CrossRef]
- Brand, L.E.; Pablo, J.; Compton, A.; Hammerschlag, N.; Mash, D.C. Cyanobacterial blooms and the occurrence of the neurotoxin, beta-N-methylamino-l-alanine (BMAA), in South Florida aquatic food webs. Harmful Algae 2010, 9, 620–635. [Google Scholar] [CrossRef]
- Purdie, E.L.; Samsudin, S.; Eddy, F.B.; Codd, G.A. Effects of the cyanobacterial neurotoxin β-N-methylamino-l-alanine on the early-life stage development of zebrafish (Danio rerio). Aquat. Toxicol. 2009, 95, 279–284. [Google Scholar] [CrossRef]
- De Munck, E.; Muñoz-Sáez, E.; Miguel, B.G.; Solas, M.T.; Ojeda, I.; Martínez, A.; Gil, C.; Arahuetes, R.M. β-N-Methylamino-l-alanine causes neurological and pathological phenotypes mimicking Amyotrophic Lateral Sclerosis (ALS): The first step towards an experimental model for sporadic ALS. Environ. Toxicol. Pharmacol. 2013, 36, 243–255. [Google Scholar] [CrossRef]
- Engskog, M.K.R.; Karlsson, O.; Haglöf, J.; Elmsjö, A.; Brittebo, E.; Arvidsson, T.; Pettersson, C. The cyanobacterial amino acid β-N-methylamino-l-alanine perturbs the intermediary metabolism in neonatal rats. Toxicology 2013, 312, 6–11. [Google Scholar] [CrossRef]
- Karamyan, V.T.; Speth, R.C. Animal models of BMAA neurotoxicity: A critical review. Life Sci. 2008, 82, 233–246. [Google Scholar] [CrossRef]
- Karlsson, O.; Berg, A.L.; Lindström, A.K.; Hanrieder, J.; Arnerup, G.; Roman, E.; Bergquist, J.; Lindquist, N.G.; Brittebo, E.B.; Andersson, M. Neonatal exposure to the cyanobacterial toxin BMAA induces changes in protein expression, and neurodegeneration in adult hippocampus. Toxicol. Sci. 2012, 130, 391–404. [Google Scholar] [CrossRef]
- Karlsson, O.; Jiang, L.; Andersson, M.; Ilag, L.L.; Brittebo, E.B. Protein association of the neurotoxin and non-protein amino acid BMAA (β-N-methylamino-l-alanine) in the liver and brain following neonatal administration in rats. Toxicol. Lett. 2014, 226, 1–5. [Google Scholar] [CrossRef]
- Goto, J.J.; Koenig, J.H.; Ikeda, K. The physiological effect of ingested β-N-methylamino-l-alanine on a glutamatergic synapse in an in vivo preparation. Comp. Biochem. Physiol. Part C 2012, 156, 171–177. [Google Scholar]
- Leeuwangh, P.; Kappers, F.I.; Dekker, M.; Koerselman, W. Toxicity of cyanobacteria in Dutch lakes and reservoirs. Aquat. Toxicol. 1983, 4, 63–72. [Google Scholar] [CrossRef]
- Falconer, I.R. Effects on human health of some toxic cyanobacteria (blue-green algae) in reservoirs, lakes and rivers. Toxic. Assess. 1989, 4, 175–184. [Google Scholar] [CrossRef]
- Repavich, W.M.; Sonzogni, W.C.; Standridge, J.H.; Wedepohl, R.E.; Meisner, L.F. Cyanobacteria (blue-green algae) in Wisconsin waters: Acute and chronic toxicity. Water Res. 1990, 24, 225–231. [Google Scholar] [CrossRef]
- Carmichael, W. Cyanobacteria secondary metabolites-the cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef]
- Negri, A.; Jones, G. Bioaccumulation of Paralytic Shellfish Poisoning (PSP) toxins from the cyanobacterium Anabaena circinalis by the freshwater mussel Alathyria condola. Toxicon 1995, 33, 667–678. [Google Scholar] [CrossRef]
- Negri, A.P.; Jones, G.J.; Hindmarsh, M. Sheep mortality associated with paralytic shellfish poisons from the cyanobacterium Anabaena circinalis. Toxicon 1995, 33, 1321–1329. [Google Scholar] [CrossRef]
- Falconer, I.R. An overview of problems caused by toxic blue–green algae (cyanobacteria) in drinking and recreational water. Environ. Toxicol. 1999, 14, 5–12. [Google Scholar] [CrossRef]
- Ernst, B.; Hoeger, S.J.; O’Brien, E.; Dietrich, D.R. Oral toxicity of the microcystin-containing cyanobacterium Planktothrix rubescens in European whitefish (Coregonus lavaretus). Aquat. Toxicol. 2006, 79, 31–40. [Google Scholar] [CrossRef]
- Brand, L.E.; Campbell, L.; Bresnan, E.K. The biology and ecology of a toxic genus. Harmful Algae 2012, 14, 156–178. [Google Scholar] [CrossRef]
- Uchiyama, M.; Konno, N. Hormonal regulation of ion and water transport in anuran amphibians. Gen. Comp. Endocrinol. 2006, 147, 54–61. [Google Scholar] [CrossRef]
- Azevedo, R.A.; Carvalho, H.F.; de Brito-Gitirana, L. Hyaluronan in the epidermal and the dermal extracellular matrix: Its role in cutaneous hydric balance and integrity of anuran integument. Micron 2007, 38, 607–610. [Google Scholar] [CrossRef]
- Maejima, S.; Yamada, T.; Hamada, T.; Matsuda, K.; Uchiyama, M. Effects of hypertonic stimuli and arginine vasotocin (AVT) on water absorption response in Japanese treefrog, Hyla japonica. Gen. Comp. Endocrinol. 2008, 157, 196–202. [Google Scholar] [CrossRef]
- Suzuki, M.; Tanaka, S. Molecular and cellular regulation of water homeostasis in anuran amphibians by aquaporins. Comp. Biochem. Physiol. Part A 2009, 153, 231–241. [Google Scholar] [CrossRef]
- Maejima, S.; Konno, N.; Matsuda, K.; Uchiyama, M. Central angiotensin II stimulates cutaneous water intake behavior via an angiotensin II type-1 receptor pathway in the Japanese tree frog Hyla japonica. Horm. Behav. 2010, 58, 457–464. [Google Scholar] [CrossRef]
- White, S.H.; Duivenvoorden, L.J.; Fabbro, L.D.; Eaglesham, G.K. Mortality and toxin bioaccumulation in Bufo marinus following exposure to Cylindrospermopsis raciborskii Cell extracts and live cultures. Environ. Pollut. 2007, 147, 158–167. [Google Scholar] [CrossRef]
- Junges, C.M.; Peltzer, P.M.; Lajmanovich, R.C.; Attademo, A.M.; Cabagna Zenklusen, M.C.; Basso, A. Toxicity of the fungicide Trifloxystrobin on tadpoles and its effect on fish-tadpole interaction. Chemosphere 2012, 87, 1348–1354. [Google Scholar] [CrossRef]
- Gillespie, G.R. Impacts of sediment loads, tadpole density, and food type on the growth and development of tadpoles of the spotted tree frog Litoria spenceri: An in-stream Experiment. Biol. Conserv. 2002, 106, 141–150. [Google Scholar] [CrossRef]
- Lamberti, G.A.; Feminella, J.W.; Pringle, C.M. Methods in Stream Ecology, 2nd ed.; Academic Press: New York, NY, USA, 2007; pp. 537–559. [Google Scholar]
- Tyler, M. Frogs and Toads as Experimental Animals; The University of Adelaide: Adelade, Australia, 2009. [Google Scholar]
- Stemple, D.; Solnica-Krezel, L.; Zwartkruis, F.; Neuhauss, S.C.; Schier, A.F.; Malicki, J.; Stainier, D.Y.; Abdelilah, S.; Rangini, Z.; Mountcastle-Shah, E.; et al. Mutations affecting development of the notochord in zebrafish. Development 1996, 123, 117–128. [Google Scholar]
- Qi, B.Q.; Beasley, S.W.; Frizelle, F.A. Evidence that the notochord may be pivotal in the development of sacral and anorectal malformations. J. Pediatr. Surg. 2003, 38, 1310–1316. [Google Scholar] [CrossRef]
- Platz, F. Structural and experimental investigations of the functional anatomy and the turgor of the notochord in the larval tail of anuran tadpoles. Ann. Anat. 2006, 188, 289–302. [Google Scholar] [CrossRef]
- Pagnon-Minot, A.; Malbouyres, M.; Haftek-Terreau, Z.; Kim, H.R.; Sasaki, T.; Thisse, C.; Thisse, B.; Ingham, P.W.; Ruggiero, F.; Le Guellec, D.; et al. A novel factor in zebrafish notochord differentiation and muscle development. Dev. Biol. 2008, 316, 21–35. [Google Scholar] [CrossRef]
- Ribes, V.; Balaskas, N.; Sasai, N.; Cruz, C.; Dessaud, E.; Cayuso, J.; Tozer, S.; Yang, L.L.; Novitch, B.; Marti, E.; et al. Distinct sonic hedgehog signaling dynamics specify floor plate and ventral neuronal progenitors in the vertevrate neural tube. Genes Dev. 2010, 24, 1186–1200. [Google Scholar] [CrossRef] [Green Version]
- Kinnear, S. Cylindrospermopsin in whole cell extracts and live cultures of Cylindrospermopsis raciborskii: Ecotoxicity, bioaccumulation and management. PhD thesis, School of Biological and Environmental Sciences, Central Queensland University, Rockhampton, QLD, Australia, July 2006. [Google Scholar]
- Ziková, A.; Lorenz, C.; Lutz, I.; Pflugmacher, S.; Kloas, W. Physiological responses of Xenopus laevis tadpoles exposed to cyanobacterial biomass containing microcystin-LR. Aquat. Toxicol. 2013, 128–129, 25–33. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Australian Code of Practice for the Care and Use of Animals for Scientific Purposes; Australian Government: Canberra, Australia, 2004.
- Gao, Y.; Cornwell, J.C.; Stoecker, D.K.; Owens, M.S. Effects of cyanobacterial-driven pH increases on sediment nutrient fluxes and coupled nitrification-denitrification in a shallow fresh water estuary. Biogeosciences 2012, 9, 2697–2710. [Google Scholar] [CrossRef]
- Marques, S.M.; Chaves, S.; Gonçalves, F.; Pereira, R. Evaluation of growth, biochemical and bioaccumulation parameters in Pelophylax perezi tadpoles, following an in-situ acute exposure to three different effluent ponds from a uranium mine. Sci. Total Environ. 2013, 445–446, 321–328. [Google Scholar] [CrossRef]
- ANZECC. Australian and New Zealand Guidelines for Fresh and Marine Water Quality; Australian and New Zealand Environment and Conservation Council: Canberra, Australia, 2000.
- Seigel, R.; Dismore, A.; Richter, S. Using well water to increase hydroperiod as a management option for pond-breeding amphibians. Wildl. Soc. Bull. 2006, 34, 1022–1027. [Google Scholar] [CrossRef]
- Economic Development and Inovation Department of Employment; Markula, A.; Csurhes, S.; Hannan-Jones, M. (Eds.) Biosecurity Queensland: Brisbane, Australia, 2010.
- Faassen, E.J.; Harkema, L.; Begeman, L.; Lurling, M. First report of (Homo)Anatoxin-a and dog neurotoxicosis after ingestion of benthic cyanobacteria in the Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar]
- Ioudina, M.; Uemura, E.; Greenlee, H.W. Glucose insufficiency alters neuronal viability and increases susceptibility to glutamate toxicity. Brain Res. 2004, 1004, 188–192. [Google Scholar]
- Berry, J.P.; Gantar, M.; Gibbs, P.D.L.; Schmale, M.C. The zebrafish (Danio rerio) embryo as a model system for identification and characterization of developmental toxins from marine and freshwater microalgae. Comp. Biochem. Physiol. Part C 2007, 145, 61–72. [Google Scholar]
- Stewart, I. Environmental risk factors for temporal lobe epilepsy—Is prenatal exposure to the marine algal neurotoxin domoic acid a potentially preventable cause? Med. Hypotheses 2010, 74, 466–481. [Google Scholar] [CrossRef]
- Karlsson, O.; Berg, C.; Brittebo, E.B.; Lindquist, N.G. Retention of the cyanobacterial neurotoxin β-N-Methylamino-l-Alanine in melanin and neuromelanin-containing cells—A possible link between Parkinson-dementia complex and pigmentary retinopathy. Pigment Cell Melanoma Res. 2009, 22, 120–130. [Google Scholar] [CrossRef]
- Müller, F.; Albert, S.; Blader, P.; Fischer, N.; Hallonet, M.; Strähle, U. Direct actin of the nodal-related signal cyclops in induction of sonic hedgehog in the ventral midline of the CNS. Development 2000, 127, 3889–3897. [Google Scholar]
- Lek, M.; Dias, J.M.; Marklund, U.; Uhde, C.W.; Kurdija, S.; Lei, Q.; Sussel, L.; Rubenstein, J.L.; Matise, M.P.; Arnold, H.H.; et al. A homeodomain feedback circuit underlies step-function interpretation of a Shh morphogen gradient during ventral neural patterning. Development 2010, 137, 4051–4060. [Google Scholar] [CrossRef]
- Qi, B.Q.; Beasley, S.W. Relationship of the notochord to foregut development in the fetal rat model of esophageal atresia. J. Pediatr. Surg. 1999, 34, 1593–1598. [Google Scholar] [CrossRef]
- McDairmid, R.; Altig, R. Tadpoles: The Biology of Anuran Larvae; The University of Chicago Press: Chicago, IL, USA, 1999; p. 436. [Google Scholar]
- Kankaanpää, H.T.; Holliday, J.; Schröder, H.; Goddard, T.J.; von Fister, R.; Carmichael, W.W. Cyanobacteria and prawn farming in Northern New South Wales, Australia—A case study on cyanobacteria diversity and hepatotoxin bioaccumulation. Toxicol. Appl. Pharmacol. 2005, 203, 243–256. [Google Scholar] [CrossRef]
- Quiblier, C.; Susanna, W.; Isidora, E.S.; Mark, H.; Aurélie, V.; Jean-François, H. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Handeland, K.; Østensvik, Ø. Microcystin poisoning in roe deer (Capreolus capreolus). Toxicon 2010, 56, 1076–1078. [Google Scholar] [CrossRef]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Contribution of hot spring cyanobacteria to the mysterious deaths of lesser flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef]
- Lopez Rodas, V.; Costas, E. Preference of mice to consume Microcystis aeruginosa (toxin-producing cyanobacteria): A possible explanation for numerous fatalities of livestock and wildlife. Res. Vet. Sci. 1999, 67, 107–110. [Google Scholar] [CrossRef]
- Soll, M.; Williams, M. Mortality of white rhinoceros (Ceratotherium simum) Suspected to be associated with the blue-green alga (Microcystis aeruginosa). J. S. Afr. Vet. Assoc. 1985, 56, 49–51. [Google Scholar]
- Stewart, I.; Seawright, A.A.; Shaw, G.R. Advances in Medicine and Biology; Hudnell, H., Ed.; Springer Science + Business Media, LLC: New York, NY, USA, 2008; Volume 619, Chapter 28; pp. 613–637. [Google Scholar]
- Jara, F.G.; Perotti, M.G. Risk of predation and behavioural response in three anuran species: Influence of tadpole size and predator type. Hydrobiologia 2010, 644, 313–324. [Google Scholar] [CrossRef]
- Kiesecker, J.; Chivers, D.P.; Marco, A.; Quilchano, C.; Anderson, M.T.; Blaustein, A.R. Identification of a disturbance signal in larval red-legged frogs, Rana aurora. Anim. Behav. 1999, 57, 1295–1300. [Google Scholar] [CrossRef]
- Mirza, R.S.; Ferrari, M.C.O.; Kiesecker, J.M.; Chivers, D.P. Responses of american toad tadpoles to predation cues: Behavioural response thresholds, threat-sensitivity and acquired predation recognition. Behaviour 2006, 143, 877–889. [Google Scholar] [CrossRef]
- Gorham, P.; Lachlan, J.M.; Hammer, U.; Kim, W. Isolation and culture of toxic strains of Anabaena flos-aquae (Lyngb) De Breb. Verh. Int. Ver. fuer Theor. und Angew. Limnol. 1964, 15, 796–804. [Google Scholar]
- Porter, K. Herpetology; W.B. Saunders Company: Philadelphia, PA, USA, 1972. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Daniels, O.; Fabbro, L.; Makiela, S. The Effects of the Toxic Cyanobacterium Limnothrix (Strain AC0243) on Bufo marinus Larvae. Toxins 2014, 6, 1021-1035. https://doi.org/10.3390/toxins6031021
Daniels O, Fabbro L, Makiela S. The Effects of the Toxic Cyanobacterium Limnothrix (Strain AC0243) on Bufo marinus Larvae. Toxins. 2014; 6(3):1021-1035. https://doi.org/10.3390/toxins6031021
Chicago/Turabian StyleDaniels, Olivia, Larelle Fabbro, and Sandrine Makiela. 2014. "The Effects of the Toxic Cyanobacterium Limnothrix (Strain AC0243) on Bufo marinus Larvae" Toxins 6, no. 3: 1021-1035. https://doi.org/10.3390/toxins6031021
APA StyleDaniels, O., Fabbro, L., & Makiela, S. (2014). The Effects of the Toxic Cyanobacterium Limnothrix (Strain AC0243) on Bufo marinus Larvae. Toxins, 6(3), 1021-1035. https://doi.org/10.3390/toxins6031021