
Links between Genetic Groups, Indole Alkaloid Profiles and Ecology within the Grass-Parasitic Claviceps purpurea Species Complex

Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Location a | Plot | Latitude/longitude | Habitat b | Host c (# sclerotia) | Genetic groups d ( data for sclerotia), & co-occurrence * |
|---|---|---|---|---|---|---|
| 2011 | 1 | 1 | 58.06502/6.78550 | Saline | Leymus arenarius (4) | |
| 2 | 58.06642/6.79428 | Saline | Leymus arenarius (5) | * | ||
| 3 | 58.06999/6.72743 | Saline | Calamagrostis epigejos (20) | |||
| 2012 | 4 | 58.07044/6.72735 | Saline | Leymus arenarius (15) | * | |
| 2011 | 2 | 1 | 58.74609/5.51021 | Saline | Ammophila arenaria (13) | * |
| 2 | 58.88869/5.60333 | Saline | Ammophila arenaria (10) | |||
| 2011 | 3 | 1 | 59.97072/10.74814 | Very wet | Molinia caerulea (8) | |
| 2 | 59.97174/10.7477 | Very wet | Molinia caerulea (20) | |||
| 3 | 59.97301/10.74723 | Very wet | Molinia caerulea (15) | |||
| 4 | 59.96967/10.74828 | Wet | Phleum pratense (5) | |||
| 2012 | 5 | 59.96972/10.74832 | Wet | Calamagrostis arundinacea (15) | ||
| 5 | 59.96972/10.74832 | Wet | Deschampsia cespitosa (11) | |||
| 6 | 59.96887/10.74753 | Wet | Elymus repens (6) | |||
| 7 | 59.96828/10.74738 | Wet | Phleum pratense (10) | |||
| 8 | 59.96832/10.7481 | Wet | Schedonorus pratensis (14) | |||
| 2011 | 4 | 1 | 59.97219/10.76052 | Dry | Phleum pratense (12) | |
| 2 | 59.97263/10.76325 | Dry | Phleum pratense (16) | |||
| 3 | 59.97296/10.76408 | Dry | Dactylis glomerata (28) | * | ||
| 3 | 59.97296/10.76408 | Dry | Elymus repens (8) | |||
| 3 | 59.97296/10.76408 | Dry | Schedonorus pratensis (13) | |||
| 2012 | 4 | 59.9723/10.75967 | Dry | Calamagrostis arundinacea (20) | ||
| 5 | 59.97898/10.75447 | Very wet | Molinia caerulea (20) | |||
| 2012 | 5 | 1 | 58.97152/9.26459 | Dry | Anthoxanthum odoratum (15) | |
| 1 | 58.97152/9.26459 | Dry | Dactylis glomerata (20) | |||
| 1 | 58.97152/9.26459 | Dry | Festuca rubra (20) | |||
| 1 | 58.97152/9.26459 | Dry | Phalaris arundinacea (15) | |||
| 1 | 58.97152/9.26459 | Dry | Poa pratensis (15) | |||
| 2 | 58.97147/9.26508 | Very wet | Molinia caerulea (20) | |||
| 3 | 58.9682/9.25837 | Dry | Calamagrostis arundinacea (29) | |||
| 3 | 58.9682/9.25837 | Dry | Calamagrostis epigejos (34) | |||
| 3 | 58.9682/9.25837 | Dry | Phleum pratense (31) | * | ||
| 2011 | 6 | 1 | 59.9838/10.66953 | Dry | Phleum pratense (40) | |
| 2 | 59.98401/10.66767 | Dry | Phleum pratense (36) | |||
| 2012 | 3 | 59.98268/10.67023 | Dry | Deschampsia cespitosa (8) | ||
| 3 | 59.98268/10.67023 | Dry | Festuca rubra (10) | |||
| 4 | 59.97965/10.66966 | Dry | Calamagrostis arundinacea (15) |
2. Results
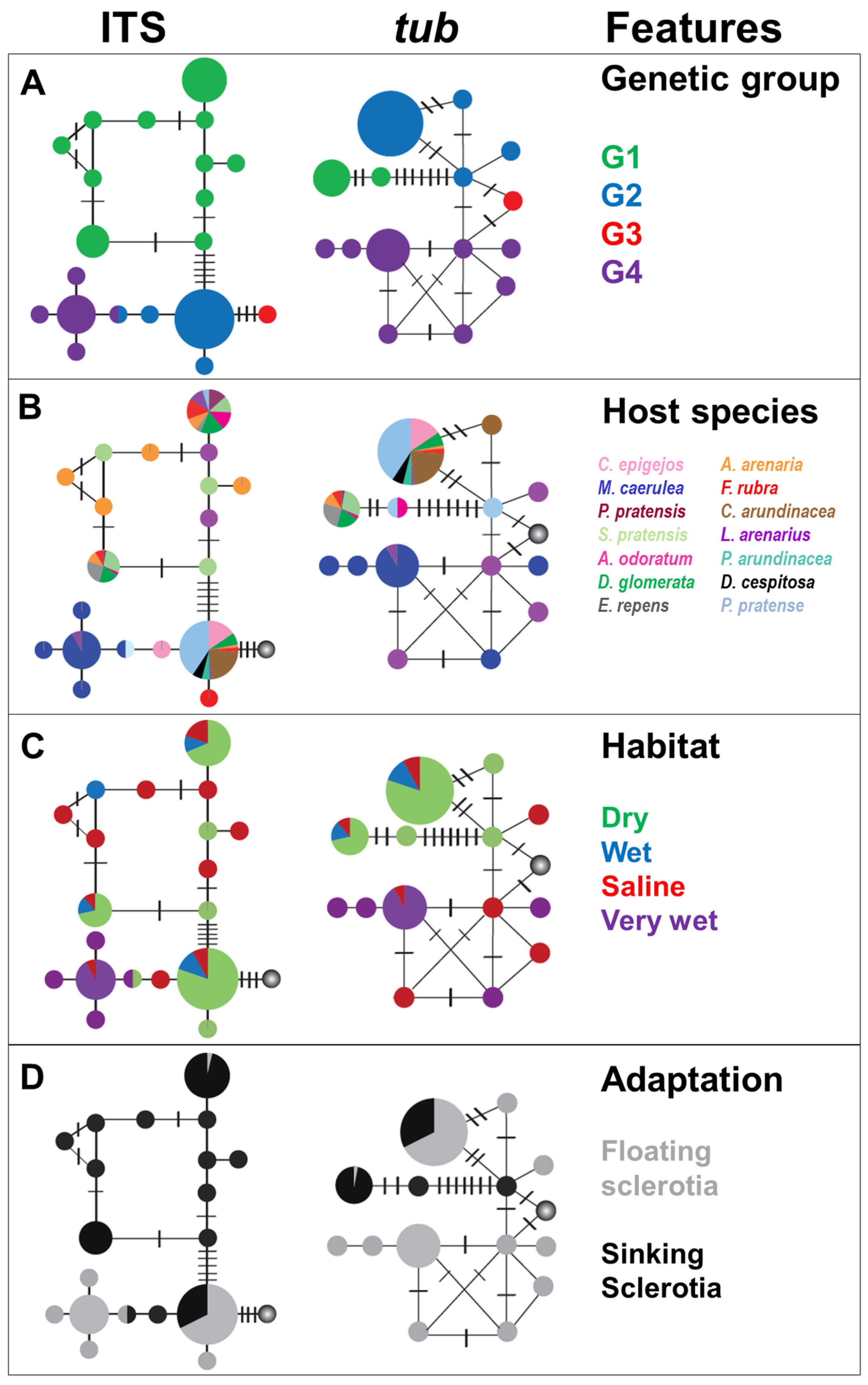
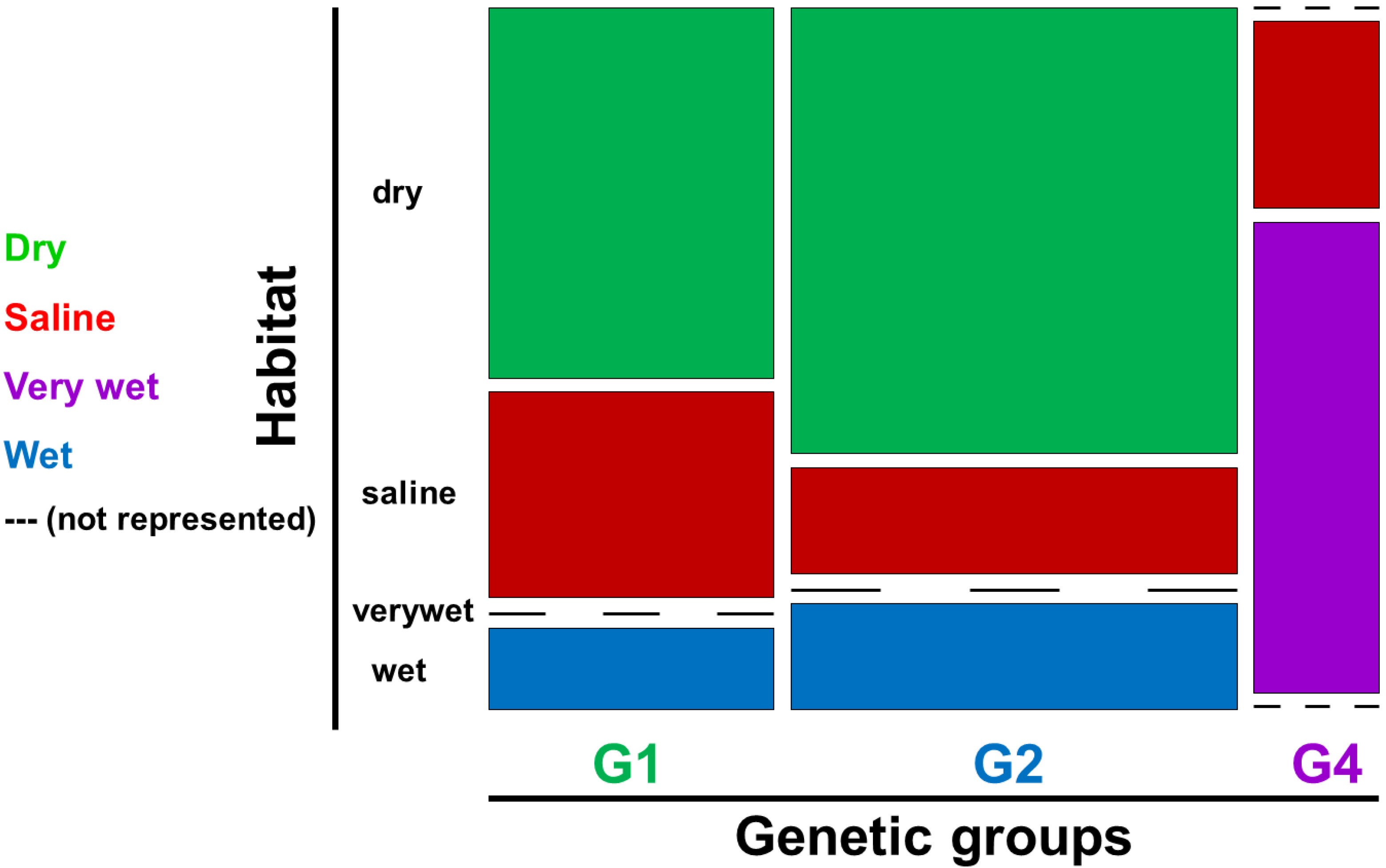
2.1. Genetic Groups



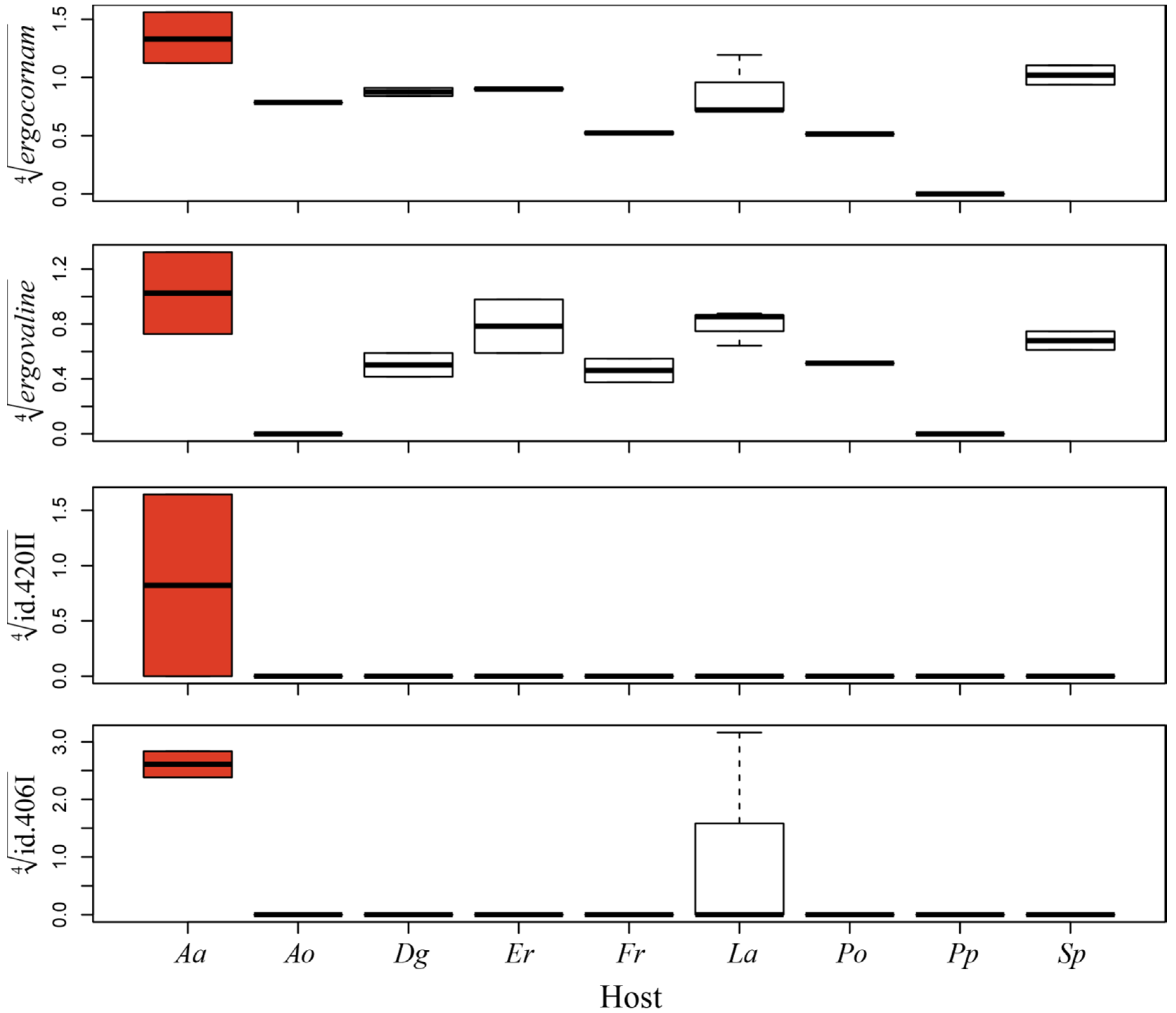
2.2. Indole Alkaloid Profiles
| Explanatory variable | Df | F.Model | R2 | Pr(>F) |
|---|---|---|---|---|
| Genetic group | 2 | 22.369 | 0.49854 | 0.001 |
| Host genus | 11 | 2.8885 | 0.46882 | 0.001 |
| Habitat | 3 | 2.4141 | 0.14134 | 0.013 |

2.3. Ecological Characteristics


3. Discussion
3.1. Genetic Groups and Speciation
3.2. Alkaloid Profiles and Speciation
3.3 Ecology and Speciation
4. Materials and Methods
4.1. Sampling and Preparation of Sclerotia
4.2. Molecular Analyses
4.3. PCR and Sequencing
4.4. Alkaloid Standards
4.5. Alkaloid Analyses
4.6. Bioinformatics and Statistical Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lorenz, N.; Haarmann, T.; Pažoutová, S.; Jung, M.; Tudzynski, P. The ergot alkaloid gene cluster: Functional analyses and evolutionary aspects. Phytochemistry 2009, 70, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Schiff, P.L. Ergot and its alkaloids. Am. J. Pharm. Educ. 2006, 70, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Komarova, E.; Tolkachev, O. The chemistry of peptide ergot alkaloids. Part 1. Classification and chemistry of ergot peptides. Pharm. Chem. J. 2001, 35, 504–513. [Google Scholar] [CrossRef]
- Schardl, C.L.; Young, C.A.; Hesse, U.; Amyotte, S.G.; Andreeva, K.; Calie, P.J.; Fleetwood, D.J.; Haws, D.C.; Moore, N.; Oeser, B.; et al. Plant-symbiotic fungi as chemical engineers: Multi-genome analysis of the Clavicipitaceae reveals dynamics of alkaloid loci. PLoS Genet 2013, 9, e1003323. [Google Scholar] [CrossRef] [PubMed]
- Panaccione, D.G. Origins and significance of ergot alkaloid diversity in fungi. FEMS Microbiol. Lett. 2005, 251, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Petersen, D.; Rolen, E.; Egge-Jacobsen, W.; Vrålstad, T. Ergosedmine, a new peptide ergot alkaloid (ergopeptine) from the ergot fungus, Claviceps purpurea parasitizing Calamagrostis arundinacea. Phytochem. Lett. 2011, 4, 79–85. [Google Scholar] [CrossRef]
- Schardl, C.L.; Panaccione, D.G.; Tudzynski, P. Ergot alkaloids—Biology and molecular biology. In The alkaloids: Chemistry and biology; Geoffrey, A.C., Ed.; Academic Press: San Diego, CA, USA, 2006; Volume 63, pp. 45–86. [Google Scholar]
- Hulvová, H.; Galuszka, P.; Frébortová, J.; Frébort, I. Parasitic fungus Claviceps as a source for biotechnological production of ergot alkaloids. Biotechnol. Adv. 2013, 31, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Botha, C.J.; Kellerman, T.S.; Fourie, N. A tremorgenic mycotoxicosis in cattle caused by Paspalum distichum (I.) infected by Claviceps paspali. J. S. Afr. Vet. Assoc. 1996, 67, 36–37. [Google Scholar] [PubMed]
- Botha, C.J.; Naude, T.W.; Moroe, M.L.; Rottinghaus, G.E. Gangrenous ergotism in cattle grazing fescue (Festuca elatior l.) in south africa. J. S. Afr. Vet. Assoc. 2004, 75, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Løken, T.; Ulvund, M.J.; Våg, T. A case of gangrenous ergotism in grazing sheep. Acta Vet. Scand. 1979, 20, 601–603. [Google Scholar] [PubMed]
- Handeland, K.; Vikøren, T. Presumptive gangrenous ergotism in free-living moose and a roe deer. J. Wildl. Dis. 2005, 41, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Flieger, M.; Wurst, M.; Shelby, R. Ergot alkaloids—Sources, structures and analytical methods. Folia Microbiol. 1997, 42, 3–30. [Google Scholar] [CrossRef]
- Panaccione, D.G.; Beaulieu, W.T.; Cook, D. Bioactive alkaloids in vertically transmitted fungal endophytes. Funct. Ecol. 2014, 28, 299–314. [Google Scholar] [CrossRef]
- Steiner, U.; Leistner, E. Ergoline alkaloids in convolvulaceous host plants originate from epibiotic clavicipitaceous fungi of the genus Periglandula. Fungal Ecol. 2012, 5, 316–321. [Google Scholar] [CrossRef]
- Uhlig, S.; Botha, C.J.; Vrålstad, T.; Rolen, E.; Miles, C.O. Indole-diterpenes and ergot alkaloids in Cynodon dactylon (Bermuda grass) infected with Claviceps cynodontis from an outbreak of tremors in cattle. J. Agric. Food Chem. 2009, 57, 11112–11119. [Google Scholar] [CrossRef] [PubMed]
- Cawdell-Smith, A.J.; Scrivener, C.J.; Bryden, W.L. Staggers in horses grazing paspalum infected with Claviceps paspali. Aust. Vet. J. 2010, 88, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Moyano, M.R.; Molina, A.M.; Lora, A.J.; Mendez, J.; Rueda, A. Tremorgenic mycotoxicosis caused by Paspalum paspaloides (michx.) scribner infected by Claviceps paspali: A case report. Vet. Med. 2010, 55, 336–338. [Google Scholar]
- Knaus, H.-G.; McManus, O.B.; Lee, S.H.; Schmalhofer, W.A.; Garcia-Calvo, M.; Helms, L.M.H.; Sanchez, M.; Giangiacomo, K.; Reuben, J.P. Tremorgenic indole alkaloids potently inhibit smooth muscle high-conductance calcium-activated potassium channels. Biochemistry 1994, 33, 5819–5828. [Google Scholar] [CrossRef]
- Uhlig, S.; Egge-Jacobsen, W.; Vrålstad, T.; Miles, C.O. Indole–diterpenoid profiles of Claviceps paspali and Claviceps purpurea from high-resolution fourier transform orbitrap mass spectrometry. Rapid Commun. Mass Spectrom. 2014, 28, 1621–1634. [Google Scholar] [CrossRef] [PubMed]
- Pažoutová, S.; Olsovska, J.; Linka, M.; Kolinska, R.; Flieger, M. Chemoraces and habitat specialization of Claviceps purpurea populations. Appl. Environ. Microbiol. 2000, 66, 5419–5425. [Google Scholar] [CrossRef] [PubMed]
- Douhan, G.W.; Smith, M.E.; Huyrn, K.L.; Westbrook, A.; Beerli, P.; Fisher, A.J. Multigene analysis suggests ecological speciation in the fungal pathogen Claviceps purpurea. Mycol. Res. 2008, 17, 2276–2286. [Google Scholar]
- Pažoutová, S.; Raybould, A.F.; Honzatko, A.; Kolinska, R. Specialised population of Claviceps purpurea from salt marsh Spartina species. Mycol. Res. 2002, 106, 210–214. [Google Scholar] [CrossRef]
- Fisher, A.J.; Gordon, T.R.; Ditomaso, J.M. Geographic distribution and diversity in Claviceps purpurea from salt marsh habitats and characterization of pacific coast populations. Mycol. Res. 2005, 109, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.J.; DiTomaso, J.M.; Gordon, T.R. Intraspecific groups of Claviceps purpurea associated with grass species in willapa bay, washington, and the prospects for biological control of invasive Spartina alterniflora. Biol. Control 2005, 34, 170–179. [Google Scholar] [CrossRef]
- Nehring, S.; Boestfleisch, C.; Buhmann, A.; Papenbrock, J. The north american toxic fungal pathogen g3 Claviceps purpurea (Fries) Tulasne is established in the german wadden sea. Biol. Invasion Rec. 2012, 1, 5–10. [Google Scholar]
- Pazoutova, S.; Kolinska, R.; Honzatko, A.; Cagas, B. Host specialization of different populations of ergot fungus (Claviceps purpurea). Czech. J. Genet. Plant Breed. UZPI 2002, 38, 75–81. [Google Scholar]
- Pažoutová, S.; Pešicová, K.; Chudíčková, M.; Šrůtka, P.; Kolařík, M. Delimitation of cryptic species inside Claviceps purpurea. Fungal Biol. 2015, 119, 7–26. [Google Scholar] [CrossRef] [PubMed]
- Young, C.A.; Schardl, C.L.; Panaccione, D.G.; Florea, S.; Takach, J.E.; Charlton, N.D.; Moore, N.; Webb, J.S.; Jaromczyk, J. Genetics, genomics and evolution of ergot alkaloid diversity. Toxins 2015, 7, 1273–1302. [Google Scholar] [CrossRef] [PubMed]
- Gerhards, N.; Neubauer, L.; Tudzynski, P.; Li, S.-M. Biosynthetic pathways of ergot alkaloids. Toxins 2014, 6, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Havemann, J.; Vogel, D.; Loll, B.; Keller, U. Cyclolization of D-lysergic acid alkaloid peptides. Chem. Biol. 2014, 21, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Haarmann, T.; Machado, C.; Lübbe, Y.; Correia, T.; Schardl, C.L.; Panaccione, D.G.; Tudzynski, P. The ergot alkaloid gene cluster in Claviceps purpurea: Extension of the cluster sequence and intra species evolution. Phytochemistry 2005, 66, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Haarmann, T.; Lorenz, N.; Tudzynski, P. Use of a nonhomologous end joining deficient strain (∆ku70) of the ergot fungus Claviceps purpurea for identification of a nonribosomal peptide synthetase gene involved in ergotamine biosynthesis. Fungal Genet. Biol. 2008, 45, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Imlach, W.L.; Finch, S.C.; Dunlop, J.; Meredith, A.L.; Aldrich, R.W.; Dalziel, J.E. The molecular mechanism of “ryegrass staggers,” a neurological disorder of K+ channels. J. Pharmacol. Exp. Ther. 2008, 327, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.; Nicholson, M.J.; Young, C.; Parker, E.J.; Scott, B. The genetic basis for indole-diterpene chemical diversity in filamentous fungi. Mycol. Res. 2008, 112, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R.A.; Sullivan, R.; Alderman, S.C.; Spatafora, J.W.; White, J.F. Claviceps purpurea var. Spartinae var nov.: An ergot adapted to the aquatic environment. Mycotaxon 2002, 81, 11–25. [Google Scholar]
- Vialle, A.; Feau, N.; Frey, P.; Bernier, L.; Hamelin, R.C. Phylogenetic species recognition reveals host-specific lineages among poplar rust fungi. Mol. Phylogenetics Evolut. 2013, 66, 628–644. [Google Scholar] [CrossRef]
- Wäli, P.P.; Wäli, P.R.; Saikkonen, K.; Tuomi, J. Is the pathogenic ergot fungus a conditional defensive mutualist for its host grass? PLoS One 2013, 8, e69249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, G.A.; Christensen, M.J.; Miles, C.O. Coevolution of fungal endophytes with grasses: The significance of secondary metabolites. In Microbial Endophytes; Bacon, C.W., White, J.F.J., Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 341–388. [Google Scholar]
- Clapham, A.R.; Tutin, T.G.; Warburg, E.F. Flora of the british isles., 2nd ed.; University Press: Cambridge, UK, 1962. [Google Scholar]
- Achenbach, L; Brix, H. Can differences in salinity tolerance explain the distribution of four genetically distinct lineages of Phragmites australis in the Mississippi River Delta? Hydrobiologia 2014, 737, 5–23. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vrålstad, T.; Knutsen, A.K.; Tengs, T.; Holst-Jensen, A. A quantitative taqman® MGB real-time polymerase chain reaction based assay for detection of the causative agent of crayfish plague Aphanomyces astaci. Vet. Microbiol. 2009, 137, 146–155. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR protocols: A guide to methods and applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Annis, S.L.; Panaccione, D.G. Presence of peptide synthetase gene transcripts and accumulation of ergopeptines in Claviceps purpurea and Neotyphodium coenophialum. Can. J. Microbiol. 1998, 44, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Petersen, D. Lactam ergot alkaloids (ergopeptams) as predominant alkaloids in sclerotia of Claviceps purpurea from norwegian wild grasses. Toxicon 2008, 52, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/nt. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evolut. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular evolution and phylogenetics; Oxford University Press: Oxford, NY, USA, 2000; p. 126. [Google Scholar]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under linux and windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A language and environment for statistical computing; 3.0.2; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Oksanen, J.; Blanchet, G.F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O'Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community ecology package. Available online: http://CRAN.R-project.org/package=vegan (accessed on 17 October 2013).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Aust. Ecol. 2001, 26, 32–46. [Google Scholar]
- Legendre, P.; Gallagher, E. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Negård, M.; Uhlig, S.; Kauserud, H.; Andersen, T.; Høiland, K.; Vrålstad, T. Links between Genetic Groups, Indole Alkaloid Profiles and Ecology within the Grass-Parasitic Claviceps purpurea Species Complex. Toxins 2015, 7, 1431-1456. https://doi.org/10.3390/toxins7051431
Negård M, Uhlig S, Kauserud H, Andersen T, Høiland K, Vrålstad T. Links between Genetic Groups, Indole Alkaloid Profiles and Ecology within the Grass-Parasitic Claviceps purpurea Species Complex. Toxins. 2015; 7(5):1431-1456. https://doi.org/10.3390/toxins7051431
Chicago/Turabian StyleNegård, Mariell, Silvio Uhlig, Håvard Kauserud, Tom Andersen, Klaus Høiland, and Trude Vrålstad. 2015. "Links between Genetic Groups, Indole Alkaloid Profiles and Ecology within the Grass-Parasitic Claviceps purpurea Species Complex" Toxins 7, no. 5: 1431-1456. https://doi.org/10.3390/toxins7051431
APA StyleNegård, M., Uhlig, S., Kauserud, H., Andersen, T., Høiland, K., & Vrålstad, T. (2015). Links between Genetic Groups, Indole Alkaloid Profiles and Ecology within the Grass-Parasitic Claviceps purpurea Species Complex. Toxins, 7(5), 1431-1456. https://doi.org/10.3390/toxins7051431