
Sphingomyelinase D/Ceramide 1-Phosphate in Cell Survival and Inflammation


{kind=link}
Abstract
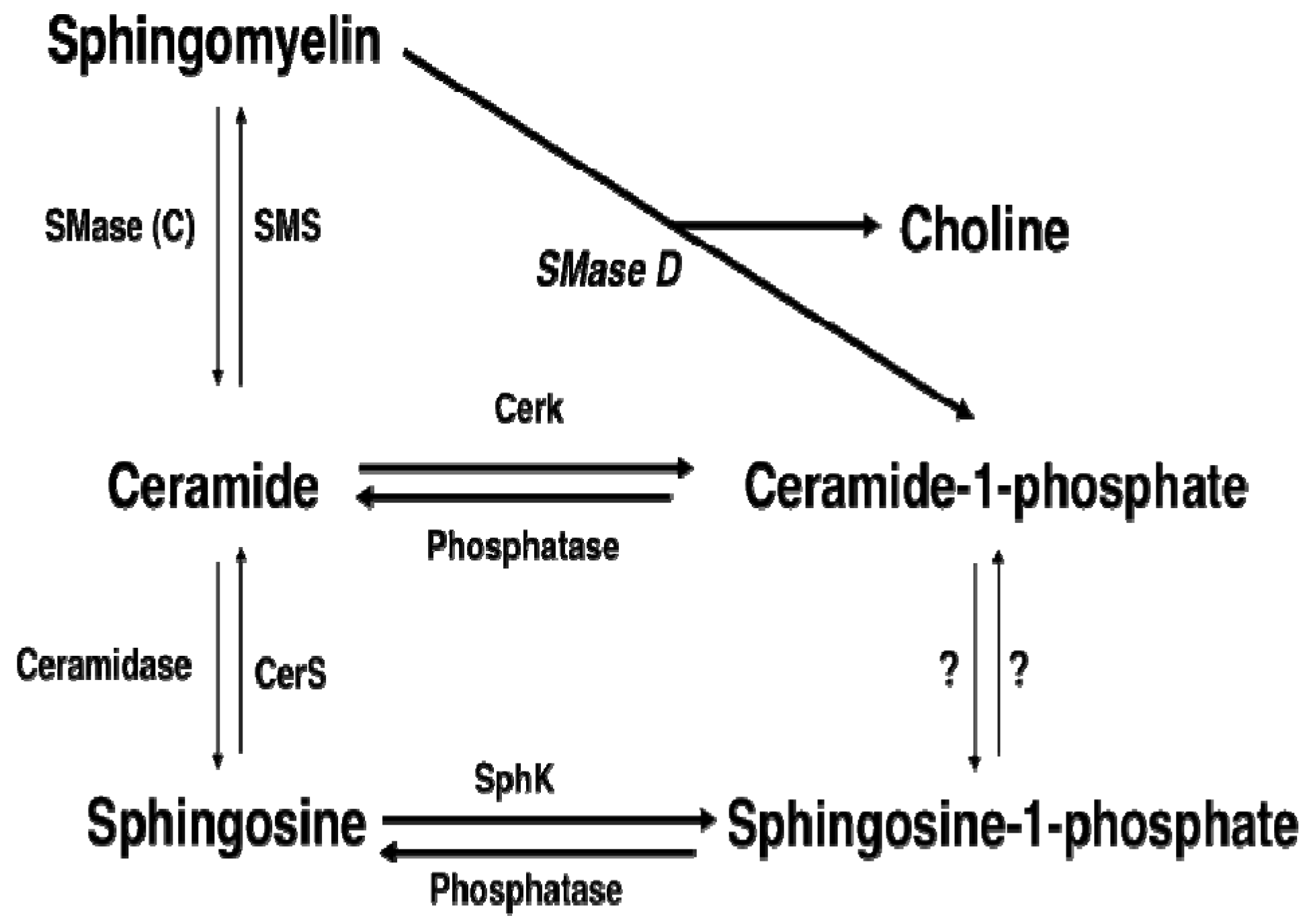
:1. Introduction

2. Biological Activities of C1P
2.1. C1P Promotes Cell Proliferation and Survival
2.2. Implication of C1P in Inflammatory Responses
2.3. Ceramide 1-Phosphate Promotes Cell Migration
3. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Zobel-Thropp, P.A.; Kerins, A.E.; Binford, G.J. Sphingomyelinase D in Sicariid spider venom is a potent insecticidal toxin. Toxicon 2012, 60, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.; Miller, M.J.; Gomez, H.F.; Warren, J.S. Loxosceles deserta spider venom induces NF-kappaB-dependent chemokine production by endothelial cells. J. Toxicol. Clin. Toxicol. 1999, 37, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Binford, G.J.; Cordes, M.H.; Wells, M.A. Sphingomyelinase D from venoms of Loxosceles spiders: Evolutionary insights from cDNA sequences and gene structure. Toxicon 2005, 45, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Gangoiti, P.; Granado, M.H.; Arana, L.; Ouro, A.; Gomez-Munoz, A. Activation of protein kinase C-alpha is essential for stimulation of cell proliferation by ceramide 1-phosphate. FEBS Lett. 2010, 584, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Munoz, A.; Duffy, P.A.; Martin, A.; O’Brien, L.; Byun, H.S.; Bittman, R.; Brindley, D.N. Short-chain ceramide-1-phosphates are novel stimulators of DNA synthesis and cell division: antagonism by cell-permeable ceramides. Mol. Pharmacol. 1995, 47, 833–839. [Google Scholar] [PubMed]
- Gomez-Munoz, A.; Frago, L.M.; Alvarez, L.; Varela-Nieto, I. Stimulation of DNA synthesis by natural ceramide 1-phosphate. Biochem. J. 1997, 325 Pt 2, 435–440. [Google Scholar] [PubMed]
- Gangoiti, P.; Granado, M.H.; Wang, S.W.; Kong, J.Y.; Steinbrecher, U.P.; Gomez-Munoz, A. Ceramide 1-phosphate stimulates macrophage proliferation through activation of the PI3-kinase/PKB, JNK and ERK1/2 pathways. Cell. Signal. 2008, 20, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Gangoiti, P.; Arana, L.; Ouro, A.; Granado, M.H.; Trueba, M.; Gomez-Munoz, A. Activation of mTOR and RhoA is a major mechanism by which Ceramide 1-phosphate stimulates macrophage proliferation. Cell. Signal. 2010, 23, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Gangoiti, P.; Bernacchioni, C.; Donati, C.; Cencetti, F.; Ouro, A.; Gomez-Munoz, A.; Bruni, P. Ceramide 1-phosphate stimulates proliferation of C2C12 myoblasts. Biochimie 2012, 94, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Arana, L.; Gangoiti, P.; Ouro, A.; Rivera, I.G.; Ordonez, M.; Trueba, M.; Lankalapalli, R.S.; Bittman, R.; Gomez-Munoz, A. Generation of reactive oxygen species (ROS) is a key factor for stimulation of macrophage proliferation by ceramide 1-phosphate. Exp. Cell Res. 2012, 318, 350–360. [Google Scholar] [CrossRef]
- Gomez-Munoz, A.; Kong, J.Y.; Salh, B.; Steinbrecher, U.P. Ceramide-1-phosphate blocks apoptosis through inhibition of acid sphingomyelinase in macrophages. J. Lipid Res. 2004, 45, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Gangoiti, P.; Granado, M.H.; Arana, L.; Ouro, A.; Gomez-Munoz, A. Involvement of nitric oxide in the promotion of cell survival by ceramide 1-phosphate. FEBS Lett. 2008, 582, 2263–2269. [Google Scholar] [CrossRef] [PubMed]
- Granado, M.H.; Gangoiti, P.; Ouro, A.; Arana, L.; Gomez-Munoz, A. Ceramide 1-phosphate inhibits serine palmitoyltransferase and blocks apoptosis in alveolar macrophages. Biochim. Biophys. Acta 2009, 1791, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Granado, M.H.; Gangoiti, P.; Ouro, A.; Arana, L.; Gonzalez, M.; Trueba, M.; Gomez-Munoz, A. Ceramide 1-phosphate (C1P) promotes cell migration Involvement of a specific C1P receptor. Cell. Signal. 2009, 21, 405–412. [Google Scholar] [CrossRef]
- Arana, L.; Ordonez, M.; Ouro, A.; Rivera, I.G.; Gangoiti, P.; Trueba, M.; Gomez-Munoz, A. Ceramide 1-phosphate induces macrophage chemoattractant protein-1 release: Involvement in ceramide 1-phosphate-stimulated cell migration. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1213–E1226. [Google Scholar] [CrossRef]
- Wijesinghe, D.S.; Lamour, N.F.; Gomez-Munoz, A.; Chalfant, C.E. Ceramide kinase and ceramide-1-phosphate. Methods Enzymol. 2007, 434, 265–292. [Google Scholar] [PubMed]
- Lamour, N.F.; Chalfant, C.E. Ceramide-1-phosphate: The “missing” link in eicosanoid biosynthesis and inflammation. Mol. Interv. 2005, 5, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Chalfant, C.E.; Spiegel, S. Sphingosine 1-phosphate and ceramide 1-phosphate: Expanding roles in cell signaling. J. Cell Sci. 2005, 118 Pt 20, 4605–4612. [Google Scholar] [CrossRef]
- Wijesinghe, D.S.; Subramanian, P.; Lamour, N.F.; Gentile, L.B.; Granado, M.H.; Bielawska, A.; Szulc, Z.; Gomez-Munoz, A.; Chalfant, C.E. Chain length specificity for activation of cPLA2alpha by C1P: Use of the dodecane delivery system to determine lipid-specific effects. J. Lipid Res. 2009, 50, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Soucek, A.; Michalec, C.; Souckova, A. Identification and characterization of a new enzyme of the group “phospholipase D” isolated from Corynebacterium ovis. Biochim. Biophys. Acta 1971, 227, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lynch, K.R. Brown recluse spider (Loxosceles reclusa) venom phospholipase D (PLD) generates lysophosphatidic acid (LPA). Biochem. J. 2005, 391 Pt 2, 317–323. [Google Scholar] [CrossRef]
- Van Meeteren, L.A.; Frederiks, F.; Giepmans, B.N.; Pedrosa, M.F.; Billington, S.J.; Jost, B.H.; Tambourgi, D.V.; Moolenaar, W.H. Spider and bacterial sphingomyelinases D target cellular lysophosphatidic acid receptors by hydrolyzing lysophosphatidylcholine. J. Biol. Chem. 2004, 279, 10833–10836. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Munoz, A. Modulation of cell signalling by ceramides. Biochim. Biophys. Acta 1998, 1391, 92–109. [Google Scholar] [CrossRef] [PubMed]
- Durgam, G.G.; Tsukahara, R.; Makarova, N.; Walker, M.D.; Fujiwara, Y.; Pigg, K.R.; Baker, D.L.; Sardar, V.M.; Parrill, A.L.; Tigyi, G.; et al. Synthesis and pharmacological evaluation of second-generation phosphatidic acid derivatives as lysophosphatidic acid receptor ligands. Bioorg. Med. Chem. Lett. 2006, 16, 633–640. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, T.M.; Pontsler, A.V.; Silva, A.R.; St Hilaire, A.; Xu, Y.; Hinshaw, J.C.; Zimmerman, G.A.; Hama, K.; Aoki, J.; Arai, H.; Prestwich, G.D. Identification of an intracellular receptor for lysophosphatidic acid (LPA): LPA is a transcellular PPARgamma agonist. Proc. Natl. Acad. Sci. USA 2003, 100, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Munoz, A.; Kong, J.Y.; Parhar, K.; Wang, S.W.; Gangoiti, P.; Gonzalez, M.; Eivemark, S.; Salh, B.; Duronio, V.; Steinbrecher, U.P. Ceramide-1-phosphate promotes cell survival through activation of the phosphatidylinositol 3-kinase/protein kinase B pathway. FEBS Lett. 2005, 579, 3744–3750. [Google Scholar] [CrossRef] [PubMed]
- Bini, F.; Frati, A.; Garcia-Gil, M.; Battistini, C.; Granado, M.; Martinesi, M.; Mainardi, M.; Vannini, E.; Luzzati, F.; Caleo, M.; et al. New signalling pathway involved in the anti-proliferative action of vitamin D(3) and its analogues in human neuroblastoma cells. A role for ceramide kinase. Neuropharmacology 2012, 63, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Mitra, P.; Maceyka, M.; Payne, S.G.; Lamour, N.; Milstien, S.; Chalfant, C.E.; Spiegel, S. Ceramide kinase regulates growth and survival of A549 human lung adenocarcinoma cells. FEBS Lett. 2007, 581, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Ito, H.; Hagiwara, K.; Yoshida, K.; Sobue, S.; Ichihara, M.; Takagi, A.; Kojima, T.; Tanaka, K.; Tamiya-Koizumi, K.; et al. ATRA inhibits ceramide kinase transcription in a human neuroblastoma cell line, SH-SY5Y cells: The role of COUP-TFI. J. Neurochem. 2010, 112, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Simanshu, D.K.; Kamlekar, R.K.; Wijesinghe, D.S.; Zou, X.; Zhai, X.; Mishra, S.K.; Molotkovsky, J.G.; Malinina, L.; Hinchcliffe, E.H.; Chalfant, C.E.; et al. Non-vesicular trafficking by a ceramide-1-phosphate transfer protein regulates eicosanoids. Nature 2013, 500, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Lankalapalli, R.S.; Ouro, A.; Arana, L.; Gomez-Munoz, A.; Bittman, R. Caged ceramide 1-phosphate analogues: Synthesis and properties. J. Org. Chem. 2009, 74, 8844–8847. [Google Scholar] [CrossRef] [PubMed]
- Pettus, B.J.; Bielawska, A.; Spiegel, S.; Roddy, P.; Hannun, Y.A.; Chalfant, C.E. Ceramide kinase mediates cytokine- and calcium ionophore-induced arachidonic acid release. J. Biol. Chem. 2003, 278, 38206–38213. [Google Scholar] [CrossRef] [PubMed]
- Pettus, B.J.; Bielawska, A.; Subramanian, P.; Wijesinghe, D.S.; Maceyka, M.; Leslie, C.C.; Evans, J.H.; Freiberg, J.; Roddy, P.; Hannun, Y.A.; et al. Ceramide 1-phosphate is a direct activator of cytosolic phospholipase A2. J. Biol. Chem. 2004, 279, 11320–11326. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, P.; Stahelin, R.V.; Szulc, Z.; Bielawska, A.; Cho, W.; Chalfant, C.E. Ceramide 1-phosphate acts as a positive allosteric activator of group IVA cytosolic phospholipase A2 alpha and enhances the interaction of the enzyme with phosphatidylcholine. J. Biol. Chem. 2005, 280, 17601–17607. [Google Scholar] [CrossRef] [PubMed]
- Lamour, N.F.; Wijesinghe, D.S.; Mietla, J.A.; Ward, K.E.; Stahelin, R.V.; Chalfant, C.E. Ceramide kinase regulates the production of tumor necrosis factor alpha (TNFalpha) via inhibition of TNFalpha-converting enzyme. J. Biol. Chem. 2011, 286, 42808–42817. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Kono, K.; Liu, H.; Shimizugawa, T.; Minekura, H.; Spiegel, S.; Kohama, T. Ceramide kinase, a novel lipid kinase. Molecular cloning and functional characterization. J. Biol. Chem. 2002, 277, 23294–23300. [Google Scholar] [CrossRef] [PubMed]
- Shinghal, R.; Scheller, R.H.; Bajjalieh, S.M. Ceramide 1-phosphate phosphatase activity in brain. J. Neurochem. 1993, 61, 2279–2285. [Google Scholar] [CrossRef] [PubMed]
- Pettus, B.J.; Kitatani, K.; Chalfant, C.E.; Taha, T.A.; Kawamori, T.; Bielawski, J.; Obeid, L.M.; Hannun, Y.A. The coordination of prostaglandin E2 production by sphingosine-1-phosphate and ceramide-1-phosphate. Mol. Pharmacol. 2005, 68, 330–335. [Google Scholar] [PubMed]
- Hinkovska-Galcheva, V.; Boxer, L.A.; Kindzelskii, A.; Hiraoka, M.; Abe, A.; Goparju, S.; Spiegel, S.; Petty, H.R.; Shayman, J.A. Ceramide 1-phosphate, a mediator of phagocytosis. J. Biol. Chem. 2005, 280, 26612–26621. [Google Scholar] [CrossRef]
- Hinkovska-Galcheva, V.T.; Boxer, L.A.; Mansfield, P.J.; Harsh, D.; Blackwood, A.; Shayman, J.A. The formation of ceramide-1-phosphate during neutrophil phagocytosis and its role in liposome fusion. J. Biol. Chem. 1998, 273, 33203–33209. [Google Scholar] [CrossRef] [PubMed]
- Mitsutake, S.; Kim, T.J.; Inagaki, Y.; Kato, M.; Yamashita, T.; Igarashi, Y. Ceramide kinase is a mediator of calcium-dependent degranulation in mast cells. J. Biol. Chem. 2004, 279, 17570–17577. [Google Scholar] [CrossRef] [PubMed]
- Mitsutake, S.; Date, T.; Yokota, H.; Sugiura, M.; Kohama, T.; Igarashi, Y. Ceramide kinase deficiency improves diet-induced obesity and insulin resistance. FEBS Lett. 2012, 586, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Wu, W.; Wysoczynski, M.; Abdel-Latif, A.; Sunkara, M.; Morris, A.; Kucia, M.; Ratajczak, J.; Ratajczak, M.Z. Conditioning for hematopoietic transplantation activates the complement cascade and induces a proteolytic environment in bone marrow: A novel role for bioactive lipids and soluble C5b-C9 as homing factors. Leukemia 2011, 26, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Schneider, G.; Abdel-Latif, A.; Mierzejewska, K.; Sunkara, M.; Borkowska, S.; Ratajczak, J.; Morris, A.J.; Kucia, M.; Ratajczak, M.Z. Ceramide-1-phosphate Regulates Migration of Multipotent Stromal Cells (MSCs) and Endothelial Progenitor Cells (EPCs)–Implications for Tissue Regeneration. Stem Cells 2013, 31, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Karapetyan, A.V.; Klyachkin, Y.M.; Selim, S.; Sunkara, M.; Ziada, K.M.; Cohen, D.A.; Zuba-Surma, E.K.; Ratajczak, J.; Smyth, S.S.; Ratajczak, M.Z.; et al. Bioactive lipids and cationic antimicrobial peptides as new potential regulators for trafficking of bone marrow-derived stem cells in patients with acute myocardial infarction. Stem Cells Dev. 2013, 22, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Ouro, A.; Arana, L.; Rivera, I.G.; Ordonez, M.; Gomez-Larrauri, A.; Presa, N.; Simon, J.; Trueba, M.; Gangoiti, P.; Bittman, R.; et al. Phosphatidic acid inhibits ceramide 1-phosphate-stimulated macrophage migration. Biochem. Pharmacol. 2014, 92, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Baudiss, K.; Ayata, C.K.; Lazar, Z.; Cicko, S.; Beckert, J.; Meyer, A.; Zech, A.; Vieira, R.P.; Bittman, R.; Gomez-Munoz, A.; et al. Ceramide-1-phosphate inhibits cigarette smoke-induced airway inflammation. Eur. Respir. J. 2015, in press. [Google Scholar]
- Gulbins, E.; Kolesnick, R. Raft ceramide in molecular medicine. Oncogene 2003, 22, 7070–7077. [Google Scholar] [CrossRef] [PubMed]
- Goggel, R.; Winoto-Morbach, S.; Vielhaber, G.; Imai, Y.; Lindner, K.; Brade, L.; Brade, H.; Ehlers, S.; Slutsky, A.S.; Schutze, S.; et al. PAF-mediated pulmonary edema: A new role for acid sphingomyelinase and ceramide. Nat. Med. 2004, 10, 155–160. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera, I.-G.; Ordoñez, M.; Presa, N.; Gomez-Larrauri, A.; Simón, J.; Trueba, M.; Gomez-Muñoz, A. Sphingomyelinase D/Ceramide 1-Phosphate in Cell Survival and Inflammation. Toxins 2015, 7, 1457-1466. https://doi.org/10.3390/toxins7051457
Rivera I-G, Ordoñez M, Presa N, Gomez-Larrauri A, Simón J, Trueba M, Gomez-Muñoz A. Sphingomyelinase D/Ceramide 1-Phosphate in Cell Survival and Inflammation. Toxins. 2015; 7(5):1457-1466. https://doi.org/10.3390/toxins7051457
Chicago/Turabian StyleRivera, Io-Guané, Marta Ordoñez, Natalia Presa, Ana Gomez-Larrauri, Jorge Simón, Miguel Trueba, and Antonio Gomez-Muñoz. 2015. "Sphingomyelinase D/Ceramide 1-Phosphate in Cell Survival and Inflammation" Toxins 7, no. 5: 1457-1466. https://doi.org/10.3390/toxins7051457
APA StyleRivera, I. -G., Ordoñez, M., Presa, N., Gomez-Larrauri, A., Simón, J., Trueba, M., & Gomez-Muñoz, A. (2015). Sphingomyelinase D/Ceramide 1-Phosphate in Cell Survival and Inflammation. Toxins, 7(5), 1457-1466. https://doi.org/10.3390/toxins7051457