Helicobacter pylori Infection-Induced Hepatoma-Derived Growth Factor Regulates the Differentiation of Human Mesenchymal Stem Cells to Myofibroblast-Like Cells


Abstract
:1. Introduction
2. Results
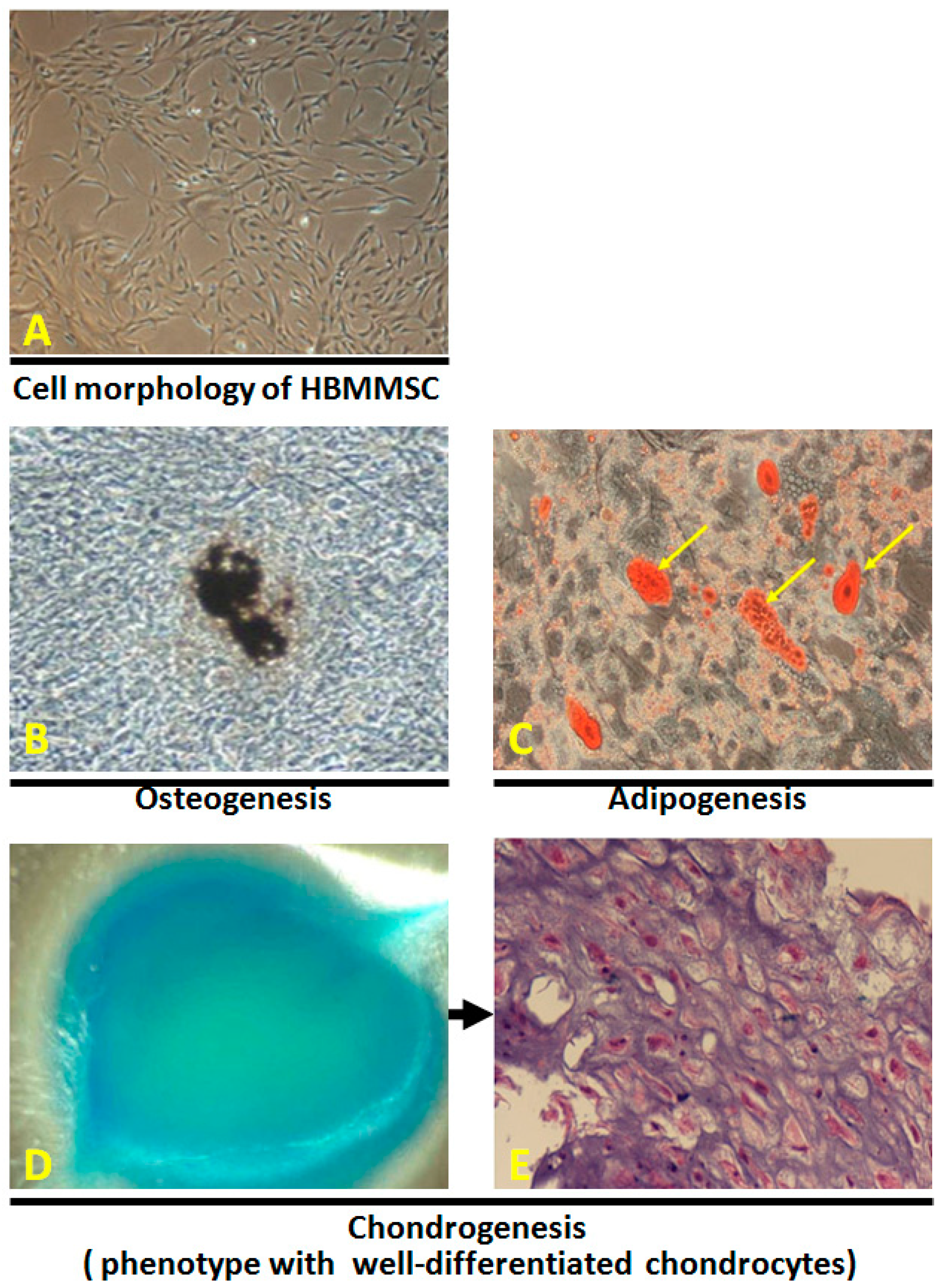
2.1. In Vitro Differentiation of Osteocytes, Adipocytes and Chondrocytes from Human Bone Marrow-Derived Mesenchymal Stem Cells
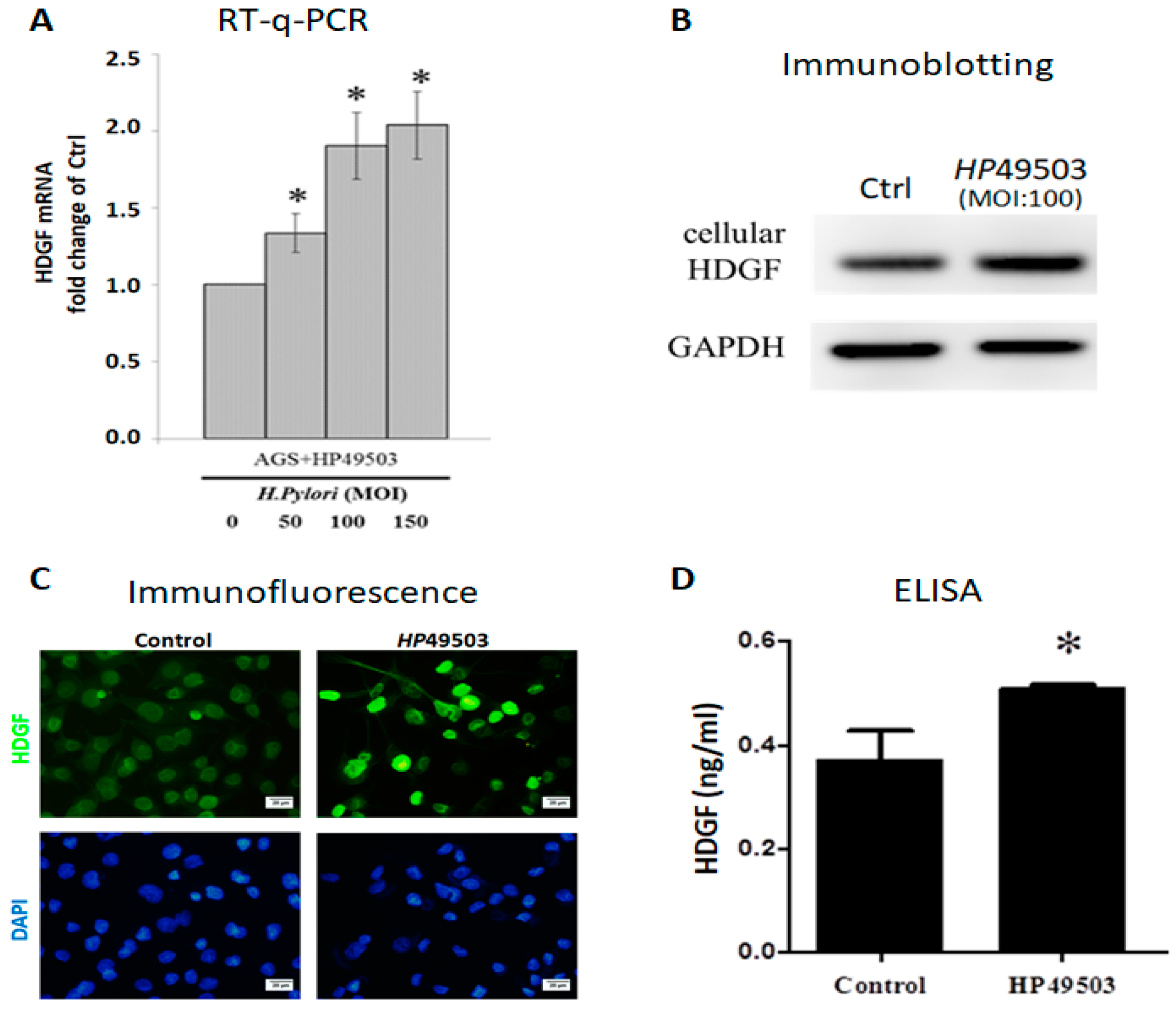
2.2. H. Pylori Infection Induces HDGF Expression in Human Gastric Cancer Cells
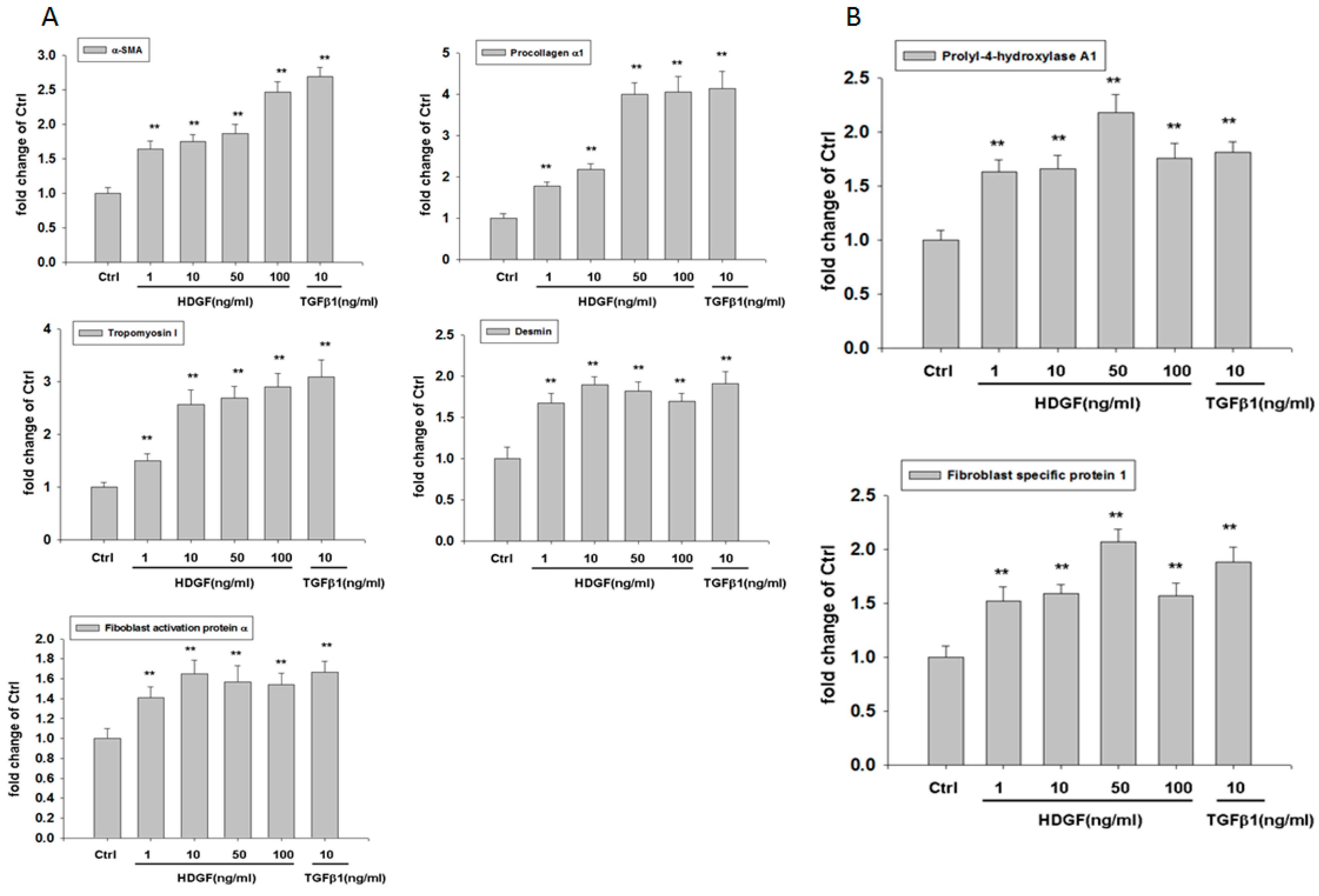
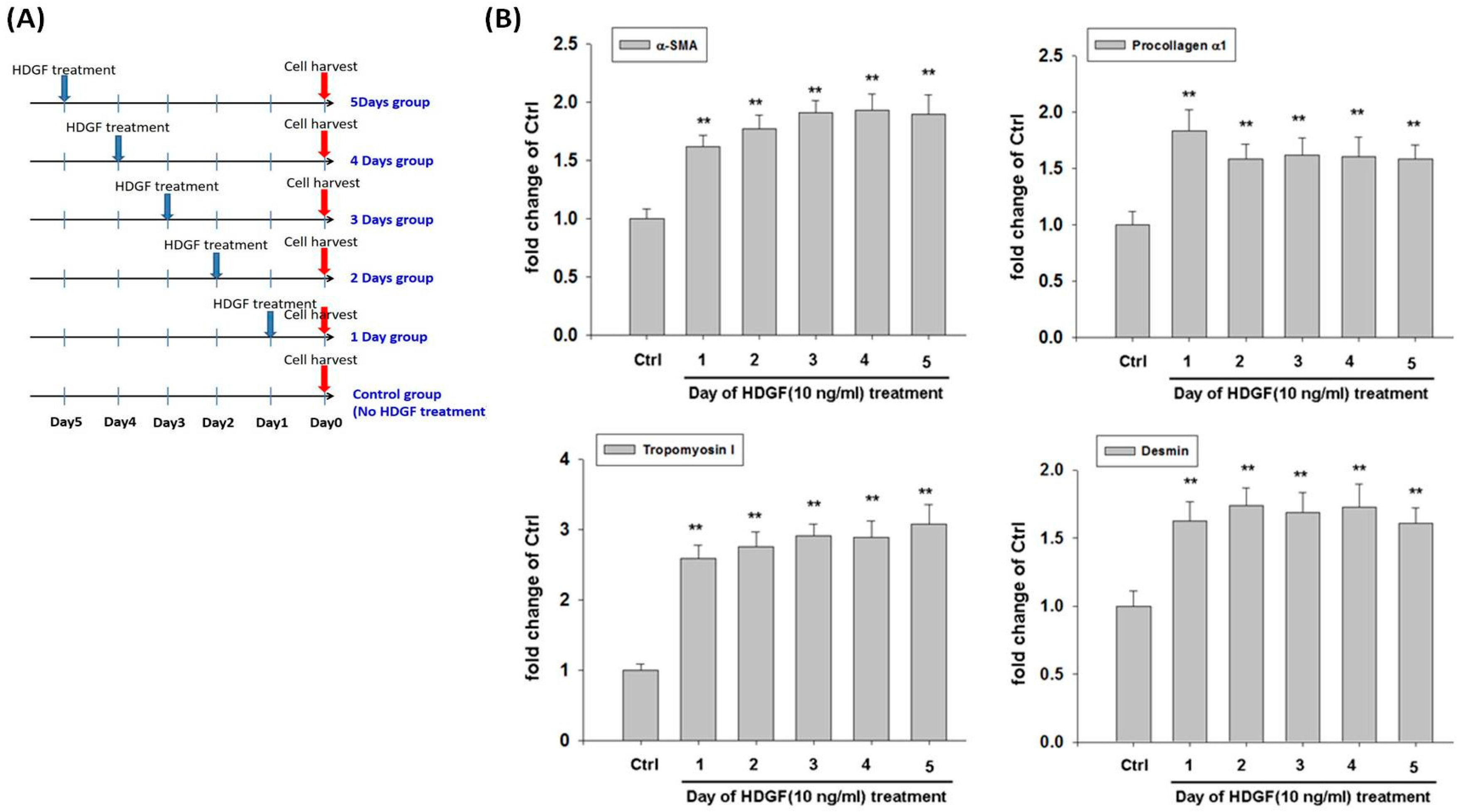
2.3. HDGF Induces Expression of Myofibroblast and Fibroblast Markers in HBMMSCs
2.4. HDGF Neutralizing Antibody and Serum Inhibits the Expression of Myofibroblast and Fibroblast Markers
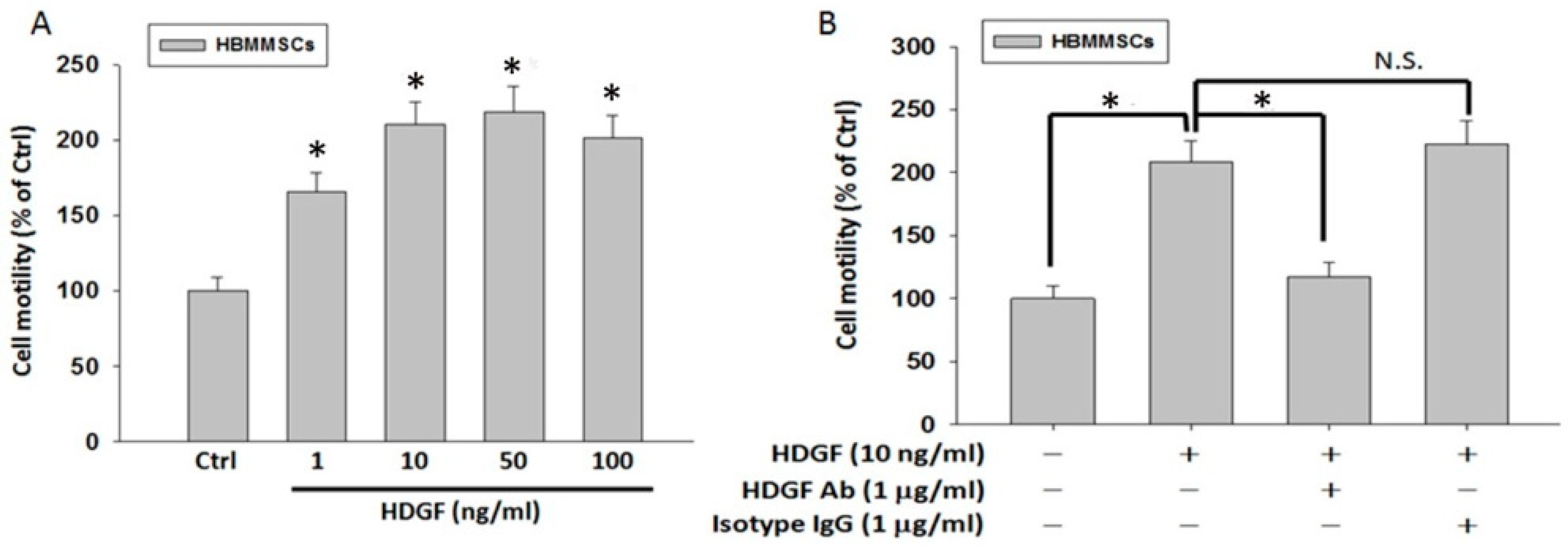
2.5. HDGF Neutralizing Antibody Inhibits HDGF-Induced HBMMSCs Recruitment
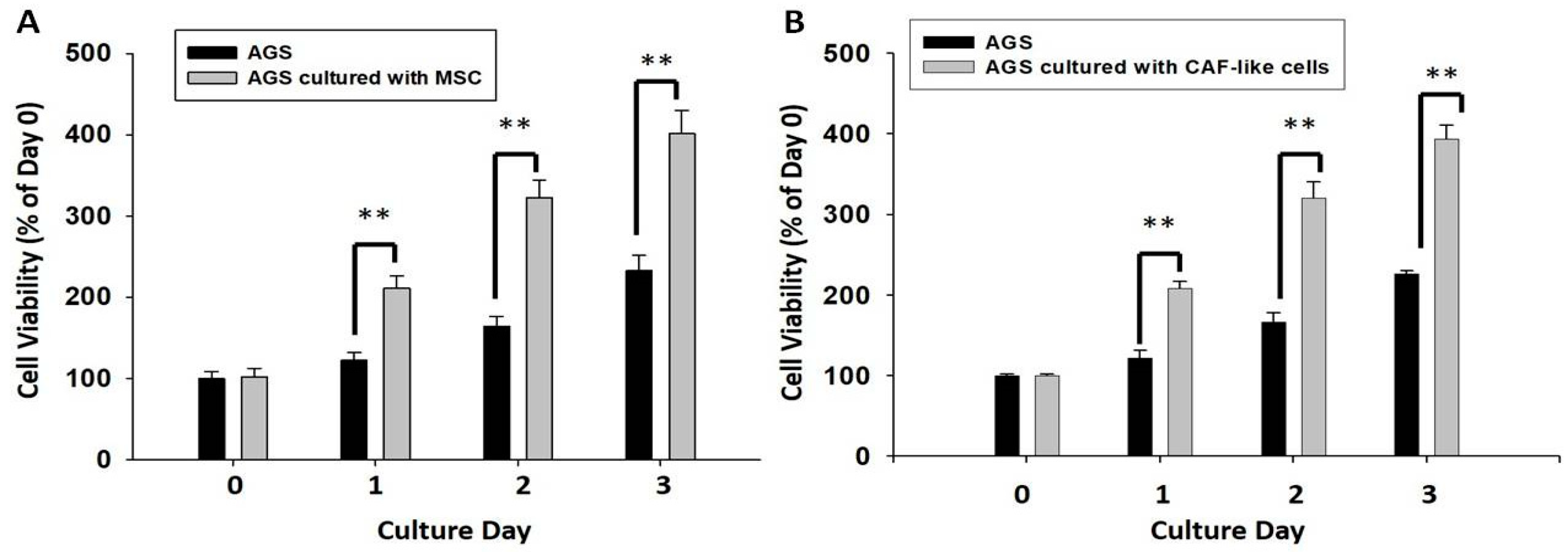
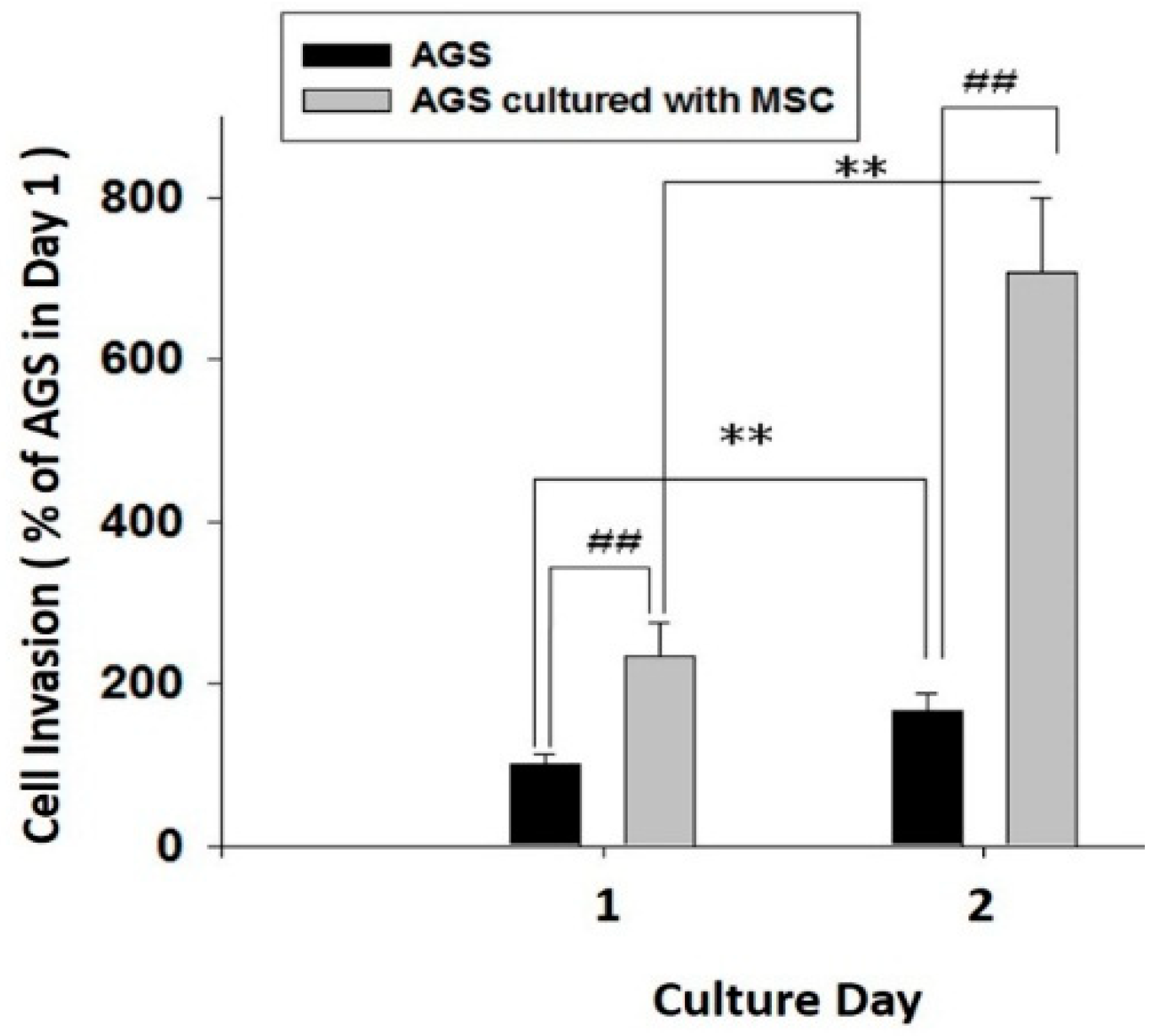
2.6. HBMMSCs Upregulate Survival and Invasive Motility in Human Gastric Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cells, Antibodies, Reagents and Enzymes
4.2. Cell Culture
4.3. H. Pylori Culture
4.4. Immunoblotting
4.5. Immunofluorescence Assay
4.6. Immunohistochemical Staining
4.7. ELISA
4.8. HBMMSCs Recruitment Assay
4.9. Invasive Motility Assay
4.10. Cell Viability Assay
4.11. Multilineage Differentiation
4.11.1. Osteogenic Differentiation
4.11.2. Adipogenic Differentiation
4.11.3. Chondrogenic Differentiation
4.11.4. Cytochemical Staining
4.11.5. Histological Analysis
4.12. Total RNA Extraction
4.13. Reverse Transcription and Real-Time PCR Assay
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shiga, K.; Hara, M.; Nagasaki, T.; Sato, T.; Takahashi, H.; Takeyama, H. Cancer-Associated Fibroblasts: Their Characteristics and Their Roles in Tumor Growth. Cancers 2015, 7, 2443–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharaishvili, G.; Simkova, D.; Bouchalova, K.; Gachechiladze, M.; Narsia, N.; Bouchal, J. The role of cancer-associated fibroblasts, solid stress and other microenvironmental factors in tumor progression and therapy resistance. Cancer Cell Int. 2014, 14, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.; Gao, J.; Wang, S.; Ye, N.; Chong, Y.; Huang, Y.; Wang, J.; Li, B.; Yin, W.; Wang, D. Cancer-associated fibroblasts promote angiogenesis in gastric cancer through galectin-1 expression. Tumour Biol. 2016, 37, 1889–1899. [Google Scholar] [CrossRef]
- Kurashige, J.; Mima, K.; Sawada, G.; Takahashi, Y.; Eguchi, H.; Sugimachi, K.; Mori, M.; Yanagihara, K.; Yashiro, M.; Hirakawa, K.; et al. Epigenetic modulation and repression of miR-200b by cancer-associated fibroblasts contribute to cancer invasion and peritoneal dissemination in gastric cancer. Carcinogenesis 2015, 36, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Fukui, H.; Hara, K.; Zhang, X.; Kitayama, Y.; Eda, H.; Tomita, T.; Oshima, T.; Kikuchi, S.; Watari, J.; et al. FGF9 from cancer-associated fibroblasts is a possible mediator of invasion and anti-apoptosis of gastric cancer cells. BMC Cancer 2015, 15, 333. [Google Scholar] [CrossRef] [PubMed]
- Togo, S.; Polanska, U.M.; Horimoto, Y.; Orimo, A. Carcinoma-associated fibroblasts are a promising therapeutic target. Cancers 2013, 5, 149–169. [Google Scholar] [CrossRef]
- Barry, F.P.; Murphy, J.M. Mesenchymal stem cells: Clinical applications and biological characterization. Int. J. Biochem. Cell Biol. 2004, 36, 568–584. [Google Scholar] [CrossRef] [PubMed]
- Loebinger, M.R.; Kyrtatos, P.G.; Turmaine, M.; Price, A.N.; Pankhurst, Q.; Lythgoe, M.F.; Janes, S.M. Magnetic resonance imaging of mesenchymal stem cells homing to pulmonary metastases using biocompatible magnetic nanoparticles. Cancer Res. 2009, 69, 8862–8867. [Google Scholar] [CrossRef] [PubMed]
- Meads, M.B.; Hazlehurst, L.A.; Dalton, W.S. The bone marrow microenvironment as a tumor sanctuary and contributor to drug resistance. Clin. Cancer Res. 2008, 14, 2519–2526. [Google Scholar] [CrossRef] [PubMed]
- Houghton, J.; Stoicov, C.; Nomura, S.; Rogers, A.B.; Carlson, J.; Li, H.; Cai, X.; Fox, J.G.; Goldenring, J.R.; Wang, T.C. Gastric cancer originating from bone marrow-derived cells. Science 2004, 306, 1568–1571. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.R.; Chae, S.S.; Jain, R.K.; Duda, D.G. Direct evidence for lineage-dependent effects of bone marrow stromal cells on tumor progression. Am. J. Cancer Res. 2011, 1, 144–154. [Google Scholar] [PubMed]
- Hung, S.C.; Pochampally, R.R.; Chen, S.C.; Hsu, S.C.; Prockop, D.J. Angiogenic effects of human multipotent stromal cell conditioned medium activate the PI3K-Akt pathway in hypoxic endothelial cells to inhibit apoptosis, increase survival, and stimulate angiogenesis. Stem Cells 2007, 25, 2363–2370. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.J.; Mishra, P.J.; Humeniuk, R.; Medina, D.J.; Alexe, G.; Mesirov, J.P.; Ganesan, S.; Glod, J.W.; Banerjee, D. Carcinoma-associated fibroblast-like differentiation of human mesenchymal stem cells. Cancer Res. 2008, 68, 4331–4339. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.J.; Kuo, F.C.; Wang, C.L.; Kuo, C.H.; Wang, S.S.; Chen, C.Y.; Huang, Y.B.; Cheng, K.H.; Yokoyama, K.K.; Chen, C.L.; et al. Suppression of IL-8-Src signalling axis by 17beta-estradiol inhibits human mesenchymal stem cells-mediated gastric cancer invasion. J. Cell Mol. Med. 2016, 20, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.J.; Kuo, F.C.; Hu, H.M.; Chen, C.Y.; Huang, Y.B.; Cheng, K.H.; Yokoyama, K.K.; Wu, D.C.; Hsieh, S.; Kuo, C.H. 17beta-Estradiol inhibition of IL-6-Src and Cas and paxillin pathway suppresses human mesenchymal stem cells-mediated gastric cancer cell motility. Transl. Res. 2014, 164, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Liu, C.J.; Lu, C.Y.; Hu, H.M.; Kuo, F.C.; Liou, Y.S.; Yang, Y.C.; Hsieh, M.C.; Lee, O.K.; Wu, D.C.; et al. 17beta-estradiol inhibits mesenchymal stem cells-induced human AGS gastric cancer cell mobility via suppression of CCL5- Src/Cas/Paxillin signaling pathway. Int. J. Med. Sci 2014, 11, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Crew, K.D.; Neugut, A.I. Epidemiology of gastric cancer. World J. Gastroenterol. 2006, 12, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Izumoto, Y.; Kambe, H.; Kuroda, T.; Mori, T.; Kawamura, K.; Yamamoto, H.; Kishimoto, T. Molecular cloning of complementary DNA for a novel human hepatoma-derived growth factor. Its homology with high mobility group-1 protein. J. Biol. Chem. 1994, 269, 25143–25149. [Google Scholar] [PubMed]
- Yamamoto, S.; Tomita, Y.; Hoshida, Y.; Takiguchi, S.; Fujiwara, Y.; Yasuda, T.; Doki, Y.; Yoshida, K.; Aozasa, K.; Nakamura, H.; et al. Expression of hepatoma-derived growth factor is correlated with lymph node metastasis and prognosis of gastric carcinoma. Clin. Cancer Res. 2006, 12, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Xu, Z.; Fang, Y.; Wang, H.; Xu, J.; Ye, J.; Zheng, S.; Zhu, Y. Hepatoma-derived growth factor involved in the carcinogenesis of gastric epithelial cells through promotion of cell proliferation by Erk1/2 activation. Cancer Sci. 2008, 99, 2120–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Choi, E.Y.; Kim, M.K.; Lee, S.H.; Jang, B.I.; Kim, T.N.; Kim, S.W.; Kim, S.W.; Song, S.K.; Kim, J.R.; et al. Hepatoma-derived growth factor regulates the bad-mediated apoptotic pathway and induction of vascular endothelial growth factor in stomach cancer cells. Oncol. Res. 2010, 19, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Pietras, K.; Ostman, A. Hallmarks of cancer: Interactions with the tumor stroma. Exp. Cell Res. 2010, 316, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Tu, S.P.; Tomita, H.; Gonda, T.; Wang, S.S.; Takashi, S.; Baik, G.H.; Shibata, W.; Diprete, B.; Betz, K.S.; et al. Bone marrow-derived myofibroblasts contribute to the mesenchymal stem cell niche and promote tumor growth. Cancer Cell 2011, 19, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Lotti, F.; Jarrar, A.M.; Pai, R.K.; Hitomi, M.; Lathia, J.; Mace, A.; Gantt, G.A., Jr.; Sukhdeo, K.; DeVecchio, J.; Vasanji, A.; et al. Chemotherapy activates cancer-associated fibroblasts to maintain colorectal cancer-initiating cells by IL-17A. J. Exp. Med. 2013, 210, 2851–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serini, G.; Gabbiani, G. Mechanisms of myofibroblast activity and phenotypic modulation. Exp. Cell Res. 1999, 250, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Lenter, M.C.; Zimmermann, R.N.; Garin-Chesa, P.; Old, L.J.; Rettig, W.J. Fibroblast activation protein, a dual specificity serine protease expressed in reactive human tumor stromal fibroblasts. J. Biol. Chem. 1999, 274, 36505–36512. [Google Scholar] [CrossRef] [PubMed]
- Strutz, F.; Okada, H.; Lo, C.W.; Danoff, T.; Carone, R.L.; Tomaszewski, J.E.; Neilson, E.G. Identification and characterization of a fibroblast marker: FSP1. J. Cell Biol. 1995, 130, 393–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, Y.; Acar, A.; Eaton, E.N.; Mellody, K.T.; Scheel, C.; Ben-Porath, I.; Onder, T.T.; Wang, Z.C.; Richardson, A.L.; Weinberg, R.A.; et al. Autocrine TGF-β and stromal cell-derived factor-1 (SDF-1) signaling drives the evolution of tumor-promoting mammary stromal myofibroblasts. Proc. Natl. Acad. Sci. USA 2010, 107, 20009–20014. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Tajima, H.; Nakanuma, S.; Sakai, S.; Makino, I.; Kinoshita, J.; Hayashi, H.; Nakamura, K.; Oyama, K.; Nakagawara, H.; et al. Angiotensin II enhances epithelial-to-mesenchymal transition through the interaction between activated hepatic stellate cells and the stromal cell-derived factor-1/CXCR4 axis in intrahepatic cholangiocarcinoma. Int. J. Oncol. 2012, 41, 573–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhoopathi, P.; Chetty, C.; Gogineni, V.R.; Gujrati, M.; Dinh, D.H.; Rao, J.S.; Lakka, S.S. MMP-2 mediates mesenchymal stem cell tropism towards medulloblastoma tumors. Gene. Ther. 2011, 18, 692–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cilley, R.E.; Zgleszewski, S.E.; Chinoy, M.R. Fetal lung development: Airway pressure enhances the expression of developmental genes. J. Pediatr. Surg. 2000, 35, 113–118; discussion 119. [Google Scholar] [CrossRef]
- Oliver, J.A.; Al-Awqati, Q. An endothelial growth factor involved in rat renal development. J. Clin. Invest. 1998, 102, 1208–1219. [Google Scholar] [CrossRef] [PubMed]
- Everett, A.D.; Narron, J.V.; Stoops, T.; Nakamura, H.; Tucker, A. Hepatoma-derived growth factor is a pulmonary endothelial cell-expressed angiogenic factor. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 286, L1194–L1201. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, H.; Yoshida, K.; Kishima, Y.; Kinoshita, T.; Yamamoto, M.; Everett, A.D.; Miyajima, A.; Nakamura, H. Hepatoma-derived growth factor is highly expressed in developing liver and promotes fetal hepatocyte proliferation. Hepatology 2002, 36, 1519–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.H.; Huang, C.C.; Liu, L.F.; Lin, P.R.; Liu, S.Y.; Chang, H.W.; Changchien, C.S.; Lee, C.M.; Chuang, J.H.; Tai, M.H. Expression of hepatoma-derived growth factor in hepatocellular carcinoma. Cancer 2003, 98, 1444–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.S.; Chao, C.C.; Su, T.L.; Yeh, S.H.; Chen, D.S.; Chen, C.T.; Chen, P.J.; Jou, Y.S. Diverse cellular transformation capability of overexpressed genes in human hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2004, 315, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Tang, X.; Lee, J.J.; Feng, L.; Everett, A.D.; Hong, W.K.; Khuri, F.R.; Mao, L. Expression of hepatoma-derived growth factor is a strong prognostic predictor for patients with early-stage non-small-cell lung cancer. J. Clin. Oncol. 2004, 22, 3230–3237. [Google Scholar] [CrossRef]
- Bernard, K.; Litman, E.; Fitzpatrick, J.L.; Shellman, Y.G.; Argast, G.; Polvinen, K.; Everett, A.D.; Fukasawa, K.; Norris, D.A.; Ahn, N.G.; et al. Functional proteomic analysis of melanoma progression. Cancer Res. 2003, 63, 6716–6725. [Google Scholar]
- Zhang, J.; Ren, H.; Yuan, P.; Lang, W.; Zhang, L.; Mao, L. Down-regulation of hepatoma-derived growth factor inhibits anchorage-independent growth and invasion of non-small cell lung cancer cells. Cancer Res. 2006, 66, 18–23. [Google Scholar] [CrossRef]
- Yoshida, K.; Tomita, Y.; Okuda, Y.; Yamamoto, S.; Enomoto, H.; Uyama, H.; Ito, H.; Hoshida, Y.; Aozasa, K.; Nagano, H.; et al. Hepatoma-derived growth factor is a novel prognostic factor for hepatocellular carcinoma. Ann. Surg. Oncol. 2006, 13, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Uyama, H.; Tomita, Y.; Nakamura, H.; Nakamori, S.; Zhang, B.; Hoshida, Y.; Enomoto, H.; Okuda, Y.; Sakon, M.; Aozasa, K.; et al. Hepatoma-derived growth factor is a novel prognostic factor for patients with pancreatic cancer. Clin. Cancer Res. 2006, 12, 6043–6048. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.C.; Tai, M.H.; Lin, J.W.; Wang, C.C.; Huang, C.C.; Hung, C.H.; Chen, C.H.; Lu, S.N.; Lee, C.M.; Changchien, C.S.; et al. Hepatoma-derived growth factor is a novel prognostic factor for gastrointestinal stromal tumors. Int. J. Cancer 2007, 121, 1059–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | Orientation | |
|---|---|---|---|
| Myofibroblast | human α smooth muscle actin (α-SMA) | ACCCACAATGTCCCCATCTA | Forward |
| human α smooth muscle actin (α-SMA) | GAAGGAATAGCCACGCTCAG | Reverse | |
| human procollagen α-I | GTGCTAAAGGTGCCAATGGT | Forward | |
| human procollagen α-I | ACCAGGTTCACCGCTGTTAC | Reverse | |
| human tropomyosin 1 | GAAGCTCGACAAGGAGAACG | Forward | |
| human tropomyosin 1 | TTTTGCAGTGACACCAGCTC | Reverse | |
| human desmin | CAAGCTGCAGGAGGAGATTC | Forward | |
| human desmin | GAGATTCAATTCTGCGCTCC | Reverse | |
| human fibroblast activation protein α (FAP-α) | CGATACCACTTACCCTGCGT | Forward | |
| human fibroblast activation protein α (FAP-α) | ATCAGTAACCCACGTGAGCC | Reverse | |
| Fibroblast | human prolyl 4-hydroxylase A1 (PHA1) | GGCAGCCAAAGCTCTGTTAC | Forward |
| human prolyl 4-hydroxylase A1 (PHA1) | AAAGCAGTCCTCAGCCGTTA | Reverse | |
| human fibroblast specific protein-1 (FSP-1) | GATGAGCAACTTGGACAGCA | Forward | |
| human fibroblast specific protein-1 (FSP-1) | CTTCCTGGGCTGCTTATCTG | Reverse | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-J.; Wang, Y.-K.; Kuo, F.-C.; Hsu, W.-H.; Yu, F.-J.; Hsieh, S.; Tai, M.-H.; Wu, D.-C.; Kuo, C.-H. Helicobacter pylori Infection-Induced Hepatoma-Derived Growth Factor Regulates the Differentiation of Human Mesenchymal Stem Cells to Myofibroblast-Like Cells. Cancers 2018, 10, 479. https://doi.org/10.3390/cancers10120479
Liu C-J, Wang Y-K, Kuo F-C, Hsu W-H, Yu F-J, Hsieh S, Tai M-H, Wu D-C, Kuo C-H. Helicobacter pylori Infection-Induced Hepatoma-Derived Growth Factor Regulates the Differentiation of Human Mesenchymal Stem Cells to Myofibroblast-Like Cells. Cancers. 2018; 10(12):479. https://doi.org/10.3390/cancers10120479
Chicago/Turabian StyleLiu, Chung-Jung, Yao-Kuang Wang, Fu-Chen Kuo, Wen-Hung Hsu, Fang-Jung Yu, Shuchen Hsieh, Ming-Hong Tai, Deng-Chyang Wu, and Chao-Hung Kuo. 2018. "Helicobacter pylori Infection-Induced Hepatoma-Derived Growth Factor Regulates the Differentiation of Human Mesenchymal Stem Cells to Myofibroblast-Like Cells" Cancers 10, no. 12: 479. https://doi.org/10.3390/cancers10120479
APA StyleLiu, C. -J., Wang, Y. -K., Kuo, F. -C., Hsu, W. -H., Yu, F. -J., Hsieh, S., Tai, M. -H., Wu, D. -C., & Kuo, C. -H. (2018). Helicobacter pylori Infection-Induced Hepatoma-Derived Growth Factor Regulates the Differentiation of Human Mesenchymal Stem Cells to Myofibroblast-Like Cells. Cancers, 10(12), 479. https://doi.org/10.3390/cancers10120479