EBV Positive Diffuse Large B Cell Lymphoma and Chronic Lymphocytic Leukemia Patients Exhibit Increased Anti-dUTPase Antibodies


Abstract
:1. Introduction
2. Results
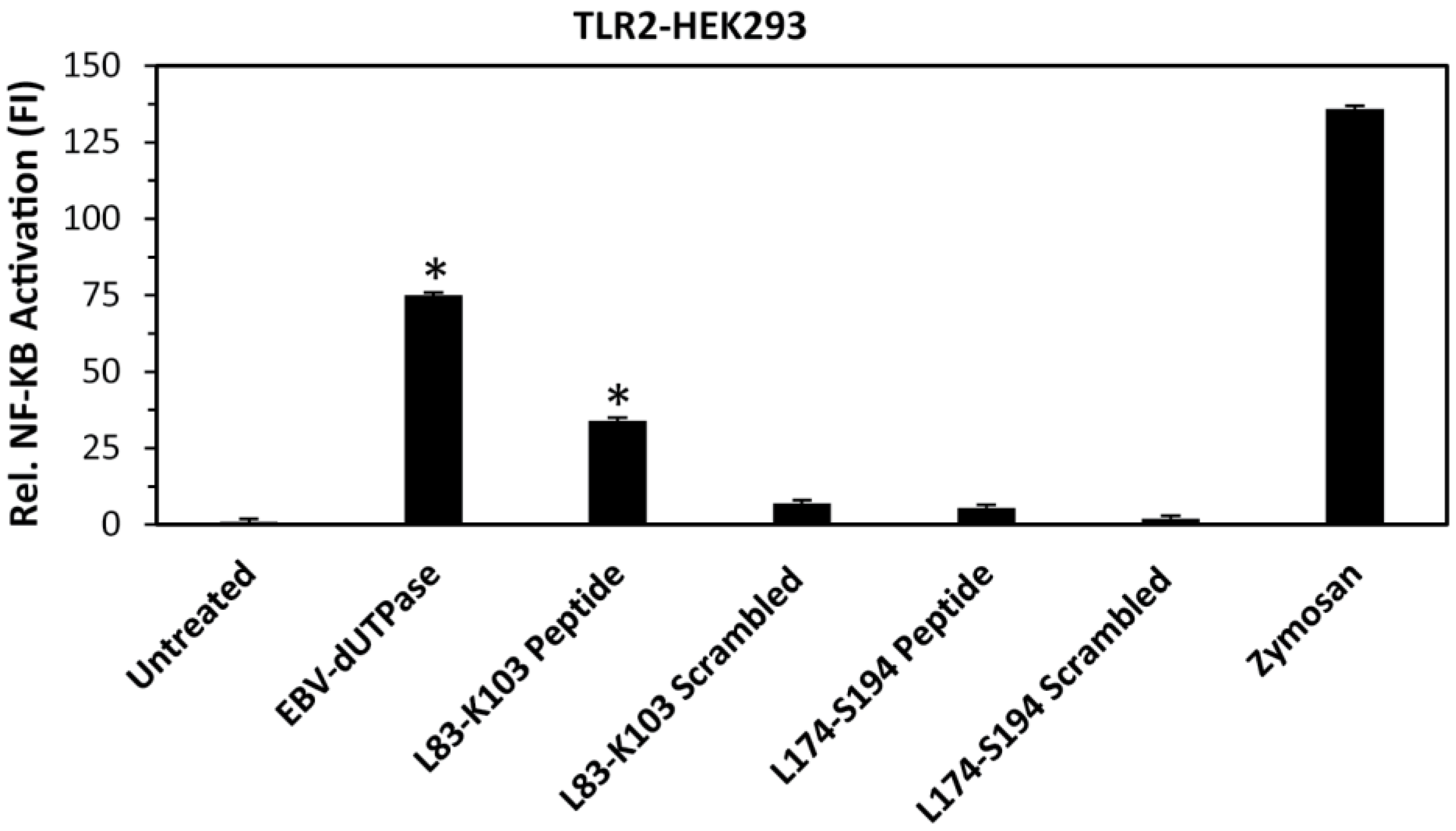
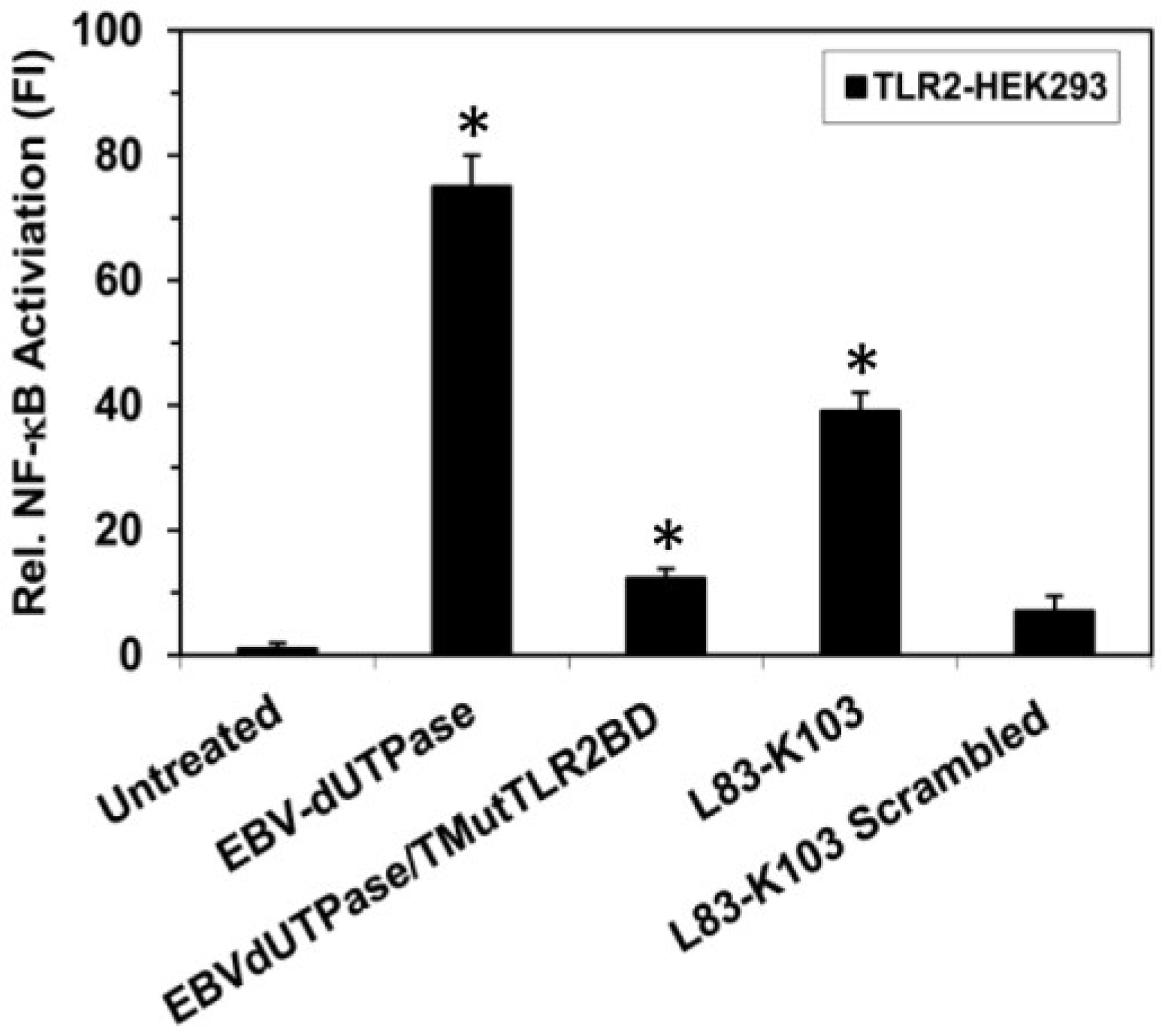
2.1. Identification of a Putative TLR2 Binding Motif within the EBV-dUTPase
2.2. Anti-EBV-Encoded dUTPase Antibody in Patients with DLBCL and Chronic Lymphocytic Leukemia (CLL)
3. Discussion
4. Materials and Methods
4.1. Construction of EBV-dUTPase Triple Mutant
4.2. Peptide Synthesis
4.3. Purification of Recombinant EBV-dUTPase Protein
4.4. EBV-dUTPase Neutralization Assays
4.5. Patients
4.6. Cell Culture
4.7. Luciferase Reporter Gene Assays
4.8. Cytokine Profile Induced by Herpesviruses-Encoded dUTPases
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TLR2 | Toll-like receptor 2 |
| DLBCL | Diffuse large B cell lymphoma |
| CLL | Chronic lymphocytic leukemia |
| EBV | Epstein-Barr virus |
| dUTPase | Deoxyuridine triphosphate nucleotidohydrolase |
| HSV-2 | Herpes simplex virus type 2 |
| HHV-6 | Human herpesvirus 6 |
| HHV-8 | Human herpesvirus 8 |
| VZV | Varicella-zoster virus |
| HEK293 | Human embryonic kidney cells |
| PBMCs | Peripheral blood mononuclear cells |
References
- Kutok, J.L.; Wan, F. Spectrum of Epstein-Barr virus-associated diseases. Annu. Rev. Pathol. Mech. Dis. 2006, 1, 375–404. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Eisen, M.B.; Davis, E.R.; Ma, C.; Lossos, I.S.; Rosenwald, A.; Boldrick, J.C.; Sabet, H.; Tran, T.; Yu, X.; et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 2000, 403, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Shannon-Lowe, C.; Rickinson, A.B.; Bell, A.I. Epstein-Barr virus-associated lymphomas. Philos. Trans. R. Soc. Lond. B Boil. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.; Yamamoto, K.; Asano, N.; Oshiro, A.; Suzuki, R.; Kagami, Y.; Morishima, Y.; Takeuchi, K.; Izumo, T.; Mori, S.; et al. Age-related EBV-associated B-cell lymphoproliferative disorders constitute a distinct clinicopathologic group: A study of 96 patients. Clin. Cancer Res. 2007, 13, 5124–5132. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, J.; Ko, Y.H.; Han, A.; Jun, H.J.; Lee, S.C.; Hwang, I.G.; Park, Y.H.; Ahn, J.S.; Jung, C.J.; et al. The impact of Epstein-Barr virus status on clinical outcome in diffuse large B-cell lymphoma. Blood 2007, 110, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y.; Yamamoto, K.; Asano, N.; Oyama, T.; Kinoshita, T.; Nakamura, S. Age-related Epstein-Barr virus-associated B-cell lymphoproliferative disorders: Special references to lymphomas surrounding this newly recognized clinicopatholoogic disease. Cancer Sci. 2008, 99, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Asano, N.; Yamamoto, K.; Tamaru, J.I.; Oyama, T.; Ishida, F.; Ohshima, K.; Yoshino, T.; Nakamura, N.; Mori, S.; Yoshie, O.; et al. Age-related Epstein-Barrvirus (EBV)-associated B-cell lymphoproliferative disorders: Comparison with EBV-positive classic Hodgkin lymphoma in elderly patients. Blood 2009, 113, 2629–2636. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.E.; His, E.D. Epstein-Barr virus-positive B-cell lymphoma of the elderly at a United States tertiary medical center: An uncommon aggressive lymphoma with a nongerminal center B-cell phenotype. Hum. Pathol. 2009, 40, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advanti, R.; Gheelmini, M.; Salles, G.A.; Zelenets, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [PubMed]
- Ghia, P.; Ferreri, A.J.M.; Caligaris-Cappio, F. Chronic lymphocytic leukemia. Crit. Rev. Oncol./Hematol. 2007, 64, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Campo, E.; Swerdlow, S.; Harris, N.L.; Pileri, S.; Stein, H.; Jaffe, E.S. The 2008 WHO classification of lymphoid neoplasms and beyond: Evolving and practical applications. Blood 2011, 117, 5019–5032. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; O’Brien, S. Richter’s transformation in chronic lymphocytic leukemia. Oncology 2012, 26, 1146–1152. [Google Scholar] [PubMed]
- Parikh, S.A.; Shanafelt, T.D. Risk factors for Richter Syndrome in chronic lymphocytic leukemia. Curr. Hematol. Malig. Rep. 2014, 9, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Vitale, C.; Ferrajoli, A. Richter syndrome in chronic lymphocytic leukemia. Curr. Hematol. Malig. Rep. 2016, 11, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Ansell, S.M.; Li, C.Y.; Lloyd, R.V.; Phyliky, R.L. Epstein-Barr virus infection in Richter’s transformation. Am. J. Hematol. 1999, 60, 99–104. [Google Scholar] [CrossRef]
- Dolcetti, R.; Carbone, A. Epstein-Barr virus infection and chronic lymphocytic leukemis: A possible progression factor? Infect. Agents Cancer 2010, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Barchino, M.J.; Sarasquete, M.E.; Panizo, C.; Morscio, J.; Martinez, A.; Alcoceba, M.; Fresquet, V.; Gonzalez-Farre, B.; Paiva, B.; Young, K.H.; et al. Richter transformation driven by Epstein-Barr virus reactivation during therapy-related immunosuppression in chronic lymphocytic leukemia. J. Pathol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Vistraop, A.G.; Huaman, F.; Narbaitz, M.; Metrebian, F.; De Matteo, E.; Preciado, M.V.; Chabay, P.A. Epstein-Barr virus lytic cycle involvement in diffuse large B cell lymphoma. Hematol. Oncol. 2018, 36, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Khanna, R.; Burrows, S.R. Role of cytotoxic T lymphocytes in Epstein-Barr virus-associated diseases. Annu. Rev. Microbiol. 2000, 54, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Hislop, A.D.; Taylor, G.S.; Sauce, D.; Rickinson, A.B. Cellular responses to viral infection in humans: Lessons from Epstein-Barr virus. Annu. Rev. Immunol. 2007, 25, 587–617. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Farre, B.; Rovira, J.; Martinez, D.; Valera, A.; Garcia-Herrera, A.; Marcos, M.A.; Sole, C.; Rouge, G.; Colomer, D.; Gonzalvo, E.; et al. In vivo intratumoral Epstein–Barr virus replication is associated with XBP1 activation and early-onset post-transplant lymphoproliferative disorders with prognostic implications. Mod. Pathol. 2014, 1599–1611. [Google Scholar] [CrossRef] [PubMed]
- Cochet, C.; Martel-Renoir, D.; Greunewald, V.; Bosq, J.; Cochet, G.; Schwaab, G.; Bernaudin, J.F.; Joab, I. Expression of the Epstein-Barr virus immediate early gene, BZLF1, in nasopharyngeal carcinoma tumor cells. Virology 1993, 197, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Martel-Renoir, A.; Grunewald, V.; Touitou, R.; Schwaab, G.; Joab, I. Qualitative analysis of the expression of Epstein-Barr virus lytic genes in nasopharyngeal biopsies. J. Gen. Virol. 1995, 76, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Montone, K.T.; Hodinka, R.L.; Salhany, K.E.; Lavi, E.; Rostami, A.; Tomaszewski, J.E. Identification of Epstein-Barr virus lytic activity in post-transplantion lymphoproliferative disease. Mod. Pathol. 1996, 9, 621–630. [Google Scholar] [PubMed]
- Xue, S.A.; Labrecque, L.G.; Lu, Q.L.; Ong, S.K.; Lampert, I.A.; Kazembe, P.; Molyneux, E.; Broadhead, R.L.; Borgstein, E.; Griffin, B.E. Promiscuous expression of Epstein-Barr virus in Burkitt’s lymphoma from the central Africian country Malawi. Int. J. Cancer 2002, 99, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Kroll, J.; Li, S.; Levi, M.; Weinberg, A. Lytic and latent EBV gene expression in transplant recipients with and without post-transplant lymphoproliferative disorder. J. Clin. Virol. 2011, 52, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Strong, M.J.; Xu, G.; Coco, J.; Baribault, C.; Vinay, D.S.; Lacey, M.R.; Strong, A.L.; Lehman, T.A.; Seddon, M.B.; Lin, L.; et al. Differences in gastric carcinoma microenvironment stratify according to EBV infection intensity: Implications for possible immune adjuvant therapy. PLoS Pathog. 2013, 9, e1003341. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.H.; Raykova, A.; Klinke, O.; Bernhardt, K.; Gartner, K.; Leung, C.S.; Geletneky, K.; Sertel, S.; Munz, C.; Feederie, R.; et al. Spontaneous lytic replication and epitheliotropism define an Epstein-Barr Virus strain found in carcinomas. Cell Rep. 2013, 5, 458–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Strong, M.J.; Wang, X.; Moss, W.N.; Concha, M.; Lin, Z.; O’Grady, T.; Baddoo, M.; Fewell, C.; Renne, R.; et al. High-throughput RNA sequencing-based virome analysis of 50 lymphoma cell lines from the cancer cell line encyclopedia project. J. Virol. 2015, 89, 713–729. [Google Scholar] [CrossRef] [PubMed]
- Tierney, R.J.; Shannon-Lowe, C.D.; Fitzsimmons, L.; Bell, A.I.; Rowe, M. Unexpected patterns of Epstein-Barr virus transcription revealed by a high throughput PCR array for absolute quantification of viral mRNA. Virology 2015, 474, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Ramayanti, O.; Juwana, H.; Verkuijlen, S.A.M.W.; Adham, M.; Pegtel, M.D.; Greijer, A.E.; Middeldorp, J. Epstein-Barr virus mRNA profiles and viral DNA methylation status in nasopharyngeal brushings from nasopharyngeal carcinoma patients reflect tumor origin. Int. J. Cancer 2017, 140, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Dolcetti, R.; Dal Col, J.; Martorelli, D.; Carbone, A.; Klein, E. Interplay among viral antigens, cellular pathways and tumor microenvironment in the pathogenesis of EBV-driven lymphomas. Semin. Cancer Biol. 2013, 23, 441–456. [Google Scholar] [CrossRef] [PubMed]
- Mui, U.N.; Haley, C.T.; Tyring, S.K. Viral oncology, molecular biology and pathogenesis. J. Clin. Med. 2017, 6, 111. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.K.; Gulley, M.L.; Feng, W.H.; Delecluse, H.J.; Holley-Guthrie, E.; Kenney, S.C. Epstein-Barr Virus lytic infection contributes to lymphoproliferative disease in a SCID mouse model. J. Virol. 2005, 79, 13993–14003. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.D.; Hedge, S.; Young, K.H.; Sullivan, R.; Rajesh, D.; Zhou, Y.; Jankowska-Gan, E.; Burlingham, W.J.; Sun, X.; Gulley, M.L.; et al. A new model of Epstein-Barr virus infection reveals an important role for early lytic viral protein expression in the development of lymphomas. J. Virol. 2011, 85, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.D.; Yu, X.; Mertz, J.E.; Gumperz, J.E.; Reinheim, E.; Zhou, Y.; Tang, W.; Burlingham, W.J.; Gulley, M.L.; Kenney, S.C. An Epstein-Barr virus (EBV) mutant with enhanced BZLF1 expression causes lymphomas with abortive lytic EBV infection in a humanized mouse model. J. Virol. 2012, 86, 7976–7987. [Google Scholar] [CrossRef] [PubMed]
- Ersing, I.; Nobre, L.; Wang, L.W.; Soday, L.; Ma, Y.; Paulo, J.A.; Narita, Y.; Ashbaugh, C.W.; Jiang, C.; Grayson, N.E.; et al. A temporal proteomic map of Epstein-Barr virus lytic replication in B cells. Cell Rep. 2017, 19, 1479–1493. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.E.; Williams, M.V. EBV-dUTPase modulates host immune responses potentially altering the tumor microenvironment in EBV-associated malignancies. J. Curr. Res. HIV/AIDS 2016, 2016, 1–9. [Google Scholar]
- Zhang, Y.; Ohyashiki, J.H.; Takaku, T.; Shinizu, N.; Okhyashiki, K. Transcriptional profiling of Epstein-Barr virus (EBV) genes and host cellular genes in nasal NK/T-cell lymphoma and chronic EBV infection. Br. J. Cancer 2006, 94, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Sommer, P.; Kremmer, E.; Bier, S.; Konig, S.; Zalud, P.; Zeppezauer, M.; Jones, J.F.; Mueller-Lantzsch, N.; Grasser, F.A. Cloning and expression of the Epstein-Barr virus encoded dUTPase: Patients with acute, reactivated or chronic virus infection develop antibodies against the enzyme. J. Gen. Virol. 1996, 77, 2795–2805. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, J.M.; Sommer, P.; Kremmer, E.; Ong, K.S.; Fung, K.; Lee, J.M.F.; Ng, M.H.; Grasser, F.A. A new lytic antibody, 7D6, detects Epstein-Barr virus dUTPase in nonkeratinizing undifferentiated nasopharyngeal carcinomas. Lab. Investig. 1998, 78, 1031–1032. [Google Scholar] [PubMed]
- Young, N.A.; Williams, M.; Jarjour, W.N.; Bruss, M.S.; Bolton, B.; Parikh, S.; Satoskar, A.; Ariza, M.E. Epstein-Barr virus (EBV) encoded dUTPase exacerbates the immune pathology of lupus nephritis in vivo. Int. J. Immunol. Immunother. 2016, 3, 023. [Google Scholar] [CrossRef]
- Fleischmann, J.; Kremmer, E.; Greenspan, J.S.; Grasser, F.A.; Niedobitek, G. Expression of viral and human dUTPase in Epstein-Barr virus-associated diseases. J. Med. Virol. 2002, 68, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.M.; Ariza, M.E.; Williams, M.V.; Jason, L.; Beqaj, S.; Fitzgerald, J.T.; Lemeshow, S.; Glaser, R. Antibody to Epstein-Barr virus deoxyuridine triphosphate nucleotidohydrolase and deoxyribonucleotide polymerase in a Chronic Fatigue Syndrome subset. PLoS ONE 2012, 7, e47891. [Google Scholar] [CrossRef] [PubMed]
- Halpin, P.; Williams, M.V.; Klimas, N.G.; Fletcher, M.A.; Barnes, Z.; Ariza, M.E. Myalgic encephalomyelitis/chronic fatigue syndrome and gulf war illness patients exhibit increased humoral responses to the herpesviruses-encoded dUTPase: Implications in disease pathophysiology. J. Med. Virol. 2017, 89, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.E.; Glaser, R.; Kaumaya, P.T.P.; Jones, C.; Williams, M. The Epstein-Barr Virus (EBV)-encoded dUTPase activates NF-κB through the TLR2 and MyD88-dependent signaling Pathway. J. Immunol. 2009, 182, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Glaser, R.; Litsky, M.L.; Padget, D.A.; Baiocchi, R.A.; Yang, E.V.; Chen, M.; Yeh, P.E.; Green-Church, K.B.; Caligiuri, M.A.; Williams, M.V. The EBV-encoded dUTPase induces immune dysregulation: Implications for the pathophysiology of EBV-associated disease. Virology 2006, 346, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Waldman, W.J.; Williams, M.V.; Lemeshow, S.A.; Binkley, P.; Guttridge, D.; Kiecolt-Glaser, J.K.; Knight, D.A.; Ladner, K.J.; Glaser, R. Epstein-Barr virus encoded dUTPase enhances proinflammatory cytokine production by macrophages in contact with endothelial cells: Evidence for depression-induced atherosclerotic risk. Brain Behav. Immun. 2008, 22, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.E.; Rivailler, P.; Glaser, R.; Chen, M.; Williams, M.V. Epstein-Barr virus encoded dUTPase containing exosomes modulate innate and adaptive immune responses in human dendritic cells and peripheral blood mononuclear cells. PLoS ONE 2013, 8, e69827. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.E.; Glaser, R.; Williams, M.V. Human herpesviruses encoded dUTPases: A family of proteins that modulate dendritic cells function and innate immunity. Front. Microbiol. 2014, 5, 504. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.V.; Cox, B.; Ariza, M.E. Herpesviruses dUTPases: A new family of Pathogen-Associated Molecular Pattern (PAMP) proteins with implications for human disease. Pathogens 2017, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Hao, S.P.; Chang, J.H.; Wu, C.C.; Tsang, N.M.; Lee, Y.; Hsu, C.L.; Ueng, S.H.; Liu, S.C.; Wei, P.C.; et al. Macrophage inflammatory protein-3α is a novel serum marker for nasopharyngeal carcinoma detection and prediction of treatment outcomes. Clin. Cancer Res. 2008, 14, 6979–6987. [Google Scholar] [CrossRef] [PubMed]
- Okudaira, T.; Yamamoto, K.; Kawakami, H.; Uchihara, J.N.; Tomita, M.; Masuda, M.; Matsuda, T.; Sairenji, T.; Iha, H.; Jeang, K.T.; et al. Transactivation of CCL20 gene by Epstein-Barr virus latent membrane protein 1. Br. J. Haematol. 2006, 132, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Baumforth, K.R.; Birgersodder, A.; Reynolds, G.M.; Wei, W.; Kapatai, G.; Flavell, J.R.; Kalk, E.; Piper, K.; Lee, S.; Machado, L.; et al. Expression of the Epstein Barr virus-encoded Epstein-Barr nuclear antigen 1 in Hodgkin’s lymphoma cells mediates up-regulation of CCL20 and the migration of regulatory T cells. Am. J. Pathol. 2008, 173, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Tarbouriech, N.; Buisson, M.; Seigneurin, J.M.; Cusack, S.; Burmeiste, W.P. The monomeric dUTPase from Epstein-Barr virus mimic trimeric dUTPases. Structure 2005, 13, 1299–1310. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.J.; Stow, N.D. New genes from old: Redeployment of dUTPase by herpesviruses. J. Virol. 2005, 79, 12880–12892. [Google Scholar] [CrossRef] [PubMed]
- Kobs-Conrad, S.; Lee, H.; DiGeorge, A.M.; Kaumaya, P.T.P. Engineered topographic Determinants with βα, βαβ and βαβα topologies show high affinity binding to native protein antigen (Lactate Dehydrogenase-C4). J. Biol. Chem. 1993, 268, 25285–25295. [Google Scholar] [PubMed]
- De Sanjose, S.; Bosch, R.; Schouten, T.; Verkuijlen, S.; Nieters, A.; Foretova, L.; Maynadie, M.; Cocco, P.L.; Staines, A.; Becker, N.; et al. Epstein-Barr virus infection and risk of lymphoma: Immunoblot analysis of antibody responses against EBV-related proteins in a large series of lymphoma subjects and matched controls. Int. J. Cancer 2007, 121, 1806–1812. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. EBV based cancer prevention and therapy in nasopharyngeal carcinoma. NPJ Precis. Oncol. 2017, 1–10. [Google Scholar] [CrossRef]
- Tosato, G.; Tanner, J.; Jones, K.D.; Revel, M.; Pike, S.E. Identification of interleukin-6 as an autocrine growth factor for Epstein-Barr virus-immortalized B cells. J. Virol. 1990, 64, 3033–3041. [Google Scholar] [PubMed]
- Scala, G.; Quinto, I.; Ruocco, M.R.; Arcucci, A.; Mallardo, M.; Caretto, P.; Forni, G.; Venuta, S. Expression of an endogenous interleukin 6 gene in human Epstein Barr virus B cell confers growth advantage and in vivo tumorigenicity. J. Exp. Med. 1990, 172, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Durandy, A.; Emilie, D.; Peuchmaur, M.; Forveille, M.; Clement, C.; Wijdenes, J.; Fischer, A. Role of IL-6 in promoting growth of human EBV-induced B-cell tumors in severe combined immunodeficient mice. J. Immunol. 1994, 152, 3561–3567. [Google Scholar]
- Jones, R.J.; Seaman, W.T.; Feng, W.H.; Barlow, E.; Dickerson, S.; Delecluse, H.J.; Kenney, S.C. Roles of lytic viral infection and IL-6 in early versus late passage lymphoblastoid cell lines and EBV-associated lymphoproliferative disease. Int. J. Cancer 2007, 121, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Mashima, R. Physiological roles of MIR-155. Immunology 2015, 145, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.W.; Visser, L.; Tan, L.P.; vanden Berg, A.; Diepstra, A. The microenvironment of Epstein-Barr virus-associated malignancies. Pathogens 2018, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Petrella, T.; Yaziji, N.; Colin, F.; Rifle, G.; Morlevat, F.; Arnould, L.; Fargeot, P.; Depret, O. Implication of the Epstein-Barr virus in the progression of chronic lymphocytic leukaemia/small lymphocytic lymphoma to Hodgkin-like lymphomas. Anticancer Res. 1997, 17, 3907–3913. [Google Scholar] [PubMed]
- Tsimberidou, A.M.; Keating, M.J.; Bueso-Ramos, C.E.; Kurzrock, R. Epstein-Barr virus in patients with chronic lymphocytic leukemia. Leuk. Lymphoma 2006, 47, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Ferrajoli, A.; Ivan, C.; Ciccone, M.; Shimizu, M.; Kita, Y.; Ohtsuka, M.; D’Abundo, L.; Qiang, J.; Lerner, S.; Nouraee, N.; et al. Epstein-Barr virus microRNAs are expressed in patients with chronic lymphocytic leukemia and correlate with overall survival. EBioMedicine 2015, 2, 572–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grywalska, E.; Rolinski, J.; Pasiarski, M.; Korona-Glowniak, I.; Maj, M.; Surdacka, A.; Grafka, A.; Stelmach-Goldys, A.; Zgurski, M.; Gozdz, S.; et al. High viral loads of Epstein-Barr virus DNA in peripheral blood of patients with chronic lymphocytic leukemia associated with unfavorable prognosis. PLoS ONE 2015, 10, e0140178. [Google Scholar] [CrossRef] [PubMed]
- Visco, C.; Falisi, E.; Young, K.H.; Pascarella, M.; Perbellini, O.; Carli, G.; Novella, E.; Rossi, D.; Giaretta, I.; Cavallini, C.; et al. Epstein-Barr virus DNA load in chronic lymphocytic leukemia is an independent predictor of clinical course and survival. Oncotarget 2015, 6, 18653–18663. [Google Scholar] [CrossRef] [PubMed]
- Glaser, R.; Pearson, G.R.; Bonneau, R.H.; Esterling, B.A.; Atkinson, C.; Kiecolt-Glaser, J.K. Stress and the memory T-cell response to the Epstein-Barr virus in healthy medical students. Health Psychol. 1993, 12, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.E.; Simanek, A.M.; Galea, S. Population levels of psychological stress, herpesvirus reactivation and HIV. AIDS Behav. 2010, 14, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.M.; Glaser, R.; Malarkey, W.B.; Beversdorf, D.Q.; Peng, J.; Kiecolt-Glaser, J.K. Inflammation and reactivation of latent herpesviruses in older adults. Brain Behav. Immun. 2012, 26, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Christian, L.M.; Iams, J.D.; Porter, K.; Glaser, R. Epstein-Barr virus reactivation during pregnancy and postpartum: Effects of race and racial discrimination. Brain Behav. Immun. 2012, 26, 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- Laichalk, L.; Thorley-Lawson, D.A. Terminal differentiation into plasma cells initiates the replicative cycle of Epstein-Barr virus in vivo. J. Virol. 2005, 79, 1296–1307. [Google Scholar] [CrossRef] [PubMed]
- Al Tabaa, Y.; Tuaillon, E.; Bollore, K.; Foulongne, V.; Petitjean, G.; Seigneurin, J.M.; Duperray, C.; Desgranges, C.; Vendrell, J.P.; et al. Functional Epstein-Barr virus reservoir in plasma cells derived from infected peripheral blood memory B cells. Blood 2009, 113, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Al Tabaa, Y.; Tuaillon, E.; Jeziorski, E.; Ouedraogo, D.E.; Bolloré, K.; Rubbo, P.A.; Foulongne, V.; Rodiere, M.; Vendrell, J.P. B-cell polyclonal activation and Epstein-Barr viral abortive lytic cycle are two key features in acute infectious mononucleosis. J. Clin. Virol. 2011, 52, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.E.; Williams, M.V. A Human Endogenous Retrovirus K dUTPase Triggers a TH1, TH17 Cytokine Response: Does it play a role in psoriasis? J. Investig. Dermatol. 2011, 131, 2419–2428. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, E.S.; Harris, N.L.; Stein, H.; Vardiman, J. Pathology and Genetics of Human Tumors of Haematopoietic and Lymphoid Tissues; IARC Press: Lyon, France, 2001. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Treatments (10 µg/mL) | IL-6 (pg/mL) | IL-1β (pg/mL) | TNFα (pg/mL) | IL-8 (pg/mL) | IL-10 (pg/mL) |
|---|---|---|---|---|---|
| Untreated | 5 ± 0.8 | 31 ± 22.8 | 5 ± 4.9 | 407 ± 5.7 | 8 ± 3.8 |
| EBV-dUTPase | 9570 ± 5.7 | 978 ± 15 | 379 ± 123 | 35,039 ± 219 | 311 ± 35 |
| Scrambled peptide L83-K103 | 369 ± 312 | 34 ± 24.4 | 9 ± 6.2 | 8738 ± 267.6 | 9 ± 5.2 |
| EBV-dUTPase peptide L83-K103 | 3272 ± 6 | 111 ± 21.5 | 45 ± 6.6 | 21,934 ± 14 | 43 ± 6.2 |
| dUTPases | Amino Acid Sequence |
|---|---|
| EBV | 81GELRLILQNQ90 |
| HHV-8 | 109GEIQVILLNK118 |
| HSV-1/2 | 102GTVMAVVAP110 |
| VZV | 130GVISALLYYR139 |
| HHV-6A | 207TDISVFLMNL116 |
| HHV-7 | 215NVISISLINL224 |
| HCMV | 173LQVPQLDVVNL183 |
| Human | 84GNVGVVLFNF93 |
| Clinical Status | Gender | % Positive EBV dUTPase Ab a | % Abnormal Reactive Ab Pattern to EBV (ARP_EBV) b | % ARP_EBV & dUTPase Seropositive c |
|---|---|---|---|---|
| Controls | Females | 11.67 (32/268) | 22.22 (10/45) | 8.88 (4/45) |
| Males | 13.98 (23/163) | 11.36 (5/44) | 4.54 (2/44) | |
| DLBCL | Females | 42.86 (9/21) | 33.33 (7/21) | 14.28 (3/21) |
| Males | 26.66 (4/15) | 20.00 (3/15) | 0.00 (0/15) | |
| CLL | Females | 34.37 (11/32) | 53.12 (17/32) | 18.75 (6/32) |
| Males | 44.12 (15/34) | 20.59 (7/34) | 14.70 (5/34) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, M.; Ariza, M.E. EBV Positive Diffuse Large B Cell Lymphoma and Chronic Lymphocytic Leukemia Patients Exhibit Increased Anti-dUTPase Antibodies. Cancers 2018, 10, 129. https://doi.org/10.3390/cancers10050129
Williams M, Ariza ME. EBV Positive Diffuse Large B Cell Lymphoma and Chronic Lymphocytic Leukemia Patients Exhibit Increased Anti-dUTPase Antibodies. Cancers. 2018; 10(5):129. https://doi.org/10.3390/cancers10050129
Chicago/Turabian StyleWilliams, Marshall, and Maria Eugenia Ariza. 2018. "EBV Positive Diffuse Large B Cell Lymphoma and Chronic Lymphocytic Leukemia Patients Exhibit Increased Anti-dUTPase Antibodies" Cancers 10, no. 5: 129. https://doi.org/10.3390/cancers10050129
APA StyleWilliams, M., & Ariza, M. E. (2018). EBV Positive Diffuse Large B Cell Lymphoma and Chronic Lymphocytic Leukemia Patients Exhibit Increased Anti-dUTPase Antibodies. Cancers, 10(5), 129. https://doi.org/10.3390/cancers10050129