Extracellular Vesicle microRNAs Contribute to the Osteogenic Inhibition of Mesenchymal Stem Cells in Multiple Myeloma


 , , and
, , and 
Abstract
:
1. Introduction
2. Results
2.1. MM EVs Are Internalized by hMSCs and Inhibit Their Osteogenic Differentiation
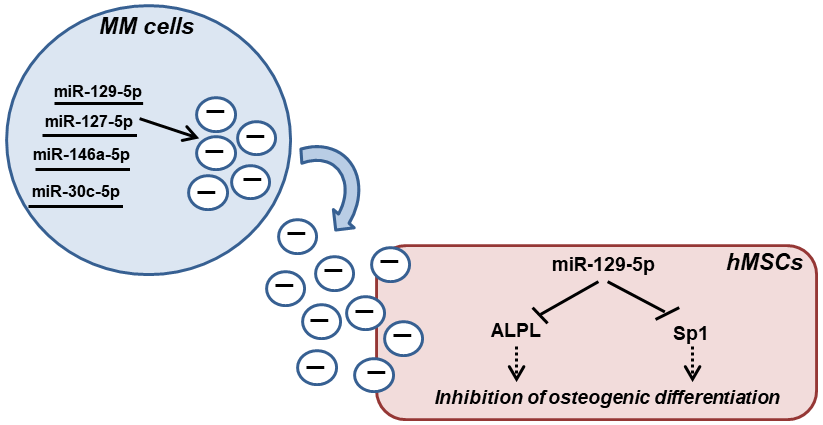
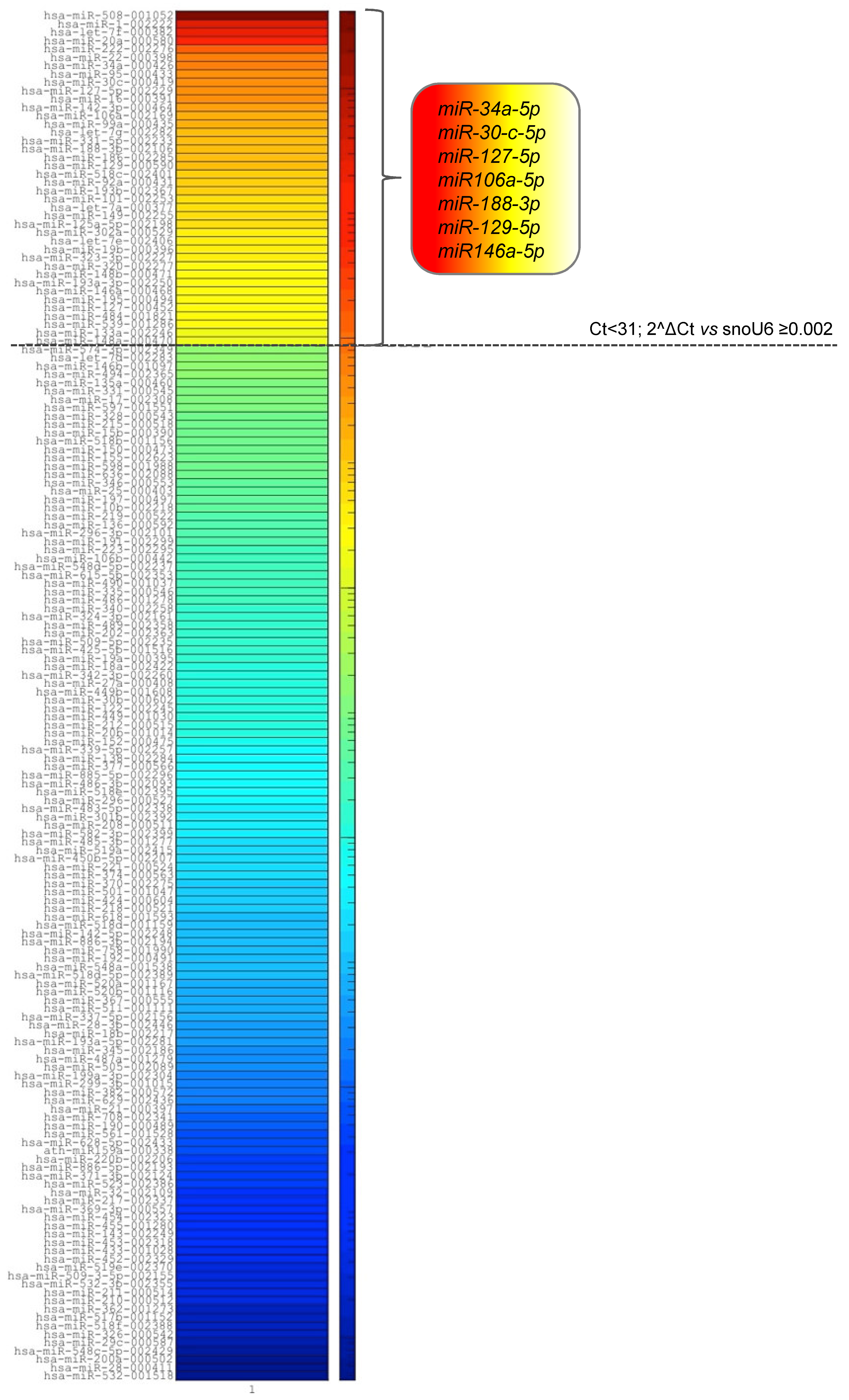
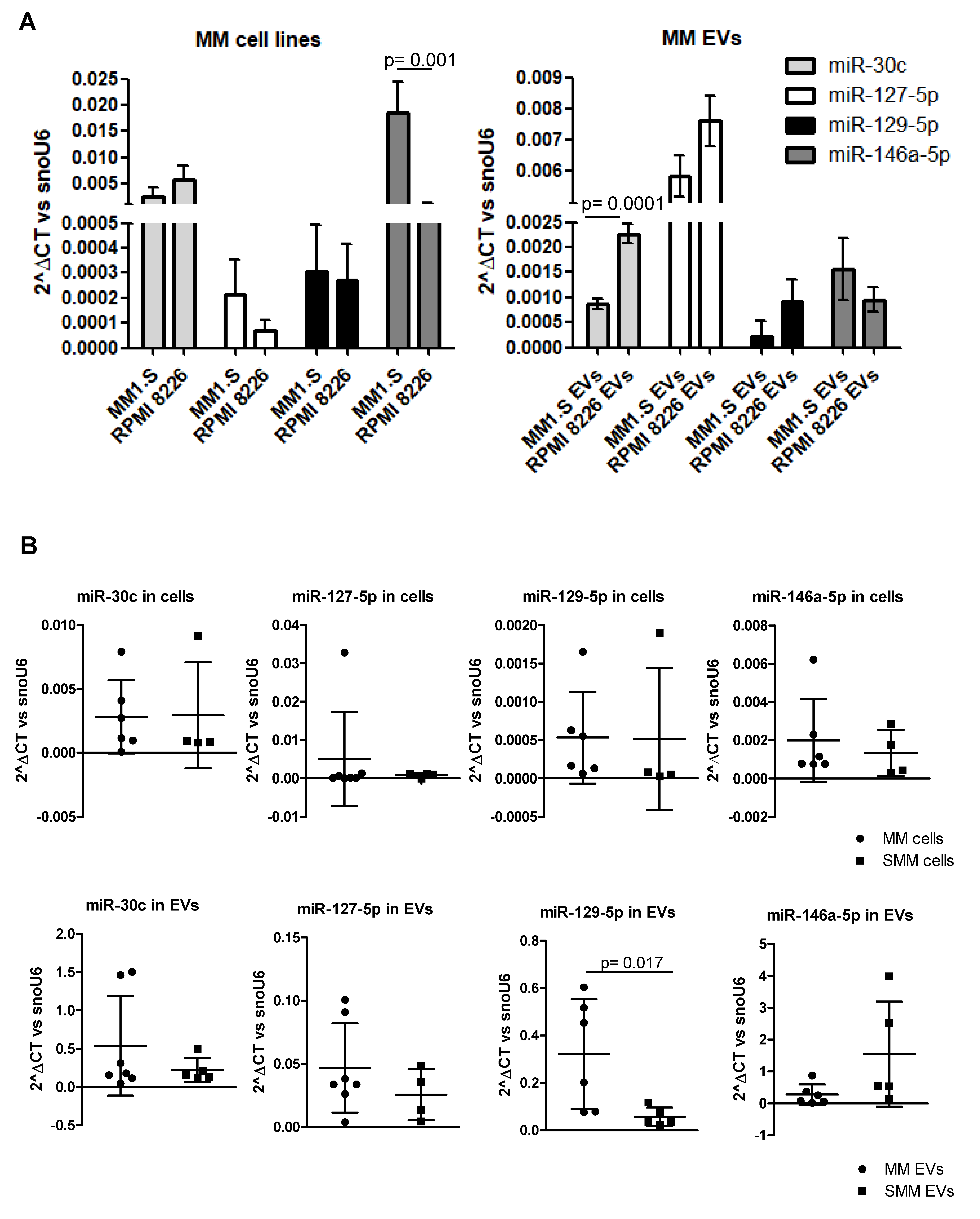
2.2. MM-EVs Contain miRNAs Involved in hMSCsOsteogenic Inhibition
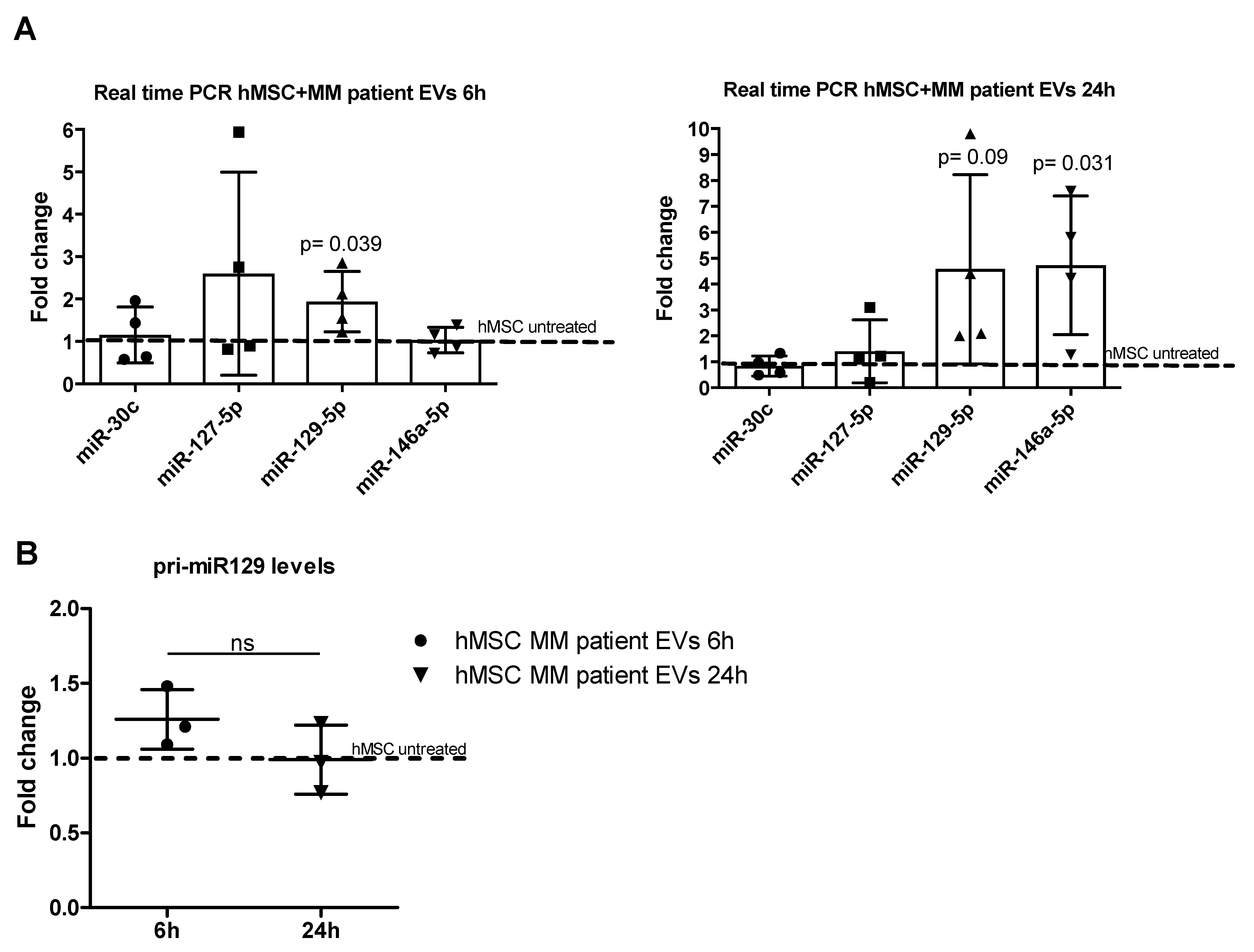
2.3. MiR-129-5p Level Increases in hMSCs afterEV Incubation
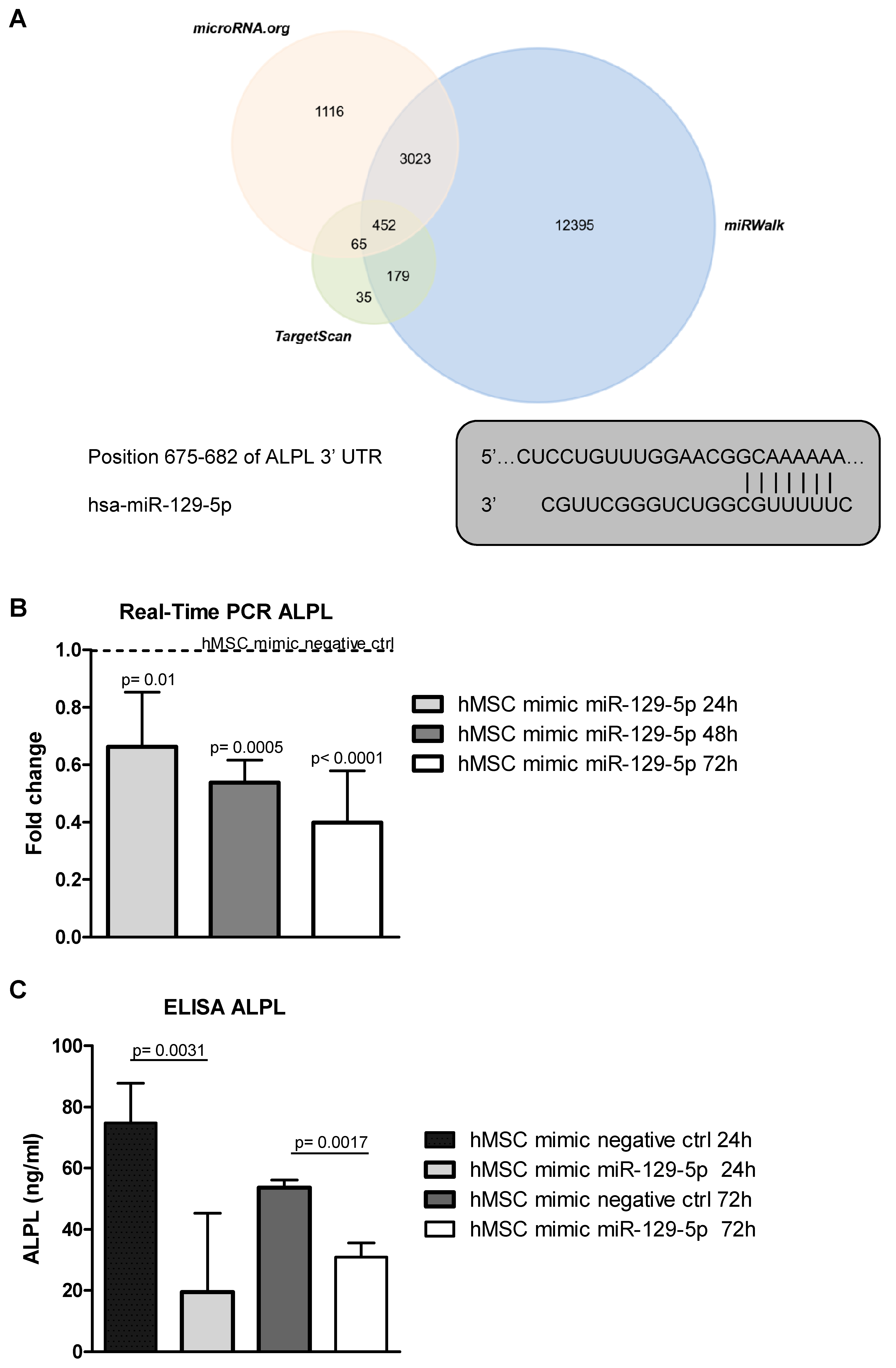
2.4. MiR-129-5p Inhibits ALPL Expression in hMSCs
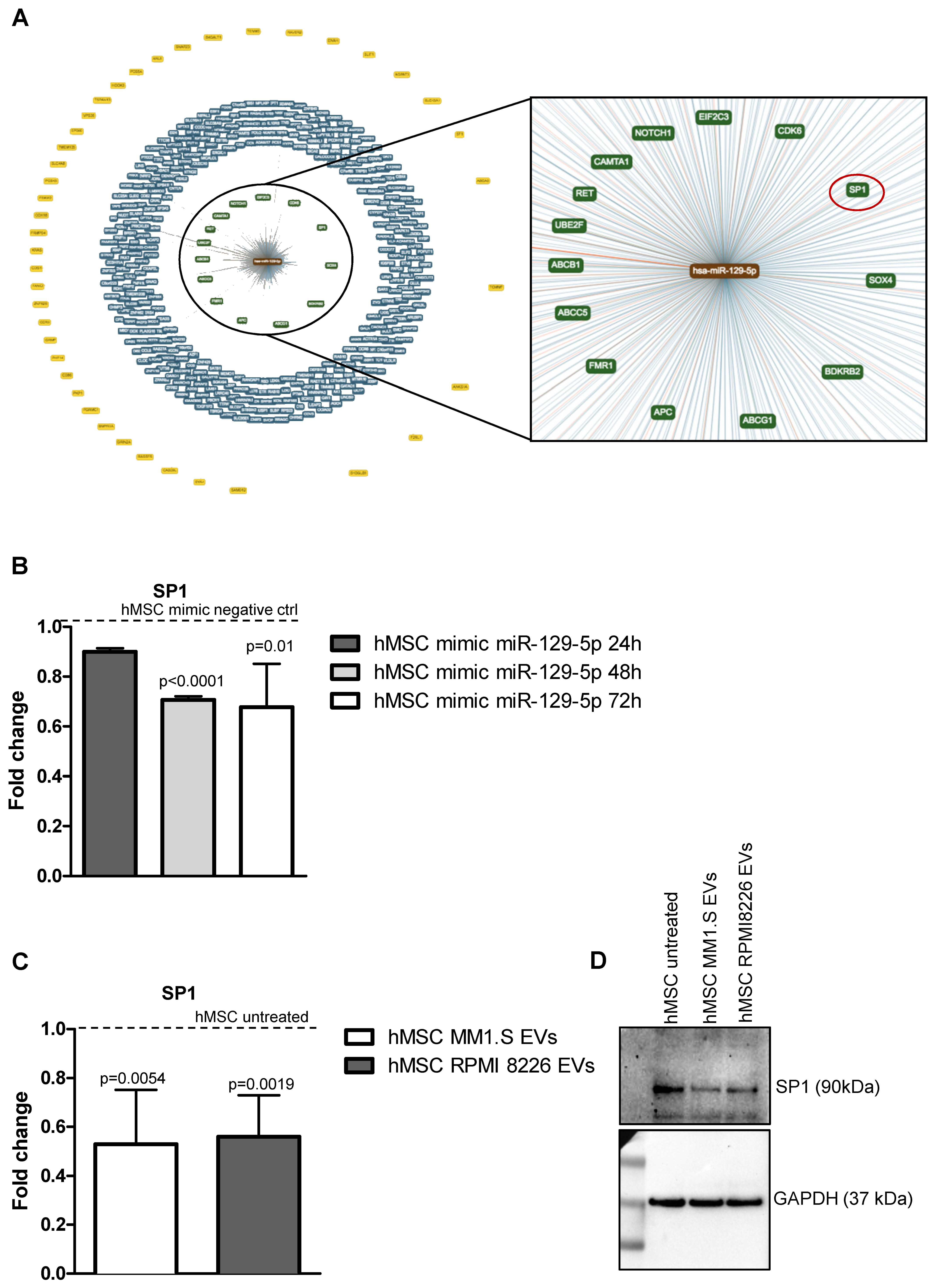
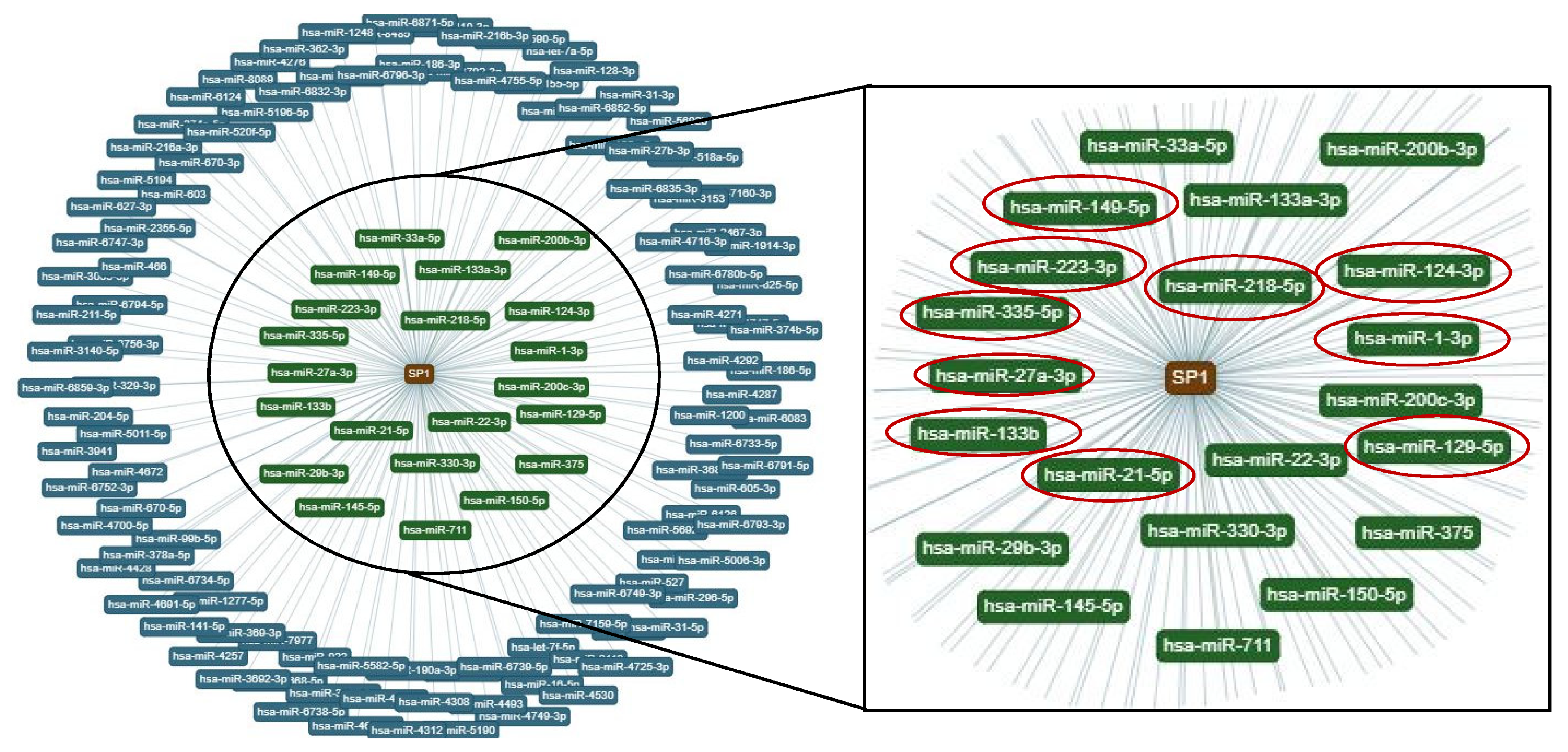
2.5. MiR-129-5p and MM EVs Inhibit the Transcription Factor SP1 Expression in hMSCs
3. Discussions
4. Materials and Methods
4.1. Cell Cultures
4.2. Purification of Primary Plasma Cells
4.3. Extracellular Vesicle Isolation
4.4. Internalization of MM-Derived EVs by hMSCs
4.5. hMSCs Treatment with EVs
4.6. MiRNA Expression Profiling
4.7. Cell Transfection
4.8. RNA Isolation and Real-Time PCR
4.9. ELISA ALPL
4.10. Western Blotting
4.11. Bioinformatics Tools
4.12. Statistical Analysis
4.13. Ethical statement:
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kazandjian, D. Multiple myeloma epidemiology and survival: A unique malignancy. Semin. Oncol. 2016, 43, 676–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateos, M.V.; Landgren, O. Mgus and smoldering multiple myeloma: Diagnosis and epidemiology. Cancer Treat. Res. 2016, 169, 3–12. [Google Scholar] [PubMed]
- Kyle, R.A.; Gertz, M.A.; Witzig, T.E.; Lust, J.A.; Lacy, M.Q.; Dispenzieri, A.; Fonseca, R.; Rajkumar, S.V.; Offord, J.R.; Larson, D.R.; et al. Review of 1027 patients with newly diagnosed multiple myeloma. Mayo Clin. Proc. 2003, 78, 21–33. [Google Scholar] [CrossRef]
- Terpos, E.; Ntanasis-Stathopoulos, I.; Gavriatopoulou, M.; Dimopoulos, M.A. Pathogenesis of bone disease in multiple myeloma: From bench to bedside. Blood Cancer J. 2018, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Brady, J.J.; Dowling, P.; Clynes, M.; O’Gorman, P. Bone disease in multiple myeloma: Pathophysiology and management. Cancer Growth Metastasis 2014, 7, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Noll, J.E.; Williams, S.A.; Tong, C.M.; Wang, H.; Quach, J.M.; Purton, L.E.; Pilkington, K.; To, L.B.; Evdokiou, A.; Gronthos, S.; et al. Myeloma plasma cells alter the bone marrow microenvironment by stimulating the proliferation of mesenchymal stromal cells. Haematologica 2014, 99, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, S.; Li, J.; Zhao, S.; Yi, Z. Roles for mirnas in osteogenic differentiation of bone marrow mesenchymal stem cells. Stem. Cell Res. Ther. 2019, 10, 197. [Google Scholar] [CrossRef]
- Arfat, Y.; Xiao, W.Z.; Ahmad, M.; Zhao, F.; Li, D.J.; Sun, Y.L.; Hu, L.; Zhihao, C.; Zhang, G.; Iftikhar, S.; et al. Role of micrornas in osteoblasts differentiation and bone disorders. Curr. Med. Chem. 2015, 22, 748–758. [Google Scholar] [CrossRef]
- Peng, S.; Gao, D.; Gao, C.; Wei, P.; Niu, M.; Shuai, C. Micrornas regulate signaling pathways in osteogenic differentiation of mesenchymal stem cells (review). Mol. Med. Rep. 2016, 14, 623–629. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (misev2018): A position statement of the international society for extracellular vesicles and update of the misev2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Moloudizargari, M.; Abdollahi, M.; Asghari, M.H.; Zimta, A.A.; Neagoe, I.B.; Nabavi, S.M. The emerging role of exosomes in multiple myeloma. Blood Rev. 2019, 38, 100595. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Giannandrea, D.; Lesma, E.; Basile, A.; Chiaramonte, R. Extracellular vesicles enhance multiple myeloma metastatic dissemination. Int. J. Mol. Sci. 2019, 20, 3236. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; De Veirman, K.; Faict, S.; Frassanito, M.A.; Ribatti, D.; Vacca, A.; Menu, E. Multiple myeloma exosomes establish a favourable bone marrow microenvironment with enhanced angiogenesis and immunosuppression. J. Pathol. 2016, 239, 162–173. [Google Scholar] [CrossRef]
- Zarfati, M.; Avivi, I.; Brenner, B.; Katz, T.; Aharon, A. Extracellular vesicles of multiple myeloma cells utilize the proteasome inhibitor mechanism to moderate endothelial angiogenesis. Angiogenesis 2019, 22, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, X.J.; Zeng, C.; Wu, P.H.; Wang, H.X.; Chen, Z.C.; Li, Q.B. Microvesicles secreted from human multiple myeloma cells promote angiogenesis. ActaPharmacol. Sin. 2014, 35, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Hong, J.; Hong, M.; Wang, Y.; Yu, T.; Zang, S.; Wu, Q. Pirna-823 delivered by multiple myeloma-derived extracellular vesicles promoted tumorigenesis through re-educating endothelial cells in the tumor environment. Oncogene 2019, 38, 5227–5238. [Google Scholar] [CrossRef] [PubMed]
- Arendt, B.K.; Walters, D.K.; Wu, X.; Tschumper, R.C.; Jelinek, D.F. Multiple myeloma dell-derived microvesicles are enriched in cd147 expression and enhance tumor cell proliferation. Oncotarget 2014, 5, 5686–5699. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; De Veirman, K.; De Beule, N.; Maes, K.; De Bruyne, E.; Van Valckenborgh, E.; Vanderkerken, K.; Menu, E. The bone marrow microenvironment enhances multiple myeloma progression by exosome-mediated activation of myeloid-derived suppressor cells. Oncotarget 2015, 6, 43992–44004. [Google Scholar] [CrossRef] [Green Version]
- Pourhanifeh, M.H.; Mahjoubin-Tehran, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; Mirzaei, H.; Asemi, Z. Micrornas and exosomes: Small molecules with big actions in multiple myeloma pathogenesis. IUBMB Life 2019. [Google Scholar] [CrossRef]
- Raimondo, S.; Saieva, L.; Vicario, E.; Pucci, M.; Toscani, D.; Manno, M.; Raccosta, S.; Giuliani, N.; Alessandro, R. Multiple myeloma-derived exosomes are enriched of amphiregulin (areg) and activate the epidermal growth factor pathway in the bone microenvironment leading to osteoclastogenesis. J. Hematol. Oncol. 2019, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, L.; De Luca, A.; Amodio, N.; Manno, M.; Raccosta, S.; Taverna, S.; Bellavia, D.; Naselli, F.; Fontana, S.; Schillaci, O.; et al. Involvement of multiple myeloma cell-derived exosomes in osteoclast differentiation. Oncotarget 2015, 6, 13772–13789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faict, S.; Muller, J.; De Veirman, K.; De Bruyne, E.; Maes, K.; Vrancken, L.; Heusschen, R.; De Raeve, H.; Schots, R.; Vanderkerken, K.; et al. Exosomes play a role in multiple myeloma bone disease and tumor development by targeting osteoclasts and osteoblasts. Blood Cancer J. 2018, 8, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Liu, H.; Li, Y.; Shao, Q.; Chen, J.; Song, J.; Fu, R. Multiple myeloma-derived exosomes inhibit osteoblastic differentiation and improve il-6 secretion of bmscs from multiple myeloma. J. Investig. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lei, Q.; Wang, H.; Xu, C.; Liu, T.; Kong, F.; Yang, C.; Yan, G.; Sun, L.; Zhao, A.; et al. Tumor-derived extracellular vesicles inhibit osteogenesis and exacerbate myeloma bone disease. Theranostics 2019, 9, 196–209. [Google Scholar] [CrossRef]
- Zhang, L.; Li, G.; Wang, K.; Wang, Y.; Dong, J.; Wang, H.; Xu, L.; Shi, F.; Cao, X.; Hu, Z.; et al. Mir-30 family members inhibit osteoblast differentiation by suppressing runx2 under unloading conditions in mc3t3-e1 cells. Biochem. Biophys. Res. Commun. 2020, 522, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Xu, H.; Han, H.; Song, S.; Zhang, X.; Ouyang, L.; Qian, C.; Hong, Y.; Qiu, Y.; Zhou, W.; et al. Exosome-mediated transfer of lncrunx2-as1 from multiple myeloma cells to mscs contributes to osteogenesis. Oncogene 2018, 37, 5508–5519. [Google Scholar] [CrossRef]
- Pinzone, J.J.; Hall, B.M.; Thudi, N.K.; Vonau, M.; Qiang, Y.W.; Rosol, T.J.; Shaughnessy, J.D., Jr. The role of dickkopf-1 in bone development, homeostasis, and disease. Blood 2009, 113, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Deniaud, E.; Baguet, J.; Mathieu, A.L.; Pages, G.; Marvel, J.; Leverrier, Y. Overexpression of sp1 transcription factor induces apoptosis. Oncogene 2006, 25, 7096–7105. [Google Scholar] [CrossRef] [Green Version]
- Haidweger, E.; Novy, M.; Rotheneder, H. Modulation of sp1 activity by a cyclin a/cdk complex. J. Mol. Biol. 2001, 306, 201–212. [Google Scholar] [CrossRef]
- Li, L.; Davie, J.R. The role of sp1 and sp3 in normal and cancer cell biology. Ann. Anat. 2010, 192, 275–283. [Google Scholar] [CrossRef]
- Zhang, X.; Li, R.; Qin, X.; Wang, L.; Xiao, J.; Song, Y.; Sheng, X.; Guo, M.; Ji, X. Sp1 plays an important role in vascular calcification both in vivo and in vitro. J. Am. Heart Assoc. 2018, 7, e007555. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yerges-Armstrong, L.M.; Chu, Y.; Zmuda, J.M.; Zhang, Y. Transcriptional regulation of frizzled-1 in human osteoblasts by sp1. PLoS ONE 2016, 11, e0163277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Xu, B.; Xu, H. Tgf-beta1 promoted chondrocyte proliferation by regulating sp1 through msc-exosomes derived mir-135b. Cell Cycle 2018, 17, 2756–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Holmstrom, K.; Qiu, W.; Ditzel, N.; Shi, K.; Hokland, L.; Kassem, M. Microrna-34a inhibits osteoblast differentiation and in vivo bone formation of human stromal stem cells. Stem Cells 2014, 32, 902–912. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Yang, K.; Xiao, L.; Guo, L.; Guo, C.; Yan, Y.; Qi, J.; Wang, F.; Ryffel, B.; Li, C.; et al. Osteoblast hypoxia-inducible factor-1alpha pathway activation restrains osteoclastogenesis via the interleukin-33-microrna-34a-notch1 pathway. Front. Immunol. 2017, 8, 1312. [Google Scholar] [CrossRef] [Green Version]
- Belaya, Z.E.; Grebennikova, T.A.; Melnichenko, G.A.; Nikitin, A.G.; Solodovnikov, A.G.; Brovkina, O.I.; Grigoriev, A.U.; Rozhinskaya, L.Y.; Dedov, I.I. Effects of endogenous hypercortisolism on bone mrna and microrna expression in humans. Osteoporos. Int. 2018, 29, 211–221. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, R.L.; Croce, C.M.; Stein, J.L.; Lian, J.B.; van Wijnen, A.J.; Stein, G.S. A program of micrornas controls osteogenic lineage progression by targeting transcription factor runx2. Proc. Natl. Acad. Sci. USA 2011, 108, 9863–9868. [Google Scholar] [CrossRef] [Green Version]
- Tu, M.; Li, Y.; Zeng, C.; Deng, Z.; Gao, S.; Xiao, W.; Luo, W.; Jiang, W.; Li, L.; Lei, G. Microrna-127-5p regulates osteopontin expression and osteopontin-mediated proliferation of human chondrocytes. Sci. Rep. 2016, 6, 25032. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Xu, L.; Zhou, F.; Liu, A.M.; Ge, H.X.; Chen, Y.Y.; Tu, M. Malat1/mir-127-5p regulates osteopontin (opn)-mediated proliferation of human chondrocytes through pi3k/akt pathway. J. Cell. Biochem. 2018, 119, 431–439. [Google Scholar] [CrossRef]
- Manochantr, S.; Marupanthorn, K.; Tantrawatpan, C.; Kheolamai, P.; Tantikanlayaporn, D.; Sanguanjit, P. The effects of bmp-2, mir-31, mir-106a, and mir-148a on osteogenic differentiation of mscs derived from amnion in comparison with mscs derived from the bone marrow. Stem Cells Int. 2017, 2017, 7257628. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Pan, Y.; Guan, P.; Li, X.; You, C. Bioinformatics analysis of col1a1 regulated by mir-129-5p as a potential therapeutic target for gastric cancer. Nan Fang Yi Ke Da XueXueBao 2019, 39, 540–546. [Google Scholar]
- Chen, Y.; Ou, Y.; Dong, J.; Yang, G.; Zeng, Z.; Liu, Y.; Liu, B.; Li, W.; He, X.; Lan, T. Osteopontin promotes collagen i synthesis in hepatic stellate cells by mirna-129-5p inhibition. Exp. Cell Res. 2018, 362, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yu, J. Mir-129-5p suppresses gastric cancer cell invasion and proliferation by inhibiting col1a1. Biochem. Cell Biol. 2018, 96, 19–25. [Google Scholar] [CrossRef]
- Musolino, C.; Oteri, G.; Allegra, A.; Mania, M.; D’Ascola, A.; Avenoso, A.; Innao, V.; Allegra, A.G.; Campo, S. Altered microrna expression profile in the peripheral lymphoid compartment of multiple myeloma patients with bisphosphonate-induced osteonecrosis of the jaw. Ann.Hematol. 2018, 97, 1259–1269. [Google Scholar] [CrossRef]
- Zhao, J.; Huang, M.; Zhang, X.; Xu, J.; Hu, G.; Zhao, X.; Cui, P. Mir-146a deletion protects from bone loss in ovx mice by suppressing rankl/opg and m-csf in bone microenvironment. J. Bone Miner. Res. 2019, 34, 2149–2161. [Google Scholar] [CrossRef]
- Ye, X.; Wang, X.B.; Wang, J.; Ming, J. Effect of microrna-146a on differentiation potential of human bone marrow mesenchymal stem cells. Zhongguo Shi Yan Xue Ye XueZaZhi 2016, 24, 596–601. [Google Scholar]
- De Veirman, K.; Wang, J.; Xu, S.; Leleu, X.; Himpe, E.; Maes, K.; De Bruyne, E.; Van Valckenborgh, E.; Vanderkerken, K.; Menu, E.; et al. Induction of mir-146a by multiple myeloma cells in mesenchymal stromal cells stimulates their pro-tumoral activity. Cancer Lett. 2016, 377, 17–24. [Google Scholar] [CrossRef]
- Hamberg, M.; Backes, C.; Fehlmann, T.; Hart, M.; Meder, B.; Meese, E.; Keller, A. Mirtargetlink-mirnas, genes and interaction networks. Int. J. Mol. Sci. 2016, 17, 564. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.W.; Rhee, D.K.; Paton, J.C.; Pyo, S. Pneumolysin-induced autophagy contributes to inhibition of osteoblast differentiation through downregulation of sp1 in human osteosarcoma cells. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2663–2673. [Google Scholar] [CrossRef]
- Yun-Feng, W.; Matsuo, N.; Sumiyoshi, H.; Yoshioka, H. Sp7/osterix up-regulates the mouse pro-alpha3(v) collagen gene (col5a3) during the osteoblast differentiation. Biochem. Biophys. Res. Commun. 2010, 394, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qiu, X.; Sun, Y.; Zhang, N.; Wang, L. Sp1-stimulated mir-545-3p inhibits osteogenesis via targeting lrp5-activated wnt/beta-catenin signaling. Biochem. Biophys. Res. Commun. 2019, 517, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Niger, C.; Lima, F.; Yoo, D.J.; Gupta, R.R.; Buo, A.M.; Hebert, C.; Stains, J.P. The transcriptional activity of osterix requires the recruitment of sp1 to the osteocalcin proximal promoter. Bone 2011, 49, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, S.; Yan, Q.; Chen, X.; Yang, Y.; Liu, X.; Wan, X. Interferon-beta induced microrna-129-5p down-regulates hpv-18 e6 and e7 viral gene expression by targeting sp1 in cervical cancer cells. PLoS ONE 2013, 8, e81366. [Google Scholar]
- Wang, W.; Yang, C.; Wang, X.Y.; Zhou, L.Y.; Lao, G.J.; Liu, D.; Wang, C.; Hu, M.D.; Zeng, T.T.; Yan, L.; et al. Microrna-129 and -335 promote diabetic wound healing by inhibiting sp1-mediated mmp-9 expression. Diabetes 2018, 67, 1627–1638. [Google Scholar] [CrossRef] [Green Version]
- Sobacchi, C.; Palagano, E.; Villa, A.; Menale, C. Soluble factors on stage to direct mesenchymal stem cells fate. Front. Bioeng. Biotechnol. 2017, 5, 32. [Google Scholar] [CrossRef]
- Roodman, G.D. Osteoblast function in myeloma. Bone 2011, 48, 135–140. [Google Scholar] [CrossRef]
- Kassen, D.; Moore, S.; Percy, L.; Herledan, G.; Bounds, D.; Rodriguez-Justo, M.; Croucher, P.; Yong, K. The bone marrow stromal compartment in multiple myeloma patients retains capability for osteogenic differentiation in vitro: Defining the stromal defect in myeloma. Br. J. Haematol. 2014, 167, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Adamik, J.; Galson, D.L.; Roodman, G.D. Osteoblast suppression in multiple myeloma bone disease. J. Bone Oncol. 2018, 13, 62–70. [Google Scholar] [CrossRef] [PubMed]
- De Matteo, M.; Brunetti, A.E.; Maiorano, E.; Cafforio, P.; Dammacco, F.; Silvestris, F. Constitutive down-regulation of osterix in osteoblasts from myeloma patients: In vitro effect of bortezomib and lenalidomide. Leuk. Res. 2010, 34, 243–249. [Google Scholar] [CrossRef]
- Bereziat, V.; Mazurier, C.; Auclair, M.; Ferrand, N.; Jolly, S.; Marie, T.; Kobari, L.; Toillon, I.; Delhommeau, F.; Feve, B.; et al. Systemic dysfunction of osteoblast differentiation in adipose-derived stem cells from patients with multiple myeloma. Cells 2019, 8, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toscani, D.; Bolzoni, M.; Accardi, F.; Aversa, F.; Giuliani, N. The osteoblastic niche in the context of multiple myeloma. Ann. N. Y. Acad. Sci. 2015, 1335, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Pan, Y.; Li, W.; Sun, C.; Liu, J.; Xu, T.; Shu, Y. Extracellular vesicles-mediated noncoding rnas transfer in cancer. J. Hematol. Oncol. 2017, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; Mancone, C.; Tarallo, R.; Battistelli, C.; Alonzi, T.; Weisz, A.; Tripodi, M. The rna-binding protein syncrip is a component of the hepatocyte exosomal machinery controlling microrna sorting. Cell Rep. 2016, 17, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, A.; Bazou, D.; O’Gorman, P. Smoldering multiple myeloma: Prevalence and current evidence guiding treatment decisions. Blood Lymphat. Cancer 2018, 8, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silbermann, R.; Roodman, G.D. Myeloma bone disease: Pathophysiology and management. J. Bone Oncol. 2013, 2, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Zangari, M.; Esseltine, D.; Lee, C.K.; Barlogie, B.; Elice, F.; Burns, M.J.; Kang, S.H.; Yaccoby, S.; Najarian, K.; Richardson, P.; et al. Response to bortezomib is associated to osteoblastic activation in patients with multiple myeloma. Br. J. Haematol. 2005, 131, 71–73. [Google Scholar] [CrossRef]
- Takeshita, H.; Kusuzaki, K.; Murata, H.; Suginoshita, T.; Hirata, M.; Hashiguchi, S.; Ashihara, T.; Gebhardt, M.C.; Mankin, H.J.; Hirasawa, Y. Osteoblastic differentiation and p-glycoprotein multidrug resistance in a murine osteosarcoma model. Br. J. Cancer 2000, 82, 1327–1331. [Google Scholar] [CrossRef]
- Mourskaia, A.A.; Amir, E.; Dong, Z.; Tiedemann, K.; Cory, S.; Omeroglu, A.; Bertos, N.; Ouellet, V.; Clemons, M.; Scheffer, G.L.; et al. Abcc5 supports osteoclast formation and promotes breast cancer metastasis to bone. Breast Cancer Res. 2012, 14, R149. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; Zhou, Y.; Xiao, Y. The regulatory roles of notch in osteocyte differentiation via the crosstalk with canonical wnt pathways during the transition of osteoblasts to osteocytes. Bone 2018, 108, 165–178. [Google Scholar] [CrossRef]
- Canalis, E.; Adams, D.J.; Boskey, A.; Parker, K.; Kranz, L.; Zanotti, S. Notch signaling in osteocytes differentially regulates cancellous and cortical bone remodeling. J. Biol. Chem. 2013, 288, 25614–25625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hassan, M.Q.; Xie, R.L.; Hawse, J.R.; Spelsberg, T.C.; Montecino, M.; Stein, J.L.; Lian, J.B.; van Wijnen, A.J.; Stein, G.S. Co-stimulation of the bone-related runx2 p1 promoter in mesenchymal cells by sp1 and ets transcription factors at polymorphic purine-rich DNA sequences (y-repeats). J. Biol. Chem. 2009, 284, 3125–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, F.; Yang, S.; Guo, Q.; Zhang, X.; Ren, D.; Lv, T.; Xu, X. Microrna-21 regulates osteogenic differentiation of periodontal ligament stem cells by targeting smad5. Sci. Rep. 2017, 7, 16608. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, M.; Liu, X.; Liu, F.; Zhu, J. Mir-27a-3p promotes the malignant phenotypes of osteosarcoma by targeting ten-eleven translocation 1. Int. J. Oncol. 2018, 52, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Zhou, S.; Ye, H.; Jiang, S.; Yu, K.; Ma, Y. The mechanism of sp1/p300 complex promotes proliferation of multiple myeloma cells through regulating iqgap1 transcription. Biomed. Pharmacother. 2019, 119, 109434. [Google Scholar] [CrossRef]
- Fulciniti, M.; Amin, S.; Nanjappa, P.; Rodig, S.; Prabhala, R.; Li, C.; Minvielle, S.; Tai, Y.T.; Tassone, P.; Avet-Loiseau, H.; et al. Significant biological role of sp1 transactivation in multiple myeloma. Clin. Cancer Res. 2011, 17, 6500–6509. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microrna target sites in mammalian mrnas. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. Mirwalk: An online resource for prediction of microrna binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microrna.Org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [Green Version]
- Pathan, M.; Keerthikumar, S.; Ang, C.S.; Gangoda, L.; Quek, C.Y.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. Funrich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| B-Actin | TCCCTTGCCATCCTAAAAAGCCACCC | CTGGGCCATTCTTCCTTAGAGAGAAG |
| OSX | TGCTTGAGGAGGAAGTTCAC | AGGTCACTGCCCACAGAGTA |
| ALPL | CCACGTCTTCACATTTGGTG | AGACTGCGCCTGGTAGTTGT |
| COL1A1 | TGTGGATGCCTCTTGGGTATC | TTTTGGCCATCTCTTCCTTCA |
| OCN | AGCAAAGGTGCAGCCTTTGT | GCGCCTGGGTCTCTTCACT |
| SP1 | GCCTCCAGACCATTAACCTCAGT | GCTCCATGATCACCTGGGGCAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raimondo, S.; Urzì, O.; Conigliaro, A.; Lo Bosco, G.; Parisi, S.; Carlisi, M.; Siragusa, S.; Raimondi, L.; De Luca, A.; Giavaresi, G.; et al. Extracellular Vesicle microRNAs Contribute to the Osteogenic Inhibition of Mesenchymal Stem Cells in Multiple Myeloma. Cancers 2020, 12, 449. https://doi.org/10.3390/cancers12020449
Raimondo S, Urzì O, Conigliaro A, Lo Bosco G, Parisi S, Carlisi M, Siragusa S, Raimondi L, De Luca A, Giavaresi G, et al. Extracellular Vesicle microRNAs Contribute to the Osteogenic Inhibition of Mesenchymal Stem Cells in Multiple Myeloma. Cancers. 2020; 12(2):449. https://doi.org/10.3390/cancers12020449
Chicago/Turabian StyleRaimondo, Stefania, Ornella Urzì, Alice Conigliaro, Giosuè Lo Bosco, Sofia Parisi, Melania Carlisi, Sergio Siragusa, Lavinia Raimondi, Angela De Luca, Gianluca Giavaresi, and et al. 2020. "Extracellular Vesicle microRNAs Contribute to the Osteogenic Inhibition of Mesenchymal Stem Cells in Multiple Myeloma" Cancers 12, no. 2: 449. https://doi.org/10.3390/cancers12020449
APA StyleRaimondo, S., Urzì, O., Conigliaro, A., Lo Bosco, G., Parisi, S., Carlisi, M., Siragusa, S., Raimondi, L., De Luca, A., Giavaresi, G., & Alessandro, R. (2020). Extracellular Vesicle microRNAs Contribute to the Osteogenic Inhibition of Mesenchymal Stem Cells in Multiple Myeloma. Cancers, 12(2), 449. https://doi.org/10.3390/cancers12020449