Impact of Age-Related Genetic Differences on the Therapeutic Outcome of Papillary Thyroid Cancer

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Patients
2.2. EMT Marker and FGFR Signaling Pathway Are Higher in Pediatric than in Adult Patients
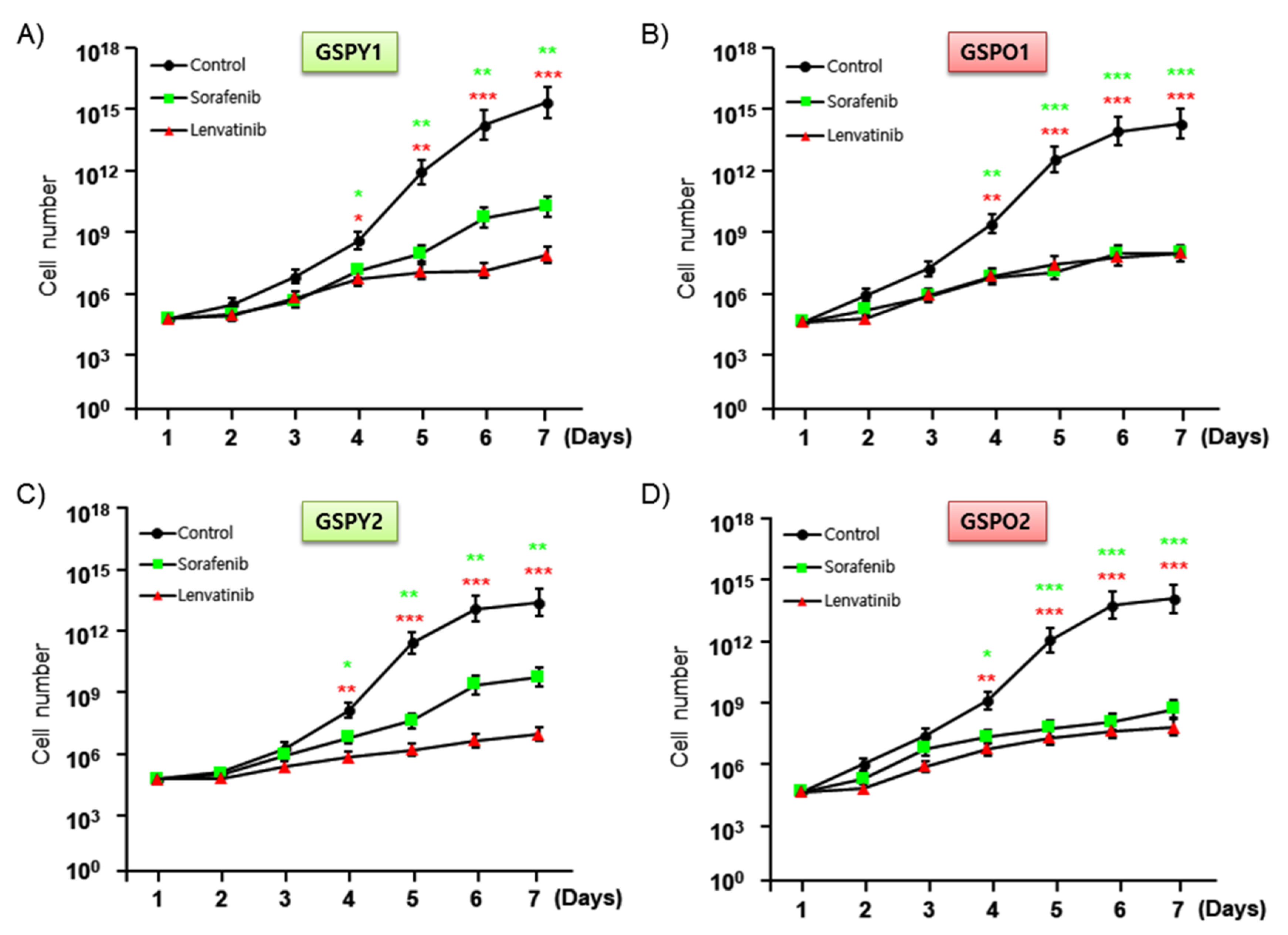
2.3. Lenvatinib More Effectively Inhibited the Proliferation of GSPY than Sorafenib
2.4. Lenvatinib Induces More Effective Reduction of EMT-Mediated FGFR Signaling Pathway in Patient-Derived Thyroid Cancer Cells
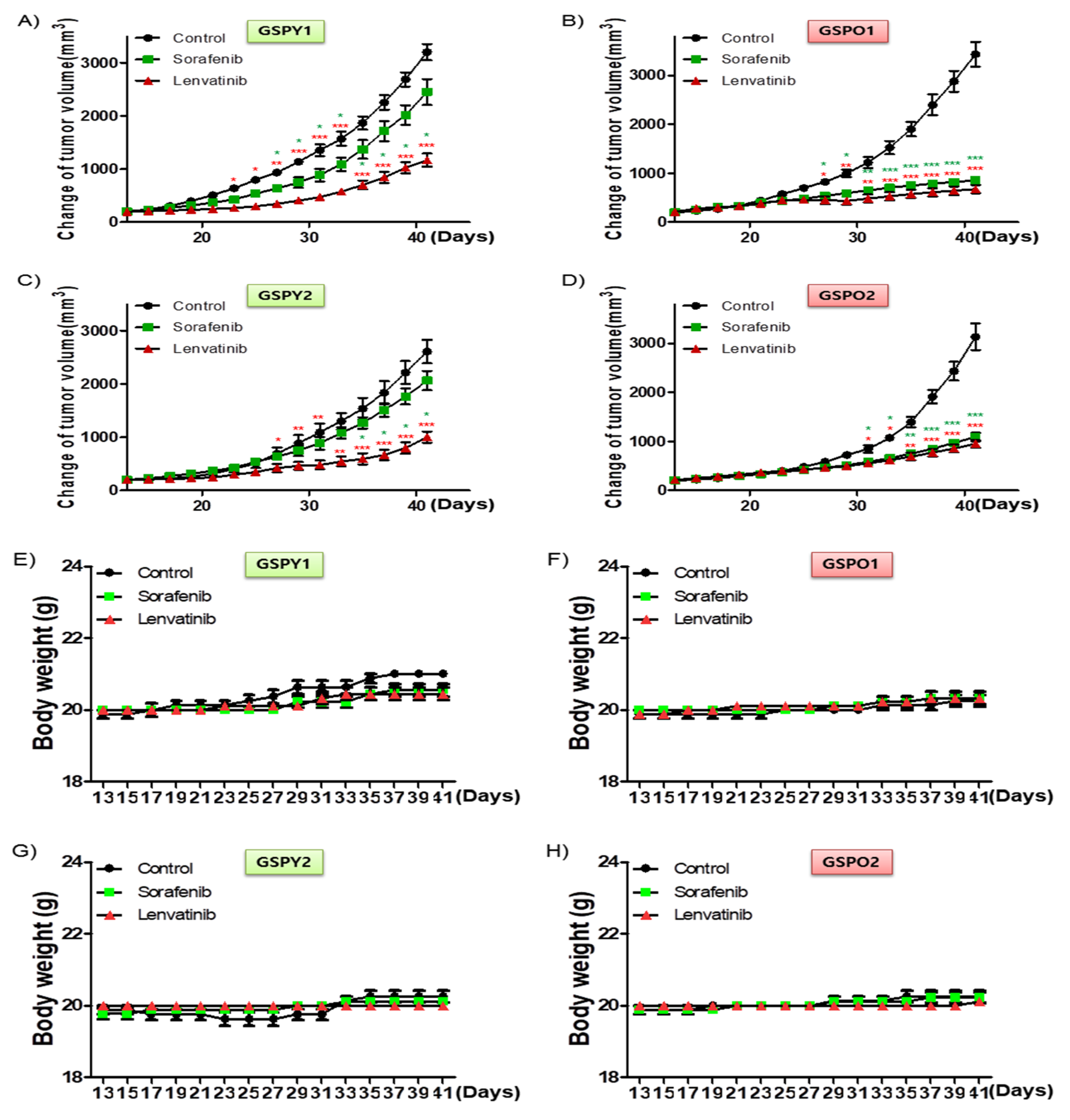
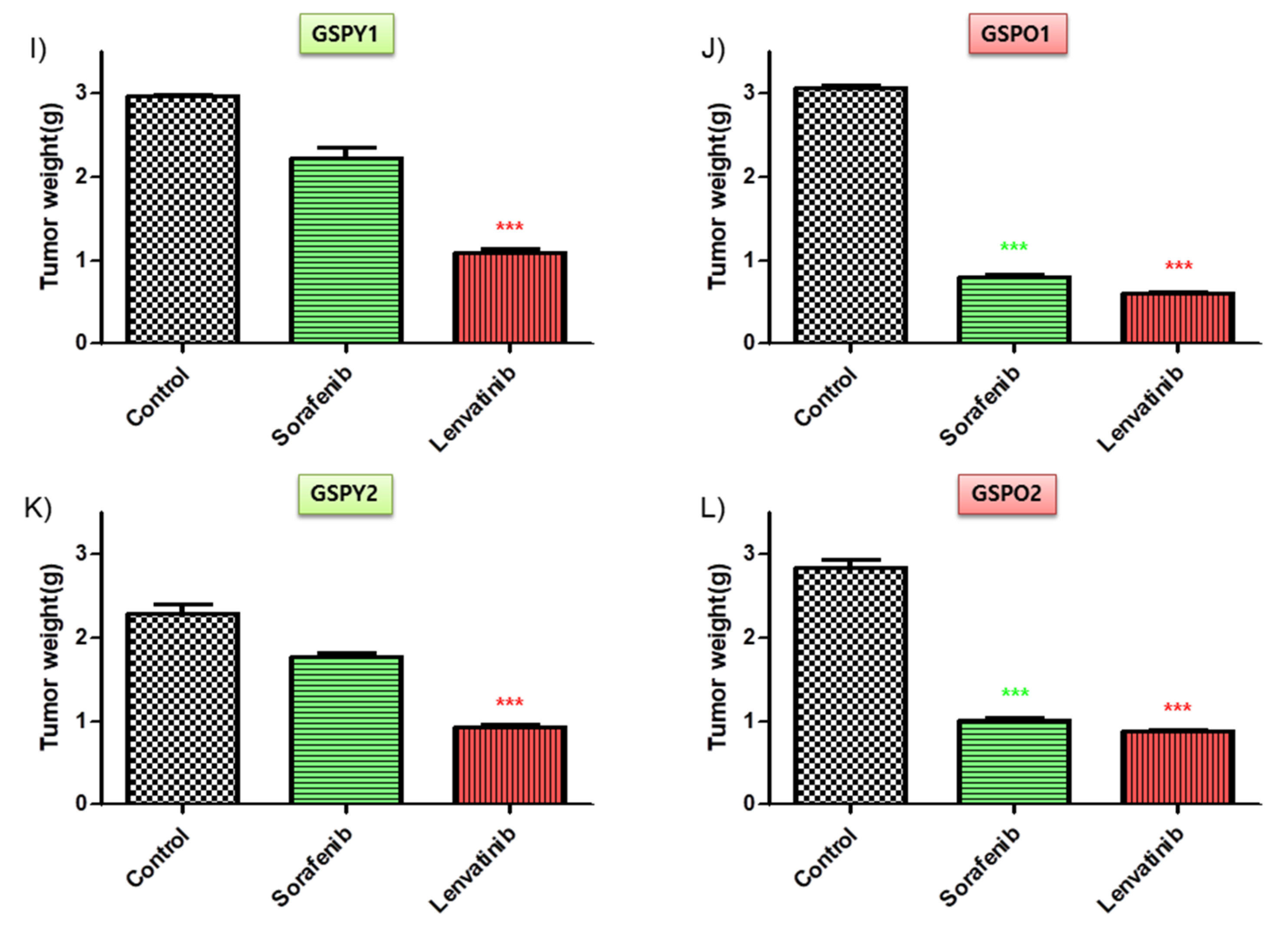
2.5. Significant Tumor Shrinkage was Induced by Lenvatinib in a Xenograft Model of The Patient-Derived PTC Cells
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Patient Tissue Specimens
4.3. Tumor Cell Isolation and Primary Culture
4.4. Cell Culture
4.5. Cell Proliferation Assay
4.6. Microarray Experiment and Data Analysis
4.7. Evaluation of Apoptotic Cell Death
4.8. Immunoblot Analysis
4.9. Immunohistochemistry
4.10. Image Analysis
4.11. Human Papillary Thyroid Cancer Cell Xenografts
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PTC | Papillary Thyroid Carcinoma |
| EMT | Epithelial-Mesenchymal Transition |
| FGFR | Fibroblast Growth Factor Receptor |
| DTC | Differentiated Thyroid Carcinoma |
| IRB | Institutional Review Board |
| DMEM/F12 | Dulbecco’s Modified Eagle’s Medium/Nutrient Mixture F-12 |
| FBS | Fetal Bovine Serum |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
| BCA | Bicinchoninic acid |
| PBS | Phosphate-buffered saline |
| GSPY | Gangnam Severance Papillary Thyroid Carcinoma—Young case |
| GSPO | Gangnam Severance Papillary Thyroid Carcinoma—OLD case |
References
- Nikita, M.E.; Jiang, W.; Cheng, S.M.; Hantash, F.M.; McPhaul, M.J.; Newbury, R.O.; Phillips, S.A.; Reitz, R.E.; Waldman, F.M.; Newfield, R.S. Mutational analysis in pediatric thyroid cancer and correlations with age, ethnicity, and clinical presentation. Thyroid Off. J. Am. Thyroid Assoc. 2016, 26, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Paulson, V.A.; Rudzinski, E.R.; Hawkins, D.S. Thyroid cancer in the pediatric population. Genes 2019, 10, 723. [Google Scholar] [CrossRef] [Green Version]
- Qian, Z.J.; Jin, M.C.; Meister, K.D.; Megwalu, U.C. Pediatric thyroid cancer incidence and mortality trends in the united states, 1973–2013. JAMA Otolaryngol. Head Neck Surg. 2019, 145, 617–623. [Google Scholar] [CrossRef]
- Ben-Shachar, R.; Eisenberg, M.; Huang, S.A.; DiStefano, J.J. Simulation of post-thyroidectomy treatment alternatives for triiodothyronine or thyroxine replacement in pediatric thyroid cancer patients. Thyroid Off. J. Am. Thyroid Assoc. 2012, 22, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Hung, W.; Sarlis, N.J. Current controversies in the management of pediatric patients with well-differentiated nonmedullary thyroid cancer: A review. Thyroid Off. J. Am. Thyroid Assoc. 2002, 12, 683–702. [Google Scholar] [CrossRef] [PubMed]
- Nikiforova, M.N.; Nikiforov, Y.E. Molecular genetics of thyroid cancer: Implications for diagnosis, treatment and prognosis. Expert Rev. Mol. Diagn. 2008, 8, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Bongarzone, I.; Fugazzola, L.; Vigneri, P.; Mariani, L.; Mondellini, P.; Pacini, F.; Basolo, F.; Pinchera, A.; Pilotti, S.; Pierotti, M.A. Age-related activation of the tyrosine kinase receptor protooncogenes ret and ntrk1 in papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 1996, 81, 2006–2009. [Google Scholar] [PubMed] [Green Version]
- Nikiforov, Y.E.; Nikiforova, M.N. Molecular genetics and diagnosis of thyroid cancer. Nat. Rev. Endocrinol. 2011, 7, 569–580. [Google Scholar] [CrossRef]
- Parisi, M.T.; Eslamy, H.; Mankoff, D. Management of differentiated thyroid cancer in children: Focus on the american thyroid association pediatric guidelines. Semin. Nucl. Med. 2016, 46, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Tracy, E.T.; Roman, S.A. Current management of pediatric thyroid disease and differentiated thyroid cancer. Curr. Opin. Oncol. 2016, 28, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Grygielewicz, P.; Dymek, B.; Bujak, A.; Gunerka, P.; Stanczak, A.; Lamparska-Przybysz, M.; Wieczorek, M.; Dzwonek, K.; Zdzalik, D. Epithelial-mesenchymal transition confers resistance to selective fgfr inhibitors in snu-16 gastric cancer cells. Gastric Cancer Off. J. Int. Gastric Cancer Assoc. Jpn. Gastric Cancer Assoc. 2016, 19, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendt, M.K.; Tian, M.; Schiemann, W.P. Deconstructing the mechanisms and consequences of tgf-beta-induced emt during cancer progression. Cell Tissue Res. 2012, 347, 85–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuhlmiller, T.J.; Miller, S.M.; Zawistowski, J.S.; Nakamura, K.; Beltran, A.S.; Duncan, J.S.; Angus, S.P.; Collins, K.A.; Granger, D.A.; Reuther, R.A.; et al. Inhibition of lapatinib-induced kinome reprogramming in erbb2-positive breast cancer by targeting bet family bromodomains. Cell Rep. 2015, 11, 390–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Settleman, J. Emt, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, S.; Gronych, J.; Warnatz, H.J.; Hutter, B.; Gröbner, S.; Ryzhova, M.; Kool, M. Recurrent met fusion genes represent a drug target in pediatric glioblastoma. Nat. Med. 2016, 22, 1314–1320. [Google Scholar]
- Wendt, M.K.; Taylor, M.A.; Schiemann, B.J.; Sossey-Alaoui, K.; Schiemann, W.P. Fibroblast growth factor receptor splice variants are stable markers of oncogenic transforming growth factor beta1 signaling in metastatic breast cancers. Breast Cancer Res. BCR 2014, 16, R24. [Google Scholar] [CrossRef] [Green Version]
- Brown, W.S.; Tan, L.; Smith, A.; Gray, N.S.; Wendt, M.K. Covalent targeting of fibroblast growth factor receptor inhibits metastatic breast cancer. Mol. Cancer Ther. 2016, 15, 2096–2106. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.M.; Lee, J. Met: Roles in epithelial-mesenchymal transition and cancer stemness. Ann. Transl. Med. 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Bernet, V.; Smallridge, R. New therapeutic options for advanced forms of thyroid cancer. Expert Opin. Emerg. Drugs 2014, 19, 225–241. [Google Scholar] [CrossRef]
- Ma, R.; Minsky, N.; Morshed, S.A.; Davies, T.F. Stemness in human thyroid cancers and derived cell lines: The role of asymmetrically dividing cancer stem cells resistant to chemotherapy. J. Clin. Endocrinol. Metab. 2014, 99, E400–E409. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The eighth edition ajcc cancer staging manual: Continuing to build a bridge from a population-based to a more “personalized” approach to cancer staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Zolotov, S. Genetic testing in differentiated thyroid carcinoma: Indications and clinical implications. Rambam Maimonides Med. J. 2016, 7, e0009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, N.; Nikitski, A.V.; Klam, E.; Hunt, J.; Witt, B.; Chadwick, B.; Nikiforov, Y.E.; Abraham, D. Molecular profile and clinical outcomes in differentiated thyroid cancer patients presenting with bone metastasis. Endocr. Pract. Off. J. Am. Coll. Endocrinol. Am. Assoc. Clin. Endocrinol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Galuppini, F.; Vianello, F.; Censi, S.; Barollo, S.; Bertazza, L.; Carducci, S.; Colato, C.; Manso, J.; Rugge, M.; Iacobone, M.; et al. Differentiated thyroid carcinoma in pediatric age: Genetic and clinical scenario. Front. Endocrinol. 2019, 10, 552. [Google Scholar] [CrossRef] [Green Version]
- Pozdeyev, N.; Gay, L.M.; Sokol, E.S.; Hartmaier, R.; Deaver, K.E.; Davis, S.; French, J.D.; Borre, P.V.; LaBarbera, D.V.; Tan, A.C.; et al. Genetic analysis of 779 advanced differentiated and anaplastic thyroid cancers. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 3059–3068. [Google Scholar] [CrossRef] [Green Version]
- Remiker, A.S.; Chuang, J.; Corathers, S.; Rutter, M.M.; Rutter, M.J.; Myer, C.M.T.; Gelfand, M.J.; Trout, A.T.; Geller, J.I. Differentiated thyroid cancer in the pediatric/adolescent population: Evolution of treatment. J. Pediatr. Hematol. Oncol. 2019, 41, 532–536. [Google Scholar] [CrossRef]
- Luster, M.; Aktolun, C.; Amendoeira, I.; Barczynski, M.; Bible, K.C.; Duntas, L.H.; Elisei, R.; Handkiewicz-Junak, D.; Hoffmann, M.; Jarzab, B.; et al. European perspective on the 2015 american thyroid association management guidelines for adult patients with thyroid nodules and differentiated thyroid cancer. Proceedings of an interactive international symposium. Thyroid Off. J. Am. Thyroid Assoc. 2018. [Google Scholar] [CrossRef]
- Diesen, D.L.; Skinner, M.A. Pediatric thyroid cancer. Semin. Pediatr. Surg. 2012, 21, 44–50. [Google Scholar] [CrossRef]
- Kuttesch, J.F., Jr. Multidrug resistance in pediatric oncology. Investig. New Drugs 1996, 14, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Fruci, D.; Cho, W.C.; Nobili, V.; Locatelli, F.; Alisi, A. Drug transporters and multiple drug resistance in pediatric solid tumors. Curr. Drug Metab. 2016, 17, 308–316. [Google Scholar] [CrossRef]
- Kennedy, R.D.; D’Andrea, A.D. DNA repair pathways in clinical practice: Lessons from pediatric cancer susceptibility syndromes. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 3799–3808. [Google Scholar] [CrossRef]
- Pastushenko, I.; Blanpain, C. Emt transition states during tumor progression and metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosland, G.V.; Dyrstad, S.E.; Tusubira, D.; Helwa, R.; Tan, T.Z.; Lotsberg, M.L.; Pettersen, I.K.N.; Berg, A.; Kindt, C.; Hoel, F.; et al. Epithelial to mesenchymal transition (emt) is associated with attenuation of succinate dehydrogenase (sdh) in breast cancer through reduced expression of sdhc. Cancer Metab. 2019, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Kang, Y. Epithelial-mesenchymal plasticity in cancer progression and metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Tsubakihara, Y.; Moustakas, A. Epithelial-mesenchymal transition and metastasis under the control of transforming growth factor beta. Int. J. Mol. Sci. 2018, 19, 3672. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Fan, D. The epithelial-mesenchymal transition and cancer stem cells: Functional and mechanistic links. Curr. Pharm. Des. 2015, 21, 1279–1291. [Google Scholar] [CrossRef]
- Haghpanah, V.; Fallah, P.; Naderi, M.; Tavakoli, R.; Soleimani, M.; Larijani, B. Cancer stem-like cell behavior in anaplastic thyroid cancer: A challenging dilemma. Life Sci. 2016, 146, 34–39. [Google Scholar] [CrossRef]
- Vicari, L.; Colarossi, C.; Giuffrida, D.; De Maria, R.; Memeo, L. Cancer stem cells as a potential therapeutic target in thyroid carcinoma. Oncol. Lett. 2016, 12, 2254–2260. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Hardin, H.; Lloyd, R.V. Cancer stem-like cells and thyroid cancer. Endocr. Relat. Cancer 2014, 21, T285–T300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borah, A.; Raveendran, S.; Rochani, A.; Maekawa, T.; Kumar, D.S. Targeting self-renewal pathways in cancer stem cells: Clinical implications for cancer therapy. Oncogenesis 2015, 4, e177. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Diaz, P.C.; Hsiao, T.H.; Chang, J.C.; Yue, D.; Tan, M.C.; Chen, H.I.; Tomlinson, G.E.; Huang, Y.; Chen, Y.; Hung, J.Y. De-regulated micrornas in pediatric cancer stem cells target pathways involved in cell proliferation, cell cycle and development. PLoS ONE 2013, 8, e61622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, F.H.; Li, Y.; Wang, Z.; Kong, D. Pancreatic cancer stem cells and emt in drug resistance and metastasis. Minerva Chir. 2009, 64, 489–500. [Google Scholar] [PubMed]
- Phi, L.T.H.; Sari, I.N.; Yang, Y.G.; Lee, S.H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer stem cells (cscs) in drug resistance and their therapeutic implications in cancer treatment. Stem Cells Int. 2018, 2018, 5416923. [Google Scholar] [CrossRef] [Green Version]
- Kroemer, G. The proto-oncogene bcl-2 and its role in regulating apoptosis. Nat. Med. 1997, 3, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.J.; Tait, S.W.G. Targeting bcl-2 regulated apoptosis in cancer. Open Biol. 2018, 8, 180002. [Google Scholar] [CrossRef]
- Reed, J.C. Bcl-2 on the brink of breakthroughs in cancer treatment. Cell Death Differ. 2018, 25, 3–6. [Google Scholar] [CrossRef] [Green Version]
- An, J.; Lv, J.; Li, A.; Qiao, J.; Fang, L.; Li, Z.; Li, B.; Zhao, W.; Chen, H.; Wang, L. Constitutive expression of bcl-2 induces epithelial-mesenchymal transition in mammary epithelial cells. BMC Cancer 2015, 15, 476. [Google Scholar] [CrossRef] [Green Version]
- Soderquist, R.S.; Crawford, L.; Liu, E.; Lu, M.; Agarwal, A.; Anderson, G.R.; Lin, K.H.; Winter, P.S.; Cakir, M.; Wood, K.C. Systematic mapping of bcl-2 gene dependencies in cancer reveals molecular determinants of bh3 mimetic sensitivity. Nat. Commun. 2018, 9, 3513. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Patients |
|---|---|
| N = 52 | |
| Age (years), mean ± SD (range) | 12.5 ± 2.9 (5–15) |
| Male:female ratio, n (%) | 11 (21.2):41 (78.8) |
| Initial presentation, n (%) | |
| Incidentaloma | 18 (34.6%) |
| Palpable mass | 33 (63.5%) |
| Thyrotoxicosis | 1 (1.9%) |
| Famliy history of thyroid cancer, n (%) | 4 (7.7%) |
| Histology, n (%) | |
| Papillary | 45 (86.5%) |
| Follicular | 6 (11.5%) |
| Medullary | 1 (1.9%) |
| Tumor size (cm), mean ± SD | 2.4 ± 1.4 |
| Extrathyroidal extension | 32 (61.5%) |
| Vascular invasion | 41 (78.8%) |
| TNM stage | |
| T stage | |
| T1:T2:T3:T4 | 13 (25.0):4 (7.7):31 (59.6):4(7.7) |
| N stage | |
| N0:N1a:N1b | 12 (23.1):9 (17.3):31 (59.6) |
| M stage | |
| M0:M1 | 50 (96.2):2 (3.8) |
| Time to recurrence (years), mean ± SD (range) | 8.3 ± 5.6 (1.1–11.7) |
| Follow up period (years), mean ± SD (range) | 9.8 ± 5.5 (4.1–13.7) |
| Variables | GSPY1 | GSPY2 | GSPO1 | GSPO2 |
|---|---|---|---|---|
| Age at Diagnosis | 4 | 13 | 52 | 47 |
| Gender | Female | Female | Female | Female |
| Primary Disease Site | Thyroid | Thyroid | Thyroid | Thyroid |
| Stage | T4aN1bM0 | T4aN1bM0 | T4aN1bM1 | T3N1bM1 |
| Primary Pathology (subtype) | Papillary thyroid cancer (conventional) | Papillary thyroid cancer (conventional) | Papillary thyroid cancer (conventional) | Papillary thyroid cancer (conventional) |
| Classification of specimen used for culture | Fresh tumor | Fresh tumor | Fresh tumor | Fresh tumor |
| Obtained from | Gangnam Severance Hospital, Seoul, Korea | Gangnam Severance Hospital, Seoul, Korea | Gangnam Severance Hospital, Seoul, Korea | Gangnam Severance Hospital, Seoul, Korea |
| Cell Line | Hisopathology | Animal | Cell Proliferation IC50 (μM) | |
|---|---|---|---|---|
| Sorafenib | Lenvatinib | |||
| GSPY1 | Thyroid cancer; Papillary | Human | 17.24 (±0.5) | 15.75 (±0.2) |
| GSPY2 | Thyroid cancer; Papillary | Human | 20.25 (±0.1) | 16.54 (±0.3) |
| GSPO1 | Thyroid cancer; Papillary | Human | 9.84 (±0.5) | 10.23 (±0.4) |
| GSPO2 | Thyroid cancer; Papillary | Human | 10.12 (±0.2) | 10.98 (±0.1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-M.; Kim, S.Y.; Park, C.S.; Chang, H.-S.; Park, K.C. Impact of Age-Related Genetic Differences on the Therapeutic Outcome of Papillary Thyroid Cancer. Cancers 2020, 12, 448. https://doi.org/10.3390/cancers12020448
Kim S-M, Kim SY, Park CS, Chang H-S, Park KC. Impact of Age-Related Genetic Differences on the Therapeutic Outcome of Papillary Thyroid Cancer. Cancers. 2020; 12(2):448. https://doi.org/10.3390/cancers12020448
Chicago/Turabian StyleKim, Seok-Mo, Soo Young Kim, Cheong Soo Park, Hang-Seok Chang, and Ki Cheong Park. 2020. "Impact of Age-Related Genetic Differences on the Therapeutic Outcome of Papillary Thyroid Cancer" Cancers 12, no. 2: 448. https://doi.org/10.3390/cancers12020448
APA StyleKim, S. -M., Kim, S. Y., Park, C. S., Chang, H. -S., & Park, K. C. (2020). Impact of Age-Related Genetic Differences on the Therapeutic Outcome of Papillary Thyroid Cancer. Cancers, 12(2), 448. https://doi.org/10.3390/cancers12020448