Aspartate-β-Hydroxylase: A Promising Target to Limit the Local Invasiveness of Colorectal Cancer

 ,
,  , ,
, ,  and
and 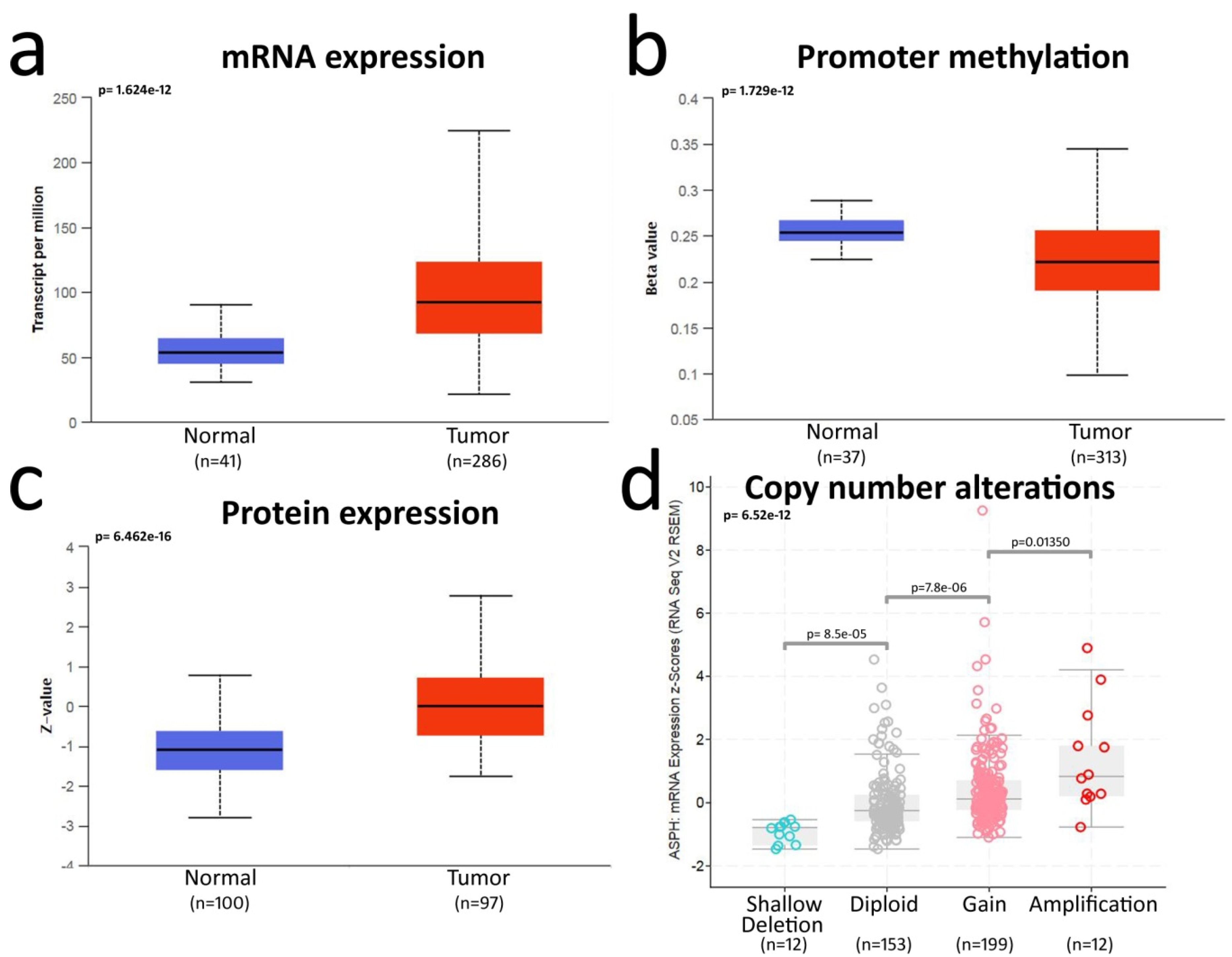{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
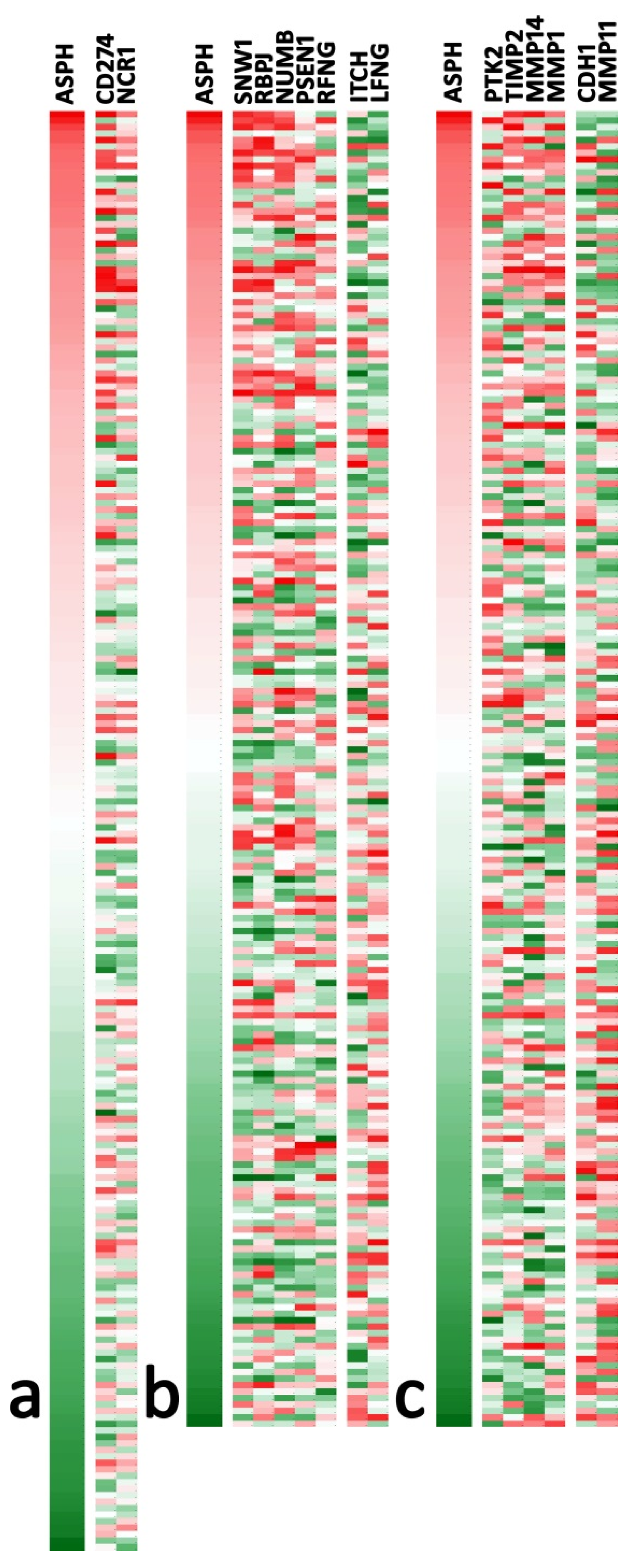
2.1. Bioinformatics Analysis Shows ASPH Upregulation in CRC
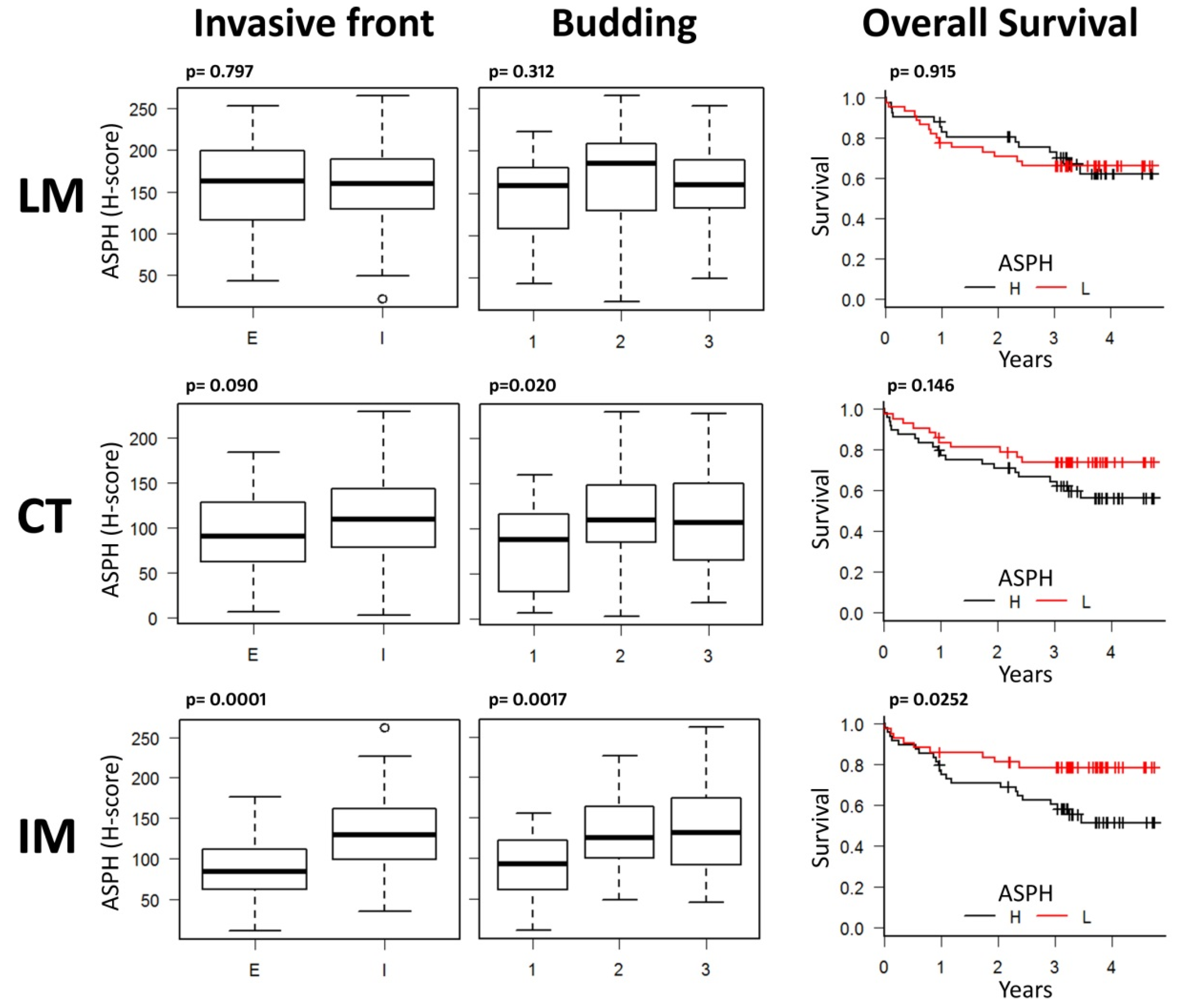
2.2. Digital Pathology Analysis Shows a Specific Role for ASPH in the Invasive Tumor Margin
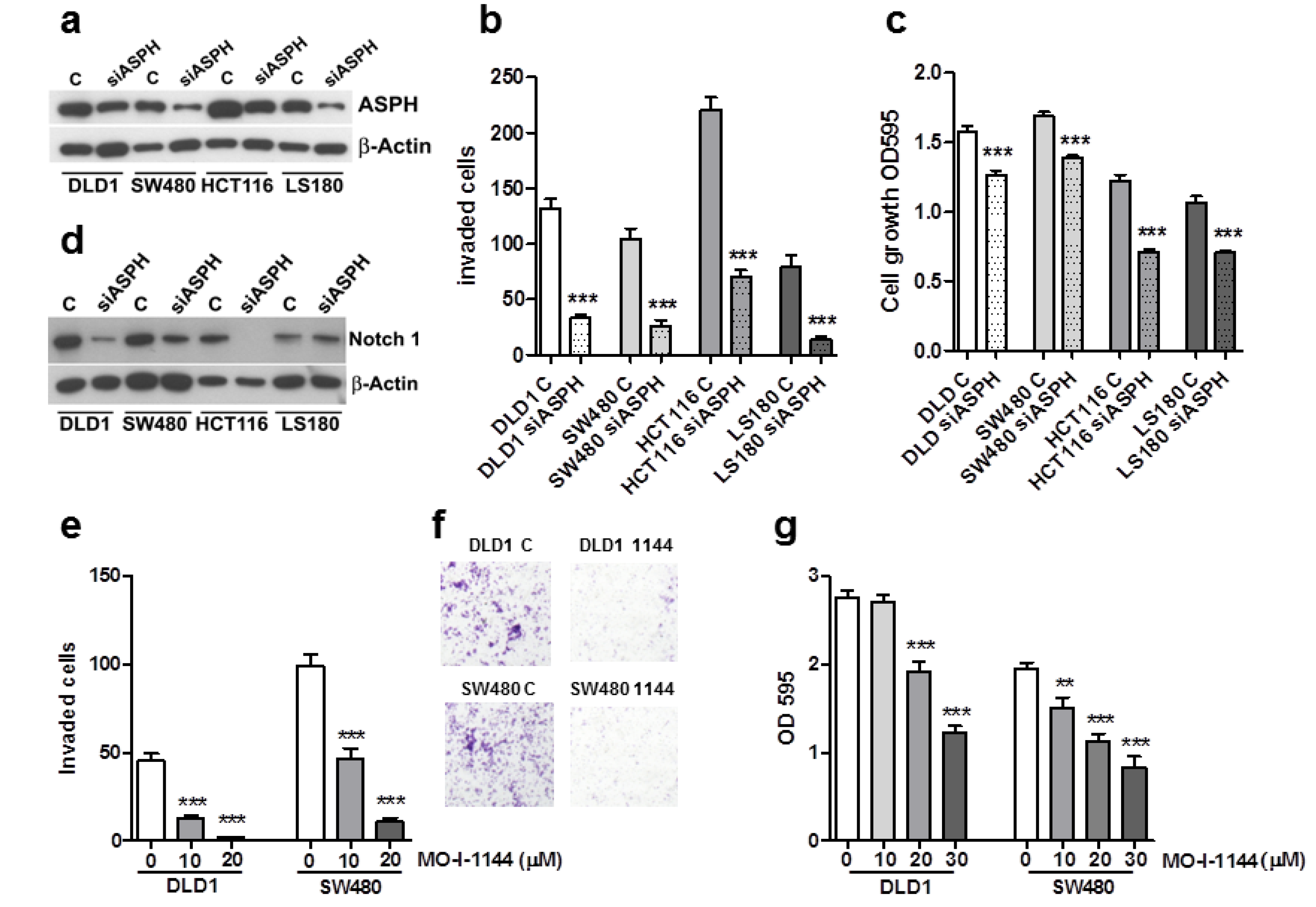
2.3. ASPH Downregulation Affects CRC Cell Line Invasion and Growth
2.4. ASPH Enzymatic Activity Inhibition Decreases CRC Cell Invasion and Growth
3. Materials and Methods
3.1. Bioinformatics Analysis
3.2. Anti-ASPH Antibody
3.3. Patient Cohort
3.4. Digital Pathology Analysis
3.5. Cell Lines
3.6. ASPH Silencing, Cell Proliferation and Invasion Assays
3.7. ASPH Inhibitors
3.8. Protein Extraction and Western Blot
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Suman, S.; Das, T.P.; Ankem, M.K.; Damodaran, C. Targeting Notch Signaling in Colorectal Cancer. Curr. Colorectal Cancer Rep. 2014, 10, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Lavaissiere, L.; Jia, S.; Nishiyama, M.; de la Monte, S.; Stern, A.M.; Wands, J.R.; Friedman, P.A. Overexpression of human aspartyl(asparaginyl)beta-hydroxylase in hepatocellular carcinoma and cholangiocarcinoma. J. Clin. Investig. 1996, 98, 1313–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Taguchi, K.; Aishima, S.; Shimada, M.; Hintz, D.; Larusso, N.; Gores, G.; Tsuneyoshi, M.; Sugimachi, K.; Wands, J.R.; et al. Clinicopathological correlates of aspartyl (asparaginyl) beta-hydroxylase over-expression in cholangiocarcinoma. Cancer Detect. Prev. 2004, 28, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Xu, B.; Bi, Y.; Wu, C.; Ru, B.; Sun, B.; Bai, X. Recent advances in research on aspartate β-hydroxylase (ASPH) in pancreatic cancer: A brief update. Bosn. J. Basic Med. Sci. 2018, 18, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Aihara, A.; Huang, C.-K.; Olsen, M.J.; Lin, Q.; Chung, W.; Tang, Q.; Dong, X.; Wands, J.R. A cell-surface β-hydroxylase is a biomarker and therapeutic target for hepatocellular carcinoma. Hepatology 2014, 60, 1302–1313. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Lin, Q.; Aihara, A.; Li, Y.; Huang, C.-K.; Chung, W.; Tang, Q.; Chen, X.; Carlson, R.; Nadolny, C.; et al. Aspartate β-Hydroxylase expression promotes a malignant pancreatic cellular phenotype. Oncotarget 2015, 6, 1231–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawton, M.; Tong, M.; Gundogan, F.; Wands, J.R.; de la Monte, S.M. Aspartyl-(asparaginyl) beta-hydroxylase, hypoxia-inducible factor-alpha and Notch cross-talk in regulating neuronal motility. Oxid. Med. Cell Longev. 2010, 3, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Ince, N.; de la Monte, S.M.; Wands, J.R. Overexpression of human aspartyl (asparaginyl) beta-hydroxylase is associated with malignant transformation. Cancer Res. 2000, 60, 1261–1266. [Google Scholar] [PubMed]
- Wang, J.; de la Monte, S.M.; Sabo, E.; Kethu, S.; Tavares, R.; Branda, M.; Simao, L.; Wands, J.R.; Resnick, M.B. Prognostic value of humbug gene overexpression in stage II colon cancer. Hum. Pathol. 2007, 38, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Dinchuk, J.E.; Henderson, N.L.; Burn, T.C.; Huber, R.; Ho, S.P.; Link, J.; T O’Neil, K.; Focht, R.J.; Scully, M.S.; Hollis, J.M. Aspartyl β-hydroxylase (Asph) and an evolutionarily conserved isoform of Asph missing the catalytic domain share exons with junctin. J. Biol. Chem. 2000, 275, 39543–39554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugli, A.; Kirsch, R.; Ajioka, Y.; Bosman, F.; Cathomas, G.; Dawson, H.; Zimaity, H.E.; Fléjou, J.-F.; Hansen, T.P.; Hartmann, A.; et al. Recommendations for reporting tumor budding in colorectal cancer based on the International Tumor Budding Consensus Conference (ITBCC) 2016. Mod. Pathol. 2017, 30, 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-K.; Iwagami, Y.; Aihara, A.; Chung, W.; de la Monte, S.; Thomas, J.-M.; Olsen, M.; Carlson, R.; Yu, T.; Dong, X.; et al. Anti-Tumor Effects of Second Generation β-Hydroxylase Inhibitors on Cholangiocarcinoma Development and Progression. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Jing, L.; Ruihua, T.; Ji, L.; Yaxiong, L.; Linjie, Y.; Dongyan, S.; Mingliang, J.; Qingsheng, H.; Junling, S. The aspartyl (asparaginyl) beta-hydroxylase in carcinomas. Front. Biosci. 2015, 20, 902–909. [Google Scholar]
- Patel, N.; Khan, A.O.; Mansour, A.; Mohamed, J.Y.; Al-Assiri, A.; Haddad, R.; Jia, X.; Xiong, Y.; Mégarbané, A.; Traboulsi, E.I.; et al. Mutations in ASPH cause facial dysmorphism, lens dislocation, anterior-segment abnormalities, and spontaneous filtering blebs, or Traboulsi syndrome. Am. J. Hum. Genet. 2014, 94, 755–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, I.; Brewitz, L.; Krojer, T.; Jensen, S.A.; Kochan, G.T.; Kershaw, N.J.; Hewitson, K.S.; McNeill, L.A.; Kramer, H.; Münzel, M.; et al. Aspartate/asparagine-β-hydroxylase crystal structures reveal an unexpected epidermal growth factor-like domain substrate disulfide pattern. Nat. Commun. 2019, 10, 4910. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benelli, R.; Costa, D.; Mastracci, L.; Grillo, F.; Olsen, M.J.; Barboro, P.; Poggi, A.; Ferrari, N. Aspartate-β-Hydroxylase: A Promising Target to Limit the Local Invasiveness of Colorectal Cancer. Cancers 2020, 12, 971. https://doi.org/10.3390/cancers12040971
Benelli R, Costa D, Mastracci L, Grillo F, Olsen MJ, Barboro P, Poggi A, Ferrari N. Aspartate-β-Hydroxylase: A Promising Target to Limit the Local Invasiveness of Colorectal Cancer. Cancers. 2020; 12(4):971. https://doi.org/10.3390/cancers12040971
Chicago/Turabian StyleBenelli, Roberto, Delfina Costa, Luca Mastracci, Federica Grillo, Mark Jon Olsen, Paola Barboro, Alessandro Poggi, and Nicoletta Ferrari. 2020. "Aspartate-β-Hydroxylase: A Promising Target to Limit the Local Invasiveness of Colorectal Cancer" Cancers 12, no. 4: 971. https://doi.org/10.3390/cancers12040971
APA StyleBenelli, R., Costa, D., Mastracci, L., Grillo, F., Olsen, M. J., Barboro, P., Poggi, A., & Ferrari, N. (2020). Aspartate-β-Hydroxylase: A Promising Target to Limit the Local Invasiveness of Colorectal Cancer. Cancers, 12(4), 971. https://doi.org/10.3390/cancers12040971