
The Functional Role of Long Non-Coding RNAs in Melanoma


Abstract
:Simple Summary
Abstract
1. Introduction
2. Protein-lncRNA Interactions in Melanoma
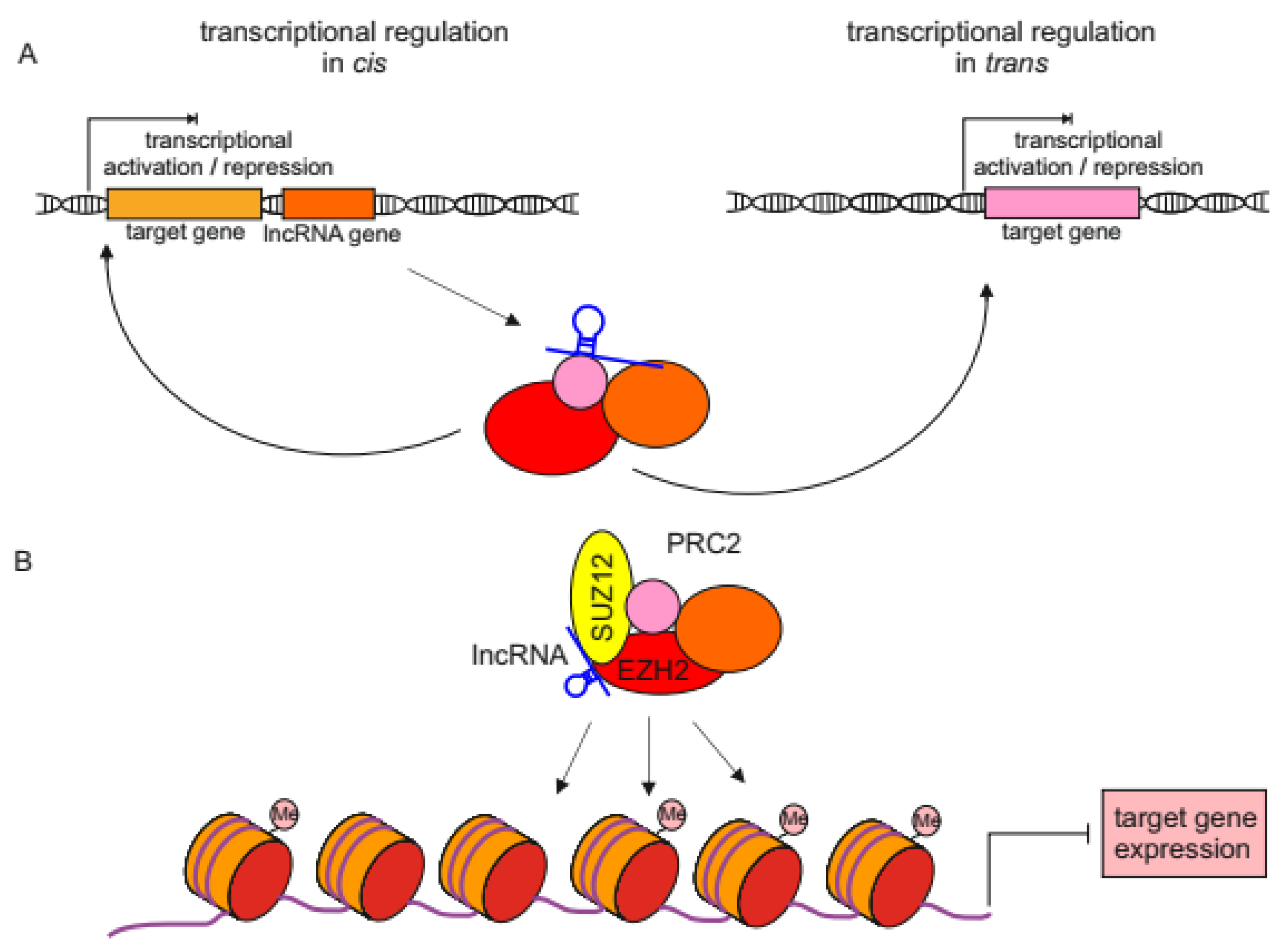
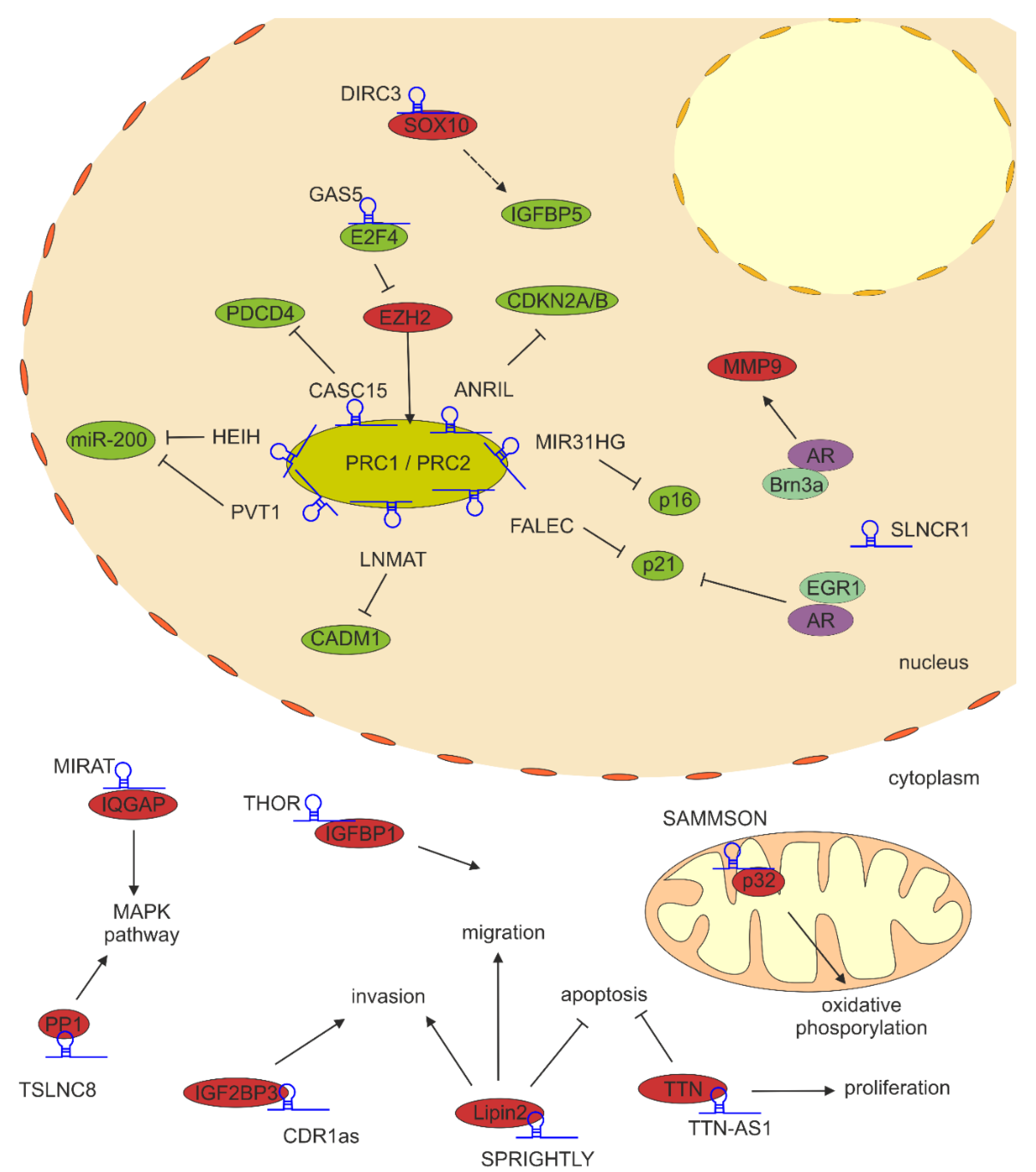
2.1. lncRNAs Regulating Transcription in Nucleus
2.2. lncRNA Scaffolding Proteins in Melanoma
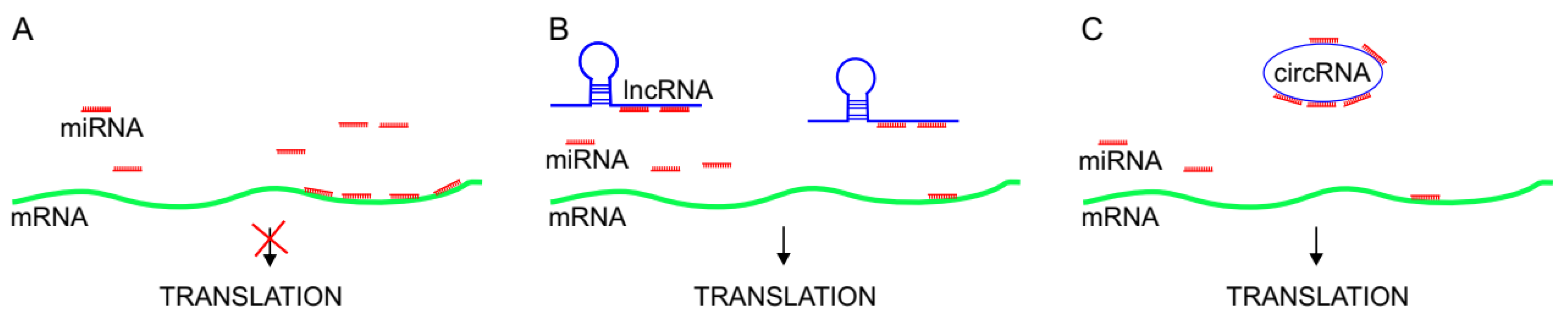
3. Competing Endogenous RNAs in Direct interactions with miRNAs
3.1. lncRNAs as miRNA Sponges in Melanoma
3.2. circRNAs as miRNA Sponges in Melanoma
3.3. The ceRNA Controversy
4. lncRNAs Involved in Melanoma Drug Resistance and Treatment Response
5. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Bollag, G.; Tsai, J.; Zhang, J.; Zhang, C.; Ibrahim, P.; Nolop, K.; Hirth, P. Vemurafenib: The first drug approved for BRAF-mutant cancer. Nat. Rev. Drug Discov. 2012, 11, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Dummer, R.; Ascierto, P.A.; Gogas, H.J.; Arance, A.; Mandala, M.; Liszkay, G.; Garbe, C.; Schadendorf, D.; Krajsova, I.; Gutzmer, R.; et al. Encorafenib plus binimetinib versus vemurafenib or encorafenib in patients with BRAF-mutant melanoma (COLUMBUS): A multicentre, open-label, randomised phase 3 trial. Lancet Oncol. 2018, 19, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Broman, K.K.; Dossett, L.A.; Sun, J.; Eroglu, Z.; Zager, J.S. Update on BRAF and MEK inhibition for treatment of melanoma in metastatic, unresectable, and adjuvant settings. Expert Opin. Drug Saf. 2019, 18, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, Z.; Ribas, A. Combination therapy with BRAF and MEK inhibitors for melanoma: Latest evidence and place in therapy. Ther. Adv. Med. Oncol. 2016, 8, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Vanella, V.; Festino, L.; Trojaniello, C.; Vitale, M.G.; Sorrentino, A.; Paone, M.; Ascierto, P.A. The Role of BRAF-Targeted Therapy for Advanced Melanoma in the Immunotherapy Era. Curr. Oncol. Rep. 2019, 21, 76. [Google Scholar] [CrossRef]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-Year Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2019, 38, 1535–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuryk, L.; Bertinato, L.; Staniszewska, M.; Pancer, K.; Wieczorek, M.; Salmaso, S.; Caliceti, P.; Garofalo, M. From Conventional Therapies to Immuno-therapy: Melanoma Treatment in Review. Cancers 2020, 12, 3057. [Google Scholar] [CrossRef] [PubMed]
- Carlino, M.S.; Larkin, J.; Long, G.V. Immune checkpoint inhibitors in melanoma. Lancet 2021, 398, 1002–1014. [Google Scholar] [CrossRef]
- Ascierto, P.A.; Ferrucci, P.F.; Fisher, R.; Del Vecchio, M.; Atkinson, V.; Schmidt, H.; Schachter, J.; Queirolo, P.; Long, G.V.; Di Giacomo, A.M.; et al. Dabrafenib, trametinib and pembrolizumab or placebo in BRAF-mutant melanoma. Nat. Med. 2019, 25, 941–946. [Google Scholar] [CrossRef]
- Ribas, A.; Dummer, R.; Puzanov, I.; VanderWalde, A.; Andtbacka, R.H.I.; Michielin, O.; Olszanski, A.J.; Malvehy, J.; Cebon, J.; Fernandez, E.; et al. Oncolytic Virotherapy Promotes Intratumoral T Cell Infiltration and Improves Anti-PD-1 Immunotherapy. Cell 2017, 170, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Ribas, A.; Lawrence, D.; Atkinson, V.; Agarwal, S.; Miller, W.H.; Carlino, M.S.; Fisher, R.; Long, G.V.; Hodi, F.S.; Tsoi, J.; et al. Combined BRAF and MEK inhibition with PD-1 blockade immunotherapy in BRAF-mutant melanoma. Nat. Med. 2019, 25, 936–940. [Google Scholar] [CrossRef]
- Trojaniello, C.; Vitale, M.G.; Ascierto, P.A. Triplet combination of BRAF, MEK and PD-1/PD-L1 blockade in melanoma: The more the better? Curr. Opin. Oncol. 2021, 33, 133–138. [Google Scholar] [CrossRef]
- Kasakovski, D.; Skrygan, M.; Gambichler, T.; Susok, L. Advances in Targeting Cutaneous Melanoma. Cancers 2021, 13, 2090. [Google Scholar] [CrossRef] [PubMed]
- Atkins, M.B.; Curiel-Lewandrowski, C.; Fisher, D.E.; Swetter, S.M.; Tsao, H.; Aguirre-Ghiso, J.A.; Soengas, M.S.; Weeraratna, A.T.; Flaherty, K.T.; Herlyn, M.; et al. Melanoma Research Foundation. The State of Melanoma: Emergent Challenges and Opportunities. Clin. Cancer Res. 2021, 27, 2678–2697. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Atkins, M.B.; Eggermont, A.M.; Gershenwald, J.E.; Grob, J.J.; Hamid, O.; Sondak, V.K.; Sosman, J.A.; Tawbi, H.A.; Weber, J.S.; et al. The “Great Debate” at Melanoma Bridge 2020: December, 5th, 2020. J. Transl. Med. 2021, 19, 142. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Yacoub, N.; Mishra, R.; White, A.; Long, Y.; Alanazi, S.; Garrett, J.T. Current Advances in the Treatment of BRAF-Mutant Melanoma. Cancers 2020, 12, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Lui, P.Y.; Jin, D.Y.; Stevenson, N.J. MicroRNA: Master controllers of intracellular signaling pathways. Cell. Mol. Life Sci. 2015, 72, 3531–3542. [Google Scholar] [CrossRef]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [Green Version]
- Dinger, M.E.; Pang, K.C.; Mercer, T.R.; Mattick, J.S. Differentiating protein-coding and noncoding RNA: Challenges and ambiguities. PLoS Comput. Biol. 2008, 4, e1000176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [Green Version]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. Conserved function of lincRNAs in vertebrate embryonic development despite rapid sequence evolution. Cell 2011, 147, 1537–5150. [Google Scholar] [CrossRef] [Green Version]
- Hezroni, H.; Koppstein, D.; Schwartz, M.G.; Avrutin, A.; Bartel, D.P.; Ulitsky, I. Principles of long noncoding RNA evolution derived from direct comparison of transcriptomes in 17 species. Cell Rep. 2015, 11, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, P.J.; Chang, H.Y. Long noncoding RNAs: Cellular address codes in development and disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, T.; Fraser, P. No-nonsense functions for long noncoding RNAs. Cell 2011, 145, 178–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spizzo, R.; Almeida, M.I.; Colombatti, A.; Calin, G.A. Long non-coding RNAs and cancer: A new frontier of translational research? Oncogene 2012, 31, 4577–4587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Liang, Y.; Han, S.; Peng, C.; Li, Y. Long Noncoding RNA and Protein Interactions: From Experimental Results to Computational Models Based on Network Methods. Int. J. Mol. Sci. 2019, 20, 1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Urrutia, E.; Bustamante Montes, L.P.; Ladrón de Guevara Cervantes, D.; Pérez-Plasencia, C.; Campos-Parra, A.D. Crosstalk Between Long Non-coding RNAs, Micro-RNAs and mRNAs: Deciphering Molecular Mechanisms of Master Regulators in Cancer. Front. Oncol. 2019, 9, 669. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Xia, J.; Xie, S.; Zou, R.; Pan, S.; Wang, Z.W.; Assaraf, Y.G.; Zhu, X. Long non-coding RNAs as a determinant of cancer drug resistance: Towards the overcoming of chemoresistance via modulation of lncRNAs. Drug Resist. Updat. 2020, 50, 100683. [Google Scholar] [CrossRef] [PubMed]
- Bande, M.; Fernandez-Diaz, D.; Fernandez-Marta, B.; Rodriguez-Vidal, C.; Lago-Baameiro, N.; Silva-Rodríguez, P.; Paniagua, L.; Blanco-Teijeiro, M.J.; Pardo, M.; Piñeiro, A. The Role of Non-Coding RNAs in Uveal Melanoma. Cancers 2020, 12, 2944. [Google Scholar] [CrossRef]
- Milán-Rois, P.; Quan, A.; Slack, F.J.; Somoza, Á. The Role of LncRNAs in Uveal Melanoma. Cancers 2021, 13, 4041. [Google Scholar] [CrossRef] [PubMed]
- Beroukhim, R.; Mermel, C.H.; Porter, D.; Wei, G.; Raychaudhuri, S.; Donovan, J.; Barretina, J.; Boehm, J.S.; Dobson, J.; Urashima, M.; et al. The landscape of somatic copy-number alteration across human cancers. Nature 2010, 463, 899–905. [Google Scholar] [CrossRef]
- Tripathi, R.; Chakraborty, P.; Varadwaj, P.K. Unraveling long non-coding RNAs through analysis of high-throughput RNA-sequencing data. Noncoding RNA Res. 2017, 2, 111–118. [Google Scholar] [CrossRef]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef]
- Ribeiro, D.M.; Zanzoni, A.; Cipriano, A.; Delli Ponti, R.; Spinelli, L.; Ballarino, M.; Bozzoni, I.; Tartaglia, G.G.; Brun, C. Protein complex scaffolding predicted as a prevalent function of long non-coding RNAs. Nucleic Acids Res. 2018, 46, 917–928. [Google Scholar] [CrossRef]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell. 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Zhao, B.; Li, D.; Yin, G. Long non-coding RNA CASC15 promotes melanoma progression by epigenetically regulating PDCD4. Cell Biosci. 2018, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Hanniford, D.; Ulloa-Morales, A.; Karz, A.; Berzoti-Coelho, M.G.; Moubarak, R.S.; Sánchez-Sendra, B.; Kloetgen, A.; Davalos, V.; Imig, J.; Wu, P.; et al. Epigenetic Silencing of CDR1as Drives IGF2BP3-Mediated Melanoma Invasion and Metastasis. Cancer Cell 2020, 37, 55–70.e15. [Google Scholar] [CrossRef] [PubMed]
- Coe, E.A.; Tan, J.Y.; Shapiro, M.; Louphrasitthiphol, P.; Bassett, A.R.; Marques, A.C.; Goding, C.R.; Vance, K.W. The MITF-SOX10 regulated long non-coding RNA DIRC3 is a melanoma tumour suppressor. PLoS Genet. 2019, 15, e1008501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, N.; Song, H.; Wang, X.; Xu, X.; Jiang, Y.; Sun, J. Up-regulation of long noncoding RNA FALEC predicts poor prognosis and promotes melanoma cell proliferation through epigenetically silencing p21. Biomed. Pharmacother. 2017, 96, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Yan, Z.; Hu, F.; Wei, W.; Yang, C.; Sun, Z. Long non-coding RNA GAS5 accelerates oxidative stress in melanoma cells by rescuing EZH2-mediated CDKN1C downregulation. Cancer Cell Int. 2020, 20, 116. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Xing, G.; Wang, Y.; Luo, Z.; Liu, G.; Meng, H. Long noncoding RNA HEIH promotes melanoma cell proliferation, migration and invasion via inhibition of miR-200b/a/429. Biosci. Rep. 2017, 37, BSR20170682. [Google Scholar] [CrossRef] [Green Version]
- Mou, K.; Zhang, X.; Mu, X.; Ge, R.; Han, D.; Zhou, Y.; Wang, L. LNMAT1 Promotes Invasion-Metastasis Cascade in Malignant Melanoma by Epigenetically Suppressing CADM1 Expression. Front. Oncol. 2019, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- Montes, M.; Nielsen, M.M.; Maglieri, G.; Jacobsen, A.; Højfeldt, J.; Agrawal-Singh, S.; Hansen, K.; Helin, K.; van de Werken, H.J.G.; Pedersen, J.S.; et al. The lncRNA MIR31HG regulates p16(INK4A) expression to modulate senescence. Nat. Commun. 2015, 6, 6967. [Google Scholar] [CrossRef] [Green Version]
- Sanlorenzo, M.; Vujic, I.; Esteve-Puig, R.; Lai, K.; Vujic, M.; Lin, K.; Posch, C.; Dimon, M.; Moy, A.; Zekhtser, M.; et al. The lincRNA MIRAT binds to IQGAP1 and modulates the MAPK pathway in NRAS mutant melanoma. Sci. Rep. 2018, 8, 10902. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, D.; Li, Y.; Li, X.; Zhao, L.; Zhang, J.; Song, Y. Effect of long non-coding RNA PVT1 on cell proliferation and migration in melanoma. Int. J. Mol. Med. 2018, 41, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Leucci, E.; Vendramin, R.; Spinazzi, M.; Laurette, P.; Fiers, M.; Wouters, J.; Radaelli, E.; Eyckerman, S.; Leonelli, C.; Vanderheyden, K.; et al. Melanoma addiction to the long non-coding RNA SAMMSON. Nature 2016, 531, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Joyce, C.E.; Buquicchio, F.; Brown, A.; Ritz, J.; Distel, R.J.; Yoon, C.H.; Novina, C.D. The lncRNA SLNCR1 Mediates Melanoma Invasion through a Conserved SRA1-like Region. Cell Rep. 2016, 15, 2025–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, K.; Carroll, J.S.; Yee, E.; Thomas, D.D.; Wert-Lamas, L.; Neier, S.C.; Sheynkman, G.; Ritz, J.; Novina, C.D. The lncRNA SLNCR Recruits the Androgen Receptor to EGR1-Bound Genes in Melanoma and Inhibits Expression of Tumor Suppressor p21. Cell Rep. 2019, 27, 2493–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazar, J.; Zhao, W.; Khalil, A.M.; Lee, B.; Shelley, J.; Govindarajan, S.S.; Yamamoto, F.; Ratnam, M.; Aftab, M.N.; Collins, S.; et al. The functional characterization of long noncoding RNA SPRY4-IT1 in human melanoma cells. Oncotarget 2014, 5, 8959–8969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosono, Y.; Niknafs, Y.S.; Prensner, J.R.; Iyer, M.K.; Dhanasekaran, S.M.; Mehra, R.; Pitchiaya, S.; Tien, J.; Escara-Wilke, J.; Poliakov, A.; et al. Oncogenic Role of THOR; a Conserved Cancer/Testis Long Non-coding RNA. Cell 2017, D171, 1559–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Fang, J.; Xiao, Z.; Deng, J.; Zhang, M.; Gu, L. Downregulation of lncRNA TSLNC8 promotes melanoma resistance to BRAF inhibitor PLX4720 through binding with PP1α to re-activate MAPK signaling. J. Cancer Res. Clin. Oncol. 2021, 147, 767–777. [Google Scholar] [CrossRef]
- Wang, Y.; Li, D.; Lu, J.; Chen, L.; Zhang, S.; Qi, W.; Li, W.; Xu, H. Long noncoding RNA TTN-AS1 facilitates tumorigenesis and metastasis by maintaining TTN expression in skin cutaneous melanoma. Cell Death Dis. 2020, 11, 664. [Google Scholar] [CrossRef]
- Chen, L.L. Linking Long Noncoding RNA Localization and Function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Y.; Li, L.; He, Y.; Zheng, D.; Yu, P.; Yu, L.; Tang, L.; Wang, Y.; Wang, Z. EZH2 RIP-seq Identifies Tissue-specific Long Non-coding RNAs. Curr. Gene Ther. 2018, 18, 275–285. [Google Scholar] [CrossRef]
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Serrano, M.; Lin, A.W.; McCurrach, M.E.; Beach, D.; Lowe, S.W. Oncogenic ras provokes premature cell senescence associated with accumulation of p53 and p16INK4a. Cell 1997, 88, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Michaloglou, C.; Vredeveld, L.C.; Soengas, M.S.; Denoyelle, C.; Kuilman, T.; van der Horst, C.M.; Majoor, D.M.; Shay, J.W.; Mooi, W.J.; Peeper, D.S. BRAFE600-associated senescence-like cell cycle arrest of human naevi. Nature 2005, 436, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Bracken, A.P.; Kleine-Kohlbrecher, D.; Dietrich, N.; Pasini, D.; Gargiulo, G.; Beekman, C.; Theilgaard-Mönch, K.; Minucci, S.; Porse, B.T.; Marine, J.C.; et al. The Polycomb group proteins bind throughout the INK4A-ARF locus and are disassociated in senescent cells. Genes Dev. 2007, 21, 525–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasmant, E.; Laurendeau, I.; Héron, D.; Vidaud, M.; Vidaud, D.; Bièche, I. Characterization of a germ-line deletion, including the entire INK4/ARF locus, in a melanoma-neural system tumor family: Identification of ANRIL, an antisense noncoding RNA whose expression coclusters with ARF. Cancer Res. 2007, 67, 3963–3969. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.B.; Kong, R.; Yin, D.D.; You, L.H.; Sun, M.; Han, L.; Xu, T.P.; Xia, R.; Yang, J.S.; De, W.; et al. Long noncoding RNA ANRIL indicates a poor prognosis of gastric cancer and promotes tumor growth by epigenetically silencing of miR-99a/miR-449a. Oncotarget 2014, 5, 2276–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meseure, D.; Vacher, S.; Lallemand, F.; Alsibai, K.D.; Hatem, R.; Chemlali, W.; Nicolas, A.; De Koning, L.; Pasmant, E.; Callens, C.; et al. Prognostic value of a newly identified MALAT1 alternatively spliced transcript in breast cancer. Br. J. Cancer. 2016, 114, 1395–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Wang, H.; Pan, H.; Shi, Y.; Li, T.; Ge, S.; Jia, R.; Zhang, H.; Fan, X. ANRIL lncRNA triggers efficient therapeutic efficacy by reprogramming the aberrant INK4-hub in melanoma. Cancer Lett. 2016, 381, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Oghabian, A.; Bodiyabadu, P.K.; Joseph, W.R.; Leung, E.Y.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Multiple Isoforms of ANRIL in Melanoma Cells: Structural Complexity Suggests Variations in Processing. Int. J. Mol. Sci. 2017, 18, 1378. [Google Scholar] [CrossRef] [PubMed]
- Lessard, L.; Liu, M.; Marzese, D.M.; Wang, H.; Chong, K.; Kawas, N.; Donovan, N.C.; Kiyohara, E.; Hsu, S.; Nelson, N.; et al. The CASC15 Long Intergenic Noncoding RNA Locus Is Involved in Melanoma Progression and Phenotype Switching. J. Invest. Dermatol. 2015, 135, 2464–2474. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zhang, L.; Huo, X.S.; Yuan, J.H.; Xu, D.; Yuan, S.X.; Zhu, N.; Zhou, W.P.; Yang, G.S.; Wang, Y.Z.; et al. Long noncoding RNA high expression in hepatocellular carcinoma facilitates tumor growth through enhancer of zeste homolog 2 in humans. Hepatology 2011, 54, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Tetzlaff, M.T.; Cui, R.; Xu, X. miR-200c inhibits melanoma progression and drug resistance through down-regulation of BMI-1. Am. J. Pathol. 2012, 181, 1823–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, D.; Shi, W.; Shao, Y.; Li, P.; Song, G. Long non-coding RNA GAS5 inhibits tumorigenesis via miR-137 in melanoma. Am. J. Transl. Res. 2017, 9, 1509–1520. [Google Scholar] [PubMed]
- Chang, C.; Lee, S.O.; Yeh, S.; Chang, T.M. Androgen receptor (AR) differential roles in hormone-related tumors including prostate, bladder, kidney, lung, breast and liver. Oncogene 2014, 33, 3225–3234. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Weidmann, C.A.; Hilimire, T.A.; Yee, E.; Hatfield, B.M.; Schneekloth, J.S., Jr.; Weeks, K.M.; Novina, C.D. Targeting the Oncogenic Long Non-coding RNA SLNCR1 by Blocking Its Sequence-Specific Binding to the Androgen Receptor. Cell Rep. 2020, 30, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ding, N.; Li, Y.; Cheng, H.; Wang, D.; Yang, Q.; Deng, Y.; Yang, Y.; Li, Y.; Ruan, X.; et al. Insulin-like growth factor binding protein 5 (IGFBP5) functions as a tumor suppressor in human melanoma cells. Oncotarget 2015, 6, 20636–20649. [Google Scholar] [CrossRef] [Green Version]
- Hoek, K.S.; Eichhoff, O.M.; Schlegel, N.C.; Döbbeling, U.; Kobert, N.; Schaerer, L.; Hemmi, S.; Dummer, R. In vivo switching of human melanoma cells between proliferative and invasive states. Cancer Res. 2008, 68, 650–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Shen, S.K.; Xiong, J.G.; Xu, Y.; Zhang, H.Q.; Liu, H.J.; Lu, Z.G. Clinical significance of long noncoding RNA SPRY4-IT1 in melanoma patients. FEBS Open Bio 2016, 6, 147–154. [Google Scholar] [CrossRef]
- Khaitan, D.; Dinger, M.E.; Mazar, J.; Crawford, J.; Smith, M.A.; Mattick, J.S.; Perera, R.J. The melanoma-upregulated long noncoding RNA SPRY4-IT1 modulates apoptosis and invasion. Cancer Res. 2011, 71, 3852–3862. [Google Scholar] [CrossRef] [Green Version]
- Garraway, L.A.; Widlund, H.R.; Rubin, M.A.; Getz, G.; Berger, A.J.; Ramaswamy, S.; Beroukhim, R.; Milner, D.A.; Granter, S.R.; Du, J.; et al. Integrative genomic analyses identify MITF as a lineage survival oncogene amplified in malignant melanoma. Nature 2005, 436, 117–122. [Google Scholar] [CrossRef]
- Fogal, V.; Richardson, A.D.; Karmali, P.P.; Scheffler, I.E.; Smith, J.W.; Ruoslahti, E. Mitochondrial p32 protein is a critical regulator of tumor metabolism via maintenance of oxidative phosphorylation. Mol. Cell Biol. 2010, 30, 1303–1318. [Google Scholar] [CrossRef] [Green Version]
- Yagi, M.; Uchiumi, T.; Takazaki, S.; Okuno, B.; Nomura, M.; Yoshida, S.; Kanki, T.; Kang, D. p32/gC1qR is indispensable for fetal development and mitochondrial translation: Importance of its RNA-binding ability. Nucleic Acids Res. 2012, 40, 9717–9737. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Crawford, S.A.; Henstridge, D.C.; Ng, I.H.; Boey, E.J.; Xu, Y.; Febbraio, M.A.; Jans, D.A.; Bogoyevitch, M.A. p32 protein levels are integral to mitochondrial and endoplasmic reticulum morphology, cell metabolism and survival. Biochem. J. 2013, 453, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Fogal, V.; Zhang, L.; Krajewski, S.; Ruoslahti, E. Mitochondrial/cell-surface protein p32/gC1qR as a molecular target in tumor cells and tumor stroma. Cancer Res. 2008, 68, 7210–7218. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–388. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, C.K.; Mosimann, C.; Fan, Z.P.; Yang, S.; Thomas, A.J.; Ablain, J.; Tan, J.L.; Fogley, R.D.; van Rooijen, E.; Hagedorn, E.J. A zebrafish melanoma model reveals emergence of neural crest identity during melanoma initiation. Science 2016, 351, aad2197. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, Z.; Liu, L.; Wang, Q.; Li, S.; Chen, D.; Hu, Z.; Yu, T.; Ding, J.; Li, J.; et al. Long noncoding RNA TSLNC8 is a tumor suppressor that inactivates the interleukin-6/STAT3 signaling pathway. Hepatology 2018, 67, 171–187. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Yu, X. Long noncoding RNA TSLNC8 suppresses cell proliferation and metastasis and promotes cell apoptosis in human glioma. Mol. Med. Rep. 2018, 18, 5536–5544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, M.; Li, Z.; Sacks, D.B. IQGAP1 is a scaffold for mitogen-activated protein kinase signaling. Mol. Cell Biol. 2005, 25, 7940–7952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, K.L.; Mazur, P.K.; Zehnder, A.M.; Zhang, J.; Zarnegar, B.; Sage, J.; Khavari, P.A. IQGAP1 scaffold-kinase interaction blockade selectively targets RAS-MAP kinase-driven tumors. Nat. Med. 2013, 19, 626–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Functional interactions among microRNAs and long noncoding RNAs. Semin. Cell. Dev. Biol. 2014, 34, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalali, S.; Bhartiya, D.; Lalwani, M.K.; Sivasubbu, S.; Scaria, V. Systematic transcriptome wide analysis of lncRNA-miRNA interactions. PLoS ONE 2013, 8, e53823. [Google Scholar] [CrossRef] [Green Version]
- Ragusa, M.; Barbagallo, C.; Brex, D.; Caponnetto, A.; Cirnigliaro, M.; Battaglia, R.; Barbagallo, D.; Di Pietro, C.; Purrello, M. Molecular Crosstalking among Noncoding RNAs: A New Network Layer of Genome Regulation in Cancer. Int. J. Genom. 2017, 2017, 4723193. [Google Scholar] [CrossRef]
- Liu, J.; Liu, T.; Wang, X.; He, A. Circles reshaping the RNA world: From waste to treasure. Mol. Cancer 2017, 16, 58. [Google Scholar] [CrossRef] [Green Version]
- Cocquerelle, C.; Mascrez, B.; Hétuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. 1993, 7, 155–1560. [Google Scholar] [CrossRef] [Green Version]
- Pasman, Z.; Been, M.D.; Garcia-Blanco, M.A. Exon circularization in mammalian nuclear extracts. RNA 1996, 2, 603–610. [Google Scholar]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Salzman, J. Circular RNA Expression: Its Potential Regulation and Function. Trends Genet. 2016, 32, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.; Xing, J.; Wang, Z.; Chen, L.; Cui, M.; Jiang, B. microRNAs and ceRNAs: RNA networks in pathogenesis of cancer. Chin. J. Cancer. Res. 2013, 25, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell. 2017, 66, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, B.; Zheng, Y.; Ma, S.; Xing, Q.; Wang, X.; Yang, B.; Yin, G.; Guan, F. BANCR contributes to the growth and invasion of melanoma by functioning as a competing endogenous RNA to upregulate Notch2 expression by sponging miR-204. Int. J. Oncol. 2017, 51, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qian, W.; Feng, F.; Cao, Q.; Li, Y.; Hou, Y.; Zhang, L.; Fan, J. Upregulated lncRNA CASC2 May Inhibit Malignant Melanoma Development Through Regulating miR-18a-5p/RUNX1. Oncol. Res. 2019, 27, 371–377. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, X.; Zhou, H.; Dan, X.; Jiang, L.; Wu, Y. Long non-coding RNA CASC2 inhibits tumorigenesis via the miR-181a/PLXNC1 axis in melanoma. Acta Biochim. Biophys. Sin. 2018, 50, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Lv, L.; Jia, J.Q.; Chen, J. The lncRNA CCAT1 Upregulates Proliferation and Invasion in Melanoma Cells via Suppressing miR-33a. Oncol. Res. 2018, 26, 201–208. [Google Scholar] [CrossRef]
- Fan, J.; Kang, X.; Zhao, L.; Zheng, Y.; Yang, J.; Li, D. Long Noncoding RNA CCAT1 Functions as a Competing Endogenous RNA to Upregulate ITGA9 by Sponging MiR-296-3p in Melanoma. Cancer Manag. Res. 2020, 12, 4699–4714. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.; Linbo, L.; Xiaomei, Z.; Hui, P. Circ_0002770, acting as a competitive endogenous RNA, promotes proliferation and invasion by targeting miR-331-3p in melanoma. Cell. Death. Dis. 2020, 11, 264. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Wang, S.S.; Wang, J.; Su, H.L.; Xu, J.H. CircRNA_0016418 expedites the progression of human skin melanoma via miR-625/YY1 axis. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10918–10930. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.Y.; Zhu, M.X.; Lu, N.H.; Liu, J.Q.; Yang, Y.W.; Zhang, Y.; Shi, Y.D.; Feng, Z.H.; Li, J.X.; Qi, F.Z.; et al. Circular RNA circ_0020710 drives tumor progression and immune evasion by regulating the miR-370-3p/CXCL12 axis in melanoma. Mol. Cancer 2020, 19, 84. [Google Scholar] [CrossRef]
- Bian, D.; Wu, Y.; Song, G. Novel circular RNA, hsa_circ_0025039 promotes cell growth, invasion and glucose metabolism in malignant melanoma via the miR-198/CDK4 axis. Biomed. Pharmacother. 2018, 108, 165–176. [Google Scholar] [CrossRef]
- Zhao, F.; Jia, Z.; Feng, Y.; Li, Z.; Feng, J. Circular RNA circ_0079593 enhances malignant melanoma progression by the regulation of the miR-573/ABHD2 axis. J. Dermatol. Sci. 2021, 102, 7–15. [Google Scholar] [CrossRef]
- Luan, W.; Shi, Y.; Zhou, Z.; Xia, Y.; Wang, J. circRNA_0084043 promote malignant melanoma progression via miR-153-3p/Snail axis. Biochem. Biophys. Res. Commun. 2018, 502, 22–29. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, J.; Wa, Q.; He, M.; Wang, X.; Zhou, J.; Cen, Y. Knockdown of circ_0084043 suppresses the development of human melanoma cells through miR-429/tribbles homolog 2 axis and Wnt/β-catenin pathway. Life Sci. 2020, 243, 117323. [Google Scholar] [CrossRef]
- Wan, N.; Yang, W.; Cheng, H.; Wang, J. FOXD3-AS1 Contributes to the Progression of Melanoma Via miR-127-3p/FJX1 Axis. Cancer Biother. Radiopharm. 2020, 35, 596–604. [Google Scholar] [CrossRef]
- Chen, X.; Gao, J.; Yu, Y.; Zhao, Z.; Pan, Y. LncRNA FOXD3-AS1 promotes proliferation; invasion and migration of cutaneous malignant melanoma via regulating miR-325/MAP3K2. Biomed. Pharmacother. 2019, 120, 109438. [Google Scholar] [CrossRef] [PubMed]
- Luan, W.; Zhou, Z.; Ni, X.; Xia, Y.; Wang, J.; Yan, Y.; Xu, B. Long non-coding RNA H19 promotes glucose metabolism and cell growth in malignant melanoma via miR-106a-5p/E2F3 axis. J. Cancer. Res. Clin. Oncol. 2018, 144, 531–542. [Google Scholar] [CrossRef]
- An, L.F.; Huang, J.W.; Han, X.; Wang, J. Downregulation of lncRNA H19 sensitizes melanoma cells to cisplatin by regulating the miR-18b/IGF1 axis. Anticancer Drugs 2020, 31, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Luan, W.; Li, R.; Liu, L.; Ni, X.; Shi, Y.; Xia, Y.; Wang, J.; Lu, F.; Xu, B. Long non-coding RNA HOTAIR acts as a competing endogenous RNA to promote malignant melanoma progression by sponging miR-152-3p. Oncotarget 2017, 8, 85401–85414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Zhang, Q.; Wang, H.; Chang, P.; Tao, K. KCNQ1OT1 promotes melanoma growth and metastasis. Aging 2018, 10, 632–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Q.; Liu, L.; Pei, H.; Zhang, J.; Chen, M.; Zhai, X. A LHFPL3-AS1/miR-580-3p/STAT3 Feedback Loop Promotes the Malignancy in Melanoma via Activation of JAK2/STAT3 Signaling. Mol. Cancer. Res. 2020, 18, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wan, H.; Zhang, X. LncRNA LHFPL3-AS1 contributes to tumorigenesis of melanoma stem cells via the miR-181a-5p/BCL2 pathway. Cell Death Dis. 2020, 11, 950. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, S.; Wong, R.P.; Ardekani, G.S.; Zhang, G.; Martinka, M.; Ong, C.J.; Li, G. Role of EIF5A2; a downstream target of Akt, in promoting melanoma cell invasion. Br. J. Cancer. 2014, 110, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Xu, W.; Zheng, Z.; Cao, Z. LINC00459 sponging miR-218 to elevate DKK3 inhibits proliferation and invasion in melanoma. Sci. Rep. 2019, 9, 19139. [Google Scholar] [CrossRef]
- Liu, Y.; He, D.; Xiao, M.; Zhu, Y.; Zhou, J.; Cao, K. Long noncoding RNA LINC00518 induces radioresistance by regulating glycolysis through an miR-33a-3p/HIF-1α negative feedback loop in melanoma. Cell Death Dis. 2021, 12, 245. [Google Scholar] [CrossRef]
- Luan, W.; Ding, Y.; Ma, S.; Ruan, H.; Wang, J.; Lu, F. Long noncoding RNA LINC00518 acts as a competing endogenous RNA to promote the metastasis of malignant melanoma via miR-204-5p/AP1S2 axis. Cell Death Dis. 2019, 10, 855. [Google Scholar] [CrossRef]
- Mu, X.; Mou, K.H.; Ge, R.; Han, D.; Zhou, Y.; Wang, L.J. Linc00961 inhibits the proliferation and invasion of skin melanoma by targeting the miR-367/PTEN axis. Int. J. Oncol. 2019, 55, 708–720. [Google Scholar] [CrossRef]
- Wu, K.; Wang, Q.; Liu, Y.L.; Xiang, Z.; Wang, Q.Q.; Yin, L.; Liu, S.L. LncRNA POU3F3 Contributes to Dacarbazine Resistance of Human Melanoma Through the MiR-650/MGMT Axis. Front. Oncol. 2021, 11, 643613. [Google Scholar] [CrossRef]
- Li, F.; Li, X.; Qiao, L.; Liu, W.; Xu, C.; Wang, X. MALAT1 regulates miR-34a expression in melanoma cells. Cell Death Dis. 2019, 10, 389. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, H.; Wang, G.; Yu, G.; Zhang, D.; Wang, Y.; Fan, W.; Yang, W. Deregulation of miR-183 promotes melanoma development via lncRNA MALAT1 regulation and ITGB1 signal activation. Oncotarget 2017, 8, 3509–3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, W.; Li, L.; Shi, Y.; Bu, X.; Xia, Y.; Wang, J.; Djangmah, H.S.; Liu, X.; You, Y.; Xu, B. Long non-coding RNA MALAT1 acts as a competing endogenous RNA to promote malignant melanoma growth and metastasis by sponging miR-22. Oncotarget 2016, 7, 63901–63912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.; Pi, X. lncRNA-MEG3 Suppresses the Proliferation and Invasion of Melanoma by Regulating CYLD Expression Mediated by Sponging miR-499-5p. Biomed. Res. Int. 2018, 2018, 2086564. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhu, L.; Li, Y.; Zheng, Z.; Lin, X.; Yang, C. LncRNA MEG3 promotes melanoma growth, metastasis and formation through modulating miR-21/E-cadherin axis. Cancer Cell Int. 2020, 20, 12. [Google Scholar] [CrossRef]
- Chen, X.; Dong, H.; Liu, S.; Yu, L.; Yan, D.; Yao, X.; Sun, W.; Han, D.; Gao, G. Long noncoding RNA MHENCR promotes melanoma progression via regulating miR-425/489-mediated PI3K-Akt pathway. Am. J. Transl. Res. 2017, 9, 90–102. [Google Scholar]
- Huo, J.; Wang, Y.; Zhang, Y.; Wang, W.; Yang, P.; Zhao, W.; Zhang, M.; Cui, L.; Zhang, D. The LncRNA MIR155HG is Upregulated by SP1 in Melanoma Cells and Drives Melanoma Progression via Modulating the MiR-485-3p/PSIP1 Axis. Anticancer Agents Med. Chem. 2021. [Google Scholar] [CrossRef]
- Guo, J.; Gan, Q.; Gan, C.; Zhang, X.; Ma, X.; Dong, M. LncRNA MIR205HG regulates melanomagenesis via the miR-299-3p/VEGFA axis. Aging 2021, 13, 5297. [Google Scholar] [CrossRef]
- Ding, F.; Lai, J.; Gao, Y.; Wang, G.; Shang, J.; Zhang, D.; Zheng, S. NEAT1/miR-23a-3p/KLF3: A novel regulatory axis in melanoma cancer progression. Cancer Cell Int. 2019, 19, 217. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.J.; Wang, H.Y.; Zhang, J.; Dai, H.Y.; Yao, Z.X.; Zheng, Z.; Meng-Yan, S.; Wu, K. NEAT1/miR-200b-3p/SMAD2 axis promotes progression of melanoma. Aging 2020, 12, 22759–22775. [Google Scholar] [CrossRef]
- Xia, Y.; Zhou, Y.; Han, H.; Li, P.; Wei, W.; Lin, N. lncRNA NEAT1 facilitates melanoma cell proliferation, migration, and invasion via regulating miR-495-3p and E2F3. J. Cell Physiol. 2019, 234, 19592–19601. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, K.; Li, J.; Wang, A.; Sun, L.; Tang, J.; Xiong, W.; Zhou, X.; Chen, X.; Zhou, J.; et al. Overexpression of long non-coding RNA NORAD promotes invasion and migration in malignant melanoma via regulating the MIR-205-EGLN2 pathway. Cancer Med. 2019, 8, 1744–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.; Menggen, Q.; Wuren, Q.; Shi, Q.; Pi, X. Long Noncoding RNA Taurine-Upregulated Gene1 (TUG1) Promotes Tumor Growth and Metastasis Through TUG1/Mir-129-5p/Astrocyte-Elevated Gene-1 (AEG-1) Axis in Malignant Melanoma. Med. Sci. Monit. 2018, 24, 1547–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, G.; Ren, L.; Wang, K.; Liu, A. Long non-coding RNA TUG1 recruits miR-29c-3p from its target gene RGS1 to promote proliferation and metastasis of melanoma cells. Int. J. Oncol. 2019, 54, 1317–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.; Tang, F.; Chen, J.; Xu, D.; Li, X.; Xu, Y.; Wang, S.; Zhou, J. Knockdown of lncRNA-UCA1 inhibits the proliferation and migration of melanoma cells through modulating the miR-28-5p/HOXB3 axis. Exp. Ther. Med. 2019, 17, 4294–4302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Sun, Q.; Zhao, L.; Wu, J.; Chen, X.; Wang, Y.; Zang, W.; Zhao, G. LncRNA UCA1-miR-507-FOXM1 axis is involved in cell proliferation, invasion and G0/G1 cell cycle arrest in melanoma. Med. Oncol. 2016, 33, 88. [Google Scholar] [CrossRef]
- Hao, Y.Q.; Liu, K.W.; Zhang, X.; Kang, S.X.; Zhang, K.; Han, W.; Li, L.; Li, Z.H. GINS2 was regulated by lncRNA XIST/miR-23a-3p to mediate proliferation and apoptosis in A375 cells. Mol. Cell Biochem. 2021, 476, 1455–1465. [Google Scholar] [CrossRef]
- Tian, K.; Sun, D.; Chen, M.; Yang, Y.; Wang, F.; Guo, T.; Shi, Z. Long Noncoding RNA X-Inactive Specific Transcript Facilitates Cellular Functions in Melanoma via miR-139-5p/ROCK1 Pathway. OncoTargets Ther. 2020, 13, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, R.; Liu, D. Long non-coding RNA ZEB1-AS1 indicates poor prognosis and promotes melanoma progression through targeting miR-1224-5p. Exp. Ther. Med. 2019, 17, 857–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Deng, J.; Zhang, L.; Zhao, J.; Zhou, F.; Liu, N.; Cai, R.; Wu, J.; Shu, B.; Qi, S. Reconstruction of lncRNA-miRNA-mRNA network based on competitive endogenous RNA reveals functional lncRNAs in skin cutaneous melanoma. BMC Cancer 2020, 20, 927. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, A.; Zhao, F.; Pechlivanis, S.; Eberle, J.; Steinle, A.; Diederichs, S.; Schadendorf, D.; Paschen, A. Tumor suppressive microRNAs miR-34a/c control cancer cell expression of ULBP2, a stress-induced ligand of the natural killer cell receptor NKG2D. Cancer Res. 2012, 72, 460–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, X.; Hao, Y.; Fang, Z.; He, Y. Potential roles of abnormally expressed long noncoding RNA UCA1 and Malat-1 in metastasis of melanoma. Melanoma Res. 2014, 24, 335–341. [Google Scholar] [CrossRef]
- Li, R.; Zhang, L.; Jia, L.; Duan, Y.; Li, Y.; Bao, L.; Sha, N. Long non-coding RNA BANCR promotes proliferation in malignant melanoma by regulating MAPK pathway activation. PLoS ONE 2014, 9, e100893. [Google Scholar] [CrossRef] [Green Version]
- Flockhart, R.J.; Webster, D.E.; Qu, K.; Mascarenhas, N.; Kovalski, J.; Kretz, M.; Khavari, P.A. BRAFV600E remodels the melanocyte transcriptome and induces BANCR to regulate melanoma cell migration. Genome Res. 2012, 22, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Galasso, M.; Morrison, C.; Minotti, L.; Corrà, F.; Zerbinati, C.; Agnoletto, C.; Baldassari, F.; Fassan, M.; Bartolazzi, A.; Vecchione, A.; et al. Loss of miR-204 expression is a key event in melanoma. Mol. Cancer 2018, 17, 71. [Google Scholar] [CrossRef] [Green Version]
- Luan, W.; Qian, Y.; Ni, X.; Bu, X.; Xia, Y.; Wang, J.; Ruan, H.; Ma, S.; Xu, B. miR-204-5p acts as a tumor suppressor by targeting matrix metalloproteinases-9 and B-cell lymphoma-2 in malignant melanoma. OncoTargets Ther. 2017, 10, 1237–1246. [Google Scholar] [CrossRef] [Green Version]
- Massi, D.; Tarantini, F.; Franchi, A.; Paglierani, M.; Di Serio, C.; Pellerito, S.; Leoncini, G.; Cirino, G.; Geppetti, P.; Santucci, M. Evidence for differential expression of Notch receptors and their ligands in melanocytic nevi and cutaneous malignant melanoma. Mod. Pathol. 2006, 19, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.R.; Yang, N. MiR-33a-5p inhibits the growth and metastasis of melanoma cells by targeting SNAI2. Neoplasma 2020, 67, 813–824. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Wei, H.; Tian, H.; Qiu, Y.; Xu, J. miR-33a is downregulated in melanoma cells and modulates cell proliferation by targeting PCTAIRE1. Oncol. Lett. 2016, 11, 2741–2746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.K.; Oommen, S.; Aubry, M.C.; Williams, B.P.; Vlahakis, N.E. Integrin α9β1 promotes malignant tumor growth and metastasis by potentiating epithelial-mesenchymal transition. Oncogene 2013, 32, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.; Li, H.; Gao, F.; Tan, Q. lncRNA H19 predicts poor prognosis in patients with melanoma and regulates cell growth, invasion, migration and epithelial-mesenchymal transition in melanoma cells. OncoTargets Ther. 2018, 11, 3583–3595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Z.; Zhao, J.; Yang, Y. Downregulation of lncRNA H19 inhibits the migration and invasion of melanoma cells by inactivating the NF-κB and PI3K/Akt signaling pathways. Mol. Med. Rep. 2018, 17, 7313–7318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balihodzic, A.; Barth, D.A.; Prinz, F.; Pichler, M. Involvement of Long Non-Coding RNAs in Glucose Metabolism in Cancer. Cancers 2021, 13, 977. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Cantile, M.; Di Bonito, M.; Tracey De Bellis, M.; Botti, G. Functional Interaction among lncRNA HOTAIR and MicroRNAs in Cancer and Other Human Diseases. Cancers 2021, 13, 570. [Google Scholar] [CrossRef]
- Tang, L.; Zhang, W.; Su, B.; Yu, B. Long noncoding RNA HOTAIR is associated with motility, invasion, and metastatic potential of metastatic melanoma. Biomed. Res. Int. 2013, 2013, 251098. [Google Scholar] [CrossRef] [Green Version]
- Cantile, M.; Scognamiglio, G.; Marra, L.; Aquino, G.; Botti, C.; Falcone, M.R.; Malzone, M.G.; Liguori, G.; Di Bonito, M.; Franco, R.; et al. HOTAIR role in melanoma progression and its identification in the blood of patients with advanced disease. J. Cell. Physiol. 2017, 232, 3422–3432. [Google Scholar] [CrossRef]
- Zeng, Q.; Wang, Q.; Chen, X.; Xia, K.; Tang, J.; Zhou, X.; Cheng, Y.; Chen, Y.; Huang, L.; Xiang, H.; et al. Analysis of lncRNAs expression in UVB-induced stress responses of melanocytes. J. Dermatol. Sci. 2016, 81, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Siwecka, N.; Rozpędek, W.; Pytel, D.; Wawrzynkiewicz, A.; Dziki, A.; Dziki, Ł.; Diehl, J.A.; Majsterek, I. Dual role of Endoplasmic Reticulum Stress-Mediated Unfolded Protein Response Signaling Pathway in Carcinogenesis. Int. J. Mol. Sci. 2019, 20, 4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Liu, L.; Zheng, M.; Li, X.; Wu, A.; Wu, Q.; Liao, C.; Zou, J.; Song, H. MEKK2 and MEKK3 suppress Hedgehog pathway-dependent medulloblastoma by inhibiting GLI1 function. Oncogene 2018, 37, 3864–3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenblatt, M.B.; Shin, D.Y.; Oh, H.; Lee, K.Y.; Zhai, B.; Gygi, S.P.; Lotinun, S.; Baron, R.; Liu, D.; Su, B.; et al. MEKK2 mediates an alternative β-catenin pathway that promotes bone formation. Proc. Natl. Acad. Sci. USA 2016, 113, E1226–E1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Greene, N.T.; Means, A.L.; Lu, P.; Jiang, A.; Schmidt, C.R.; Chakravarthy, A.B.; Merchant, N.B.; Washington, M.K.; Zhang, B.; Shyr, Y.; et al. Four jointed box 1 promotes angiogenesis and is associated with poor patient survival in colorectal carcinoma. PLoS ONE 2013, 8, e69660. [Google Scholar] [CrossRef] [Green Version]
- Luan, W.; Ding, Y.; Yuan, H.; Ma, S.; Ruan, H.; Wang, J.; Lu, F.; Bu, X. Long non-coding RNA LINC00520 promotes the proliferation and metastasis of malignant melanoma by inducing the miR-125b-5p/EIF5A2 axis. J. Exp. Clin. Cancer Res. 2020, 39, 96. [Google Scholar] [CrossRef]
- Siena, Á.D.D.; Plaça, J.R.; Araújo, L.F.; de Barros, I.I.; Peronni, K.; Molfetta, G.; de Biagi, C.A.O., Jr.; Espreafico, E.M.; Sousa, J.F.; Silva, W.A., Jr. Whole transcriptome analysis reveals correlation of long noncoding RNA ZEB1-AS1 with invasive profile in melanoma. Sci. Rep. 2019, 9, 11350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caenepeel, S.; Cooke, K.; Wadsworth, S.; Huang, G.; Robert, L.; Moreno, B.H.; Parisi, G.; Cajulis, E.; Kendall, R.; Beltran, P.; et al. MAPK pathway inhibition induces MET and GAB1 levels, priming BRAF mutant melanoma for rescue by hepatocyte growth factor. Oncotarget 2017, 8, 17795–17809. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Cai, J.; Wang, Q.; Wang, Y.; Liu, M.; Yang, J.; Zhou, J.; Kang, C.; Li, M.; Jiang, C. Long Noncoding RNA NEAT1, Regulated by the EGFR Pathway, Contributes to Glioblastoma Progression Through the WNT/β-Catenin Pathway by Scaffolding EZH2. Clin. Cancer. Res. 2018, 24, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Peng, C.; Li, D.; Zhang, D.; Li, X.; Cui, F.; Chen, Y.; He, Q. E2F3 promotes cancer growth and is overexpressed through copy number variation in human melanoma. OncoTargets Ther. 2018, 11, 5303–5313. [Google Scholar] [CrossRef] [Green Version]
- Lo, R.S.; Witte, O.N. Transforming growth factor-beta activation promotes genetic context-dependent invasion of immortalized melanocytes. Cancer Res. 2008, 68, 4248–4257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Gao, Y.; Li, J.; Zhou, Y.; Yuan, J.; Guan, H.; Yao, P. LncRNA MEG3 repressed malignant melanoma progression via inactivating Wnt signaling pathway. J. Cell. Biochem. 2018, 119, 7498–7505. [Google Scholar] [CrossRef] [PubMed]
- Urbanik, T.; Koehler, B.C.; Wolpert, L.; Elßner, C.; Scherr, A.L.; Longerich, T.; Kautz, N.; Welte, S.; Hövelmeyer, N.; Jäger, D.; et al. CYLD deletion triggers nuclear factor-κB-signaling and increases cell death resistance in murine hepatocytes. World J. Gastroenterol. 2014, 20, 17049–17064. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Jono, H.; Komatsu, K.; Woo, C.H.; Lee, J.; Miyata, M.; Matsuno, T.; Xu, X.; Huang, Y.; Zhang, W.; et al. CYLD negatively regulates transforming growth factor-β-signalling via deubiquitinating Akt. Nat. Commun. 2012, 3, 771. [Google Scholar] [CrossRef] [Green Version]
- Kourtidis, A.; Lu, R.; Pence, L.J.; Anastasiadis, P.Z. A central role for cadherin signaling in cancer. Exp. Cell. Res. 2017, 358, 378–385. [Google Scholar] [CrossRef]
- Chen, Y.; Soong, J.; Mohanty, S.; Xu, L.; Scott, G. The neural guidance receptor Plexin C1 delays melanoma progression. Oncogene 2013, 32, 4941–4949. [Google Scholar] [CrossRef] [Green Version]
- Kuphal, S.; Lodermeyer, S.; Bataille, F.; Schuierer, M.; Hoang, B.H.; Bosserhoff, A.K. Expression of Dickkopf genes is strongly reduced in malignant melanoma. Oncogene 2006, 25, 5027–5036. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.F.; Yan, S.; Wu, S.F. MicroRNA-153-3p suppress cell proliferation and invasion by targeting SNAI1 in melanoma. Biochem. Biophys. Res. Commun. 2017, 487, 140–145. [Google Scholar] [CrossRef]
- Wang, J.; Park, J.S.; Wei, Y.; Rajurkar, M.; Cotton, J.L.; Fan, Q.; Lewis, B.C.; Ji, H.; Mao, J. TRIB2 acts downstream of Wnt/TCF in liver cancer cells to regulate YAP and C/EBPα function. Mol. Cell. 2013, 51, 211–252. [Google Scholar] [CrossRef] [Green Version]
- Varum, S.; Baggiolini, A.; Zurkirchen, L.; Atak, Z.K.; Cantù, C.; Marzorati, E.; Bossart, R.; Wouters, J.; Häusel, J.; Tuncer, E.; et al. Yin Yang 1 Orchestrates a Metabolic Program Required for Both Neural Crest Development and Melanoma Formation. Cell. Stem. Cell. 2019, 24, 637–653. [Google Scholar] [CrossRef] [Green Version]
- Thomson, D.W.; Dinger, M.E. Endogenous microRNA sponges: Evidence and controversy. Nat. Rev. Genet. 2016, 17, 272–283. [Google Scholar] [CrossRef]
- Denzler, R.; Agarwal, V.; Stefano, J.; Stoffel, M. Assessing the ceRNA Hypothesis with Quantitative Measurements of miRNA and Target Abundance. Mol. Cell 2014, 54, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Denzler, R.; McGeary, S.E.; Title, A.C.; Agarwal, V.; Bartel, D.P.; Stoffel, M. Impact of MicroRNA Levels, Target-Site Complementarity, and Cooperativity on Competing Endogenous RNA-Regulated Gene Expression. Mol. Cell 2016, 64, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.Y.; Kingston, E.R.; Kleaveland, B.; Lin, D.H.; Stubna, M.W.; Bartel, D.P. The ZSWIM8 ubiquitin ligase mediates target-directed microRNA degradation. Science 2020, 370, eabc9359. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Shao, T.J.; Bofill-De Ros, X.; Lian, C.; Villanueva, P.; Dai, L.; Gu, S. AGO-bound mature miRNAs are oligouridylated by TUTs and subsequently degraded by DIS3L2. Nat. Commun. 2020, 11, 2765. [Google Scholar] [CrossRef] [PubMed]
- Kleaveland, B.; Shi, C.Y.; Stefano, J.; Bartel, D.P. A Network of Noncoding Regulatory RNAs Acts in the Mammalian Brain. Cell 2018, 174, 350–362.e17. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 35, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecozzi, N.; Vera, O.; Karreth, F.A. Squaring the circle: circRNAs in melanoma. Oncogene 2021, 40, 5559–5566. [Google Scholar] [CrossRef]
- Li, L.; Wang, D.; Xue, M.; Mi, X.; Liang, Y.; Wang, P. 3’UTR shortening identifies high-risk cancers with targeted dysregulation of the ceRNA network. Sci. Rep. 2014, 4, 5406. [Google Scholar] [CrossRef]
- Guttman, M.; Rinn, J.L. Modular regulatory principles of large non-coding RNAs. Nature 2012, 482, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.Y.; Sirey, T.; Honti, F.; Graham, B.; Piovesan, A.; Merkenschlager, M.; Webber, C.; Ponting, C.P.; Marques, A.C. Extensive microRNA-mediated crosstalk between lncRNAs and mRNAs in mouse embryonic stem cells. Genome Res. 2015, 25, 655–666. [Google Scholar] [CrossRef] [Green Version]
- Gloss, B.S.; Dinger, M.E. The specificity of long noncoding RNA expression. Biochim. Biophys. Acta 2016, 1859, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.B.; Johnston, R.L.; Inostroza-Ponta, M.; Fox, A.H.; Fortini, E.; Moscato, P.; Dinger, M.E.; Mattick, J.S. Genome-wide analysis of long noncoding RNA stability. Genome Res. 2012, 22, 885–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Yang, S.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Xu, J.; Xia, K.; Chang, Y.; et al. Emerging role of exosome-derived long non-coding RNAs in tumor microenvironment. Mol. Cancer 2018, 17, 82. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Lin, X.; Zhang, L.; Hong, W.; Zhang, Y. Long noncoding RNA X-inactive specific transcript promotes malignant melanoma progression and oxaliplatin resistance. Melanoma Res. 2019, 29, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Ursic, K.; Kos, S.; Kamensek, U.; Cemazar, M.; Scancar, J.; Bucek, S.; Kranjc, S.; Staresinic, B.; Sersa, G. Comparable effectiveness and immunomodulatory actions of oxaliplatin and cisplatin in electrochemotherapy of murine melanoma. Bioelectrochemistry 2018, 119, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, T.; Rutkowski, P.; Michalak, M.; Kozak, K.; Guglas, K.; Ryś, M.; Galus, Ł.; Woźniak, S.; Ługowska, I.; Gos, A.; et al. Plasma lncRNA expression profile as a prognostic tool in BRAF-mutant metastatic melanoma patients treated with BRAF inhibitor. Oncotarget 2019, 10, 3879–3893. [Google Scholar] [CrossRef] [Green Version]
- Sousa, J.F.; Torrieri, R.; Silva, R.R.; Pereira, C.G.; Valente, V.; Torrieri, E.; Peronni, K.C.; Martins, W.; Muto, N.; Francisco, G.; et al. Novel primate-specific genes, RMEL 1, 2 and 3, with highly restricted expression in melanoma, assessed by new data mining tool. PLoS ONE 2010, 5, 13510. [Google Scholar] [CrossRef]
- Cardoso, C.; Serafim, R.B.; Kawakami, A.; Gonçalves Pereira, C.; Roszik, J.; Valente, V.; Vazquez, V.L.; Fisher, D.E.; Espreafico, E.M. The lncRNA RMEL3 protects immortalized cells from serum withdrawal-induced growth arrest and promotes melanoma cell proliferation and tumor growth. Pigment Cell Melanoma Res. 2019, 32, 303–314. [Google Scholar] [CrossRef]
- Goedert, L.; Pereira, C.G.; Roszik, J.; Plaça, J.R.; Cardoso, C.; Chen, G.; Deng, W.; Yennu-Nanda, V.G.; Silva, W.A., Jr.; Davies, M.A.; et al. RMEL3, a novel BRAFV600E-associated long noncoding RNA, is required for MAPK and PI3K signaling in melanoma. Oncotarget 2016, 7, 36711–36718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Sabnis, A.J.; Chan, E.; Olivas, V.; Cade, L.; Pazarentzos, E.; Asthana, S.; Neel, D.; Yan, J.J.; Lu, X.; et al. The Hippo effector YAP promotes resistance to RAF- and MEK-targeted cancer therapies. Nat. Genet. 2015, 47, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Joung, J.; Engreitz, J.M.; Konermann, S.; Abudayyeh, O.O.; Verdine, V.K.; Aguet, F.; Gootenberg, J.S.; Sanjana, N.E.; Wright, J.B.; Fulco, C.P.; et al. Genome-scale activation screen identifies a lncRNA locus regulating a gene neighbourhood. Nature 2017, 548, 343–346. [Google Scholar] [CrossRef]
- Atianand, M.K.; Caffrey, D.R.; Fitzgerald, K.A. Immunobiology of Long Noncoding RNAs. Annu. Rev. Immunol. 2017, 35, 177–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Satpathy, A.T.; Chang, H.Y. Gene regulation in the immune system by long noncoding RNAs. Nat. Immunol. 2017, 18, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Flores-Concha, M.; Oñate, Á.A. Long Non-coding RNAs in the Regulation of the Immune Response and Trained Immunity. Front. Genet. 2020, 11, 718. [Google Scholar] [CrossRef] [PubMed]
- Ping, S.; Wang, S.; He, J.; Chen, J. Identification and Validation of Immune-Related lncRNA Signature as a Prognostic Model for Skin Cutaneous Melanoma. Pharmgenomics Pers. Med. 2021, 14, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ba, H.J.; Wen, X.Z.; Zhou, M.; Küçük, C.; Tamagnone, L.; Wei, L.; You, H. A prognostic model for melanoma patients on the basis of immune-related lncRNAs. Aging 2021, 13, 6554–6564. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Chen, Z.; Chen, Y.; Wang, X.; Tang, N. MIR155HG is a prognostic biomarker and associated with immune infiltration and immune checkpoint molecules expression in multiple cancers. Cancer Med. 2019, 8, 7161–7173. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Yan, Y.; Ren, Y.; Hu, Y.; Wang, Y.; Chen, L.; Zhi, Z.; Zheng, Y.; Shao, Y.; Liu, J. LncRNA SAMMSON Mediates Adaptive Resistance to RAF Inhibition in BRAF-Mutant Melanoma Cells. Cancer Res. 2021, 81, 2918–2929. [Google Scholar] [CrossRef] [PubMed]
- Vidaurre, S.; Fitzpatrick, C.; Burzio, V.A.; Briones, M.; Villota, C.; Villegas, J.; Echenique, J.; Oliveira-Cruz, L.; Araya, M.; Borgna, V.; et al. Down-regulation of the antisense mitochondrial non-coding RNAs (ncRNAs) is a unique vulnerability of cancer cells and a potential target for cancer therapy. J. Biol. Chem. 2014, 289, 27182–27198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobos-González, L.; Silva, V.; Araya, M.; Restovic, F.; Echenique, J.; Oliveira-Cruz, L.; Fitzpatrick, C.; Briones, M.; Villegas, J.; Villota, C.; et al. Targeting antisense mitochondrial ncRNAs inhibits murine melanoma tumor growth and metastasis through reduction in survival and invasion factors. Oncotarget 2016, 7, 58331–58350. [Google Scholar] [CrossRef] [PubMed]
- Varas-Godoy, M.; Lladser, A.; Farfan, N.; Villota, C.; Villegas, J.; Tapia, J.C.; Burzio, L.O.; Burzio, V.A.; Valenzuela, P.D.T. In vivo knockdown of antisense non-coding mitochondrial RNAs by a lentiviral-encoded shRNA inhibits melanoma tumor growth and lung colonization. Pigment Cell Melanoma Res. 2018, 31, 64–72. [Google Scholar] [CrossRef]
- Kalhori, M.R.; Khodayari, H.; Khodayari, S.; Vesovic, M.; Jackson, G.; Farzaei, M.H.; Bishayee, A. Regulation of Long Non-Coding RNAs by Plant Secondary Metabolites: A Novel Anticancer Therapeutic Approach. Cancers 2021, 13, 1274. [Google Scholar] [CrossRef] [PubMed]
- Arun, G.; Diermeier, S.D.; Spector, D.L. Therapeutic Targeting of Long Non-Coding RNAs in Cancer. Trends Mol. Med. 2018, 24, 257–277. [Google Scholar] [CrossRef]
- Chen, J.; Li, P.; Chen, Z.; Wang, S.; Tang, S.; Chen, X.; Chen, Z.; Zhou, J. Elevated LINC01550 induces the apoptosis and cell cycle arrest of melanoma. Med. Oncol. 2021, 38, 32. [Google Scholar] [CrossRef]
- Wang, B.J.; Ding, H.W.; Ma, G.A. Long Noncoding RNA PVT1 Promotes Melanoma Progression via Endogenous Sponging miR-26b. Oncol. Res. 2018, 26, 675–681. [Google Scholar] [CrossRef]
- Ke, L.; Yang, D.C.; Wang, Y.; Ding, Y.; Gao, G. AnnoLnc2: The one-stop portal to systematically annotate novel lncRNAs for human and mouse. Nucleic Acids Res. 2020, 48, W230–W238. [Google Scholar] [CrossRef]
- Volders, P.J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. LNCipedia 5: Towards a reference set of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Liu, L.; Jiang, S.; Li, Q.; Feng, C.; Du, Q.; Zou, D.; Xiao, J.; Zhang, Z.; Ma, L. LncExpDB: An expression database of human long non-coding RNAs. Nucleic Acids Res. 2021, 49, D962–D968. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, X.; Shang, S.; Guo, S.; Wang, P.; Sun, D.; Gan, J.; Sun, J.; Zhang, Y.; Wang, J.; et al. LincSNP 3.0: An updated database for linking functional variants to human long non-coding RNAs, circular RNAs and their regulatory elements. Nucleic Acids Res. 2021, 49, D1244–D1250. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Xu, Q.; Liu, M.; Hu, H.; Xie, Y.; Zuo, Z.; Ren, J. lnCAR: A Comprehensive Resource for lncRNAs from Cancer Arrays. Cancer Res. 2019, 79, 2076–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. circBase: A database for circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef] [Green Version]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Guan, Z.; Wei, S.; Tan, R.; Li, P.; Yan, L. Identification of Long Non-coding and Messenger RNAs Differentially Expressed Between Primary and Metastatic Melanoma. Front. Genet. 2019, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium. Pan-cancer analysis of whole genomes. Nature 2020, 578, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Bierhoff, H. Analysis of lncRNA-Protein Interactions by RNA-Protein Pull-Down Assays and RNA Immunoprecipitation (RIP). Methods Mol. Biol. 2018, 1686, 241–250. [Google Scholar] [CrossRef]
- Hafner, M.; Landthaler, M.; Burger, L.; Khorshid, M.; Hausser, J.; Berninger, P.; Rothballer, A.; Ascano, M., Jr.; Jungkamp, A.C.; Munschauer, M.; et al. Transcriptome-wide identification of RNA-binding protein and microRNA target sites by PAR-CLIP. Cell 2010, 141, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; La Russa, M.; Qi, L.S. CRISPR/Cas9 in Genome Editing and Beyond. Annu. Rev. Biochem. 2016, 85, 227–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| lncRNA | Expression in Melanoma a | Biding Partner (s) | The Effect of lncRNA-Protein(s) Interaction | Mode of Action (in cis/in trans) | References |
|---|---|---|---|---|---|
| ANRIL and circANRIL | upregulated | CBX7 (PRC1) | ANRIL binds to PRC1 and represses the expression of the CDKN2A/B locus | in cis | [42] |
| CASC15 | upregulated | EZH2 | CASC15 binds to EZH2 and together silence the expression of PDCD4 tumor suppressor gene | in trans | [43] |
| CDR1as | downregulated | IGF2BP3 | CDR1as inhibits pro-invasive properties of IGF2BP3 which are unleashed after CDR1as loss | N/A | [44] |
| DIRC3 | downregulated | SOX10 | acts locally to block SOX10 chromatin binding at melanoma regulatory elements and activate IGFBP5 expression | in cis | [45] |
| FALEC | upregulated | EZH2 | FALEC-EZH2 complex binds to p21 promoter and inhibits p21 expression | in trans | [46] |
| GAS5 | downregulated | E2F4 | GAS5 recruits E2F4 transcription factor to EZH2 promoter, which represses EZH2 expression | in trans | [47] |
| HEIH | upregulated | EZH2 | HEIH-EZH2 repressive complex binds to the promoter of miR-200 family tumor suppressor miRNAs | in trans | [48] |
| LNMAT1 | upregulated | EZH2 | LNMAT1-EZH2 binds to CADM1 promoter and inhibits expression of this tumor suppressor gene | in trans | [49] |
| MIR31HG | upregulated | SUZ12 and EZH2 | MIR31HG binds to PRC2 represses the expression of p16 tumor suppressor gene | in cis | [50] |
| MIRAT | upregulated | IQGAP1 | MIRAT binds to and stabilizes IQGAP1, which promotes MAPK signaling | N/A | [51] |
| PVT1 | upregulated | EZH2 | PVT1-EZH2 binds to miR-200c promoter and inhibits expression of this miRNA | in trans | [52] |
| SAMMSON | upregulated | p32 | SAMMSON stabilizes the mitochondrial protein p32, required for oxidative phosphorylation | N/A | [53] |
| SLNCR1 | upregulated | Brn3a and AR AR and EGR1 | binds to MMP9 promoter and activates MMP9 transcription represses EGR1-inducible expression of p21 | in trans in trans | [54] [55] |
| SPRY4-IT1/SPRIGHTLY | upregulated | lipin 2 | SPRIGHTLY binding to lipin 2 blocks its phosphatidic acid phosphatase activity | N/A | [56] |
| THOR | upregulated | IGF2BP1 | directly binds to and stabilizes IGF2BP1; this interaction promotes melanoma growth | in trans | [57] |
| TSLNC8 | upregulated | PP1α | TSLNC8 triggers PP1α accumulation in cytoplasm which results in reactivation of MAPK signaling | N/A | [58] |
| TTN-AS1 | upregulated | TTN | High expression of TTN-AS1 and TTN induces melanoma proliferation, and suppresses cell apoptosis, | N/A | [59] |
| lncRNA | Function | Expression in Melanoma a | miRNA-mRNA Axis | References |
|---|---|---|---|---|
| BANCR | oncogene | upregulated | miR-204-5p/notch2 | [107] |
| CASC2 | tumor suppressor | downregulated | miR-18a-5p/RUNX1 miR-181a/PLXNC1 | [108] [109] |
| CCAT1 | oncogene | upregulated | miR-33a/N/A miR-296-3p/ITGA9 | [110] [111] |
| circ_0002770 | oncogene | upregulated | miR-331-3p/DUSP5, TGFBR1 | [112] |
| circ_0016418 | oncogene | upregulated | miR-625/YY1 | [113] |
| circ_0020710 | oncogene | upregulated | miR-370-3p/CXCL12 | [114] |
| circ_0025039 | oncogene | upregulated | miR-198/CDK4 | [115] |
| circ_0079593 | oncogene | upregulated | miR-573/ABHD2 | [116] |
| circ_0084043 | oncogene | upregulated | miR-153-3p/Snail miR-429/TRIB2 | [117] [118] |
| FOXD3-AS1 | oncogene | upregulated | miR-127-3p/FJX1 miR-325/MAP3K2 | [119] [120] |
| H19 | oncogene | upregulated | miR-106a-5p/E2F3 miR-18b/IGF1 | [121] [122] |
| HOTAIR | oncogene | upregulated | miR-152-3p/c-MET | [123] |
| KCNQ1OT1 | oncogene | upregulated | miR-153/MET/MYC | [124] |
| LHFPL3-AS1 | oncogene | upregulated | miR-580-3p/STAT3 miR-181a-5p/BCL2 | [125] [126] |
| LINC00520 | oncogene | upregulated | miR-125b-5p/EIF5A2 | [127] |
| LINC00459 | tumor suppressor | downregulated | miR-218/DKK3 | [128] |
| LINC00518 | oncogene | upregulated | miR-33a-3p/HIF-1α miR-204-5p/AP1S2 | [129] [130] |
| LINC00961 | tumor suppressor | downregulated | miR-367/PTEN | [131] |
| LINC01158 | oncogene | upregulated | miR-650/MGMT | [132] |
| MALAT1 | oncogene | upregulated | miR-34a/c-Myc, MET miR-183/ITGB1 miR-22/MMP14, Snail | [133] [134] [135] |
| MEG3 | tumor suppressor | downregulated | miR-499-5p/CYLD miR-21/E-cadherin | [136] [137] |
| MHENCR | oncogene | upregulated | miR-425/IGF1 miR-489/SPIN1 | [138] |
| MIR155HG | oncogene | upregulated | miR-485-3p/PSIP1 | [139] |
| MIR205HG | oncogene | upregulated | miR-299-3p/VEGFA | [140] |
| NEAT1 | oncogene | upregulated | miR-23a-3p/KLF3 miR-200b-3p/SMAD2 miR-495-3p/E2F3 | [141] [142] [143] |
| NORAD | oncogene | upregulated | miR-205/EGLN2 | [144] |
| TUG1 | oncogene | upregulated | miR-129-5p/AEG1 miR-29c-3p/RGS1 | [145] [146] |
| UCA1 | oncogene | upregulated | miR-28/HOXB3 miR-507/FOXM1 | [147] [148] |
| XIST | oncogene | upregulated | miR-23a-3p/GINS2 39-5p/ROCK1 | [149] [150] |
| ZEB1-AS1 | oncogene | upregulated | miR-1224-5p/N/A | [151] |
| lncRNA | Expression in Melanoma a | Association with Melanoma Treatment Outcomes | References |
|---|---|---|---|
| BANCR | upregulated | high expression correlated with melanoma stage (p = 0.017, n = 103), and lower OS (p < 0.01, n = 72) | [156] |
| CASC15 | upregulated | high expression associated with TNM stage (p = 0.013), distal metastasis (p = 0.018) and lymphatic metastasis (p = 0.006) (n = 461) | [43] |
| CCAT1 | upregulated | high expression linked to worse OS of melanoma patients (p = 0.038, n = 30) | [110] |
| CDR1as | downregulated | low expression correlated with shorter PFS (0.0008) and OS (p = 0.0023) of melanoma patients (n = 53) | [44] |
| circ_0025039 | upregulated | patients with high circ_0025039 expression were characterized with shorter OS (p < 0.05, n = 43) | [115] |
| circ_0002770 | upregulated | high expression correlated with shorter OS of melanoma patients (p < 0.05, n = 20) | [112] |
| circ_0020710 | upregulated | expression correlated with the advanced Breslow depth (p = 0.012) and Clark level (p = 0.034); high expression characterized with shorter OS (p = 0.036) in melanoma patients (n = 88) | [114] |
| circ_0079593 | upregulated | low expression correlated with longer survival time (p = 0.004); expression associated with melanoma clinical stage (p = 0.028) and Breslow thickness (p = 0.014) of melanoma patients (n = 47) | [116] |
| circ_0084043 | upregulated | high expression correlated with clinical stage of melanoma (p < 0.01, n = 30) and decreased OS of melanoma patients (p < 0.05, n = 33) | [117] |
| FALEC | upregulated | high expression linked to poorer patients OS (p < 0.001) and TNM stage (p = 0.012) of melanoma patients (n = 78) | [46] |
| GAS5 | downregulated | low expression correlated with the TNM staging of melanoma patients (p < 0.05, n = 94) | [74] |
| H19 | upregulated | high expression associated with advanced tumor invasion and TNM stage (p < 0.001), distal (p = 0.015) and lymph (p = 0.048) node metastases and shorter OS (p < 0.05) of melanoma patients (n = 30) | [122,164] |
| HEIH | upregulated | high expression associated with advanced clinical stages; might predict poor clinical outcomes in melanoma patients. (66 patients with melanoma and 42 patients with benign nevi, p = 0.026) | [48] |
| HOTAIR | upregulated | identified in serum of melanoma patients (n = 34); correlated with melanoma incidence | [170] |
| IGF2AS MEG3 Zeb2NAT | downregulated downregulated upregulated | combined expression levels of these three lncRNAs measured in plasma patients might serve as prognostic markers of response to vemurafenib treatment in melanoma patients | [209] |
| LHFPL3-AS1 | upregulated | high expression in melanoma patients (n = 52) associated with TNM (p = 0.009) stage and distant metastasis (p = 0.012) | [125] |
| LINC00518 | upregulated | high expression was an independent risk factor for the prognosis of melanoma patients (OS: p = 0.03, n = 458) | [130] |
| LINC00459 | downregulated | low expression correlated with decreased OS (p = 0.032) and FPS (0.009) of melanoma patients (n = 126) | [128] |
| LINC00520 | upregulated | high expression closely related to the clinical stage of melanoma (p < 0.01) and OS (p < 0.05) of melanoma patients (n = 38) | [176] |
| LINC01550 | downregulated | high expression associated with increased OS (p = 0.015, n = 541) and DFS (p = 0.042, n = 380) in melanoma patients | [227] |
| MEG3 | downregulated | low expression correlated with poor prognosis for melanoma patients (OS: p = 0.001, n = 42) | [136] |
| MHENCR | upregulated | high expression indicated poor OS of melanoma patients (p = 0.017, n = 30) | [138] |
| MIR205HG | upregulated | high expression of MIR205HG associated with lower OS of melanoma patients (p = 3.5e-6, n = 521) | [140] |
| PVT1 | upregulated | high expression negatively correlated with OS (p = 0.021) of melanoma patients (n = 35). | [228] |
| RMEL3 | upregulated | identified mutations associated with poor patient survival rates (p < 0.05, n = 38) | [211] |
| SAMMSON | upregulated | high expression negatively correlated with cytotoxic effects of vemurafenib and pimasertib in melanoma cell lines | [53] |
| SLNCR1 | upregulated | high expression was associated with shorter OS in melanoma patients (p = 0.043; n = 213) | [54] |
| SPRIGHTLY | upregulated | identified in plasma of melanoma patients (n = 70), associated with tumor stage (p < 0.001); high expression significantly reduced OS rates (p < 0.001) of melanoma patients | [79] |
| TTN-AS1 | upregulated | high expression correlated with poor OS of melanoma patients (p = 0.048, n = 165) | [59] |
| TUG1 | upregulated | Overexpressed might serve as a prognostic marker of poor prognosis for melanoma patients (OS: p = 0.026, n = 48) | [146] |
| UCA1 | upregulated | high expression positively correlated with melanoma stage (p = 0.046, n = 63) | [155] |
| ZEB1-AS1 | upregulated | high expression is correlated with decreased OS of melanoma patients (p = 0.05, n = 46). | [151] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wozniak, M.; Czyz, M. The Functional Role of Long Non-Coding RNAs in Melanoma. Cancers 2021, 13, 4848. https://doi.org/10.3390/cancers13194848
Wozniak M, Czyz M. The Functional Role of Long Non-Coding RNAs in Melanoma. Cancers. 2021; 13(19):4848. https://doi.org/10.3390/cancers13194848
Chicago/Turabian StyleWozniak, Michal, and Malgorzata Czyz. 2021. "The Functional Role of Long Non-Coding RNAs in Melanoma" Cancers 13, no. 19: 4848. https://doi.org/10.3390/cancers13194848
APA StyleWozniak, M., & Czyz, M. (2021). The Functional Role of Long Non-Coding RNAs in Melanoma. Cancers, 13(19), 4848. https://doi.org/10.3390/cancers13194848