
(In)Distinctive Role of Long Non-Coding RNAs in Common and Rare Ovarian Cancers

 ,
,  ,
,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Common Versus Rare Ovarian Cancers

{kind=link}
{kind=link}
| Cancer Type | Common or Rare | Molecular Alterations | Altered Pathways | Prognosis | Reference |
|---|---|---|---|---|---|
| Epithelial | |||||
| High-Grade Serous Ovarian Carcinoma (HGSCO) | Common | Genomic Instability, BRCA1, BRCA2, TP53, | PI3K-AKT | Almost 30% of Patients Die within 5-years of Diagnosis | [3,19,20] |
| Low-Grade Serous Ovarian Carcinoma (LGSOC) | Rare | KRAS, BRAF, ERBB2, PIK3CA, FFAR1, USP9X and EIF1AX | MAPK and AKT-mTOR | Better than The High-Grade Serous Cancer | [22] |
| Endometrioid Carcinoma (EC) | Common | CTNNB1, CDKN2A, PIK3CA, KRAS, ARID1A, PTEN, and PPP2R1A | WNT, MAPK/RAS and PI3K | Good | [19,20] |
| Ovarian Clear Cell Carcinoma (OCCC) | Rare | ARI1D1A, PI3KCA, PPP2R1A and KRAS | PI3K and mTOR | Favorable Compared with The Serous Cancer | [23,24,25] |
| Mucinous ovarian carcinoma (MOC) | Rare | High Microsatellite Instability (MSI-H), KRAS, CTNNB1, or APC | WNT | Worse than Advanced Stage Serous Cancer | [23,26,27,28] |
| Non-Epithelial | |||||
| Sex-Cord Stromal Tumors (Granulosa Cell Tumors) | Rare | FOXL2 | PI3K/AKT, TGF-β, and Notch | Good | [29,30,31,32] |
| Sex-Cord Stromal Tumors (Sertoli–Leydig Cell Tumors) | Rare | DICER1, FOXL2 | Unknown | Good | [33] |
| Germ-Cell Tumors | Rare | KIT, KRAS | Unknown | Unknown | [34] |
Availability of Cell Culture Models for Studying Common and Rare Ovarian Cancers
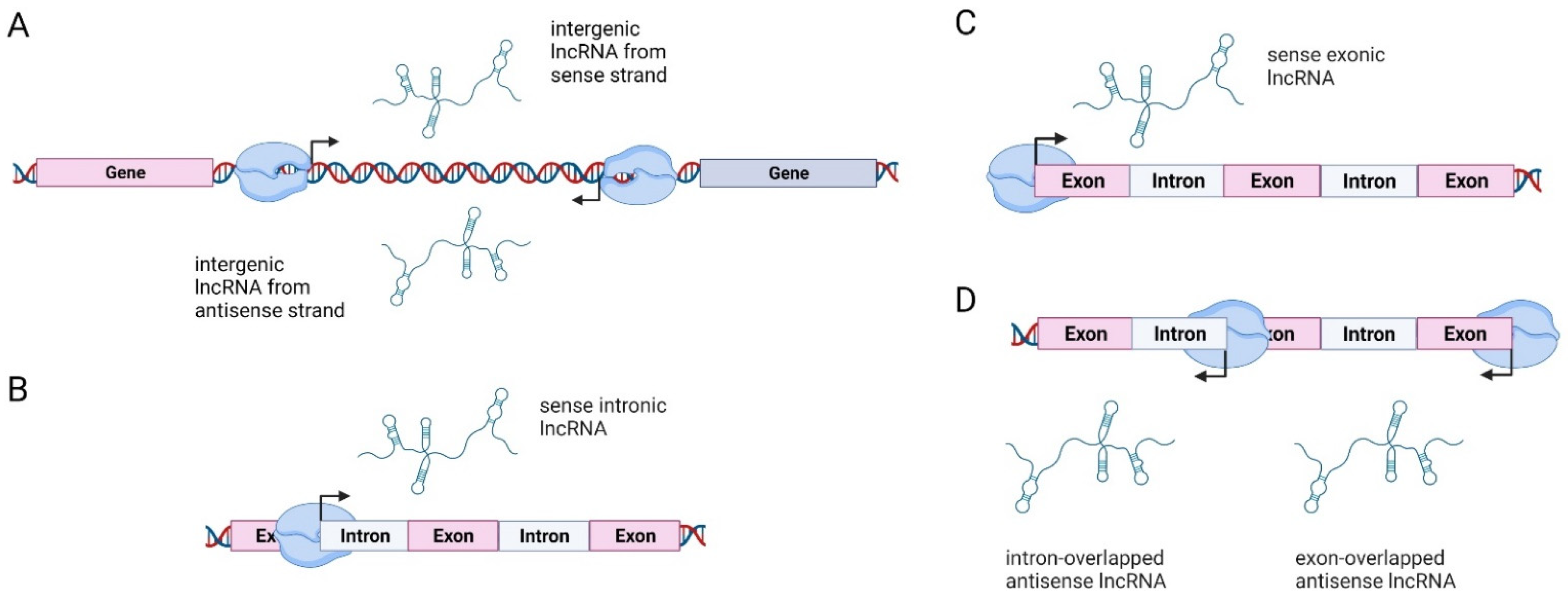
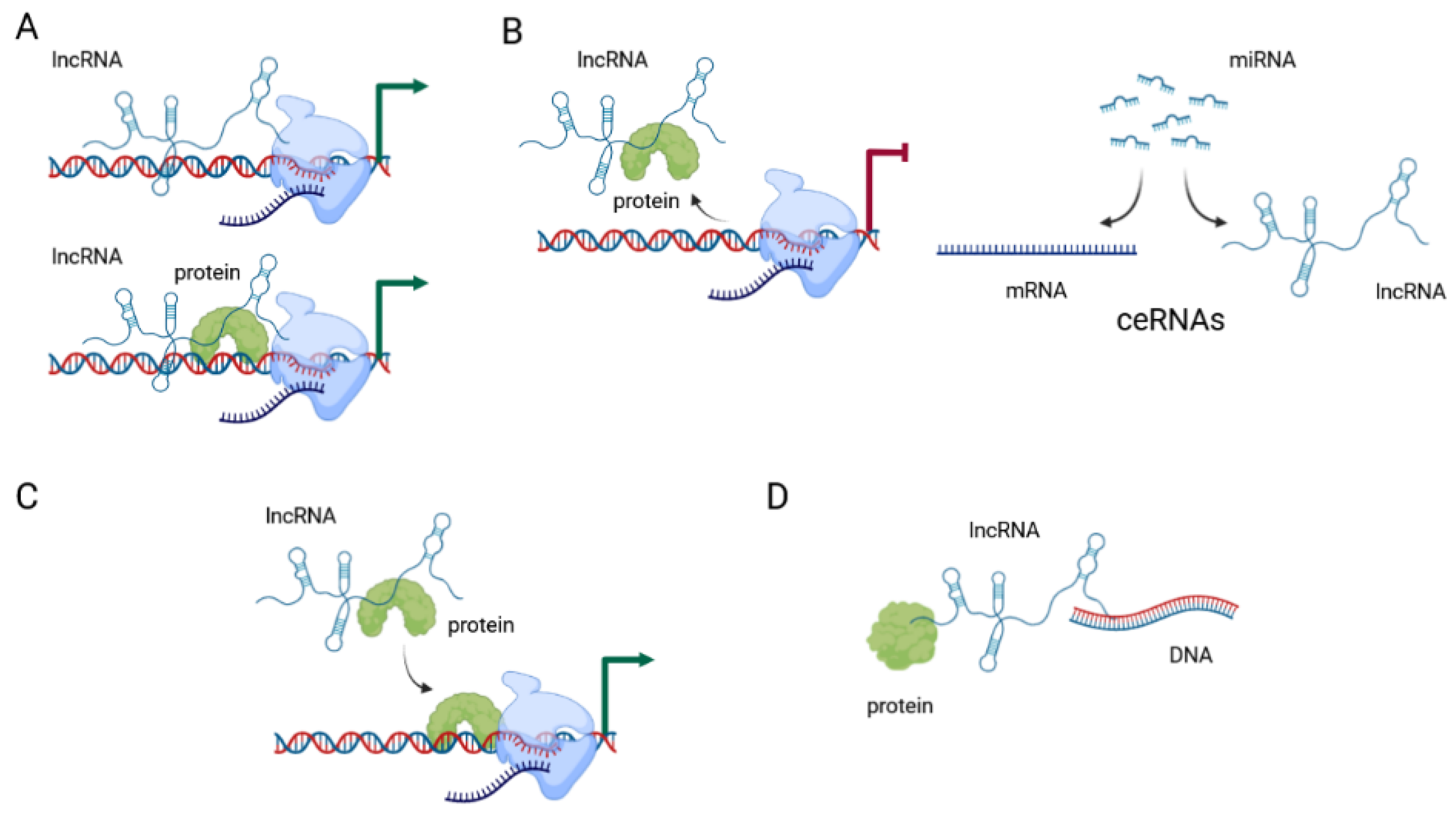
3. Long Non-Coding RNAs: Classification and Mechanisms of Action
4. Intracellular and Extracellular Compartmentalization of lncRNAs
5. The Role of lncRNAs in Various Biological Processes Related to Ovarian Carcinogenesis
6. LncRNAs Associated with Common and Rare Ovarian Cancers
6.1. H19
6.2. HOTAIR
6.3. MALAT1/NEAT2
6.4. MEG3
6.5. NEAT1
6.6. UCA1
6.7. XIST
| lncRNA | Target | OC Type | Mode of Action | Effect | Role | Reference |
|---|---|---|---|---|---|---|
| H19 | miR-370-3p | EC, HGSOC | ceRNA | Promotes TGFB1-Induced EMT | Oncogene | [144] |
| miR-324-5p | EC | ceRNA | Promotes Warburg Effect through PKM2 | Oncogene | [104] | |
| miR-19b | GCT | ceRNA | Increased Expression of CTGF, Resulting in Cell Proliferation and Reduced Rate of Apoptosis | Oncogene | [155] | |
| let-7 | EC, OCSC | ceRNA | Promotes Tumor Cell Migration and Invasion | Oncogene | [145] | |
| HOTAIR | PRC2 | / | Scaffold | The trimethylation of the H3K27 Histone and Consequent Gene Silencing | Oncogene | [156] |
| MALAT1 | miR-22 | EOC | ceRNA | Increased Cell Proliferation, Migration, Invasion, Tumor Growth, and Metastasis | Oncogene | [179] |
| miR-506 | OC | ceRNA | Upregulation of iASPP and Cell Proliferation | Oncogene | [180] | |
| miR-200c | EC, EOC | ceRNA | Increased Invasive Capacity | Oncogene | [181,238] | |
| miR-143-3p | EOC | ceRNA | Upregulation of CMPK | Oncogene | [182] | |
| miR-200a | EC | ceRNA | Promotes Autophagy and Invasion | Oncogene | [96] | |
| miR-211 | EC, HGSOC | ceRNA | Upregulation of PHF19, Leading to OC Progression | Oncogene | [183] | |
| miR-503-5p | EOC | ceRNA | Promotes Proliferation and Inhibits Apoptosis Through the JAK2-STAT3 Pathway | Oncogene | [184] | |
| miR-1271-5p | HGSOC, EC | ceRNA | Upregulation of E2F5 Expression, Mediates DPP-Resistant OC Development | Oncogene | [188] | |
| SRSF1 | EOC | Scaffold | Downregulation of RBFOX2, Leading to Alternative Splicing of KIF1B Leading to Production of the Pro-Apoptotic Isoform | Oncogene | [185] | |
| YAP | EC | ceRNA | Inhibition of Nucleus-Cytoplasm Translocation, Resulting in Enhanced Activity and Promotion of Stemness Phenotype | Oncogene | [189] | |
| AMPK | GCT | Unknown | Proliferation, Viability, Inhibition of Autophagy, Downregulation of AMPK | Oncogene | [191] | |
| multiple | GCT | Signal Molecule | Downregulation of p21, p53, p-JNK, p-ERK1/2; Upregulation of CDK2, Cyclin D1, p-P38 MAPK | Oncogene | [187] | |
| MEG3 | ATG3 | EOC | Scaffold | Protects ATG3 mRNA from Degradation, Induces Autophagy | Tumor Suppressor | [95] |
| miR-214 | EOC | ceRNA | Reduction of Resistance to Cisplatin | Tumor Suppressor | [203] | |
| PTEN | EOC | Unknown | Upregulation of PTEN, Inhibition of Cell Proliferation, Induction of Apoptosis, Cell Cycle Block | Tumor Suppressor | [200] | |
| miR-219a-5p | OC | ceRNA | Downregulation of EGRF, Inhibition of Proliferation and Induction of Apoptosis | Tumor Suppressor | [239] | |
| miR-30e-3p | OC | ceRNA | Upregulation of LAMA4, Reduced Proliferation, Migration, and Invasion of OC Cells | Tumor Suppressor | [199] | |
| miR-205-5p | OC | ceRNA | Inhibition of Cell Viability, Migration, and Invasion, Induction of Apoptosis | Tumor Suppressor | [240] | |
| NEAT1 | miR-34a-5p | OC | ceRNA | Promotes Proliferation by Upregulating BCL2 | Oncogene | [208] |
| miR-124-3p | OC | ceRNA | Promotes Cell Proliferation and Invasion, NEAT1 Expression is Stabilized by HuR Protein | Oncogene | [209] | |
| miR-382-3p | OC | ceRNA | Promotes ROCK1-Mediated Metastasis | Oncogene | [210] | |
| miR-506 | EOC | ceRNA | Promotes Cell Proliferation and Migration, NEAT1 is Stabilized by LIN28B | Oncogene | [211] | |
| miR-1321 | OC | ceRNA | Increased Expression of TJP3, Enhances EMT, Invasion, and Migration | Oncogene | [212] | |
| miR-4500 | OC | ceRNA | Increased Expression of BZW1, Enhances Cell Proliferation, Colony Formation, Migration, Invasion, and Glycolysis, Reduces Apoptosis | Oncogene | [213] | |
| miR-365 | EC, HGSOC | ceRNA | Increased Expression of FGF9, Promotes Cell Proliferation and Angiogenesis | Oncogene | [214] | |
| UCA1 | miR-485-5p | EOC | ceRNA | Increased Expression of MMP14, Possible Role in Metastasis of OC | Oncogene | [221] |
| AMOT | EOC | ceRNA | Enhances AMOT-YAP Interaction, Activation of YAP Target Genes, | Oncogene | [223] | |
| miR-143 | OC | ceRNA | Upregulation of FOSL2, Increased Cisplatin Resistance | Oncogene | [227] | |
| miR-654-5p | OC | ceRNA | Upregulation of SIK2, Resistance to Paclitaxel | Oncogene | [228] | |
| miR-129 | OC | ceRNA | Upregulation of ABCB1, Resistance to Paclitaxel | Oncogene | [229] | |
| XIST | miR-149-3p | EOC | ceRNA | Upregulation of FOXP3 Leading to OC Cell Proliferation | Oncogene | [235] |
| miR-101-3p | HGSOC | ceRNA | Upregulation of C/EBPα and KLF6 Leading to Macrophage Polarization to Affect Cell Proliferation of OC | Oncogene | [237] | |
| miR-214-3p | HGSOC | ceRNA | Suppression of Cell Proliferation, Invasion, Increased Chemosensitivity, Inhibition of Tumor Growth In Vivo | Tumor Suppressor | [236] | |
| miR-106a | OC | ceRNA | Decrease in Cell Proliferation and Activation of Apoptosis, In Vivo Tumor Growth Deceleration | Tumor Suppressor | [234] |
6.8. Other lncRNAs Involved in Rare Ovarian Cancers
| lncRNA | Target | OC Type | Mode of Action | Effect | Role | Reference |
|---|---|---|---|---|---|---|
| GAS5 | IL6 | GCT | Unknown | Upregulation of IL6, Decreased Apoptosis | Oncogene | [241] |
| HCP5 | miR-27a-3p | GCT | ceRNA | Proliferation, Inhibition of Apoptosis | Oncogene | [242] |
| NPTN-IT1 | NF90 | GCT | Scaffold | Reduced Cell Viability and Migration, Increased Apoptosis | Tumor Suppressor | [243] |
| LINC00324 | miR-214-5p | IOT | ceRNA | Proliferation, Decreased Apoptosis | Oncogene | [244] |
| NBAT1 | ERK1/2 and AKT Signaling Pathways | Serous and Other | Unknown | Inhibition of Cell Proliferation, Invasion, and Migration | Tumor Suppressor | [245] |
7. Circulating lncRNAs as Diagnostic and Prognostic Biomarkers for OCs
8. Strategies for Targeting lncRNAs as a Treatment for OCs
9. Online Resources for lncRNA Research
10. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BEP | bleomycin, etoposide, and cisplatin |
| CAF | cancer-associated fibroblasts |
| ceRNA | competing endogenous RNA |
| CSC | cancer stem cells |
| EC | endometroid carcinoma |
| EMT | epithelial to mesenchymal transition |
| EOC | epithelial ovarian cancer |
| FIGO | Fédération Internationale de Gynécologie et d’Obstétrique |
| GCT | granulosa cell tumor |
| HGSOC | high-grade serous ovarian cancer |
| HOTAIR | HOX transcript antisense RNA |
| IOT | immature ovarian teratocarcinoma |
| LGSOC | low-grade serous ovarian cancer |
| lincRNA | long intergenic non-coding RNA |
| lncRNA | long non-coding RNA |
| MALAT1 | metastasis-associated lung adenocarcinoma transcript 1 |
| MSC | mesenchymal stem cell |
| MET | mesenchymal to epithelial transition |
| miRNA/miR | microRNA |
| MOC | mucinous ovarian carcinoma |
| NAT | natural antisense transcript |
| ncRNA | non-coding RNA |
| OC | ovarian cancer |
| OCCC | ovarian clear cell carcinoma |
| OCSC | ovarian cancer stem cells |
| PCOS | polycystic ovary syndrome |
| PRC | Polycomb Repressive Complex |
| RNAi | RNA interference |
| ROC | rare ovarian cancer |
| siRNA | small interfering RNA |
| shRNA | short hairpin RNA |
| XIST | X-inactive specific transcript |
References
- Di Fiore, R.; Suleiman, S.; Ellul, B.; O’Toole, S.; Savona-Ventura, C.; Felix, A.; Napolioni, V.; Conlon, N.; Kahramanoglu, I.; Azzopardi, M.J. GYNOCARE Update: Modern Strategies to Improve Diagnosis and Treatment of Rare Gynecologic Tumors—Current Challenges and Future Directions. Cancers 2021, 13, 493. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Lisio, M.-A.; Fu, L.; Goyeneche, A.; Gao, Z.-H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Research UK. Available online: https://www.cancerresearchuk.org/home (accessed on 17 August 2021).
- Debuquoy, C.; Romeo, C.; Vanacker, H.; Ray-Coquard, I. Rare ovarian tumors: An update on diagnosis and treatment. Int. J. Gynecol. Cancer 2020, 30, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Jayde, V.; White, K.; Blomfield, P. Symptoms and diagnostic delay in ovarian cancer: A summary of the literature. Contemp. Nurse 2010, 34, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Li, Y.; Li, J.; Gao, Z.; Yang, Z.; Li, Y.; Liu, H.; Fan, T. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front. Oncol. 2020, 10, 8817. [Google Scholar] [CrossRef] [PubMed]
- Salamini-Montemurri, M.; Lamas-Maceiras, M.; Barreiro-Alonso, A.; Vizoso-Vázquez, Á.; Rodríguez-Belmonte, E.; Quindós-Varela, M.; Cerdán, M.E. The Challenges and Opportunities of LncRNAs in Ovarian Cancer Research and Clinical Use. Cancers 2020, 12, 1020. [Google Scholar] [CrossRef] [Green Version]
- Zhan, L.; Li, J.; Wei, B. Long non-coding RNAs in ovarian cancer. J. Exp. Clin. Cancer Res. 2018, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-Y.; Lu, A.-Q.; Chen, L.-J. LncRNAs in ovarian cancer. Clin. Chim. Acta 2019, 490, 17–27. [Google Scholar] [CrossRef]
- Oncul, S.; Amero, P.; Rodriguez-Aguayo, C.; Calin, G.; Sood, A.K.; Lopez-Berestein, G. Long non-coding RNAs in ovarian cancer: Expression profile and functional spectrum. RNA Biol. 2020, 17, 1523–1534. [Google Scholar] [CrossRef]
- Braga, E.A.; Fridman, M.V.; Moscovtsev, A.A.; Filippova, E.A.; Dmitriev, A.A.; Kushlinskii, N.E. LncRNAs in Ovarian Cancer Progression, Metastasis, and Main Pathways: ceRNA and Alternative Mechanisms. Int. J. Mol. Sci. 2020, 21, 8855. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, V.; Tan, Y.Q.; Wu, M.M.; Ma, L.; Zhu, T.; Lobie, P.E.; Pandey, V. Long non-coding RNAs in recurrent ovarian cancer: Theranostic perspectives. Cancer Lett. 2021, 502, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Zamaraev, A.V.; Volik, P.I.; Sukhikh, G.T.; Kopeina, G.S.; Zhivotovsky, B. Long non-coding RNAs: A view to kill ovarian cancer. Biochim. Biophys. Acta Bioenerg. 2021, 1876, 188584. [Google Scholar] [CrossRef]
- Seborova, K.; Vaclavikova, R.; Rob, L.; Soucek, P.; Vodicka, P. Non-Coding RNAs as Biomarkers of Tumor Progression and Metastatic Spread in Epithelial Ovarian Cancer. Cancers 2021, 13, 1839. [Google Scholar] [CrossRef] [PubMed]
- Keyvani, V.; Farshchian, M.; Esmaeili, S.-A.; Yari, H.; Moghbeli, M.; Nezhad, S.-R.K.; Abbaszadegan, M.R. Ovarian cancer stem cells and targeted therapy. J. Ovarian Res. 2019, 12, 120. [Google Scholar] [CrossRef] [Green Version]
- Dubeau, L.; Drapkin, R. Coming into focus: The nonovarian origins of ovarian cancer. Ann. Oncol. 2013, 24, viii28–viii35. [Google Scholar] [CrossRef] [PubMed]
- WHO. Female Genital Tumours; WHO: Geneva, Switzerland, 2020; ISBN 978-92-832-4504-9. [Google Scholar]
- Hollis, R.L.; Gourley, C. Genetic and molecular changes in ovarian cancer. Cancer Biol. Med. 2016, 13, 236–247. [Google Scholar] [CrossRef] [Green Version]
- Rojas, V.; Hirshfield, K.M.; Ganesan, S.; Rodriguez-Rodriguez, L. Molecular Characterization of Epithelial Ovarian Cancer: Implications for Diagnosis and Treatment. Int. J. Mol. Sci. 2016, 17, 2113. [Google Scholar] [CrossRef] [Green Version]
- Peres, L.C.; Cushing-Haugen, K.L.; Köbel, M.; Harris, H.R.; Berchuck, A.; Rossing, M.A.; Schildkraut, J.M.; Doherty, J. Invasive Epithelial Ovarian Cancer Survival by Histotype and Disease Stage. J. Natl. Cancer Inst. 2019, 111, 60–68. [Google Scholar] [CrossRef]
- Gadducci, A.; Cosio, S. Therapeutic Approach to Low-Grade Serous Ovarian Carcinoma: State of Art and Perspectives of Clinical Research. Cancers 2020, 12, 1336. [Google Scholar] [CrossRef] [PubMed]
- Mandilaras, V.; Karakasis, K.; Clarke, B.; Oza, A.; Lheureux, S. Rare tumors in gynaecological cancers and the lack of therapeutic options and clinical trials. Expert Opin. Orphan Drugs 2016, 5, 71–83. [Google Scholar] [CrossRef]
- Zorn, K.; Bonome, T.; Gangi, L.; Chandramouli, G.V.; Awtrey, C.S.; Gardner, G.J.; Barrett, J.C.; Boyd, J.; Birrer, M.J. Gene Expression Profiles of Serous, Endometrioid, and Clear Cell Subtypes of Ovarian and Endometrial Cancer. Clin. Cancer Res. 2005, 11, 6422–6430. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, Y.; Tokunaga, H.; Saito, S.; Shimokawa, K.; Katsuoka, F.; Bin, L.; Kojima, K.; Nagasaki, M.; Yamamoto, M.; Yaegashi, N.; et al. Identification of somatic genetic alterations in ovarian clear cell carcinoma with next generation sequencing. Genes Chromosom. Cancer 2018, 57, 51–60. [Google Scholar] [CrossRef]
- Anglesio, M.S.; Kommoss, S.; Tolcher, M.C.; Clarke, B.; Galletta, L.; Porter, H.; Damaraju, S.; Fereday, S.; Winterhoff, B.J.; Kalloger, S.; et al. Molecular characterization of mucinous ovarian tumours supports a stratified treatment approach with HER2 targeting in 19% of carcinomas. J. Pathol. 2012, 229, 111–120. [Google Scholar] [CrossRef]
- Di Fiore, R.; Suleiman, S.; Pentimalli, F.; O’Toole, S.; O’Leary, J.; Ward, M.; Conlon, N.; Sabol, M.; Ozretić, P.; Erson-Bensan, A.; et al. Could MicroRNAs Be Useful Tools to Improve the Diagnosis and Treatment of Rare Gynecological Cancers? A Brief Overview. Int. J. Mol. Sci. 2021, 22, 3822. [Google Scholar] [CrossRef]
- Babaier, A.; Ghatage, P. Mucinous Cancer of the Ovary: Overview and Current Status. Diagnostics 2020, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Manchana, T.; Ittiwut, C.; Mutirangura, A.; Kavanagh, J.J. Targeted therapies for rare gynaecological cancers. Lancet Oncol. 2010, 11, 685–693. [Google Scholar] [CrossRef]
- Kommoss, S.; Anglesio, M.S.; Mackenzie, R.; Yang, W.; Senz, J.; Ho, J.; Bell, L.; Lee, S.; Lorette, J.; Huntsman, D.G.; et al. FOXL2 molecular testing in ovarian neoplasms: Diagnostic approach and procedural guidelines. Mod. Pathol. 2013, 26, 860–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Bao, R.; Peng, S.; Zhang, C. The molecular mechanism of ovarian granulosa cell tumors. J. Ovarian Res. 2018, 11, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis-Banke, S.E.; Lerdrup, M.; Kleine-Kohlbrecher, D.; Mohammad, F.; Sidoli, S.; Jensen, O.N.; Yanase, T.; Nakamura, T.; Iwase, A.; Stylianou, A.; et al. Mutant FOXL2C134W Hijacks SMAD4 and SMAD2/3 to Drive Adult Granulosa Cell Tumors. Cancer Res. 2020, 80, 3466–3479. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Yang, W.; Mo, F.; Senz, J.; Yap, D.; Anglesio, M.S.; Gilks, B.; Morin, G.B.; Huntsman, D.G. The Oncogenic Roles of DICER1 RNase IIIb Domain Mutations in Ovarian Sertoli-Leydig Cell Tumors. Neoplasia 2015, 17, 650–660. [Google Scholar] [CrossRef] [Green Version]
- Maoz, A.; Matsuo, K.; Ciccone, M.A.; Matsuzaki, S.; Klar, M.; Roman, L.D.; Sood, A.K.; Gershenson, D.M. Molecular Pathways and Targeted Therapies for Malignant Ovarian Germ Cell Tumors and Sex Cord–Stromal Tumors: A Contemporary Review. Cancers 2020, 12, 1398. [Google Scholar] [CrossRef] [PubMed]
- Hollis, R.L.; Thomson, J.P.; Stanley, B.; Churchman, M.; Meynert, A.M.; Rye, T.; Bartos, C.; Iida, Y.; Croy, I.; Mackean, M.; et al. Molecular stratification of endometrioid ovarian carcinoma predicts clinical outcome. Nat. Commun. 2020, 11, 4995. [Google Scholar] [CrossRef] [PubMed]
- Bower, M.; Fife, K.; Holden, L.; Paradinas, F.; Rustin, G.; Newlands, E. Chemotherapy for ovarian germ cell tumours. Eur. J. Cancer 1996, 32, 593–597. [Google Scholar] [CrossRef]
- Simone, C.G.; Markham, M.J.; Dizon, D.S. Chemotherapy in ovarian germ cell tumors: A systematic review. Gynecol. Oncol. 2016, 141, 602–607. [Google Scholar] [CrossRef]
- Goulvent, T.; Ray-Coquard, I.; Borel, S.; Haddad, V.; Devouassoux-Shisheboran, M.; Vacher-Lavenu, M.-C.; Pujade-Laurraine, E.; Savina, A.; Maillet, D.; Gillet, G.; et al. DICER1andFOXL2mutations in ovarian sex cord-stromal tumours: A GINECO Group study. Histopathology 2016, 68, 279–285. [Google Scholar] [CrossRef]
- Ray-Coquard, I.; Weber, B.; Lotz, J.P.; Tournigand, C.; Provençal, J.; Mayeur, D.; Treilleux, I.; Paraiso, D.; Duvillard, P.; Pujade-Lauraine, É. Management of rare ovarian cancers: The experience of the French website «Observatory for rare malignant tumours of the ovaries» by the GINECO group: Interim analysis of the first 100 patients. Gynecol. Oncol. 2010, 119, 53–59. [Google Scholar] [CrossRef]
- Jamieson, S.; Fuller, P. Molecular Pathogenesis of Granulosa Cell Tumors of the Ovary. Endocr. Rev. 2012, 33, 109–144. [Google Scholar] [CrossRef]
- Nasioudis, D.; Mastroyannis, S.A.; Haggerty, A.F.; Ko, E.M.; Latif, N.A. Ovarian Sertoli-Leydig and granulosa cell tumor: Comparison of epidemiology and survival outcomes. Arch. Gynecol. Obstet. 2020, 302, 1–6. [Google Scholar] [CrossRef]
- Aust, S.; Eberst, L.; Tredan, O.; Rousset-Jablonski, C.; Treilleux, I.; Méeus, P.; Chopin, N.; Beurrier, F.; Charreton, A.; Véronique, D.; et al. Detailed overview on rare malignant ovarian tumors. Bull. Cancer 2020, 107, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Anglesio, M.S.; Wiegand, K.C.; Melnyk, N.; Chow, C.; Salamanca, C.; Prentice, L.M.; Senz, J.; Yang, W.; Spillman, M.A.; Cochrane, D.R.; et al. Type-Specific Cell Line Models for Type-Specific Ovarian Cancer Research. PLoS ONE 2013, 8, e72162. [Google Scholar] [CrossRef]
- Domcke, S.; Sinha, R.; Levine, D.A.; Sander, C.; Schultz, N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat. Commun. 2013, 4, 2126. [Google Scholar] [CrossRef]
- Ince, T.A.; Sousa, A.D.; Jones, M.A.; Harrell, J.C.; Agoston, E.S.; Krohn, M.; Selfors, L.; Liu, W.; Chen, K.; Yong, M.; et al. Characterization of twenty-five ovarian tumour cell lines that phenocopy primary tumours. Nat. Commun. 2015, 6, 7419. [Google Scholar] [CrossRef] [Green Version]
- Barnes, B.M.; Nelson, L.; Tighe, A.; Burghel, G.J.; Lin, I.-H.; Desai, S.; McGrail, J.C.; Morgan, R.D.; Taylor, S.S. Distinct transcriptional programs stratify ovarian cancer cell lines into the five major histological subtypes. Genome Med. 2021, 13, 1–19. [Google Scholar] [CrossRef]
- Papp, E.; Hallberg, D.; Konecny, G.E.; Bruhm, D.C.; Adleff, V.; Noë, M.; Kagiampakis, I.; Palsgrove, D.; Conklin, D.; Kinose, Y.; et al. Integrated Genomic, Epigenomic, and Expression Analyses of Ovarian Cancer Cell Lines. Cell Rep. 2018, 25, 2617–2633. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Engqvist, H.; Parris, T.Z.; Biermann, J.; Rönnerman, E.W.; Larsson, P.; Sundfeldt, K.; Kovács, A.; Karlsson, P.; Helou, K. Integrative genomics approach identifies molecular features associated with early-stage ovarian carcinoma histotypes. Sci. Rep. 2020, 10, 7946. [Google Scholar] [CrossRef] [PubMed]
- Bairoch, A. The Cellosaurus, a Cell-Line Knowledge Resource. J. Biomol. Tech. 2018, 29, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Nusinow, D.P.; Szpyt, J.; Ghandi, M.; Rose, C.M.; McDonald, E.R.; Kalocsay, M.; Jané-Valbuena, J.; Gelfand, E.; Schweppe, D.K.; Jedrychowski, M.; et al. Quantitative Proteomics of the Cancer Cell Line Encyclopedia. Cell 2020, 180, 387–402.e16. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue of Somatic Mutations in Cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15, R17–R29. [Google Scholar] [CrossRef] [Green Version]
- Aprile, M.; Katopodi, V.; Leucci, E.; Costa, V. LncRNAs in Cancer: From garbage to Junk. Cancers 2020, 12, 3220. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Merkel, A.; Gonzalez, D.; Lagarde, J.; et al. The GENCODE v7 Catalogue of Human Long Non-Coding RNAs: Analysis of Their Structure, Evolution and Expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Kapranov, P.; Drenkow, J.; Dike, S.; Brubaker, S.; Patel, S.; Long, J.; Stern, D.; Tammana, H.; Helt, G.; et al. Transcriptional Maps of 10 Human Chromosomes at 5-Nucleotide Resolution. Science 2005, 308, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Y.-H.; Pan, X.; Liu, M.; Wang, S.; Huang, T.; Cai, Y.-D. Tissue Expression Difference between mRNAs and lncRNAs. Int. J. Mol. Sci. 2018, 19, 3416. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Sun, Z. Novel lincRNA Discovery and Tissue-Specific Gene Expression across 30 Normal Human Tissues. Genes 2021, 12, 614. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 924–933. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Balas, M.; Johnson, A.M. Exploring the mechanisms behind long noncoding RNAs and cancer. Non-Coding RNA Res. 2018, 3, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Khandelwal, A.; Bacolla, A.; Vasquez, K.M.; Jain, A. Long non-coding RNA: A new paradigm for lung cancer. Mol. Carcinog. 2015, 54, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Aznaourova, M.; Schmerer, N.; Schmeck, B.; Schulte, L.N. Disease-Causing Mutations and Rearrangements in Long Non-coding RNA Gene Loci. Front. Genet. 2020, 11, 11. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nat. Cell Biol. 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Grote, P. Beyond the RNA-dependent function of LncRNA genes. eLife 2020, 9, 60583. [Google Scholar] [CrossRef] [PubMed]
- Creamer, K.M.; Lawrence, J.B. XIST RNA: A window into the broader role of RNA in nuclear chromosome architecture. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160360. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Ebert, M.S.; Sharp, P.A. Emerging Roles for Natural MicroRNA Sponges. Curr. Biol. 2010, 20, R858–R861. [Google Scholar] [CrossRef] [Green Version]
- Xing, J.; Liu, H.; Jiang, W.; Wang, L. LncRNA-Encoded Peptide: Functions and Predicting Methods. Front. Oncol. 2021, 10. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, Y.; Jia, L.; Li, W. Long Noncoding RNA H19 Promotes Osteoblast Differentiation Via TGF-β1/Smad3/HDAC Signaling Pathway by Deriving miR-675. Stem Cells 2015, 33, 3481–3492. [Google Scholar] [CrossRef]
- Bocchetti, M.; Scrima, M.; Melisi, F.; Luce, A.; Sperlongano, R.; Caraglia, M.; Zappavigna, S.; Cossu, A. LncRNAs and Immunity: Coding the Immune System with Noncoding Oligonucleotides. Int. J. Mol. Sci. 2021, 22, 1741. [Google Scholar] [CrossRef] [PubMed]
- Aillaud, M.; Schulte, L.N. Emerging Roles of Long Noncoding RNAs in the Cytoplasmic Milieu. Non-Coding RNA 2020, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Gusic, M.; Prokisch, H. ncRNAs: New Players in Mitochondrial Health and Disease? Front. Genet. 2020, 11, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcucci, V.; Stacker, S.; Achen, M. Control of Gene Expression by Exosome-Derived Non-Coding RNAs in Cancer Angiogenesis and Lymphangiogenesis. Biomolecules 2021, 11, 249. [Google Scholar] [CrossRef]
- Tassinari, M.; Gandellini, P. Noncoding RNAs in the Interplay between Tumor Cells and Cancer-Associated Fibroblasts: Signals to Catch and Targets to Hit. Cancers 2021, 13, 709. [Google Scholar] [CrossRef]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef] [Green Version]
- Carlevaro-Fita, J.; Lanzós, A.; Feuerbach, L.; Hong, C.; Mas-Ponte, D.; Pedersen, J.S.; PCAWG Drivers and Functional Interpretation Group; Johnson, R.; PCAWG Consortium Cancer. LncRNA Census reveals evidence for deep functional conservation of long noncoding RNAs in tumorigenesis. Commun. Biol. 2020, 3, 56. [Google Scholar] [CrossRef]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long noncoding RNAs in cancer metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef]
- May, J.M.; Bylicky, M.; Chopra, S.; Coleman, C.N.; Aryankalayil, M.J. Long and short non-coding RNA and radiation response: A review. Transl. Res. 2021, 233, 162–179. [Google Scholar] [CrossRef]
- McCabe, E.M.; Rasmussen, T.P. lncRNA involvement in cancer stem cell function and epithelial-mesenchymal transitions. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Zeng, S.; Liu, S.; Feng, J.; Gao, J.; Xue, F. Upregulation of lncRNA AB073614 functions as a predictor of epithelial ovarian cancer prognosis and promotes tumor growth in vitro and in vivo. Cancer Biomark. 2019, 24, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Yang, P.; Gao, Y. Long non-coding RNA EPB41L4A-AS2 suppresses progression of ovarian cancer by sequestering microRNA-103a to upregulate transcription factor RUNX1T1. Exp. Physiol. 2019, 105, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, Z.; Huang, Y.; Wang, K.; Xie, Y.; Yang, N. LncRNA GAS5 inhibits the proliferation and invasion of ovarian clear cell carcinoma via the miR -31-5p/ ARID1A axis. Kaohsiung J. Med Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Luo, C.; Meng, H.; Li, S.; Nie, S.; Jiang, Y.; Wan, Y.; Li, H.; Cheng, W. Upregulated LINC00565 Accelerates Ovarian Cancer Progression By Targeting GAS6. OncoTargets Ther. 2019, 12, 10011–10022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Wang, Y.; Xu, Y.; He, X.; Li, Y. Silencing the long noncoding RNA, TINCR, a molecular sponge of miR-335, inhibits the malignant phenotype of epithelial ovarian cancer via FGF2 suppression. Int. J. Oncol. 2019, 55, 1110–1124. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Gao, H.; Li, X.; Zhu, Y.; Peng, S.; Yu, J.; Zhan, G.; Wang, J.; Liu, N.; Guo, X. LncRNA TPT1-AS1 promotes tumorigenesis and metastasis in epithelial ovarian cancer by inducingTPT1 expression. Cancer Sci. 2019, 110, 1587–1598. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Li, H.; Li, F.; Liu, P.; Zhao, X. Long Noncoding RNA MNX1-AS1 Knockdown Inhibits Cell Proliferation and Migration in Ovarian Cancer. Cancer Biother. Radiopharm. 2017, 32, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, C.; Lu, Z.; Chen, L.; Wang, J.; Li, Y.; Ma, H. Upregulation of the long non-coding RNA SPRY4-IT1 indicates a poor prognosis and promotes tumorigenesis in ovarian cancer. Biomed. Pharmacother. 2017, 88, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, S.; Fu, C.; Wang, X.; Liu, G.; Dai, C.; Gong, Z.; Xu, H.; Fu, Z.; Xu, P.; et al. LncRNA KB-1471A8.2 Overexpression Suppresses Cell Proliferation and Migration and Antagonizes the Paclitaxel Resistance of Ovarian Cancer Cells. Cancer Biother. Radiopharm. 2019, 34, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Gao, S.; Zheng, Y.; Yao, M.; Ruan, F. LncRNA CASC15 Functions As An Unfavorable Predictor Of Ovarian Cancer Prognosis And Inhibits Tumor Progression Through Regulation Of miR-221/ARID1A Axis. OncoTargets Ther. 2019, 12, 8725–8736. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.; Zhang, X.; Zhao, Y.; Qian, H.; Wang, H.; He, C.; Liu, X.; Guo, T.; Lin, M.; Yu, H.; et al. Role of lncRNA-ATB in ovarian cancer and its mechanisms of action. Exp. Ther. Med. 2019, 19, 965–971. [Google Scholar] [CrossRef]
- Li, J.; Yang, C.; Li, Y.; Chen, A.; Li, L.; You, Z. LncRNA GAS5 suppresses ovarian cancer by inducing inflammasome formation. Biosci. Rep. 2018, 38, 20171150. [Google Scholar] [CrossRef] [Green Version]
- Xiu, Y.-L.; Sun, K.-X.; Chen, X.; Chen, S.; Zhao, Y.; Guo, Q.-G.; Zong, Z.-H. Upregulation of the lncRNA Meg3 induces autophagy to inhibit tumorigenesis and progression of epithelial ovarian carcinoma by regulating activity of ATG3. Oncotarget 2017, 8, 31714–31725. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Zhang, L.; Mei, Z.; Jiang, Y.; Yi, Y.; Liu, L.; Meng, Y.; Zhou, L.; Zeng, J.; Wu, H.; et al. Interaction of E3 Ubiquitin Ligase MARCH7 with Long Noncoding RNA MALAT1 and Autophagy-Related Protein ATG7 Promotes Autophagy and Invasion in Ovarian Cancer. Cell. Physiol. Biochem. 2018, 47, 654–666. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, X.; Tian, H.; Zhang, Z.; Tian, Y. Knockdown of long non-coding RNA HOTAIR increases cisplatin sensitivity in ovarian cancer by inhibiting cisplatin-induced autophagy. Off. J. Balk. Union Oncol. 2018, 23, 1396–1401. [Google Scholar]
- Chen, S.; Wu, D.-D.; Sang, X.-B.; Wang, L.-L.; Zong, Z.-H.; Sun, K.-X.; Liu, B.-L.; Zhao, Y. The lncRNA HULC functions as an oncogene by targeting ATG7 and ITGB1 in epithelial ovarian carcinoma. Cell Death Dis. 2017, 8, e3118. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.-H.; Du, X.; Sun, F.-D.; Wang, P.-C.; Li, M. Cisplatin suppresses tumor proliferation by inhibiting autophagy in ovarian cancer via long non-coding RNA RP11-135L22.1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Hamblin, M.R. Regulation of Glycolysis by Non-coding RNAs in Cancer: Switching on the Warburg Effect. Mol. Ther.-Oncolytics 2020, 19, 218–239. [Google Scholar] [CrossRef]
- Liu, Y.; He, X.; Chen, Y.; Cao, D. Long non-coding RNA LINC00504 regulates the Warburg effect in ovarian cancer through inhibition of miR-1244. Mol. Cell. Biochem. 2020, 464, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Lee, J.; Haemmerle, M.; Ling, H.; Previs, R.A.; Pradeep, S.; Wu, S.; Ivan, C.; Ferracin, M.; Dennison, J.B.; et al. Long Noncoding RNA Ceruloplasmin Promotes Cancer Growth by Altering Glycolysis. Cell Rep. 2015, 13, 2395–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Ji, G.; Le, X.; Wang, C.; Xu, L.; Feng, M.; Zhang, Y.; Yang, H.; Xuan, Y.; Yang, Y.; et al. Long Noncoding RNA LINC00092 Acts in Cancer-Associated Fibroblasts to Drive Glycolysis and Progression of Ovarian Cancer. Cancer Res. 2017, 77, 1369–1382. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Zhou, Y.; Chen, W.; Chen, L.; Lu, J.; He, F.; Li, X.; Zhao, L. Ginsenoside 20(S)-Rg3 Prevents PKM2-Targeting miR-324-5p from H19 Sponging to Antagonize the Warburg Effect in Ovarian Cancer Cells. Cell. Physiol. Biochem. 2018, 51, 1340–1353. [Google Scholar] [CrossRef]
- Qiu, J.-J.; Lin, X.-J.; Tang, X.-Y.; Zheng, T.-T.; Lin, Y.-Y.; Hua, K.-Q. Exosomal Metastasis-Associated Lung Adenocarcinoma Transcript 1 Promotes Angiogenesis and Predicts Poor Prognosis in Epithelial Ovarian Cancer. Int. J. Biol. Sci. 2018, 14, 1960–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Yang, F.; Qi, X.; Li, Q.; Wang, D.; Yi, T.; Yin, R.; Zhao, X.; Zhong, X.; Bian, C. LncRNA DANCR promotes tumor growth and angiogenesis in ovarian cancer through direct targeting of miR-145. Mol. Carcinog. 2019, 58, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lin, J.; Pan, J.; Qing, Q.; Li, D.; Liao, J.; Sun, C.; Zhou, H. LncRNA HNF1A-AS1 Promotes Ovarian Cancer Growth by Countering MiR-214-Mediated Suppression of the Sema 4D/Plexin B1 Pathway; Social Science Research Network: Rochester, NY, USA, 2019. [Google Scholar]
- Wu, M.; Fu, P.; Qu, L.; Liu, J.; Lin, A. Long Noncoding RNAs, New Critical Regulators in Cancer Immunity. Front. Oncol. 2020, 10, 550987. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Wang, W.; Gu, C.; Chen, C.; Zeng, B.; Yang, Y.; Ji, P.; Sun, J.; Wu, J.; Lu, W.; et al. Long non-coding RNA HOTTIP enhances IL-6 expression to potentiate immune escape of ovarian cancer cells by upregulating the expression of PD-L1 in neutrophils. J. Exp. Clin. Cancer Res. 2019, 38, 411. [Google Scholar] [CrossRef] [PubMed]
- Garg, M. Epithelial-mesenchymal transition–activating transcription factors—Multifunctional regulators in cancer. World J. Stem Cells 2013, 5, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Vafaee, F.; Colvin, E.K.; Mok, S.C.; Howell, V.M.; Samimi, G. Functional prediction of long non-coding RNAs in ovarian cancer-associated fibroblasts indicate a potential role in metastasis. Sci. Rep. 2017, 7, 10374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Liu, S.; Tao, Y. Regulating tumor suppressor genes: Post-translational modifications. Signal Transduct. Target. Ther. 2020, 5, 1–25. [Google Scholar] [CrossRef]
- Zhang, H.; Han, W. Protein Post-translational Modifications in Head and Neck Cancer. Front. Oncol. 2020, 10, 571944. [Google Scholar] [CrossRef]
- Yao, R.-W.; Wang, Y.; Chen, L.-L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Pérez-Cadahía, B.; Drobic, B.; Khan, P.; Shivashankar, C.C.; Davie, J. Current understanding and importance of histone phosphorylation in regulating chromatin biology. Curr. Opin. Drug Discov. Dev. 2010, 13, 613–622. [Google Scholar]
- Esteller, M. Epigenetics in Cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yang, Y.; Zhou, N.; Tang, K.; Lau, W.B.; Lau, B.; Wang, W.; Xu, L.; Yang, Z.; Huang, S.; et al. Epigenetics in ovarian cancer: Premise, properties, and perspectives. Mol. Cancer 2018, 17, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, A.K.; Xi, Y.; McCarthy, R.; Allton, K.; Akdemir, K.C.; Patel, L.; Aronow, B.; Lin, C.; Li, W.; Yang, L.; et al. LncPRESS1 Is a p53-Regulated LncRNA that Safeguards Pluripotency by Disrupting SIRT6-Mediated De-acetylation of Histone H3K56. Mol. Cell 2016, 64, 967–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.-T.; He, J.; Liang, Q.; Ren, L.-L.; Yan, T.-T.; Yu, T.-C.; Tang, J.; Bao, Y.-J.; Hu, Y.; Lin, Y.; et al. LncRNA GClnc1 Promotes Gastric Carcinogenesis and May Act as a Modular Scaffold of WDR5 and KAT2A Complexes to Specify the Histone Modification Pattern. Cancer Discov. 2016, 6, 784–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.-L.; Scott, A.D.; Zhou, D.C.; Wang, L.-B.; Weerasinghe, A.; Elmas, A.; Liu, R.; Wu, Y.; Wendl, M.C.; Wyczalkowski, M.A.; et al. Spatially interacting phosphorylation sites and mutations in cancer. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Mereiter, S.; Balmaña, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Abbott, K.L.; Lim, J.-M.; Wells, L.; Benigno, B.B.; McDonald, J.F.; Pierce, M. Identification of candidate biomarkers with cancer-specific glycosylation in the tissue and serum of endometrioid ovarian cancer patients by glycoproteomic analysis. Proteomics 2010, 10, 470–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, S.; Williams, C.C.; Dimapasoc, L.M.; Ro, G.S.; Ozcan, S.; Miyamoto, S.; Lebrilla, C.B.; An, H.J.; Leiserowitz, G.S. Isomer-specific chromatographic profiling yields highly sensitive and specific potential N-glycan biomarkers for epithelial ovarian cancer. J. Chromatogr. A 2013, 1279, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Saldova, R.F.; Wormald, M.; Dwek, R.A.; Rudd, P.M. Glycosylation Changes on Serum Glycoproteins in Ovarian Cancer May Contribute to Disease Pathogenesis. Dis. Markers 2008, 25, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ferguson, J.F.; Xue, C.; Ballantyne, R.L.; Silverman, I.; Gosai, S.J.; Serfecz, J.; Morley, M.P.; Gregory, B.D.; Li, M.; et al. Tissue-Specific RNA-Seq in Human Evoked Inflammation Identifies Blood and Adipose LincRNA Signatures of Cardiometabolic Diseases. Arter. Thromb. Vasc. Biol. 2014, 34, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Lanzillotti, C.; De Mattei, M.; Mazziotta, C.; Taraballi, F.; Rotondo, J.C.; Tognon, M.; Martini, F. Long Non-coding RNAs and MicroRNAs Interplay in Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 646032. [Google Scholar] [CrossRef] [PubMed]
- Nombela-Arrieta, C.; Ritz, J.; Silberstein, L.E. The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol. 2011, 12, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Bhaskar, B.; Mekala, N.; Baadhe, R.R.; Rao, P. Role of Signaling Pathways in Mesenchymal Stem Cell Differentiation. Curr. Stem Cell Res. Ther. 2014, 9, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Sikora, M.; Marycz, K.; Smieszek, A. Small and Long Non-coding RNAs as Functional Regulators of Bone Homeostasis, Acting Alone or Cooperatively. Mol. Ther.-Nucleic Acids 2020, 21, 792–803. [Google Scholar] [CrossRef]
- Wagner, E.; Luther, G.; Zhu, G.; Luo, Q.; Shi, Q.; Kim, S.H.; Gao, J.; Huang, E.; Gao, Y.; Yang, K.; et al. Defective Osteogenic Differentiation in the Development of Osteosarcoma. Sarcoma 2011, 2011, 325238. [Google Scholar] [CrossRef]
- Qiao, L.; Xu, Z.-L.; Zhao, T.-J.; Ye, L.-H.; Zhang, X.-D. Dkk-1 secreted by mesenchymal stem cells inhibits growth of breast cancer cells via depression of Wnt signalling. Cancer Lett. 2008, 269, 67–77. [Google Scholar] [CrossRef]
- Muehlberg, F.L.; Song, Y.-H.; Krohn, A.; Pinilla, S.P.; Droll, L.H.; Leng, X.; Seidensticker, M.; Ricke, J.; Altman, A.M.; Devarajan, E.; et al. Tissue-resident stem cells promote breast cancer growth and metastasis. Carcinogenesis 2009, 30, 589–597. [Google Scholar] [CrossRef] [Green Version]
- McLean, K.; Gong, Y.; Choi, Y.; Deng, N.; Yang, K.; Bai, S.; Cabrera, L.; Keller, E.; McCauley, L.; Cho, K.; et al. Human ovarian carcinoma–associated mesenchymal stem cells regulate cancer stem cells and tumorigenesis via altered BMP production. J. Clin. Investig. 2011, 121, 3206–3219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthaman, K.; Yee, F.C.; Cheyyatraivendran, S.; Biswas, A.; Choolani, M.; Bongso, A. Human umbilical cord wharton’s jelly stem cell (hWJSC) extracts inhibit cancer cell growth in vitro. J. Cell. Biochem. 2012, 113, 2027–2039. [Google Scholar] [CrossRef]
- Zhu, P.; Chen, M.; Wang, L.; Ning, Y.; Liang, J.; Zhang, H.; Xu, C.; Chen, S.; Yao, L. Systemic mesenchymal stem cells reduce growth rate of cisplatin-resistant ovarian cancer. Int. J. Clin. Exp. Pathol. 2013, 6, 2506–2514. [Google Scholar] [PubMed]
- Bu, S.; Wang, Q.; Zhang, Q.; Sun, J.; He, B.; Xiang, C.; Liu, Z.; Lai, D. Human endometrial mesenchymal stem cells exhibit intrinsic anti-tumor properties on human epithelial ovarian cancer cells. Sci. Rep. 2016, 6, 37019. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Hu, Z.; Feng, Y.; Hu, X.; Yuan, J.; Zhao, S.D.; Zhang, Y.; Yang, L.; Shan, W.; He, Q.; et al. Comprehensive Genomic Characterization of Long Non-coding RNAs across Human Cancers. Cancer Cell 2015, 28, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Akrami, R.; Jacobsen, A.; Hoell, J.; Schultz, N.; Sander, C.; Larsson, E. Comprehensive Analysis of Long Non-Coding RNAs in Ovarian Cancer Reveals Global Patterns and Targeted DNA Amplification. PLoS ONE 2013, 8, e80306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Hou, Y.; Li, K.; Li, Z.; Wang, W.; Xie, H.; Rong, Z.; Lou, G. Identification of a six-lncRNA signature associated with recurrence of ovarian cancer. Sci. Rep. 2017, 7, 752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.T.; Choi, B.H.; Niikawa, N.; Lee, T.S.; Chang, S.I. Frequent loss of imprinting of the H19 and IGF-II genes in ovarian tumors. Am. J. Med. Genet. 1998, 80, 391–395. [Google Scholar] [CrossRef]
- Zhu, Z.; Song, L.; He, J.; Sun, Y.; Liu, X.; Zou, X. Ectopic expressed long non-coding RNA H19 contributes to malignant cell behavior of ovarian cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 10082–10091. [Google Scholar]
- Li, J.; Huang, Y.; Deng, X.; Luo, M.; Wang, X.; Hu, H.; Liu, C.; Zhong, M. Long noncoding RNA H19 promotes transforming growth factor-β-induced epithelial–mesenchymal transition by acting as a competing endogenous RNA of miR-370-3p in ovarian cancer cells. OncoTargets Ther. 2018, 11, 427–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Zhou, J.; Gao, Y.; Ghazal, S.; Lu, L.; Bellone, S.; Yang, Y.; Liu, N.; Zhao, X.; Santin, A.D.; et al. Regulation of tumor cell migration and invasion by the H19/let-7 axis is antagonized by metformin-induced DNA methylation. Oncogene 2015, 34, 3076–3084. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, Y.; He, J.; Sun, H.; Jin, Z. Long non-coding RNA H19 mediates ovarian cancer cell cisplatin-resistance and migration during EMT. Int. J. Clin. Exp. Pathol. 2019, 12, 2506–2515. [Google Scholar]
- Sajadpoor, Z.; Amini-Farsani, Z.; Teimori, H.; Shamsara, M.; Sangtarash, M.H.; Ghasemi-Dehkordi, P.; Yadollahi, F. Valproic Acid Promotes Apoptosis and Cisplatin Sensitivity Through Downregulation of H19 Noncoding RNA in Ovarian A2780 Cells. Appl. Biochem. Biotechnol. 2018, 185, 1132–1144. [Google Scholar] [CrossRef]
- Zheng, Z.-G.; Xu, H.; Suo, S.-S.; Xu, X.-L.; Ni, M.-W.; Gu, L.-H.; Chen, W.; Wang, L.-Y.; Zhao, Y.; Tian, B.; et al. The Essential Role of H19 Contributing to Cisplatin Resistance by Regulating Glutathione Metabolism in High-Grade Serous Ovarian Cancer. Sci. Rep. 2016, 6, 26093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Li, T.-L.; Zhang, H.-B.; Deng, J.-L.; Zhang, R.; Sun, H.; Wan, Z.-R.; Liu, Y.-Z.; Zhu, Y.-S.; Wang, G. Polymorphisms in IGF2/H19 gene locus are associated with platinum-based chemotherapeutic response in Chinese patients with epithelial ovarian cancer. Pharmacogenomics 2019, 20, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Rachmilewitz, J.; Elkin, M.; Rosensaft, J.; Gelman-Kohan, Z.; Ariel, I.; Lustig, O.; Schneider, T.; Goshen, R.; Biran, H.; De Groot, N. H19 expression and tumorigenicity of choriocarcinoma derived cell lines. Oncogene 1995, 11, 863–870. [Google Scholar]
- Ross, J.A.; Schmidt, P.T.; Perentesis, J.P.; Davies, S.M. Genomic Imprinting of H19 and Insulin-like Growth Factor-2 in Pediatric Germ Cell Tumors. Cancer 1999, 85, 1389–1394. [Google Scholar] [CrossRef]
- Schneider, D.T.; Schuster, A.; Fritsch, M.K.; Hu, J.; Olson, T.; Lauer, S.; Göbel, U.; Perlman, E. Multipoint imprinting analysis indicates a common precursor cell for gonadal and nongonadal pediatric germ cell tumors. Cancer Res. 2001, 61, 7268–7276. [Google Scholar]
- Sievers, S.; Alemazkour, K.; Zahn, S.; Perlman, E.; Gillis, A.J.M.; Looijenga, L.; Göbel, U.; Schneider, D.T. IGF2/H19 imprinting analysis of human germ cell tumors (GCTs) using the methylation-sensitive single-nucleotide primer extension method reflects the origin of GCTs in different stages of primordial germ cell development. Genes Chromosom. Cancer 2005, 44, 256–264. [Google Scholar] [CrossRef]
- Miura, K.; Obama, M.; Yun, K.; Masuzaki, H.; Ikeda, Y.; Yoshimura, S.; Akashi, T.; Niikawa, N.; Ishimaru, T.; Jinno, Y. Meth-ylation Imprinting of H19 and SNRPN Genes in Human Benign Ovarian Teratomas. Am. J. Hum. Genet. 1999, 65, 1359–1367. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yan, X.; Liu, K.; Wu, M.; Li, Z.; Wang, Y.; Zhong, X.; Qin, L.; Huang, C.; Wei, X. lncRNA H19 acts as a ceRNA to regulate the expression of CTGF by targeting miR-19b in polycystic ovary syndrome. Braz. J. Med Biol. Res. 2020, 53, e9266. [Google Scholar] [CrossRef]
- Cantile, M.; Di Bonito, M.; De Bellis, M.T.; Botti, G. Functional Interaction among lncRNA HOTAIR and MicroRNAs in Cancer and Other Human Diseases. Cancers 2021, 13, 570. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.-J.; Wang, Y.; Ding, J.-X.; Jin, H.-Y.; Yang, G.; Hua, K.-Q. The long non-coding RNA HOTAIR promotes the proliferation of serous ovarian cancer cells through the regulation of cell cycle arrest and apoptosis. Exp. Cell Res. 2015, 333, 238–248. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Song, T.; Zou, Y.; Jiang, J.; Fang, L.; Li, P. HOTAIR is a potential target for the treatment of cisplatin-resistant ovarian cancer. Mol. Med. Rep. 2015, 12, 2211–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Guo, J.; Cai, E.; Cai, J.; Wen, Y.; Lu, S.; Li, X.; Han, Q.; Jiang, J.; Li, T.; et al. HOTAIR maintains the stemness of ovarian cancer stem cells via the miR-206/TBX3 axis. Exp. Cell Res. 2020, 395, 112218. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Hussain, I.; Ansari, K.; Kasiri, S.; Bashyal, A.; Mandal, S.S. Antisense Transcript Long Noncoding RNA (lncRNA) HOTAIR is Transcriptionally Induced by Estradiol. J. Mol. Biol. 2013, 425, 3707–3722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Liao, L.-M.; Liu, A.-W.; Wu, J.-B.; Cheng, X.-L.; Lin, J.-X.; Zheng, M. Overexpression of long noncoding RNA HOTAIR predicts a poor prognosis in patients with cervical cancer. Arch. Gynecol. Obstet. 2014, 290, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Liu, X.-F.; Wang, Y.-C.; Li, N.-D.; Liao, S.-J.; Yu, M.; Liang, C.-Z.; Tu, J.-C. Prognostic value of abnormally expressed lncRNAs in ovarian carcinoma: A systematic review and meta-analysis. Oncotarget 2017, 8, 23927–23936. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.-J.; Lin, Y.-Y.; Ye, L.-C.; Ding, J.-X.; Feng, W.-W.; Jin, H.-Y.; Zhang, Y.; Li, Q.; Hua, K.-Q. Overexpression of long non-coding RNA HOTAIR predicts poor patient prognosis and promotes tumor metastasis in epithelial ovarian cancer. Gynecol. Oncol. 2014, 134, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wen, W.; Zhao, S.; Wang, J.; Chen, J.; Wang, Y.; Zhang, Q. Prognostic role of HOTAIR in four estrogen-dependent malignant tumors: A meta-analysis. OncoTargets Ther. 2015, 8, 1471–1482. [Google Scholar] [CrossRef] [Green Version]
- Saeedi, N.; Ghorbian, S. Analysis of clinical important of LncRNA-HOTAIR gene variations and ovarian cancer susceptibility. Mol. Biol. Rep. 2020, 47, 7421–7427. [Google Scholar] [CrossRef]
- Wu, H.; Shang, X.; Shi, Y.; Yang, Z.; Zhao, J.; Yang, M.; Li, Y.; Xu, S. Genetic variants of lncRNA HOTAIR and risk of epithelial ovarian cancer among Chinese women. Oncotarget 2016, 7, 41047–41052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Lei, H.; Luo, F.; Li, Y.; Xie, L. The effect of lncRNA HOTAIR on chemoresistance of ovarian cancer through regulation of HOXA. Biol. Chem. 2018, 399, 485–497. [Google Scholar] [CrossRef]
- Teschendorff, A.E.; Lee, S.-H.; Jones, A.; Fiegl, H.; Kalwa, M.; Wagner, W.; Chindera, K.; Evans, I.; Dubeau, L.; Orjalo, A.; et al. HOTAIR and its surrogate DNA methylation signature indicate carboplatin resistance in ovarian cancer. Genome Med. 2015, 7, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özeş, A.R.; Wang, Y.; Zong, X.; Fang, F.; Pilrose, J.; Nephew, K.P. Therapeutic targeting using tumor specific peptides inhibits long non-coding RNA HOTAIR activity in ovarian and breast cancer. Sci. Rep. 2017, 7, 894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, N.; Moriam, S.; Umer, M.; Phan, H.-P.; Salomon, C.; Kline, R.; Nguyen, N.-T.; Shiddiky, M.J.A. Naked-eye and electrochemical detection of isothermally amplified HOTAIR long non-coding RNA. Analyst 2018, 143, 3021–3028. [Google Scholar] [CrossRef] [PubMed]
- Soda, N.; Umer, M.; Kashaninejad, N.; Kasetsirikul, S.; Kline, R.; Salomon, C.; Nguyen, N.-T.; Shiddiky, M.J.A. PCR-Free Detection of Long Non-Coding HOTAIR RNA in Ovarian Cancer Cell Lines and Plasma Samples. Cancers 2020, 12, 2233. [Google Scholar] [CrossRef]
- Zhu, K.; Ren, Q.; Zhao, Y. lncRNA MALAT1 overexpression promotes proliferation, migration and invasion of gastric cancer by activating the PI3K/AKT pathway. Oncol. Lett. 2019, 17, 5335–5342. [Google Scholar] [CrossRef] [Green Version]
- Pang, E.-J.; Yang, R.; Fu, X.-B.; Liu, Y.-F. Overexpression of long non-coding RNA MALAT1 is correlated with clinical progression and unfavorable prognosis in pancreatic cancer. Tumor Biol. 2015, 36, 2403–2407. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, D.; Li, Y.; Pan, X.; Zhang, X.; Kuang, B.; Zhou, M.; Li, X.; Xiong, W.; Li, G.; et al. The Long Noncoding RNA MALAT-1 is A Novel Biomarker in Various Cancers: A Meta-analysis Based on the GEO Database and Literature. J. Cancer 2016, 7, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Su, Y.; He, X.; Zhao, W.; Wu, C.; Zhang, W.; Si, X.; Dong, B.; Zhao, L.; Gao, Y.; et al. Plasma long non-coding RNA MALAT1 is associated with distant metastasis in patients with epithelial ovarian cancer. Oncol. Lett. 2016, 12, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, X.; Lv, H.; Wen, Q.; Li, J.; Tan, L.; Li, J.; Sheng, X. The Long Noncoding RNA MALAT-1 Is Highly Expressed in Ovarian Cancer and Induces Cell Growth and Migration. PLoS ONE 2016, 11, e0155250. [Google Scholar] [CrossRef]
- Zou, A.; Liu, R.; Wu, X. Long non-coding RNA MALAT1 is up-regulated in ovarian cancer tissue and promotes SK-OV-3 cell proliferation and invasion. Neoplasma 2016, 63, 865–872. [Google Scholar] [CrossRef]
- Pei, C.; Gong, X.; Zhang, Y. LncRNA MALAT-1 promotes growth and metastasis of epithelial ovarian cancer via sponging microrna. Am. J. Transl. Res. 2020, 12, 6977–6987. [Google Scholar] [PubMed]
- Lei, R.; Xue, M.; Zhang, L.; Lin, Z. Long noncoding RNA MALAT1-regulated microRNA 506 modulates ovarian cancer growth by targeting iASPP. OncoTargets Ther. 2016, 10, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Pa, M.; Naizaer, G.; Seyiti, A.; Kuerbang, G. Long Noncoding RNA MALAT1 Functions as a Sponge of MiR-200c in Ovarian Cancer. Oncol. Res. 2017. [Google Scholar] [CrossRef]
- Lin, Q.; Guan, W.; Ren, W.; Zhang, L.; Zhang, J.; Xu, G. MALAT1 affects ovarian cancer cell behavior and patient survival. Oncol. Rep. 2018, 39, 2644–2652. [Google Scholar] [CrossRef]
- Tao, F.; Tian, X.; Ruan, S.; Shen, M.; Zhang, Z. miR-211 sponges lncRNA MALAT1 to suppress tumor growth and progression through inhibiting PHF19 in ovarian carcinoma. FASEB J. 2018, 32, 6330–6343. [Google Scholar] [CrossRef]
- Sun, Q.; Li, Q.; Xie, F. LncRNA-MALAT1 regulates proliferation and apoptosis of ovarian cancer cells by targeting miR-503-5p. OncoTargets Ther. 2019, 12, 6297–6307. [Google Scholar] [CrossRef] [Green Version]
- Gordon, M.A.; Babbs, B.; Cochrane, D.R.; Bitler, B.; Richer, J.K. The long non-coding RNA MALAT1 promotes ovarian cancer progression by regulating RBFOX2-mediated alternative splicing. Mol. Carcinog. 2019, 58, 196–205. [Google Scholar] [CrossRef]
- Jin, Y.; Feng, S.-J.; Qiu, S.; Shao, N.; Zheng, J.-H. LncRNA MALAT1 promotes proliferation and metastasis in epithelial ovarian cancer via the PI3K-AKT pathway. Eur. Rev. Med Pharmacol. Sci. 2017, 21, 3176–3184. [Google Scholar] [PubMed]
- Li, Y.; Liu, Y.-D.; Chen, S.-L.; Chen, X.; Ye, D.-S.; Zhou, X.-Y.; Zhe, J.; Zhang, J. Down-regulation of long non-coding RNAMALAT1inhibits granulosa cell proliferation in endometriosis by up-regulating P21 via activation of the ERK/MAPK pathway. Mol. Hum. Reprod. 2018, 25, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Han, L.; Hu, D. LncRNA MALAT1 Regulates the Progression and Cisplatin Resistance of Ovarian Cancer Cells via Modulating miR-1271-5p/E2F5 Axis. Cancer Manag. Res. 2020, 12, 9999–10010. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Y.; Zhong, W.; Cheng, H.; Tian, Z. The Long Non-Coding RNA MALAT1 Enhances Ovarian Cancer Cell Stemness by Inhibiting YAP Translocation from Nucleus to Cytoplasm. Med. Sci. Monit. 2020, 26, e922012. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Wang, A.; Zhang, Y.; Xu, X.; Zhang, X. Knockdown of MALAT1 enhances chemosensitivity of ovarian cancer cells to cisplatin through inhibiting the Notch1 signaling pathway. Exp. Cell Res. 2018, 366, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, P.; Li, Y.; Zhao, N.; Han, H. The AMPK-mTOR axis requires increased MALAT1 expression for promoting granulosa cell proliferation in endometriosis. Exp. Ther. Med. 2020, 21, 21. [Google Scholar] [CrossRef]
- Shi, D.; Zhang, Y.; Lu, R.; Zhang, Y. The long non-coding RNA MALAT1 interacted with miR-218 modulates choriocarcinoma growth by targeting Fbxw. Biomed. Pharmacother. 2018, 97, 543–550. [Google Scholar] [CrossRef]
- Al-Rugeebah, A.; Alanazi, M.; Parine, N.R. MEG3: An Oncogenic Long Non-coding RNA in Different Cancers. Pathol. Oncol. Res. 2019, 25, 859–874. [Google Scholar] [CrossRef]
- Zhang, J.-J.; Guo, S.-H.; Jia, B.-Q. Down-regulation of long non-coding RNA MEG3 serves as an unfavorable risk factor for survival of patients with breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5143–5147. [Google Scholar]
- Qin, R.; Chen, Z.; Ding, Y.; Hao, J.; Hu, J.; Guo, F. Long non-coding RNA MEG3 inhibits the proliferation of cervical carcinoma cells through the induction of cell cycle arrest and apoptosis. Neoplasma 2013, 60, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Guo, X.; Xia, J.; Shan, T.; Gu, C.; Liang, Z.; Zhao, W.; Jin, S. MiR-148a regulates MEG3 in gastric cancer by targeting DNA methyltransferase. Med. Oncol. 2014, 31, 879. [Google Scholar] [CrossRef]
- Lu, K.-H.; Li, W.; Liu, X.-H.; Sun, M.; Zhang, M.-L.; Wu, W.-Q.; Xie, W.-P.; Hou, Y.-Y. Long non-coding RNA MEG3 inhibits NSCLC cells proliferation and induces apoptosis by affecting p53 expression. BMC Cancer 2013, 13, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, X.; Li, J.; Yang, L.; Chen, Z.; Zhao, Q.; Tan, L.; Zhou, Y.; Li, J.; Chen, Z.; Zhao, Q.; et al. Promoter hypermethylation influences the suppressive role of maternally expressed 3, a long non-coding RNA, in the development of epithelial ovarian cancer. Oncol. Rep. 2014, 32, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, Y.; Ding, L.; Yu, L.; Zhang, B.; Wei, D. LncRNA MEG3 suppressed the progression of ovarian cancer via sponging miR-30e-3p and regulating LAMA4 expression. Cancer Cell Int. 2020, 20, 181. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, W.; He, Y.; Xia, Q.; Liu, S. LncRNA MEG3 impacts proliferation, invasion, and migration of ovarian cancer cells through regulating PTEN. Inflamm. Res. 2018, 67, 927–936. [Google Scholar] [CrossRef]
- Buttarelli, M.; De Donato, M.; Raspaglio, G.; Babini, G.; Ciucci, A.; Martinelli, E.; Baccaro, P.; Pasciuto, T.; Fagotti, A.; Scambia, G.; et al. Clinical Value of lncRNA MEG3 in High-Grade Serous Ovarian Cancer. Cancers 2020, 12, 966. [Google Scholar] [CrossRef] [Green Version]
- El-Khazragy, N.; Mohammed, H.F.; Yassin, M.; Elghoneimy, K.; Bayoumy, W.; Hewety, A.; El Magdoub, H.M.; Elayat, W.; Zaki, W.; Safwat, G.; et al. Tissue-based long non-coding RNAs “PVT1, TUG1 and MEG3” signature predicts Cisplatin resistance in ovarian Cancer. Genomics 2020, 112, 4640–4646. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Xu, X.; Li, L. Curcumin suppresses cisplatin resistance development partly via modulating extracellular vesicle-mediated transfer of MEG3 and miR-214 in ovarian cancer. Cancer Chemother. Pharmacol. 2017, 79, 479–487. [Google Scholar] [CrossRef]
- Chen, J.; Peng, X.; Dai, Y. The Long Non-Coding RNA (lncRNA) AGAP2-AS1 is Upregulated in Ovarian Carcinoma and Negatively Regulates lncRNA MEG. Med. Sci. Monit. 2019, 25, 4699–4704. [Google Scholar] [CrossRef]
- Chen, Z.-J.; Zhang, Z.; Xie, B.-B.; Zhang, H.-Y. Clinical significance of up-regulated lncRNA NEAT1 in prognosis of ovarian cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3373–3377. [Google Scholar]
- Yang, C.; Li, Z.; Li, Y.; Xu, R.; Wang, Y.; Tian, Y.; Chen, W. Long non-coding RNA NEAT1 overexpression is associated with poor prognosis in cancer patients: A systematic review and meta-analysis. Oncotarget 2016, 8, 2672–2680. [Google Scholar] [CrossRef] [Green Version]
- Thankachan, S.; Bhardwaj, B.K.; Venkatesh, T.; Suresh, P.S. Long Non-coding RNA NEAT1 as an Emerging Biomarker in Breast and Gynecologic Cancers: A Systematic Overview. Reprod. Sci. 2021, 28, 2436–2447. [Google Scholar] [CrossRef]
- Ding, N.; Wu, H.; Tao, T.; Peng, E. NEAT1 regulates cell proliferation and apoptosis of ovarian cancer by miR-34a-5p/BCL. OncoTargets Ther. 2017, 10, 4905–4915. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.; Liu, J.; Zhang, Z.; Liu, L. HuR-regulated lnc RNA NEAT 1 stability in tumorigenesis and progression of ovarian cancer. Cancer Med. 2016, 5, 1588–1598. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Fu, X.; Lu, Z. Long non-coding RNA NEAT1 promoted ovarian cancer cells’ metastasis through regulation of miR-382-3p/ROCK1 axial. Cancer Sci. 2018, 109, 2188–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, W.; Yu, D.; Jun, Z.; Yachen, D.; Weiwei, W.; Midie, X.; Xingzhu, J.; Xiaohua, W. Long noncoding RNA NEAT1, regulated by LIN28B, promotes cell proliferation and migration through sponging miR-506 in high-grade serous ovarian cancer. Cell Death Dis. 2018, 9, 861. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zhang, L.; Yang, H.; Luo, K.; Qing, C. Long non-coding RNA NEAT1 promotes ovarian cancer cell invasion and migration by interacting with miR-1321 and regulating tight junction protein 3 expression. Mol. Med. Rep. 2020, 22, 3429–3439. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Sun, X.; Huang, Y.; Si, Q.; Li, M. Long non-coding RNA NEAT1 modifies cell proliferation, colony formation, apoptosis, migration and invasion via the miR-4500/BZW1 axis in ovarian cancer. Mol. Med. Rep. 2020, 22, 3347–3357. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Yi, K.; Yang, L. LncRNA NEAT1 promotes proliferation of ovarian cancer cells and angiogenesis of co-incubated human umbilical vein endothelial cells by regulating FGF9 through sponging miR-365: An Experimental Study. Medicine 2021, 100, e23423. [Google Scholar] [CrossRef]
- Zhu, M.; Yang, L.; Wang, X. NEAT1 Knockdown Suppresses the Cisplatin Resistance in Ovarian Cancer by Regulating miR-770-5p/PARP1 Axis. Cancer Manag. Res. 2020, 12, 7277–7289. [Google Scholar] [CrossRef]
- An, J.; Lv, W.; Zhang, Y. LncRNA NEAT1 contributes to paclitaxel resistance of ovarian cancer cells by regulating ZEB1 expression via miR-194. OncoTargets Ther. 2017, 10, 5377–5390. [Google Scholar] [CrossRef] [Green Version]
- Nitusca, D.; Marcu, A.; Dema, A.; Balacescu, L.; Balacescu, O.; Bardan, R.; Cumpanas, A.; Sirbu, I.; Petrut, B.; Seclaman, E.; et al. Long Noncoding RNA NEAT1 as a Potential Candidate Biomarker for Prostate Cancer. Life 2021, 11, 320. [Google Scholar] [CrossRef]
- Swellam, M.; El Magdoub, H.M.; Shawki, M.; Adel, M.; Hefny, M.M.; El-Shazly, S.S. Clinical impact of LncRNA XIST and LncRNA NEAT1 for diagnosis of high-risk group breast cancer patients. Curr. Probl. Cancer 2021, 100709. [Google Scholar] [CrossRef]
- Pils, D.; Tong, D.; Hager, G.; Obermayr, E.; Aust, S.; Heinze, G.; Kohl, M.; Schuster, E.; Wolf, A.; Sehouli, J.; et al. A combined blood based gene expression and plasma protein abundance signature for diagnosis of epithelial ovarian cancer—A study of the OVCAD consortium. BMC Cancer 2013, 13, 178. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.-H.; Hou, L.-K.; Pan, X.; Wu, C.-Y.; Huang, H.; Li, B.; Nie, W. Long non-coding RNA UCA1 is a predictive biomarker of cancer. Oncotarget 2016, 7, 44442–44447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Jiang, Y.; Wan, Y.; Zhang, L.; Qiu, J.; Zhou, S.; Cheng, W. UCA1 functions as a competing endogenous RNA to suppress epithelial ovarian cancer metastasis. Tumor Biol. 2016, 37, 10633–10641. [Google Scholar] [CrossRef] [PubMed]
- Moss, N.M.; Barbolina, M.V.; Liu, Y.; Sun, L.; Munshi, H.G.; Stack, M.S. Ovarian Cancer Cell Detachment and Multicellular Aggregate Formation Are Regulated by Membrane Type 1 Matrix Metalloproteinase: A Potential Role in I.p. Metastatic Dissemination. Cancer Res. 2009, 69, 7121–7129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Spindler, T.J.; Fonseca, M.A.D.S.; Corona, R.I.; Seo, J.-H.; Dezem, F.S.; Li, L.; Lee, J.M.; Long, H.W.; Sellers, T.A.; et al. Super-Enhancer-Associated LncRNA UCA1 Interacts Directly with AMOT to Activate YAP Target Genes in Epithelial Ovarian Cancer. iScience 2019, 17, 242–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.R.; Zhao, M.Y.; Sun, L.; Yang, B.C.; Hei, K.W.; Du, X.; Li, Y.M. Expression of IncRNA UCA1 in ovarian cancer and its clinical significance. Eur. J. Gynaecol. Oncol. 2017, 38, 191–195. [Google Scholar]
- Zhang, L.; Cao, X.; Zhang, L.; Zhang, X.; Sheng, H.; Tao, K. UCA1 overexpression predicts clinical outcome of patients with ovarian cancer receiving adjuvant chemotherapy. Cancer Chemother. Pharmacol. 2016, 77, 629–634. [Google Scholar] [CrossRef]
- Wang, F.; Zhou, J.; Xie, X.; Hu, J.; Chen, L.; Hu, Q.; Guo, H.; Yu, C. Involvement of SRPK1 in cisplatin resistance related to long non-coding RNA UCA1 in human ovarian cancer cells. Neoplasma 2015, 62, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Niu, H.; Qin, Q.; Yang, S.; Wang, Q.; Yu, C.; Wei, Z.; Jin, Z.; Wang, X.; Yang, A.; et al. lncRNA UCA1 Mediates Resistance to Cisplatin by Regulating the miR-143/FOSL2-Signaling Pathway in Ovarian Cancer. Mol. Ther.—Nucleic Acids 2019, 17, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-Y.; Wang, X.-L.; Dang, Y.; Zhu, X.-Z.; Zhang, Y.-H.; Cai, B.-X.; Zheng, L. Long non-coding RNA UCA1 promotes the progression of paclitaxel resistance in ovarian cancer by regulating the miR-654-5p/SIK2 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 591–603. [Google Scholar] [CrossRef]
- Wang, J.; Ye, C.; Liu, J.; Hu, Y. UCA1 confers paclitaxel resistance to ovarian cancer through miR-129/ABCB1 axis. Biochem. Biophys. Res. Commun. 2018, 501, 1034–1040. [Google Scholar] [CrossRef]
- Pintacuda, G.; Young, A.N.; Cerase, A. Function by Structure: Spotlights on Xist Long Non-coding RNA. Front. Mol. Biosci. 2017, 4, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Kong, F.; Xing, L.; Jin, Z.; Li, Z. Prognostic and clinicopathological value of long noncoding RNA XIST in cancer. Clin. Chim. Acta 2018, 479, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Mei, X.; Tang, D. Long non-coding RNA XIST is down-regulated and correlated to better prognosis in ovarian cancer. Math. Biosci. Eng. 2020, 17, 2070–2081. [Google Scholar] [CrossRef] [PubMed]
- Zuo, K.; Zhao, Y.; Zheng, Y.; Chen, D.; Liu, X.; Du, S.; Liu, Q. Long non-coding RNA XIST promotes malignant behavior of epithelial ovarian cancer. OncoTargets Ther. 2019, 12, 7261–7267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Yuan, D.; Zhang, W.; Zhu, D.; Xiao, A.; Mao, G.; Jiang, W.; Lin, M.; Wang, J. Upregulation of long noncoding RNA XIST has anticancer effects on ovarian cancer through sponging miR-106a. Hum. Cell 2021, 34, 579–587. [Google Scholar] [CrossRef]
- Jiang, R.; Zhang, H.; Zhou, J.; Wang, J.; Xu, Y.; Zhang, H.; Gu, Y.; Fu, F.; Shen, Y.; Zhang, G.; et al. Inhibition of long non-coding RNA XIST upregulates microRNA-149-3p to repress ovarian cancer cell progression. Cell Death Dis. 2021, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Qi, S.; Xie, C.; Li, C.; Wang, P.; Liu, D. Upregulation of long non-coding RNA XIST has anticancer effects on epithelial ovarian cancer cells through inverse downregulation of hsa-miR-214-3p. J. Gynecol. Oncol. 2018, 29, e99. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, Z.; Ma, R.; Zhang, Y.; Zhao, L.; Yan, Y.; Lv, X.; Zhang, L.; Su, P.; Bi, J.; et al. lncRNA-Xist/miR-101-3p/KLF6/C/EBPα axis promotes TAM polarization to regulate cancer cell proliferation and migration. Mol. Ther.-Nucleic Acids 2021, 23, 536–551. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, C.; Chen, R.; Xiong, H.; Qiu, F.; Liu, S.; Zhang, M.; Wang, F.; Wang, Y.; Zhou, X.; et al. Disrupting MALAT1/miR-200c sponge decreases invasion and migration in endometrioid endometrial carcinoma. Cancer Lett. 2016, 383, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, M.; Zhao, S. lncRNA MEG3 modified epithelial-mesenchymal transition of ovarian cancer cells by sponging miR-219a-5p and regulating EGFR. J. Cell. Biochem. 2019, 120, 17709–17722. [Google Scholar] [CrossRef] [PubMed]
- Tao, P.; Yang, B.; Zhang, H.; Sun, L.; Wang, Y.; Zheng, W. The overexpression of lncRNA MEG3 inhibits cell viability and invasion and promotes apoptosis in ovarian cancer by sponging miR-205-5p. Int. J. Clin. Exp. Pathol. 2020, 13, 869–879. [Google Scholar]
- Wang, C.; Yue, S.; Jiang, Y.; Mao, Y.; Zhao, Z.; Liu, X.; Zhang, X.; Pei, D.; Li, Y. LncRNA GAS5 is upregulated in polycystic ovary syndrome and regulates cell apoptosis and the expression of IL-1. J. Ovarian Res. 2020, 13, 145. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, X.; An, Y.; Liu, B.; Lu, M. LncRNA HCP5 promotes cell proliferation and inhibits apoptosis via miR-27a-3p/IGF-1 axis in human granulosa-like tumor cell line KGN. Mol. Cell. Endocrinol. 2020, 503, 110697. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Zhang, W.; Meng, J.; Ma, L.; Li, A. LncRNA-LET inhibits cell viability, migration and EMT while induces apoptosis by up-regulation of TIMP2 in human granulosa-like tumor cell line KGN. Biomed. Pharmacother. 2018, 100, 250–256. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, M.; Xie, L.; Wu, S.; Zhong, Y. LINC00324 facilitates cell proliferation through competing for miR-214-5p in immature ovarian teratocarcinoma. Int. J. Mol. Med. 2020, 47, 397–407. [Google Scholar] [CrossRef]
- Yan, C.; Jiang, Y.; Wan, Y.; Zhang, L.; Liu, J.; Zhou, S.; Cheng, W. Long noncoding RNA NBAT-1 suppresses tumorigenesis and predicts favorable prognosis in ovarian cancer. OncoTargets Ther. 2017, 10, 1993–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, J.; Leão, R.; Jerónimo, C.; Henrique, R. Liquid Biopsies in the Clinical Management of Germ Cell Tumor Patients: State-of-the-Art and Future Directions. Int. J. Mol. Sci. 2021, 22, 2654. [Google Scholar] [CrossRef]
- Wang, X.; Kong, D.; Wang, C.; Ding, X.; Zhang, L.; Zhao, M.; Chen, J.; Xu, X.; Hu, X.; Yang, J.; et al. Circulating microRNAs as novel potential diagnostic biomarkers for ovarian cancer: A systematic review and updated meta-analysis. J. Ovarian Res. 2019, 12, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Liu, S.; Liu, Y.; Lin, X.; Zheng, T.; Liu, X.; Qiu, J.; Hua, K. Circulating serum exosomal aHIF is a novel prognostic predictor for epithelial ovarian cancer. OncoTargets Ther. 2019, 12, 7699–7711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Zhang, H. Serum lncRNA LOXL1-AS1 is a diagnostic and prognostic marker for epithelial ovarian cancer. J. Gene Med. 2020, 22, e3233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, C.; Huang, Z.; Li, Z.; Zhang, Q.; He, Y. In Silico screening of circulating tumor DNA, circulating microRNAs, and long non-coding RNAs as diagnostic molecular biomarkers in ovarian cancer: A comprehensive meta-analysis. PLoS ONE 2021, 16, e0250717. [Google Scholar] [CrossRef]
- Markou, A.; Smilkou, S.; Tsaroucha, E.; Lianidou, E. The Effect of Genomic DNA Contamination on the Detection of Circulating Long Non-Coding RNAs: The Paradigm of MALAT1. Diagnostics 2021, 11, 1160. [Google Scholar] [CrossRef]
- Cai, T.; Zhang, Q.; Wu, B.; Wang, J.; Li, N.; Zhang, T.; Wang, Z.; Luo, J.; Guo, X.; Ding, X.; et al. LncRNA-encoded microproteins: A new form of cargo in cell culture-derived and circulating extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12123. [Google Scholar] [CrossRef]
- Kim, S.S.; Baek, G.O.; Son, J.; Ahn, H.R.; Yoon, M.K.; Cho, H.J.; Yoon, J.H.; Nam, S.W.; Cheong, J.Y.; Eun, J.W. Early detection of hepatocellular carcinoma via liquid biopsy: Panel of small extracellular vesicle-derived long noncoding RNAs identified as markers. Mol. Oncol. 2021, 15, 2715–2731. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, S.; Xi, X.; Zhang, M.; Liu, X.; Tang, W.; Cai, P.; Xing, S.; Bao, P.; Jin, Y.; et al. Integrative analysis of long extracellular RNAs reveals a detection panel of noncoding RNAs for liver cancer. Theranostics 2021, 11, 181–193. [Google Scholar] [CrossRef]
- Yu, X.; Wang, R.; Han, C.; Wang, Z.; Jin, X. A Panel of Urinary Long Non-coding RNAs Differentiate Bladder Cancer from Urocystitis. J. Cancer 2020, 11, 781–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.; Du, L.; Wang, L.; Jiang, X.; Zhang, S.; Li, J.; Yan, K.; Duan, W.; Zhao, Y.; Wang, L.; et al. Expression signatures of exosomal long non-coding RNAs in urine serve as novel non-invasive biomarkers for diagnosis and recurrence prediction of bladder cancer. Mol. Cancer 2018, 17, 142. [Google Scholar] [CrossRef]
- Barwal, T.S.; Sharma, U.; Vasquez, K.M.; Prakash, H.; Jain, A. A panel of circulating long non-coding RNAs as liquid biopsy biomarkers for breast and cervical cancers. Biochimie 2020, 176, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Dong, C.; Liu, G.; Li, Y.; Liu, L. Panel of seven long noncoding RNA as a candidate prognostic biomarker for ovarian cancer. OncoTargets Ther. 2017, 10, 2805–2813. [Google Scholar] [CrossRef] [Green Version]
- Hyter, S.; Hirst, J.; Pathak, H.; Pessetto, Z.Y.; Koestler, D.C.; Raghavan, R.; Pei, D.; Godwin, A.K. Developing a genetic signature to predict drug response in ovarian cancer. Oncotarget 2018, 9, 14828–14848. [Google Scholar] [CrossRef]
- Song, J.; Zhang, W.; Wang, S.; Liu, K.; Song, F.; Ran, L. A panel of 7 prognosis-related long non-coding RNAs to improve platinum-based chemoresistance prediction in ovarian cancer. Int. J. Oncol. 2018, 53, 866–876. [Google Scholar] [CrossRef]
- Wang, L.; Hu, Y.; Xiang, X.; Qu, K.; Teng, Y. Identification of long non-coding RNA signature for paclitaxel-resistant patients with advanced ovarian cancer. Oncotarget 2017, 8, 64191–64202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arun, G.; Diermeier, S.D.; Spector, D.L. Therapeutic Targeting of Long Non-Coding RNAs in Cancer. Trends Mol. Med. 2018, 24, 257–277. [Google Scholar] [CrossRef]
- Jiang, M.-C.; Ni, J.-J.; Cui, W.-Y.; Wang, B.-Y.; Zhuo, W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am J Cancer Res 2019, 9, 1354–1366. [Google Scholar]
- Sandy, P.; Ventura, A.; Jacks, T. Mammalian RNAi: A practical guide. Biotechniques 2005, 39, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taxman, D.J.; Moore, C.B.; Guthrie, E.H.; Huang, M.T.-H. Short Hairpin RNA (shRNA): Design, Delivery, and Assessment of Gene Knockdown. Methods Mol. Biol. 2010, 629, 139–156. [Google Scholar] [CrossRef] [Green Version]
- Richard, J.L.C.; Eichhorn, P.J.A. Platforms for Investigating LncRNA Functions. SLAS Technol. 2018, 23, 493–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dizaji, B.F. Strategies to target long non-coding RNAs in cancer treatment: Progress and challenges. Egypt. J. Med. Hum. Genet. 2020, 21, 41. [Google Scholar] [CrossRef]
- Walder, R.Y.; Walder, J.A. Role of RNase H in hybrid-arrested translation by antisense oligonucleotides. Proc. Natl. Acad. Sci. USA 1988, 85, 5011–5015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arun, G.; Diermeier, S.; Akerman, M.; Chang, K.-C.; Wilkinson, J.E.; Hearn, S.; Kim, Y.; MacLeod, A.R.; Krainer, A.; Norton, L.; et al. Differentiation of mammary tumors and reduction in metastasis uponMalat1lncRNA loss. Genes Dev. 2016, 30, 34–51. [Google Scholar] [CrossRef] [Green Version]
- Gutschner, T.; Hämmerle, M.; Eißmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Groß, M.; et al. The Noncoding RNA MALAT1 Is a Critical Regulator of the Metastasis Phenotype of Lung Cancer Cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurreck, J.; Wyszko, E.; Gillen, C.; Erdmann, V.A. Design of antisense oligonucleotides stabilized by locked nucleic acids. Nucleic Acids Res. 2002, 30, 1911–1918. [Google Scholar] [CrossRef] [Green Version]
- Sarma, K.; Levasseur, P.; Aristarkhov, A.; Lee, J.T. Locked nucleic acids (LNAs) reveal sequence requirements and kinetics of Xist RNA localization to the X chromosome. Proc. Natl. Acad. Sci. USA 2010, 107, 22196–22201. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhang, X.; Chen, S.; Zhang, S. Natural antisense transcripts in the biological hallmarks of cancer: Powerful regulators hidden in the dark. J. Exp. Clin. Cancer Res. 2020, 39, 187. [Google Scholar] [CrossRef]
- Modarresi, F.; Faghihi, M.A.; Lopez-Toledano, M.; Fatemi, R.P.; Magistri, M.; Brothers, S.; Van Der Brug, M.P.; Wahlestedt, C. Inhibition of natural antisense transcripts in vivo results in gene-specific transcriptional upregulation. Nat. Biotechnol. 2012, 30, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.; Chong, K.Y.; Hartwich, T.M.P.; Bi, F.; Witham, A.K.; Patrick, D.; Morrisson, M.J.; Cady, S.L.; Cerchia, A.P.; Kelk, D.; et al. Ovarian BDNF promotes survival, migration, and attachment of tumor precursors originated from p53 mutant fallopian tube epithelial cells. Oncogenesis 2020, 9, 55. [Google Scholar] [CrossRef]
- Hagedorn, P.H.; Persson, R.; Funder, E.D.; Albæk, N.; Diemer, S.L.; Hansen, D.J.; Møller, M.R.; Papargyri, N.; Christiansen, H.; Hansen, B.R.; et al. Locked nucleic acid: Modality, diversity, and drug discovery. Drug Discov. Today 2018, 23, 101–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, C.J.; Maier, V.K.; Davey, R.; Brennan, J.; Li, G.; Brothers, J.; Schwartz, B.; Gordo, S.; Kasper, A.; Okamoto, T.R.; et al. Gene activation of SMN by selective disruption of lncRNA-mediated recruitment of PRC2 for the treatment of spinal muscular atrophy. Proc. Natl. Acad. Sci. USA 2017, 114, E1509–E1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousard, A.; Raposo, A.C.; Zylicz, J.; Picard, C.; Pires, V.B.; Qi, Y.; Gil, C.; Syx, L.; Chang, H.Y.; Heard, E.; et al. The role of Xist -mediated Polycomb recruitment in the initiation of X-chromosome inactivation. EMBO Rep. 2019, 20, e48019. [Google Scholar] [CrossRef]
- Antorod, S.W.S.; Joyce, G.F. Mechanism and Utility of an RNA-Cleaving DNA Enzyme. Biochemistry 1998, 37, 13330–13342. [Google Scholar] [CrossRef]
- Franzen, S. Expanding the catalytic repertoire of ribozymes and deoxyribozymes beyond RNA substrates. Curr. Opin. Mol. Ther. 2010, 12, 223–232. [Google Scholar]
- Sednev, M.V.; Mykhailiuk, V.; Choudhury, P.; Halang, J.; Sloan, K.E.; Bohnsack, M.T.; Höbartner, C. N 6 -Methyladenosine-Sensitive RNA-Cleaving Deoxyribozymes. Angew. Chem. Int. Ed. 2018, 57, 15117–15121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liu, C.; Zhang, S.; Yan, H.; Zhang, L.; Jiang, A.; Liu, Y.; Feng, Y.; Li, D.; Guo, Y.; et al. N6-methyladenosine modification of MALAT1 promotes metastasis via reshaping nuclear speckles. Dev. Cell 2021, 56, 702–715.e8. [Google Scholar] [CrossRef]
- Patil, D.P.; Chen, C.K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m6A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive Analysis of mRNA Methylation Reveals Enrichment in 3′ UTRs and near Stop Codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Yao, S.; Zhou, Y.; Liu, Y.; Huang, P.; Zhou, A.; Liu, J.; Che, L.; Li, J. Long noncoding RNA GAS5 inhibits progression of colorectal cancer by interacting with and triggering YAP phosphorylation and degradation and is negatively regulated by the m6A reader YTHDF1. Mol. Cancer 2019, 18, 143. [Google Scholar] [CrossRef]
- Hu, X.; Peng, W.-X.; Zhou, H.; Jiang, J.; Zhou, X.; Huang, D.; Mo, Y.-Y.; Yang, L. IGF2BP2 regulates DANCR by serving as an N6-methyladenosine reader. Cell Death Differ. 2019, 27, 1782–1794. [Google Scholar] [CrossRef] [Green Version]
- Garikipati, V.; Uchida, S. Elucidating the Functions of Non-Coding RNAs from the Perspective of RNA Modifications. Non-Coding RNA 2021, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ji, J.; Shen, J.; Lu, X. When Long Noncoding RNAs Meet Genome Editing in Pluripotent Stem Cells. Stem Cells Int. 2017, 2017, 3250624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, S.; Li, X. Application of CRISPR-Cas9 for Long Noncoding RNA Genes in Cancer Research. Hum. Gene Ther. 2019, 30, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Morelli, E.; Gulla’, A.; Amodio, N.; Taiana, E.; Neri, A.; Fulciniti, M.; Munshi, N.C. CRISPR Interference (CRISPRi) and CRISPR Activation (CRISPRa) to Explore the Oncogenic lncRNA Network. Methods Mol. Biol. 2021, 2348, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Li, W.; Liu, J.; Chen, C.-H.; Liao, Q.; Xu, P.; Xu, H.; Xiao, T.; Cao, Z.; Peng, J.; et al. Genome-scale deletion screening of human long non-coding RNAs using a paired-guide RNA CRISPR–Cas9 library. Nat. Biotechnol. 2016, 34, 1279–1286. [Google Scholar] [CrossRef]
- Liu, S.J.; Horlbeck, M.A.; Cho, S.W.; Birk, H.S.; Malatesta, M.; He, D.; Attenello, F.J.; Villalta, J.E.; Cho, M.Y.; Chen, Y.; et al. CRISPRi-based genome-scale identification of functional long noncoding RNA loci in human cells. Science 2017, 355, eaah7111. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Cai, G.; Liu, X.; Zhang, Y.; Wang, Y.; Zhou, L.; Sui, H.; Li, Q. MALAT1 regulates the transcriptional and translational levels of proto-oncogene RUNX2 in colorectal cancer metastasis. Cell Death Dis. 2019, 10, 378. [Google Scholar] [CrossRef]
- Colognori, D.; Sunwoo, H.; Kriz, A.J.; Wang, C.-Y.; Lee, J.T. Xist Deletional Analysis Reveals an Interdependency between Xist RNA and Polycomb Complexes for Spreading along the Inactive X. Mol. Cell 2019, 74, 101–117.e10. [Google Scholar] [CrossRef] [Green Version]
- Attenello, F.J.; Tsung, K.; Bishara, I.; Loh, Y.E.; Chen, T.C. In vivo CRISPR screening for novel noncoding RNA functional targets in glioblastoma models. J. Neurosci. Res. 2021, 99, 2029–2045. [Google Scholar] [CrossRef]
- Chen, B.; Deng, S.; Ge, T.; Ye, M.; Yu, J.; Lin, S.; Ma, W.; Songyang, Z. Live cell imaging and proteomic profiling of endogenous NEAT1 lncRNA by CRISPR/Cas9-mediated knock-in. Protein Cell 2020, 11, 641–660. [Google Scholar] [CrossRef]
- Zhen, S.; Hua, L.; Liu, Y.-H.; Sun, X.-M.; Jiang, M.-M.; Chen, W.; Zhao, L.; Li, X. Inhibition of long non-coding RNA UCA1 by CRISPR/Cas9 attenuated malignant phenotypes of bladder cancer. Oncotarget 2017, 8, 9634–9646. [Google Scholar] [CrossRef] [Green Version]
- Qiu, H.; Wang, X.; Guo, R.; Liu, Q.; Wang, Y.; Yuan, Z.; Li, J.; Shi, H. HOTAIR rs920778 polymorphism is associated with ovarian cancer susceptibility and poor prognosis in a Chinese population. Futur. Oncol. 2017, 13, 347–355. [Google Scholar] [CrossRef]
- Wang, V.; Wu, W. MicroRNA-based therapeutics for cancer. BioDrugs 2009, 23, 15–23. [Google Scholar] [CrossRef]
- Lin, H.; Shen, L.; Lin, Q.; Dong, C.; Maswela, B.; Illahi, G.S.; Wu, X. SNHG5 enhances Paclitaxel sensitivity of ovarian cancer cells through sponging miR-23a. Biomed. Pharmacother. 2020, 123, 109711. [Google Scholar] [CrossRef] [PubMed]
- Majem, B.; Parrilla, A.; Jiménez, C.J.; Suárez-Cabrera, L.; Barber, M.; Marín, A.; Castellvi, J.; Tamayo, G.; Moreno-Bueno, G.; Ponce, J.; et al. MicroRNA-654-5p suppresses ovarian cancer development impacting on MYC, WNT and AKT pathways. Oncogene 2019, 38, 6035–6050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutschner, T.; Richtig, G.; Haemmerle, M.; Pichler, M. From biomarkers to therapeutic targets—the promises and perils of long non-coding RNAs in cancer. Cancer Metastasis Rev. 2017, 37, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, A.; Carninci, P. From bench to bedside: The long journey of long non-coding RNAs. Curr. Opin. Syst. Biol. 2017, 3, 119–124. [Google Scholar] [CrossRef]
- Watrin, M.; Von Pelchrzim, F.; Dausse, E.; Schroeder, R.; Toulme, J.-J. In Vitro Selection of RNA Aptamers Derived from a Genomic Human Library against the TAR RNA Element of HIV-1. Biochemistry 2009, 48, 6278–6284. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Chang, L.-C.; Chen, K.-B.; Wang, S.-C. Aptamer-guided targeting of the intracellular long-noncoding RNA HOTAIR. Am. J. Cancer Res. 2021, 11, 945–954. [Google Scholar]
- Xu, A.; Huang, M.-F.; Zhu, D.; Gingold, J.A.; Bazer, D.A.; Chang, B.; Wang, D.; Lai, C.-C.; Lemischka, I.R.; Zhao, R.; et al. LncRNA H19 Suppresses Osteosarcomagenesis by Regulating snoRNAs and DNA Repair Protein Complexes. Front. Genet. 2021, 11, 611823. [Google Scholar] [CrossRef]
- Rahbarizadeh, F.; Ahmadvand, D.; Sharifzadeh, Z. Nanobody; an Old Concept and New Vehicle for Immunotargeting. Immunol. Investig. 2011, 40, 299–338. [Google Scholar] [CrossRef] [PubMed]
- Van Audenhove, I.; Gettemans, J. Nanobodies as Versatile Tools to Understand, Diagnose, Visualize and Treat Cancer. EBioMedicine 2016, 8, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawez, F.; Duray, E.; Hu, Y.; Vandenameele, J.; Romão, E.; Vincke, C.; Dumoulin, M.; Galleni, M.; Muyldermans, S.; Vandevenne, M. Combinatorial Design of a Nanobody that Specifically Targets Structured RNAs. J. Mol. Biol. 2018, 430, 1652–1670. [Google Scholar] [CrossRef]
- Brown, J.A.; Bulkley, D.; Wang, J.; Valenstein, M.L.; Yario, T.A.; Steitz, T.A.; Steitz, J.A. Structural insights into the stabilization of MALAT1 noncoding RNA by a bipartite triple helix. Nat. Struct. Mol. Biol. 2014, 21, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Schmidt, B.F.; Bruchez, M.P.; McManus, C.J. Structural analyses of NEAT1 lncRNAs suggest long-range RNA interactions that may contribute to paraspeckle architecture. Nucleic Acids Res. 2018, 46, 3742–3752. [Google Scholar] [CrossRef]
- Somarowthu, S.; Legiewicz, M.; Chillón, I.; Marcia, M.; Liu, F.; Pyle, A.M. HOTAIR Forms an Intricate and Modular Secondary Structure. Mol. Cell 2015, 58, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Donlic, A.; Hargrove, A.E. Targeting RNA in mammalian systems with small molecules. Wiley Interdiscip. Rev. RNA 2018, 9, e1477. [Google Scholar] [CrossRef]
- Fatemi, R.P.; Velmeshev, D.; Faghihi, M.A. De-repressing LncRNA-Targeted Genes to Upregulate Gene Expression: Focus on Small Molecule Therapeutics. Mol. Ther.-Nucleic Acids 2014, 3, e196. [Google Scholar] [CrossRef]
- Fatemi, R.P.; Salah-Uddin, S.; Modarresi, F.; Khoury, N.; Wahlestedt, C.; Faghihi, M.A. Screening for Small-Molecule Modulators of Long Noncoding RNA-Protein Interactions Using AlphaScreen. J. Biomol. Screen. 2015, 20, 1132–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donlic, A.; Morgan, B.S.; Xu, J.L.; Liu, A.; Roble, J.C.; Hargrove, A.E. Discovery of Small Molecule Ligands for MALAT1 by Tuning an RNA-Binding Scaffold. Angew. Chem. Int. Ed. 2018, 57, 13242–13247. [Google Scholar] [CrossRef] [PubMed]
- Abulwerdi, F.A.; Xu, W.; Ageeli, A.A.; Yonkunas, M.; Arun, G.; Nam, H.; Schneekloth, J.S.; Dayie, T.K.; Spector, D.; Baird, N.; et al. Selective Small-Molecule Targeting of a Triple Helix Encoded by the Long Noncoding RNA, MALAT1. ACS Chem. Biol. 2019, 14, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Denichenko, P.; Mogilevsky, M.; Cléry, A.; Welte, T.; Biran, J.; Shimshon, O.; Barnabas, G.D.; Danan-Gotthold, M.; Kumar, S.; Yavin, E.; et al. Specific inhibition of splicing factor activity by decoy RNA oligonucleotides. Nat. Commun. 2019, 10, 1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-J.; Kim, J.; Li, S.; Zaia, J.; Yee, J.-K.; Anderson, J.; Akkina, R.; Rossi, J.J. Long-Term Inhibition of HIV-1 Infection in Primary Hematopoietic Cells by Lentiviral Vector Delivery of a Triple Combination of Anti-HIV shRNA, Anti-CCR5 Ribozyme, and a Nucleolar-Localizing TAR Decoy. Mol. Ther. 2005, 12, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tan, Z.; Zhang, Y.; Zhang, Z.; Hu, Q.; Liang, K.; Jun, Y.; Ye, Y.; Li, Y.-C.; Li, C.; et al. A noncoding RNA modulator potentiates phenylalanine metabolism in mice. Science 2021, 373, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, A.; Czerniak, A.; Levy, T.; Amiur, S.; Gallula, J.; Matouk, I.; Abu-Lail, R.; Sorin, V.; Birman, T.; De Groot, N.; et al. Development of targeted therapy for ovarian cancer mediated by a plasmid expressing diphtheria toxin under the control of H19 regulatory sequences. J. Transl. Med. 2009, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Lavie, O.; Edelman, D.; Levy, T.; Fishman, A.; Hubert, A.; Segev, Y.; Raveh, E.; Gilon, M.; Hochberg, A. A phase 1/2a, dose-escalation, safety, pharmacokinetic, and preliminary efficacy study of intraperitoneal administration of BC-819 (H19-DTA) in subjects with recurrent ovarian/peritoneal cancer. Arch. Gynecol. Obstet. 2017, 295, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Abildgaard, C.; Canto, L.M.D.; Steffensen, K.D.; Rogatto, S.R. Long Non-coding RNAs Involved in Resistance to Chemotherapy in Ovarian Cancer. Front. Oncol. 2020, 9, 1549. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Zhang, Y.; Xu, D.; Zhou, X.; Peng, P.; Pan, Z.; Xiao, N.; Yao, J.; Li, Z. Research Progress on Long Non-Coding RNA and Radiotherapy. Med. Sci. Monit. 2019, 25, 5757–5770. [Google Scholar] [CrossRef]
- Wang, F.; Zuroske, T.; Watts, J.K. RNA therapeutics on the rise. Nat. Rev. Drug Discov. 2020, 19, 441–442. [Google Scholar] [CrossRef] [PubMed]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Liu, F.; Yang, J.; Liu, X.; Meng, Y.; Deng, X.; Peng, C.; Tian, G.; Zhou, L. Probing lncRNA–Protein Interactions: Data Repositories, Models, and Algorithms. Front. Genet. 2020, 10, 1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yotsukura, S.; Duverle, D.; Hancock, T.; Natsume-Kitatani, Y.; Mamitsuka, H. Computational recognition for long non-coding RNA (lncRNA): Software and databases. Brief. Bioinform. 2017, 18, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Pinkney, H.R.; Wright, B.M.; Diermeier, S.D. The lncRNA Toolkit: Databases and In Silico Tools for lncRNA Analysis. Non-Coding RNA 2020, 6, 49. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- RNAcentral Consortium; Sweeney, B.; Petrov, A.; Ribas, C.; Finn, R.D.; Bateman, A.; Szymanski, M.; Karlowski, W.M.; Seemann, S.; Gorodkin, J.; et al. RNAcentral 2021: Secondary structure integration, improved sequence search and new member databases. Nucleic Acids Res. 2021, 49, D212–D220. [Google Scholar] [CrossRef]
- Volders, P.-J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. LNCipedia 5: Towards a reference set of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Cao, J.; Liu, L.; Du, Q.; Li, Z.; Zou, D.; Bajic, V.B.; Zhang, Z. LncBook: A curated knowledgebase of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D128–D134. [Google Scholar] [CrossRef] [Green Version]
- Amaral, P.P.; Clark, M.B.; Gascoigne, D.K.; Dinger, M.E.; Mattick, J.S. lncRNAdb: A reference database for long noncoding RNAs. Nucleic Acids Res. 2011, 39, D146–D151. [Google Scholar] [CrossRef] [Green Version]
- Bhartiya, D.; Pal, K.; Ghosh, S.; Kapoor, S.; Jalali, S.; Panwar, B.; Jain, S.; Sati, S.; Sengupta, S.; Sachidanandan, C.; et al. lncRNome: A comprehensive knowledgebase of human long noncoding RNAs. Database 2013, 2013, bat034. [Google Scholar] [CrossRef] [Green Version]
- The GTEx Consortium The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 2020, 369, 1318–1330. [CrossRef]
- Clough, E.; Barrett, T. The Gene Expression Omnibus Database. Methods Mol. Biol. 2016, 1418, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Papatheodorou, I.; Moreno, P.; Manning, J.; Fuentes, A.M.-P.; George, N.; Fexova, S.; Fonseca, N.; Füllgrabe, A.; Green, M.; Huang, N.; et al. Expression Atlas update: From tissues to single cells. Nucleic Acids Res. 2019, 48, D77–D83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S.; Zhang, L.; Guo, J.; Niu, Y.; Wu, Y.; Li, H.; Zhao, L.; Li, X.; Teng, X.; Sun, X.; et al. NONCODEV5: A comprehensive annotation database for long non-coding RNAs. Nucleic Acids Res. 2018, 46, D308–D314. [Google Scholar] [CrossRef] [PubMed]
- Dinger, M.E.; Pang, K.C.; Mercer, T.R.; Crowe, M.L.; Grimmond, S.M.; Mattick, J.S. NRED: A database of long noncoding RNA expression. Nucleic Acids Res. 2009, 37, D122–D126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Liu, L.; Jiang, S.; Li, Q.; Feng, C.; Du, Q.; Zou, D.; Xiao, J.; Zhang, Z.; Ma, L. LncExpDB: An expression database of human long non-coding RNAs. Nucleic Acids Res. 2021, 49, D962–D968. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.; Xu, K.; Jin, X.; Li, J.; Shi, Y.; Zhang, M.; Jin, X.; Li, Y.; Xu, J.; Li, X. LncSpA: LncRNA Spatial Atlas of Expression across Normal and Cancer Tissues. Cancer Res. 2020, 80, 2067–2071. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Han, L.; Roebuck, P.; Diao, L.; Liu, L.; Yuan, Y.; Weinstein, J.N.; Liang, H. TANRIC: An Interactive Open Platform to Explore the Function of lncRNAs in Cancer. Cancer Res. 2015, 75, 3728–3737. [Google Scholar] [CrossRef] [Green Version]
- Bao, Z.; Yang, Z.; Huang, Z.; Zhou, Y.; Cui, Q.; Dong, D. LncRNADisease 2.0: An updated database of long non-coding RNA-associated diseases. Nucleic Acids Res. 2019, 47, D1034–D1037. [Google Scholar] [CrossRef]
- Gao, Y.; Shang, S.; Guo, S.; Li, X.; Zhou, H.; Liu, H.; Sun, Y.; Wang, J.; Wang, P.; Zhi, H.; et al. Lnc2Cancer 3.0: An updated resource for experimentally supported lncRNA/circRNA cancer associations and web tools based on RNA-seq and scRNA-seq data. Nucleic Acids Res. 2021, 49, D1251–D1258. [Google Scholar] [CrossRef]
- Miao, Y.-R.; Liu, W.; Zhang, Q.; Guo, A.-Y. lncRNASNP2: An updated database of functional SNPs and mutations in human and mouse lncRNAs. Nucleic Acids Res. 2018, 46, D276–D280. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; An, G.; Zhao, C.; Ouyang, Z.; Bo, X.; Shu, W. Lnc2Catlas: An atlas of long noncoding RNAs associated with risk of cancers. Sci. Rep. 2018, 8, 1909. [Google Scholar] [CrossRef] [Green Version]
- Paraskevopoulou, M.D.; Karagkouni, D.; Vlachos, I.S.; Tastsoglou, S.; Hatzigeorgiou, A.G. microCLIP super learning framework uncovers functional transcriptome-wide miRNA interactions. Nat. Commun. 2018, 9, 3601. [Google Scholar] [CrossRef] [PubMed]
- Kirk, J.M.; Kim, S.O.; Inoue, K.; Smola, M.J.; Lee, D.M.; Schertzer, M.D.; Wooten, J.S.; Baker, A.R.; Sprague, D.; Collins, D.W.; et al. Functional classification of long non-coding RNAs by k-mer content. Nat. Genet. 2018, 50, 1474–1482. [Google Scholar] [CrossRef]
- Zhou, Y.-K.; Shen, Z.-A.; Yu, H.; Luo, T.; Gao, Y.; Du, P.-F. Predicting lncRNA–Protein Interactions With miRNAs as Mediators in a Heterogeneous Network Model. Front. Genet. 2020, 10, 1341. [Google Scholar] [CrossRef]
- Ma, L.; Cao, J.; Liu, L.; Li, Z.; Shireen, H.; Pervaiz, N.; Batool, F.; Raza, R.Z.; Zou, D.; Bao, Y.; et al. Community Curation and Expert Curation of Human Long Noncoding RNAs with LncRNAWiki and LncBook. Curr. Protoc. Bioinform. 2019, 67, e82. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.K.; Doxtater, K.; Keramatnia, F.; Zacheaus, C.; Yallapu, M.; Jaggi, M.; Chauhan, S.C. Role of lncRNAs in ovarian cancer: Defining new biomarkers for therapeutic purposes. Drug Discov. Today 2018, 23, 1635–1643. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabol, M.; Calleja-Agius, J.; Di Fiore, R.; Suleiman, S.; Ozcan, S.; Ward, M.P.; Ozretić, P. (In)Distinctive Role of Long Non-Coding RNAs in Common and Rare Ovarian Cancers. Cancers 2021, 13, 5040. https://doi.org/10.3390/cancers13205040
Sabol M, Calleja-Agius J, Di Fiore R, Suleiman S, Ozcan S, Ward MP, Ozretić P. (In)Distinctive Role of Long Non-Coding RNAs in Common and Rare Ovarian Cancers. Cancers. 2021; 13(20):5040. https://doi.org/10.3390/cancers13205040
Chicago/Turabian StyleSabol, Maja, Jean Calleja-Agius, Riccardo Di Fiore, Sherif Suleiman, Sureyya Ozcan, Mark P. Ward, and Petar Ozretić. 2021. "(In)Distinctive Role of Long Non-Coding RNAs in Common and Rare Ovarian Cancers" Cancers 13, no. 20: 5040. https://doi.org/10.3390/cancers13205040
APA StyleSabol, M., Calleja-Agius, J., Di Fiore, R., Suleiman, S., Ozcan, S., Ward, M. P., & Ozretić, P. (2021). (In)Distinctive Role of Long Non-Coding RNAs in Common and Rare Ovarian Cancers. Cancers, 13(20), 5040. https://doi.org/10.3390/cancers13205040