
Galectins in Glioma: Current Roles in Cancer Progression and Future Directions for Improving Treatment


{kind=link}
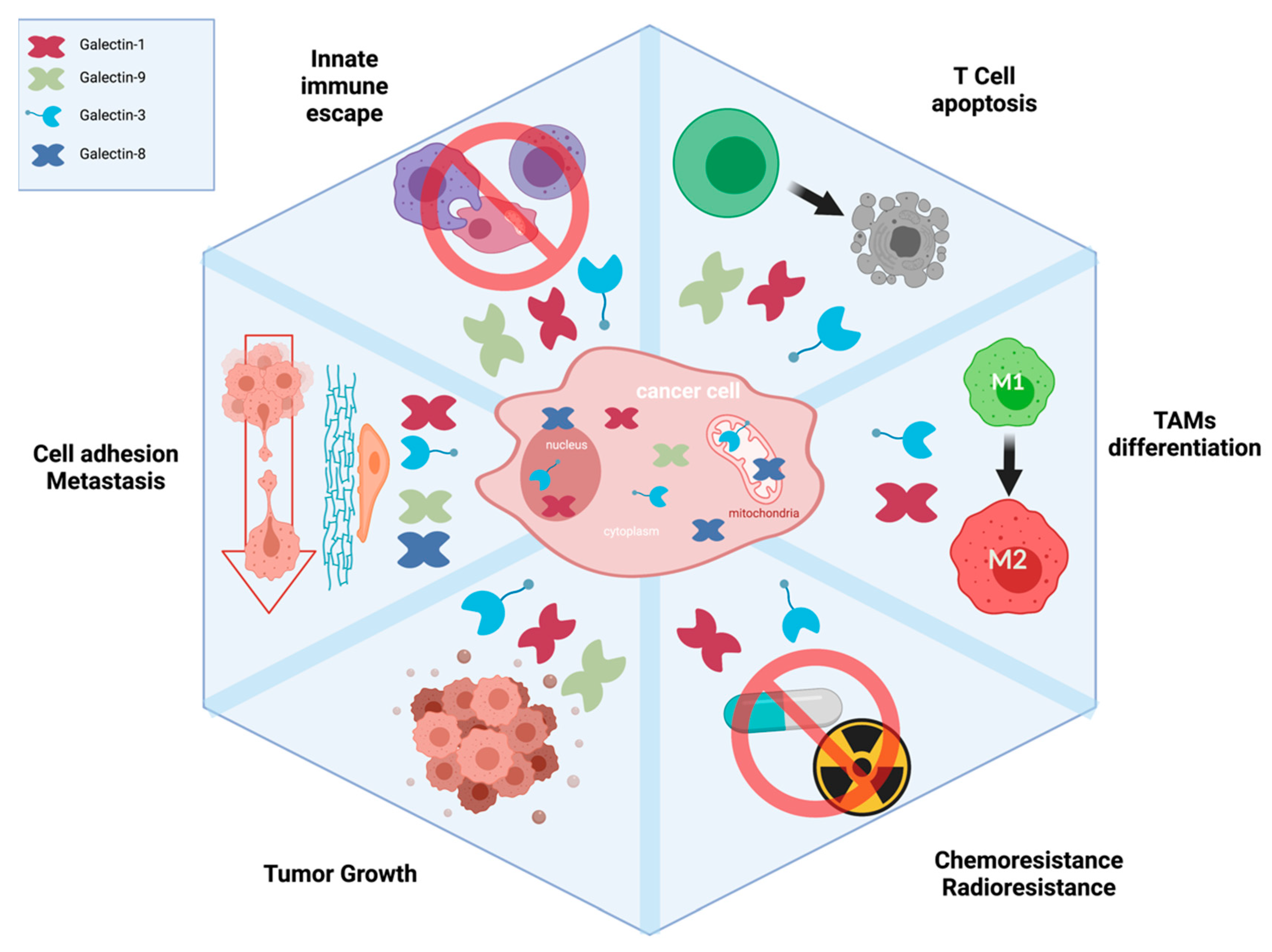{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Galectins and Their Implication in Cancer Progression
3. The Case of Galectin-1
4. The Case of Galectin-3
5. The Case of Galectin-8
6. The Case of Galectin-9
7. Approaches for Targeting and Delivery of Galectins
8. Conclusions
Funding
Conflicts of Interest
Abbreviations
| BBB | blood–brain barrier |
| GBM | glioblastoma |
| MoA | mechanism of action |
| TMZ | telozolomide |
| ICI’s | immune checkpoint inhibitors |
| VEGF | vascular endothelial growth factor |
| GBS | glycan-binding site |
| CRD | carbohydrate recognition domain |
| TME | tumor microenvironment |
| Nbs | nanobodies |
References
- Ostrom, Q.T.; Bauchet, L.; Davis, F.G.; Deltour, I.; Fisher, J.L.; Langer, C.E.; Pekmezci, M.; Schwartzbaum, J.A.; Turner, M.C.; Walsh, K.M.; et al. The Epidemiology of Glioma in Adults: A “State of the Science” Review. Neuro-Oncol. 2014, 16, 896–913. [Google Scholar] [CrossRef] [Green Version]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO Classification of Tumours of the Central Nervous System. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Smith-Cohn, M.; Cohen, A.L.; Colman, H. Glioma Subclassifications and Their Clinical Significance. Neurother. J. Am. Soc. Exp. Neurother. 2017, 14, 284–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hata, N.; Mizoguchi, M.; Kuga, D.; Hatae, R.; Akagi, Y.; Sangatsuda, Y.; Amemiya, T.; Michiwaki, Y.; Fujioka, Y.; Takigawa, K.; et al. First-Line Bevacizumab Contributes to Survival Improvement in Glioblastoma Patients Complementary to Temozolomide. J. Neurooncol. 2020, 146, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.K.; Sulman, E.P.; Wen, P.Y.; Kurz, S.C. Novel Therapies for Glioblastoma. Curr. Neurol. Neurosci. Rep. 2020, 20, 19. [Google Scholar] [CrossRef] [PubMed]
- Teichberg, V.I.; Silman, I.; Beitsch, D.D.; Resheff, G. A Beta-D-Galactoside Binding Protein from Electric Organ Tissue of Electrophorus Electricus. Proc. Natl. Acad. Sci. USA 1975, 72, 1383–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, K.; Hirabayashi, J. Galectins: A Family of Animal Lectins That Decipher Glycocodes. J. Biochem. 1996, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vasta, G.R.; Ahmed, H.; Nita-Lazar, M.; Banerjee, A.; Pasek, M.; Shridhar, S.; Guha, P.; Fernández-Robledo, J.A. Galectins as Self/Non-Self Recognition Receptors in Innate and Adaptive Immunity: An Unresolved Paradox. Front. Immunol. 2012, 3, 199. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Gao, J.; Si, Y.; Cui, L.; Song, C.; Wang, Y.; Wu, R.; Tai, G.; Zhou, Y. Galectin-10: A New Structural Type of Prototype Galectin Dimer and Effects on Saccharide Ligand Binding. Glycobiology 2018, 28, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grozdanovic, M.M.; Doyle, C.B.; Liu, L.; Maybruck, B.; Kwatia, M.A.; Thiyagarajan, N.; Acharya, K.R.; Ackerman, S.J. Charcot-Leyden Crystal Protein/Galectin-10 Interacts with Cationic Ribonucleases and Is Required for Eosinophil Granulogenesis. J. Allergy Clin. Immunol. 2020, 146, 377–389.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persson, E.K.; Verstraete, K.; Heyndrickx, I.; Gevaert, E.; Aegerter, H.; Percier, J.-M.; Deswarte, K.; Verschueren, K.H.G.; Dansercoer, A.; Gras, D.; et al. Protein Crystallization Promotes Type 2 Immunity and Is Reversible by Antibody Treatment. Science 2019, 364, eaaw4295. [Google Scholar] [CrossRef] [PubMed]
- Mey, A.; Leffler, H.; Hmama, Z.; Normier, G.; Revillard, J.P. The Animal Lectin Galectin-3 Interacts with Bacterial Lipopolysaccharides via Two Independent Sites. J. Immunol. Baltim. 1996, 156, 1572–1577. [Google Scholar]
- Gauthier, L.; Rossi, B.; Roux, F.; Termine, E.; Schiff, C. Galectin-1 Is a Stromal Cell Ligand of the Pre-B Cell Receptor (BCR) Implicated in Synapse Formation between Pre-B and Stromal Cells and in Pre-BCR Triggering. Proc. Natl. Acad. Sci. USA 2002, 99, 13014–13019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepur, A.; Salomonsson, E.; Nilsson, U.J.; Leffler, H. Ligand Induced Galectin-3 Protein Self-Association. J. Biol. Chem. 2012, 287, 21751–21756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yıldırım, C.; Vogel, D.Y.S.; Hollander, M.R.; Baggen, J.M.; Fontijn, R.D.; Nieuwenhuis, S.; Haverkamp, A.; de Vries, M.R.; Quax, P.H.A.; Garcia-Vallejo, J.J.; et al. Galectin-2 Induces a Proinflammatory, Anti-Arteriogenic Phenotype in Monocytes and Macrophages. PLoS ONE 2015, 10, e0124347. [Google Scholar] [CrossRef] [PubMed]
- Advedissian, T.; Proux-Gillardeaux, V.; Nkosi, R.; Peyret, G.; Nguyen, T.; Poirier, F.; Viguier, M.; Deshayes, F. E-Cadherin Dynamics Is Regulated by Galectin-7 at Epithelial Cell Surface. Sci. Rep. 2017, 7, 17086. [Google Scholar] [CrossRef] [Green Version]
- Girotti, M.R.; Salatino, M.; Dalotto-Moreno, T.; Rabinovich, G.A. Sweetening the Hallmarks of Cancer: Galectins as Multifunctional Mediators of Tumor Progression. J. Exp. Med. 2020, 217, e20182041. [Google Scholar] [CrossRef] [PubMed]
- Camby, I.; Belot, N.; Rorive, S.; Lefranc, F.; Maurage, C.A.; Lahm, H.; Kaltner, H.; Hadari, Y.; Ruchoux, M.M.; Brotchi, J.; et al. Galectins Are Differentially Expressed in Supratentorial Pilocytic Astrocytomas, Astrocytomas, Anaplastic Astrocytomas and Glioblastomas, and Significantly Modulate Tumor Astrocyte Migration. Brain Pathol. Zur. Switz. 2001, 11, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Rorive, S.; Belot, N.; Decaestecker, C.; Lefranc, F.; Gordower, L.; Micik, S.; Maurage, C.A.; Kaltner, H.; Ruchoux, M.M.; Danguy, A.; et al. Galectin-1 Is Highly Expressed in Human Gliomas with Relevance for Modulation of Invasion of Tumor Astrocytes into the Brain Parenchyma. Glia 2001, 33, 241–255. [Google Scholar] [CrossRef]
- Toussaint, L.G.; Nilson, A.E.; Goble, J.M.; Ballman, K.V.; James, C.D.; Lefranc, F.; Kiss, R.; Uhm, J.H. Galectin-1, a Gene Preferentially Expressed at the Tumor Margin, Promotes Glioblastoma Cell Invasion. Mol. Cancer 2012, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Han, B.; Meng, X.; Duan, C.; Yang, C.; Wu, Z.; Magafurov, D.; Zhao, S.; Safin, S.; Jiang, C.; et al. Immunogenomic Analysis Reveals LGALS1 Contributes to the Immune Heterogeneity and Immunosuppression in Glioma. Int. J. Cancer 2019, 145, 517–530. [Google Scholar] [CrossRef]
- Chou, S.-Y.; Yen, S.-L.; Huang, C.-C.; Huang, E.-Y. Galectin-1 Is a Poor Prognostic Factor in Patients with Glioblastoma Multiforme after Radiotherapy. BMC Cancer 2018, 18, 105. [Google Scholar] [CrossRef] [Green Version]
- Shih, W.; Yamada, S. N-Cadherin as a Key Regulator of Collective Cell Migration in a 3D Environment. Cell Adhes. Migr. 2012, 6, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Eto, K.; Puzon-McLaughlin, W.; Sheppard, D.; Sehara-Fujisawa, A.; Zhang, X.P.; Takada, Y. RGD-Independent Binding of Integrin Alpha9beta1 to the ADAM-12 and -15 Disintegrin Domains Mediates Cell-Cell Interaction. J. Biol. Chem. 2000, 275, 34922–34930. [Google Scholar] [CrossRef] [Green Version]
- Paulus, W.; Baur, I.; Beutler, A.S.; Reeves, S.A. Diffuse Brain Invasion of Glioma Cells Requires Beta 1 Integrins. Lab. Investig. J. Tech. Methods Pathol. 1996, 75, 819–826. [Google Scholar]
- D′Abaco, G.M.; Kaye, A.H. Integrins: Molecular Determinants of Glioma Invasion. J. Clin. Neurosci. Off. J. Neurosurg. Soc. Australas. 2007, 14, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.R.; Wu, M.Y.; Dai, L.J.; Huang, Y.; Shan, M.Y.; Ma, S.N.; Wang, J.; Peng, H.; Ding, Y.; Zhang, Q.F.; et al. Nuclear FAM289-Galectin-1 Interaction Controls FAM289-Mediated Tumor Promotion in Malignant Glioma. J. Exp. Clin. Cancer Res. 2019, 38, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozeki, Y.; Matsui, T.; Yamamoto, Y.; Funahashi, M.; Hamako, J.; Titani, K. Tissue Fibronectin Is an Endogenous Ligand for Galectin-1. Glycobiology 1995, 5, 255–261. [Google Scholar] [CrossRef]
- Moiseeva, E.P.; Spring, E.L.; Baron, J.H.; de Bono, D.P. Galectin 1 Modulates Attachment, Spreading and Migration of Cultured Vascular Smooth Muscle Cells via Interactions with Cellular Receptors and Components of Extracellular Matrix. J. Vasc. Res. 1999, 36, 47–58. [Google Scholar] [CrossRef]
- Moiseeva, E.P.; Williams, B.; Goodall, A.H.; Samani, N.J. Galectin-1 Interacts with Beta-1 Subunit of Integrin. Biochem. Biophys. Res. Commun. 2003, 310, 1010–1016. [Google Scholar] [CrossRef]
- Fortin, S.; Le Mercier, M.; Camby, I.; Spiegl-Kreinecker, S.; Berger, W.; Lefranc, F.; Kiss, R. Galectin-1 Is Implicated in the Protein Kinase C ε/Vimentin-Controlled Trafficking of Integrin-Β1 in Glioblastoma Cells. Brain Pathol. Zur. Switz. 2010, 20, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Camby, I.; Belot, N.; Lefranc, F.; Sadeghi, N.; de Launoit, Y.; Kaltner, H.; Musette, S.; Darro, F.; Danguy, A.; Salmon, I.; et al. Galectin-1 Modulates Human Glioblastoma Cell Migration into the Brain Through Modifications to the Actin Cytoskeleton and Levels of Expression of Small GTPases. J. Neuropathol. Exp. Neurol. 2002, 61, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Bros, M.; Haas, K.; Moll, L.; Grabbe, S. RhoA as a Key Regulator of Innate and Adaptive Immunity. Cells 2019, 8, 733. [Google Scholar] [CrossRef] [Green Version]
- Le Mercier, M.; Fortin, S.; Mathieu, V.; Kiss, R.; Lefranc, F. Galectins and Gliomas. Brain Pathol. Zur. Switz. 2010, 20, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, W. Knockdown of Galectin-1 Facilitated Cisplatin Sensitivity by Inhibiting Autophagy in Neuroblastoma Cells. Chem. Biol. Interact. 2019, 297, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-F.; Wu, J.; Luo, J.-H.; Li, K.-S.; Wang, F.; Huang, W.; Wu, Y.; Gao, S.-P.; Zhang, X.-M.; Zhang, P.-N. SNHG22 Overexpression Indicates Poor Prognosis and Induces Chemotherapy Resistance via the MiR-2467/Gal-1 Signaling Pathway in Epithelial Ovarian Carcinoma. Aging 2019, 11, 8204–8216. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, A.; Li, S.; Tao, X.; Sheng, B.; Chetry, M.; Zhu, X. Predictive Role of Galectin-1 and Integrin A5β1 in Cisplatin-Based Neoadjuvant Chemotherapy of Bulky Squamous Cervical Cancer. Biosci. Rep. 2017, 37, BSR20170958. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Zhang, P.; Shi, B.; Zhou, M.; Jiang, H.; Zhang, H.; Pan, X.; Gao, H.; Sun, H.; Li, Z. Galectin-1 Overexpression Promotes Progression and Chemoresistance to Cisplatin in Epithelial Ovarian Cancer. Cell Death Dis. 2014, 5, e991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, L.-Y.; Tang, S.-J.; Sun, G.-H.; Chou, T.-Y.; Yeh, T.-S.; Yu, S.-L.; Sun, K.-H. Galectin-1 Promotes Lung Cancer Progression and Chemoresistance by Upregulating P38 MAPK, ERK, and Cyclooxygenase-2. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 4037–4047. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Brachova, P.; Yang, S.; Xiong, Z.; Zhang, Y.; Thiel, K.W.; Leslie, K.K. Knockdown of MTDH Sensitizes Endometrial Cancer Cells to Cell Death Induction by Death Receptor Ligand TRAIL and HDAC Inhibitor LBH589 Co-Treatment. PLoS ONE 2011, 6, e20920. [Google Scholar] [CrossRef] [PubMed]
- van der Burg, S.H.; Arens, R.; Ossendorp, F.; van Hall, T.; Melief, C.J.M. Vaccines for Established Cancer: Overcoming the Challenges Posed by Immune Evasion. Nat. Rev. Cancer 2016, 16, 219–233. [Google Scholar] [CrossRef] [PubMed]
- RodrÍguez, E.; Schetters, S.T.T.; van Kooyk, Y. The Tumour Glyco-Code as a Novel Immune Checkpoint for Immunotherapy. Nat. Rev. Immunol. 2018, 18, 204–211. [Google Scholar] [CrossRef]
- Alard, E.; Butnariu, A.-B.; Grillo, M.; Kirkham, C.; Zinovkin, D.A.; Newnham, L.; Macciochi, J.; Pranjol, M.Z.I. Advances in Anti-Cancer Immunotherapy: Car-T Cell, Checkpoint Inhibitors, Dendritic Cell Vaccines, and Oncolytic Viruses, and Emerging Cellular and Molecular Targets. Cancers 2020, 12, 1826. [Google Scholar] [CrossRef] [PubMed]
- Berois, N.; Osinaga, E. Glycobiology of Neuroblastoma: Impact on Tumor Behavior, Prognosis, and Therapeutic Strategies. Front. Oncol. 2014, 4, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagnoni, A.J.; Giribaldi, M.L.; Blidner, A.G.; Cutine, A.M.; Gatto, S.G.; Morales, R.M.; Salatino, M.; Abba, M.C.; Croci, D.O.; Mariño, K.V.; et al. Galectin-1 Fosters an Immunosuppressive Microenvironment in Colorectal Cancer by Reprogramming CD8+ Regulatory T Cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2102950118. [Google Scholar] [CrossRef]
- Van Woensel, M.; Mathivet, T.; Wauthoz, N.; Rosière, R.; Garg, A.D.; Agostinis, P.; Mathieu, V.; Kiss, R.; Lefranc, F.; Boon, L.; et al. Sensitization of Glioblastoma Tumor Micro-Environment to Chemo- and Immunotherapy by Galectin-1 Intranasal Knock-down Strategy. Sci. Rep. 2017, 7, 1217. [Google Scholar] [CrossRef]
- Lowenstein, P.R.; Baker, G.J.; Castro, M.G. Cracking the Glioma-NK Inhibitory Code: Toward Successful Innate Immunotherapy. Oncoimmunology 2014, 3, e965573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, D.; Comba, A.; Faisal, S.M.; Kadiyala, P.; Baker, G.J.; Alghamri, M.S.; Doherty, R.; Zamler, D.; Nuñez, G.; Castro, M.G.; et al. A Novel MiR1983-TLR7-IFNβ Circuit Licenses NK Cells to Kill Glioma Cells, and Is under the Control of Galectin-1. Oncoimmunology 2021, 10, 1939601. [Google Scholar] [CrossRef] [PubMed]
- Perillo, N.L.; Pace, K.E.; Seilhamer, J.J.; Baum, L.G. Apoptosis of T Cells Mediated by Galectin-1. Nature 1995, 378, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Matarrese, P.; Tinari, A.; Mormone, E.; Bianco, G.A.; Toscano, M.A.; Ascione, B.; Rabinovich, G.A.; Malorni, W. Galectin-1 Sensitizes Resting Human T Lymphocytes to Fas (CD95)-Mediated Cell Death via Mitochondrial Hyperpolarization, Budding, and Fission. J. Biol. Chem. 2005, 280, 6969–6985. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.-T.; Rabinovich, G.A. Galectins as Modulators of Tumour Progression. Nat. Rev. Cancer 2005, 5, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, D.K.; Aguilera, T.; Cao, H.; Kwok, S.; Kong, C.; Bloomstein, J.; Wang, Z.; Rangan, V.S.; Jiang, D.; Eyben, R.V.; et al. Galectin-1-Driven T Cell Exclusion in the Tumor Endothelium Promotes Immunotherapy Resistance. J. Clin. Investig. 2019, 129, 5553–5567. [Google Scholar] [CrossRef] [Green Version]
- Capalbo, C.; Filetti, M.; Scafetta, G.; Marchetti, P.; Bartolazzi, A. Galectins Role as Predictive Markers for Anti-PD-1-Based Immunotherapy in Non-Small Cell Lung Cancer. J. Clin. Oncol. 2019, 37 (Suppl. S8), 111. [Google Scholar] [CrossRef]
- Verschuere, T.; Toelen, J.; Maes, W.; Poirier, F.; Boon, L.; Tousseyn, T.; Mathivet, T.; Gerhardt, H.; Mathieu, V.; Kiss, R.; et al. Glioma-Derived Galectin-1 Regulates Innate and Adaptive Antitumor Immunity. Int. J. Cancer 2014, 134, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Roff, C.F.; Wang, J.L. Endogenous Lectins from Cultured Cells. Isolation and Characterization of Carbohydrate-Binding Proteins from 3T3 Fibroblasts. J. Biol. Chem. 1983, 258, 10657–10663. [Google Scholar] [CrossRef]
- Barondes, S.H.; Cooper, D.N.; Gitt, M.A.; Leffler, H. Galectins. Structure and Function of a Large Family of Animal Lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [CrossRef]
- Fortuna-Costa, A.; Gomes, A.M.; Kozlowski, E.O.; Stelling, M.P.; Pavão, M.S.G. Extracellular Galectin-3 in Tumor Progression and Metastasis. Front. Oncol. 2014, 4, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nangia-Makker, P.; Hogan, V.; Raz, A. Galectin-3 and Cancer Stemness. Glycobiology 2018, 28, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagher, S.F.; Wang, J.L.; Patterson, R.J. Identification of Galectin-3 as a Factor in Pre-MRNA Splicing. Proc. Natl. Acad. Sci. USA 1995, 92, 1213–1217. [Google Scholar] [CrossRef] [Green Version]
- Califice, S.; Castronovo, V.; Bracke, M.; van den Brûle, F. Dual Activities of Galectin-3 in Human Prostate Cancer: Tumor Suppression of Nuclear Galectin-3 vs. Tumor Promotion of Cytoplasmic Galectin-3. Oncogene 2004, 23, 7527–7536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binh, N.H.; Satoh, K.; Kobayashi, K.; Takamatsu, M.; Hatano, Y.; Hirata, A.; Tomita, H.; Kuno, T.; Hara, A. Galectin-3 in Preneoplastic Lesions of Glioma. J. Neurooncol. 2013, 111, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.-I.; Kim, E.-G.; Lee, E.-J.; Hong, S.-Y.; Yoon, C.-S.; Hong, M.-J.; Park, S.-J.; Woo, R.-S.; Baik, T.-K.; Song, D.-Y. Neuroanatomical Distribution of Galectin-3 in the Adult Rat Brain. J. Mol. Histol. 2017, 48, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Seguin, L.; Odouard, S.; Corlazzoli, F.; Haddad, S.A.; Moindrot, L.; Calvo Tardón, M.; Yebra, M.; Koval, A.; Marinari, E.; Bes, V.; et al. Macropinocytosis Requires Gal-3 in a Subset of Patient-Derived Glioblastoma Stem Cells. Commun. Biol. 2021, 4, 718. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-M.; Yang, Y.-Z.; Zhang, T.-Z.; Qin, C.-F.; Li, X.-N. LGALS3 Is a Poor Prognostic Factor in Diffusely Infiltrating Gliomas and Is Closely Correlated With CD163+ Tumor-Associated Macrophages. Front. Med. 2020, 7, 182. [Google Scholar] [CrossRef]
- Wang, H.; Song, X.; Huang, Q.; Xu, T.; Yun, D.; Wang, Y.; Hu, L.; Yan, Y.; Chen, H.; Lu, D.; et al. LGALS3 Promotes Treatment Resistance in Glioblastoma and Is Associated with Tumor Risk and Prognosis. Cancer Epidemiol. Prev. Biomark. 2019, 28, 760–769. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Zhang, S.; Chen, J.; Li, D. Increased LGALS3 Expression Independently Predicts Shorter Overall Survival in Patients with the Proneural Subtype of Glioblastoma. Cancer Med. 2019, 8, 2031–2040. [Google Scholar] [CrossRef]
- Chen, H.; Zheng, Z.; Yuan, B.; Liu, Z.; Jing, J.; Wang, S.-S. The Effect of Galectin-3 Genetic Variants on the Susceptibility and Prognosis of Gliomas in a Chinese Population. Neurosci. Lett. 2012, 518, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Takashima, Y.; Kawaguchi, A.; Hayano, A.; Yamanaka, R. CD276 and the Gene Signature Composed of GATA3 and LGALS3 Enable Prognosis Prediction of Glioblastoma Multiforme. PLoS ONE 2019, 14, e0216825. [Google Scholar] [CrossRef]
- Saleh, A.; Marhuenda, E.; Fabre, C.; Hassani, Z.; de Weille, J.; Boukhaddaoui, H.; Guelfi, S.; Maldonado, I.L.; Hugnot, J.-P.; Duffau, H.; et al. A Novel 3D Nanofibre Scaffold Conserves the Plasticity of Glioblastoma Stem Cell Invasion by Regulating Galectin-3 and Integrin-Β1 Expression. Sci. Rep. 2019, 9, 14612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikemori, R.Y.; Machado, C.M.L.; Furuzawa, K.M.; Nonogaki, S.; Osinaga, E.; Umezawa, K.; de Carvalho, M.A.; Verinaud, L.; Chammas, R. Galectin-3 up-Regulation in Hypoxic and Nutrient Deprived Microenvironments Promotes Cell Survival. PLoS ONE 2014, 9, e111592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akahani, S.; Nangia-Makker, P.; Inohara, H.; Kim, H.R.; Raz, A. Galectin-3: A Novel Antiapoptotic Molecule with a Functional BH1 (NWGR) Domain of Bcl-2 Family. Cancer Res. 1997, 57, 5272–5276. [Google Scholar]
- Shalom-Feuerstein, R.; Levy, R.; Makovski, V.; Raz, A.; Kloog, Y. Galectin-3 Regulates RasGRP4-Mediated Activation of N-Ras and H-Ras. Biochim. Biophys. Acta 2008, 1783, 985–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukumori, T.; Takenaka, Y.; Oka, N.; Yoshii, T.; Hogan, V.; Inohara, H.; Kanayama, H.-O.; Kim, H.-R.C.; Raz, A. Endogenous Galectin-3 Determines the Routing of CD95 Apoptotic Signaling Pathways. Cancer Res. 2004, 64, 3376–3379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenaka, Y.; Fukumori, T.; Yoshii, T.; Oka, N.; Inohara, H.; Kim, H.-R.C.; Bresalier, R.S.; Raz, A. Nuclear Export of Phosphorylated Galectin-3 Regulates Its Antiapoptotic Activity in Response to Chemotherapeutic Drugs. Mol. Cell. Biol. 2004, 24, 4395–4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nangia-Makker, P.; Balan, V.; Raz, A. Regulation of Tumor Progression by Extracellular Galectin-3. Cancer Microenviron. 2008, 1, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon-Alonso, M.; Bruger, A.M.; van der Bruggen, P. Extracellular Galectins as Controllers of Cytokines in Hematological Cancer. Blood 2018, 132, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Demotte, N.; Wieërs, G.; Smissen, P.V.D.; Moser, M.; Schmidt, C.; Thielemans, K.; Squifflet, J.-L.; Weynand, B.; Carrasco, J.; Lurquin, C.; et al. A Galectin-3 Ligand Corrects the Impaired Function of Human CD4 and CD8 Tumor-Infiltrating Lymphocytes and Favors Tumor Rejection in Mice. Cancer Res. 2010, 70, 7476–7488. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.; Jiang, Y.; Li, Z.; Wu, L.; Santiago, U.; Zou, H.; Cai, C.; Sharma, V.; Guan, Y.; McCarl, L.H.; et al. Chitinase-3-like 1 Protein Complexes Modulate Macrophage-Mediated Immune Suppression in Glioblastoma. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Gordon-Alonso, M.; Hirsch, T.; Wildmann, C.; van der Bruggen, P. Galectin-3 Captures Interferon-Gamma in the Tumor Matrix Reducing Chemokine Gradient Production and T-Cell Tumor Infiltration. Nat. Commun. 2017, 8, 793. [Google Scholar] [CrossRef] [PubMed]
- Curti, B.D.; Koguchi, Y.; Leidner, R.S.; Rolig, A.S.; Sturgill, E.R.; Sun, Z.; Wu, Y.; Rajamanickam, V.; Bernard, B.; Hilgart-Martiszus, I.; et al. Enhancing Clinical and Immunological Effects of Anti-PD-1 with Belapectin, a Galectin-3 Inhibitor. J. Immunother. Cancer 2021, 9, e002371. [Google Scholar] [CrossRef] [PubMed]
- Sturgill, E.R.; Rolig, A.S.; Linch, S.N.; Mick, C.; Kasiewicz, M.J.; Sun, Z.; Traber, P.G.; Shlevin, H.; Redmond, W.L. Galectin-3 Inhibition with Belapectin Combined with Anti-OX40 Therapy Reprograms the Tumor Microenvironment to Favor Anti-Tumor Immunity. Oncoimmunology 2021, 10, 1892265. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, P.; Zhang, Y.; Han, L.; Hu, Z.; Cai, Z.; Cai, J. Inhibition of Galectin-3 Augments the Antitumor Efficacy of PD-L1 Blockade in Non-Small-Cell Lung Cancer. FEBS Open Bio 2021, 11, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Hadari, Y.R.; Paz, K.; Dekel, R.; Mestrovic, T.; Accili, D.; Zick, Y. Galectin-8. A New Rat Lectin, Related to Galectin-4. J. Biol. Chem. 1995, 270, 3447–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadari, Y.R.; Eisenstein, M.; Zakut, R.; Zick, Y. Galectin-8: On the Road from Structure to Function. Trends Glycosci. Glycotechnol. 1997, 9, 103–112. [Google Scholar] [CrossRef]
- Levy, Y.; Arbel-Goren, R.; Hadari, Y.R.; Eshhar, S.; Ronen, D.; Elhanany, E.; Geiger, B.; Zick, Y. Galectin-8 Functions as a Matricellular Modulator of Cell Adhesion. J. Biol. Chem. 2001, 276, 31285–31295. [Google Scholar] [CrossRef] [Green Version]
- Ferragut, F.; Cagnoni, A.J.; Colombo, L.L.; Sánchez Terrero, C.; Wolfenstein-Todel, C.; Troncoso, M.F.; Vanzulli, S.I.; Rabinovich, G.A.; Mariño, K.V.; Elola, M.T. Dual Knockdown of Galectin-8 and Its Glycosylated Ligand, the Activated Leukocyte Cell Adhesion Molecule (ALCAM/CD166), Synergistically Delays in Vivo Breast Cancer Growth. Biochim. Biophys. Acta—Mol. Cell Res. 2019, 1866, 1338–1352. [Google Scholar] [CrossRef]
- Levy, Y.; Ronen, D.; Bershadsky, A.D.; Zick, Y. Sustained Induction of ERK, Protein Kinase B, and P70 S6 Kinase Regulates Cell Spreading and Formation of F-Actin Microspikes upon Ligation of Integrins by Galectin-8, a Mammalian Lectin. J. Biol. Chem. 2018, 293, 7265. [Google Scholar] [CrossRef] [Green Version]
- Cárcamo, C.; Pardo, E.; Oyanadel, C.; Bravo-Zehnder, M.; Bull, P.; Cáceres, M.; Martínez, J.; Massardo, L.; Jacobelli, S.; González, A.; et al. Galectin-8 Binds Specific Beta1 Integrins and Induces Polarized Spreading Highlighted by Asymmetric Lamellipodia in Jurkat T Cells. Exp. Cell Res. 2006, 312, 374–386. [Google Scholar] [CrossRef]
- Thurston, T.L.M.; Wandel, M.P.; von Muhlinen, N.; Foeglein, Á.; Randow, F. Galectin 8 Targets Damaged Vesicles for Autophagy to Defend Cells against Bacterial Invasion. Nature 2012, 482, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Staring, J.; von Castelmur, E.; Blomen, V.A.; van den Hengel, L.G.; Brockmann, M.; Baggen, J.; Thibaut, H.J.; Nieuwenhuis, J.; Janssen, H.; van Kuppeveld, F.J.M.; et al. PLA2G16 Represents a Switch between Entry and Clearance of Picornaviridae. Nature 2017, 541, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Levy, Y.; Auslender, S.; Eisenstein, M.; Vidavski, R.R.; Ronen, D.; Bershadsky, A.D.; Zick, Y. It Depends on the Hinge: A Structure-Functional Analysis of Galectin-8, a Tandem-Repeat Type Lectin. Glycobiology 2006, 16, 463–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reticker-Flynn, N.E.; Malta, D.F.B.; Winslow, M.M.; Lamar, J.M.; Xu, M.J.; Underhill, G.H.; Hynes, R.O.; Jacks, T.E.; Bhatia, S.N. A Combinatorial Extracellular Matrix Platform Identifies Cell-Extracellular Matrix Interactions That Correlate with Metastasis. Nat. Commun. 2012, 3, 1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, Y.; Kanzawa, T.; Sawaya, R.; Kondo, S. The Role of Autophagy in Cancer Development and Response to Therapy. Nat. Rev. Cancer 2005, 5, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Jandrey, E.H.F.; Bezerra, M.; Inoue, L.T.; Furnari, F.B.; Camargo, A.A.; Costa, É.T. A Key Pathway to Cancer Resilience: The Role of Autophagy in Glioblastomas. Front. Oncol. 2021, 11, 652133. [Google Scholar] [CrossRef] [PubMed]
- Metz, C.; Döger, R.; Riquelme, E.; Cortés, P.; Holmes, C.; Shaughnessy, R.; Oyanadel, C.; Grabowski, C.; González, A.; Soza, A. Galectin-8 Promotes Migration and Proliferation and Prevents Apoptosis in U87 Glioblastoma Cells. Biol. Res. 2016, 49, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shatz-Azoulay, H.; Vinik, Y.; Isaac, R.; Kohler, U.; Lev, S.; Zick, Y. The Animal Lectin Galectin-8 Promotes Cytokine Expression and Metastatic Tumor Growth in Mice. Sci. Rep. 2020, 10, 7375. [Google Scholar] [CrossRef] [PubMed]
- Portella, L.; Bello, A.M.; Scala, S. CXCL12 Signaling in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2021, 1302, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.L.; Guimaraes, J.C.; Auf der Maur, P.; De Silva, D.; Trefny, M.P.; Okamoto, R.; Bruno, S.; Schmidt, A.; Mertz, K.; Volkmann, K.; et al. Hepatic Stellate Cells Suppress NK Cell-Sustained Breast Cancer Dormancy. Nature 2021, 594, 566–571. [Google Scholar] [CrossRef]
- Chen, I.X.; Chauhan, V.P.; Posada, J.; Ng, M.R.; Wu, M.W.; Adstamongkonkul, P.; Huang, P.; Lindeman, N.; Langer, R.; Jain, R.K. Blocking CXCR4 Alleviates Desmoplasia, Increases T-Lymphocyte Infiltration, and Improves Immunotherapy in Metastatic Breast Cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 4558–4566. [Google Scholar] [CrossRef] [Green Version]
- Feig, C.; Jones, J.O.; Kraman, M.; Wells, R.J.B.; Deonarine, A.; Chan, D.S.; Connell, C.M.; Roberts, E.W.; Zhao, Q.; Caballero, O.L.; et al. Targeting CXCL12 from FAP-Expressing Carcinoma-Associated Fibroblasts Synergizes with Anti–PD-L1 Immunotherapy in Pancreatic Cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 20212–20217. [Google Scholar] [CrossRef] [Green Version]
- Tribulatti, M.V.; Mucci, J.; Cattaneo, V.; Agüero, F.; Gilmartin, T.; Head, S.R.; Campetella, O. Galectin-8 Induces Apoptosis in the CD4(High)CD8(High) Thymocyte Subpopulation. Glycobiology 2007, 17, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Norambuena, A.; Metz, C.; Vicuña, L.; Silva, A.; Pardo, E.; Oyanadel, C.; Massardo, L.; González, A.; Soza, A. Galectin-8 Induces Apoptosis in Jurkat T Cells by Phosphatidic Acid-Mediated ERK1/2 Activation Supported by Protein Kinase A down-Regulation. J. Biol. Chem. 2009, 284, 12670–12679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tribulatti, M.V.; Cattaneo, V.; Hellman, U.; Mucci, J.; Campetella, O. Galectin-8 Provides Costimulatory and Proliferative Signals to T Lymphocytes. J. Leukoc. Biol. 2009, 86, 371–380. [Google Scholar] [CrossRef]
- Prato, C.A.; Carabelli, J.; Campetella, O.; Tribulatti, M.V. Galectin-8 Enhances T Cell Response by Promotion of Antigen Internalization and Processing. iScience 2020, 23, 101278. [Google Scholar] [CrossRef] [PubMed]
- Tribulatti, M.V.; Figini, M.G.; Carabelli, J.; Cattaneo, V.; Campetella, O. Redundant and Antagonistic Functions of Galectin-1, -3, and -8 in the Elicitation of T Cell Responses. J. Immunol. Baltim. Md 1950 2012, 188, 2991–2999. [Google Scholar] [CrossRef]
- Türeci, O.; Schmitt, H.; Fadle, N.; Pfreundschuh, M.; Sahin, U. Molecular Definition of a Novel Human Galectin Which Is Immunogenic in Patients with Hodgkin’s Disease. J. Biol. Chem. 1997, 272, 6416–6422. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Nishi, N.; Shoji, H.; Seki, M.; Hashidate, T.; Hirabayashi, J.; Kasai Ki, K.; Hata, Y.; Suzuki, S.; Hirashima, M.; et al. Functional Analysis of the Carbohydrate Recognition Domains and a Linker Peptide of Galectin-9 as to Eosinophil Chemoattractant Activity. Glycobiology 2002, 12, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Nagae, M.; Nishi, N.; Murata, T.; Usui, T.; Nakamura, T.; Wakatsuki, S.; Kato, R. Crystal Structure of the Galectin-9 N-Terminal Carbohydrate Recognition Domain from Mus Musculus Reveals the Basic Mechanism of Carbohydrate Recognition. J. Biol. Chem. 2006, 281, 35884–35893. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Ming, H.; Wang, Y.; Yang, Y.; Yi, L.; Li, T.; Ma, H.; Tong, L.; Zhang, L.; Liu, P.; et al. Molecular and Clinical Characterization of Galectin-9 in Glioma through 1,027 Samples. J. Cell. Physiol. 2020, 235, 4326–4334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Anderson, A.C.; Schubart, A.; Xiong, H.; Imitola, J.; Khoury, S.J.; Zheng, X.X.; Strom, T.B.; Kuchroo, V.K. The Tim-3 Ligand Galectin-9 Negatively Regulates T Helper Type 1 Immunity. Nat. Immunol. 2005, 6, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Fourcade, J.; Sun, Z.; Benallaoua, M.; Guillaume, P.; Luescher, I.F.; Sander, C.; Kirkwood, J.M.; Kuchroo, V.; Zarour, H.M. Upregulation of Tim-3 and PD-1 Expression Is Associated with Tumor Antigen–Specific CD8+ T Cell Dysfunction in Melanoma Patients. J. Exp. Med. 2010, 207, 2175–2186. [Google Scholar] [CrossRef] [PubMed]
- Sakuishi, K.; Apetoh, L.; Sullivan, J.M.; Blazar, B.R.; Kuchroo, V.K.; Anderson, A.C. Targeting Tim-3 and PD-1 Pathways to Reverse T Cell Exhaustion and Restore Anti-Tumor Immunity. J. Exp. Med. 2010, 207, 2187–2194. [Google Scholar] [CrossRef] [PubMed]
- Wolf, Y.; Anderson, A.C.; Kuchroo, V.K. TIM3 Comes of Age as an Inhibitory Receptor. Nat. Rev. Immunol. 2020, 20, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Ohashi, P.S. Clinical Blockade of PD1 and LAG3—Potential Mechanisms of Action. Nat. Rev. Immunol. 2015, 15, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A Guide to Cancer Immunotherapy: From T Cell Basic Science to Clinical Practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The Blockade of Immune Checkpoints in Cancer Immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Sun, L.; Li, C.-F.; Wang, Y.-H.; Yao, J.; Li, H.; Yan, M.; Chang, W.-C.; Hsu, J.-M.; Cha, J.-H.; et al. Galectin-9 Interacts with PD-1 and TIM-3 to Regulate T Cell Death and Is a Target for Cancer Immunotherapy. Nat. Commun. 2021, 12, 832. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, A.M.; Rudkjøbing, S.J.; Sørensen, M.D.; Dahlrot, R.H.; Kristensen, B.W. Expression and Prognostic Value of the Immune Checkpoints Galectin-9 and PD-L1 in Glioblastomas. J. Neuropathol. Exp. Neurol. 2021, 80, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Han, H.; He, X.; Li, S.; Wu, C.; Yu, C.; Wang, S. Expression of the Galectin-9-Tim-3 Pathway in Glioma Tissues Is Associated with the Clinical Manifestations of Glioma. Oncol. Lett. 2016, 11, 1829–1834. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.; Wang, X.; Wang, F.; Feng, E.; You, G. Galectin-9: A Predictive Biomarker Negatively Regulating Immune Response in Glioma Patients. World Neurosurg. 2019, 132, e455–e462. [Google Scholar] [CrossRef]
- Lv, R.; Bao, Q.; Li, Y. Regulation of M1-type and M2-type Macrophage Polarization in RAW264.7 Cells by Galectin-9. Mol. Med. Rep. 2017, 16, 9111–9119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enninga, E.A.L.; Nevala, W.K.; Holtan, S.G.; Leontovich, A.A.; Markovic, S.N. Galectin-9 Modulates Immunity by Promoting Th2/M2 Differentiation and Impacts Survival in Patients with Metastatic Melanoma. Melanoma Res. 2016, 26, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, H.; Yu, X.; Collins, P.M.; Bum-Erdene, K. Galectin-3 Inhibitors: A Patent Review (2008-Present). Expert Opin. Ther. Pat. 2014, 24, 1053–1065. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, H.; Bum-Erdene, K.; Bohari, M.H.; Yu, X. Galectin-1 Inhibitors and Their Potential Therapeutic Applications: A Patent Review. Expert Opin. Ther. Pat. 2016, 26, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Natarajamurthy, S.H.; Sistla, S.; Dharmesh, S.M. Disruption of Galectin-3 and Galectin-3 Binding Protein (G3BP) Interaction by Dietary Pectic Polysaccharides (DPP)—Arrest of Metastasis, Inhibition of Proliferation and Induction of Apoptosis. Int. J. Biol. Macromol. 2019, 139, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Zhao, Z.; Lin, Z.; Geng, J.; Guan, Y.; Song, C.; Zhou, Y.; Tai, G. Selective Effects of Ginseng Pectins on Galectin-3-Mediated T Cell Activation and Apoptosis. Carbohydr. Polym. 2019, 219, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Téllez-Sanz, R.; García-Fuentes, L.; Vargas-Berenguel, A. Human Galectin-3 Selective and High Affinity Inhibitors. Present State and Future Perspectives. Curr. Med. Chem. 2013, 20, 2979–2990. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-C.; Lin, H.-Y.; Tu, Z.; Kuo, Y.-H.; Hsu, S.-T.D.; Lin, C.-H. Dissecting the Structure-Activity Relationship of Galectin-Ligand Interactions. Int. J. Mol. Sci. 2018, 19, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetterberg, F.R.; Peterson, K.; Johnsson, R.E.; Brimert, T.; Håkansson, M.; Logan, D.T.; Leffler, H.; Nilsson, U.J. Monosaccharide Derivatives with Low-Nanomolar Lectin Affinity and High Selectivity Based on Combined Fluorine-Amide, Phenyl-Arginine, Sulfur-π, and Halogen Bond Interactions. ChemMedChem 2018, 13, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Patel, B.; Kishor, C.; Houston, T.A.; Shatz-Azoulay, H.; Zick, Y.; Vinik, Y.; Blanchard, H. Rational Design and Synthesis of Methyl-β-d-Galactomalonyl Phenyl Esters as Potent Galectin-8N Antagonists. J. Med. Chem. 2020, 63, 11573–11584. [Google Scholar] [CrossRef]
- Sampson, J.H.; Gunn, M.D.; Fecci, P.E.; Ashley, D.M. Brain Immunology and Immunotherapy in Brain Tumours. Nat. Rev. Cancer 2020, 20, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Kipnis, J. Multifaceted Interactions between Adaptive Immunity and the Central Nervous System. Science 2016, 353, 766–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa de Olza, M.; Navarro Rodrigo, B.; Zimmermann, S.; Coukos, G. Turning up the Heat on Non-Immunoreactive Tumours: Opportunities for Clinical Development. Lancet Oncol. 2020, 21, e419–e430. [Google Scholar] [CrossRef]
- Aigner, A.; Kögel, D. Nanoparticle/SiRNA-Based Therapy Strategies in Glioma: Which Nanoparticles, Which SiRNAs? Nanomedicine 2018, 13, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F.; Messaoudi, K.; Lemaire, L.; Benoit, J.-P.; Lagarce, F. Combined Anti-Galectin-1 and Anti-EGFR SiRNA-Loaded Chitosan-Lipid Nanocapsules Decrease Temozolomide Resistance in Glioblastoma: In Vivo Evaluation. Int. J. Pharm. 2015, 481, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Van Woensel, M.; Wauthoz, N.; Rosière, R.; Mathieu, V.; Kiss, R.; Lefranc, F.; Steelant, B.; Dilissen, E.; Van Gool, S.W.; Mathivet, T.; et al. Development of SiRNA-Loaded Chitosan Nanoparticles Targeting Galectin-1 for the Treatment of Glioblastoma Multiforme via Intranasal Administration. J. Control. Release Off. J. Control. Release Soc. 2016, 227, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Sharanek, A.; Burban, A.; Hernandez-Corchado, A.; Madrigal, A.; Fatakdawala, I.; Najafabadi, H.S.; Soleimani, V.D.; Jahani-Asl, A. Transcriptional Control of Brain Tumor Stem Cells by a Carbohydrate Binding Protein. Cell Rep. 2021, 36, 109647. [Google Scholar] [CrossRef] [PubMed]
- Goett, H.; Kolodziej, M.; Pons-Kühnemann, J.; Schwarm, F.; Koch, C.; Uhl, E.; Stein, M. P11.38 The Galectin-1 Inhibitor OTX-008 Has Antiproliferative Effects on Human Glioblastoma Cells Independent of MGMT-Promotor Status. Neuro -Oncol. 2019, 21 (Suppl. S3), iii51. [Google Scholar] [CrossRef]
- Vladoiu, M.C.; Labrie, M.; St-Pierre, Y. Intracellular Galectins in Cancer Cells: Potential New Targets for Therapy (Review). Int. J. Oncol. 2014, 44, 1001–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fecci, P.E.; Sampson, J.H. The Current State of Immunotherapy for Gliomas: An Eye toward the Future. J. Neurosurg. 2019, 131, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Pothin, E.; Lesuisse, D.; Lafaye, P. Brain Delivery of Single-Domain Antibodies: A Focus on VHH and VNAR. Pharmaceutics 2020, 12, 937. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-López, E.; Schuhmacher, A.J. Transportation of Single-Domain Antibodies through the Blood–Brain Barrier. Biomolecules 2021, 11, 1131. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Bourgeois, J.-P.; Celli, S.; Glacial, F.; Sourd, A.-M.L.; Mecheri, S.; Weksler, B.; Romero, I.; Couraud, P.-O.; Rougeon, F.; et al. Cell-Penetrating Anti-GFAP VHH and Corresponding Fluorescent Fusion Protein VHH-GFP Spontaneously Cross the Blood-Brain Barrier and Specifically Recognize Astrocytes: Application to Brain Imaging. FASEB J. 2012, 26, 3969–3979. [Google Scholar] [CrossRef] [PubMed]
- Rutgers, K.S.; Nabuurs, R.J.A.; van den Berg, S.A.A.; Schenk, G.J.; Rotman, M.; Verrips, C.T.; van Duinen, S.G.; Maat-Schieman, M.L.; van Buchem, M.A.; de Boer, A.G.; et al. Transmigration of Beta Amyloid Specific Heavy Chain Antibody Fragments across the in Vitro Blood-Brain Barrier. Neuroscience 2011, 190, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Fatehi, D.; Baral, T.N.; Abulrob, A. In Vivo Imaging of Brain Cancer Using Epidermal Growth Factor Single Domain Antibody Bioconjugated to Near-Infrared Quantum Dots. J. Nanosci. Nanotechnol. 2014, 14, 5355–5362. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, K.; Iqbal, U.; Tanha, J.; MacKenzie, R.; Moreno, M.; Stanimirovic, D. Single-Domain Antibodies as Therapeutic and Imaging Agents for the Treatment of CNS Diseases. Antibodies 2019, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajarrag, S.; St-Pierre, Y. Galectins in Glioma: Current Roles in Cancer Progression and Future Directions for Improving Treatment. Cancers 2021, 13, 5533. https://doi.org/10.3390/cancers13215533
Ajarrag S, St-Pierre Y. Galectins in Glioma: Current Roles in Cancer Progression and Future Directions for Improving Treatment. Cancers. 2021; 13(21):5533. https://doi.org/10.3390/cancers13215533
Chicago/Turabian StyleAjarrag, Samy, and Yves St-Pierre. 2021. "Galectins in Glioma: Current Roles in Cancer Progression and Future Directions for Improving Treatment" Cancers 13, no. 21: 5533. https://doi.org/10.3390/cancers13215533
APA StyleAjarrag, S., & St-Pierre, Y. (2021). Galectins in Glioma: Current Roles in Cancer Progression and Future Directions for Improving Treatment. Cancers, 13(21), 5533. https://doi.org/10.3390/cancers13215533