
RhoGDI2-Mediated Rac1 Recruitment to Filamin A Enhances Rac1 Activity and Promotes Invasive Abilities of Gastric Cancer Cells

 ,
,  and
and 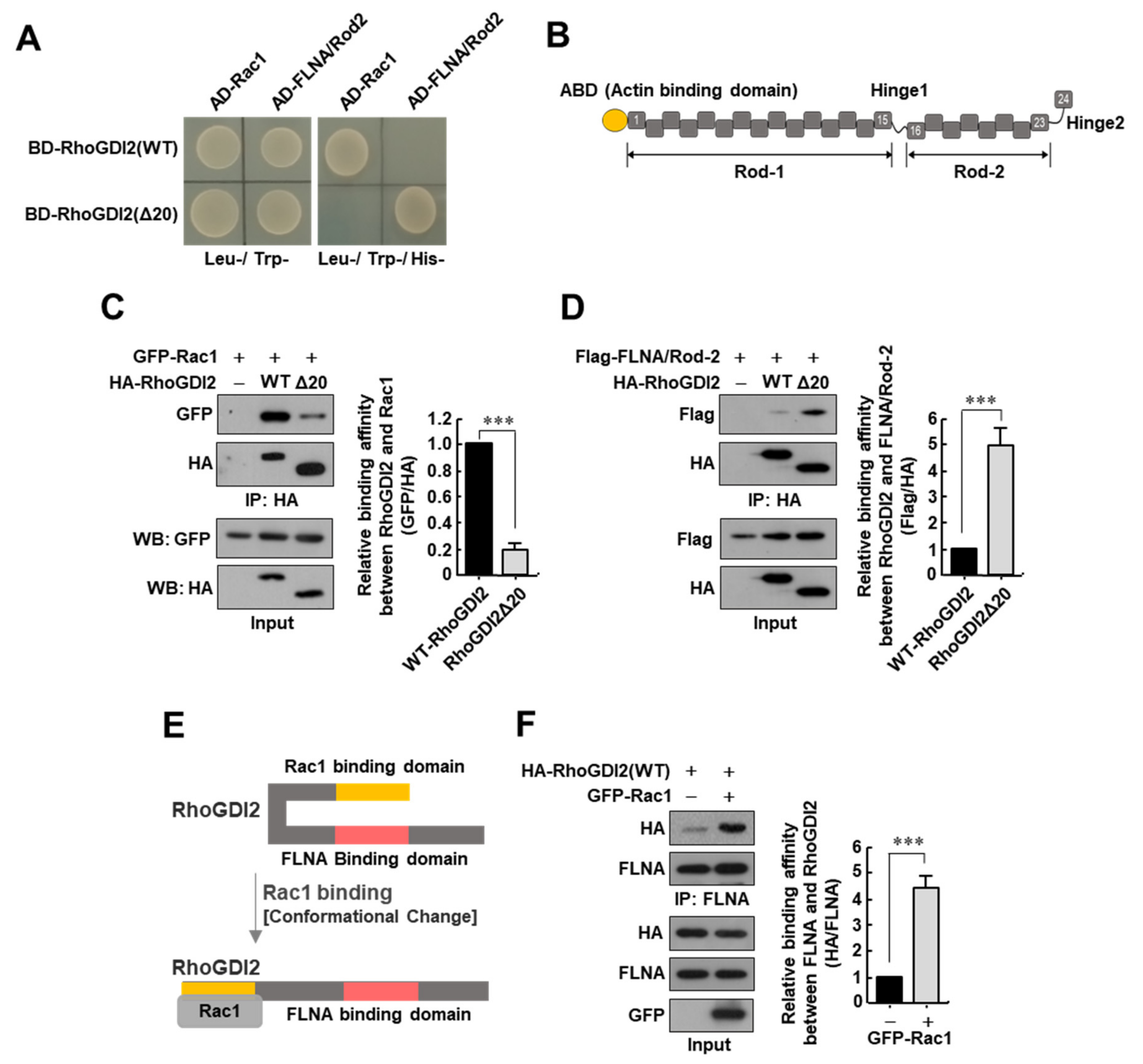{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Gene Cloning and Transfection
2.3. Antibodies and Western Blot Analysis
2.4. Yeast Two-Hybrid Screening
2.5. Immunoprecipitation
2.6. RNA Interference Experiments and Transfection
2.7. Rac Activity Assay
2.8. Migration Assay
2.9. Invasion Assay
2.10. Proliferation Assay
2.11. Statical Analysis
3. Results
3.1. Interaction between RhoGDI2 and Rac1 Is a Prerequisite for RhoGDI2 to Interact with Filamin A
3.2. RhoGDI2 Plays an Important Role in the Interaction between Rac1 and Filamin A and Rac1 Activation in Gastric Cancer Cells
3.3. Filamin A Is Required for Rac1 Activation and the Invasive Ability of Gastric Cancer Cells
3.4. Trio Is Critical for Rac1 Activation and Invasive Ability of Gastric Cancer Cells
3.5. Expression of RhoGDI2 and Filamin A could Be a Poor Prognostic Marker for Gastric Cancer Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jamal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duraes, C.; Almeida, G.M.; Seruca, R.; Oliveira, C.; Caeneiro, F. Biomarkers for gastric cancer: Prognostic, predictive or targets of therapy? Virchows Arch. 2014, 464, 367–378. [Google Scholar] [CrossRef]
- Schmidt, A.; Hall, A. Guanine nucleotide exchange factors for Rho GTPases: Turning on the switch. Genes Dev. 2002, 16, 1587–1609. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.Y.; Zheng, Y. Rho GTPase-activation proteins in cell regulation. Trends Cell Biol. 2003, 13, 13–22. [Google Scholar] [CrossRef]
- Fukomoto, Y.; Kaibuchi, K.; Hori, Y.; Fujioka, H.; Araki, S.; Ueda, T.; Kikuchi, A.; Takai, Y. Molecular cloning and characterization of a novel type of regulatory protein (GDI) for the Rho proteins, ras p21-like small GTP-binding proteins. Oncogene 1990, 5, 1321–1328. [Google Scholar]
- Scherle, P.; Behrens, T.; Staudt, L.M. Ly-GDI, a GDP-dissociation inhibitor of the RhoA GTP-binding protein, is expressed preferentially in lymphocytes. Proc. Natl. Acad. Sci. USA 1993, 90, 7568–7572. [Google Scholar] [CrossRef] [Green Version]
- Zalcman, G.; Closson, V.; Camonis, J.; Honore, N.; Rousseau-Merck, M.F.; Tavitian, A.; Olofsson, B. RhoGDI-3 is a new GDP dissociation inhibitor (GDI). Identification of a non-cytosolic GDI protein interacting with the small GTP-binding proteins RhoB and RhoG. J. Biol. Chem. 1996, 271, 30366–30374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dovas, A.; Couchman, J.R. RhoGDI: Multiple functions in the regulation of Rho family GTPase activities. Biochem. J. 2005, 390, 1–9. [Google Scholar] [CrossRef]
- DerMardirossian, C.; Bokoch, G.M. GDIs: Central regulatory molecules in Rho GTPase activation. Trends Cell Biol. 2005, 15, 356–363. [Google Scholar] [CrossRef]
- Hart, M.J.; Maru, Y.; Leonard, D.; Witte, O.N.; Evans, T.; Cerione, R.A. A GDP dissociation inhibitor that serves as a GTPase inhibitor for the Ras-like protein CDC42Hs. Science 1992, 258, 812–815. [Google Scholar] [CrossRef]
- Chuang, T.H.; Xu, X.; Knaus, U.G.; Hart, M.J.; Bokoch, G.M. GDP dissociation inhibitor prevents intrinsic and GTPase activating protein-stimulated GTP hydrolysis by the Rac GTP-binding protein. J. Biol. Chem. 1993, 268, 775–778. [Google Scholar] [CrossRef]
- Gildea, J.J.; Seraj, M.J.; Oxford, G.; Harding, M.A.; Hampton, G.M.; Moskaluk, C.A.; Frierson, H.F.; Conaway, M.R.; Theodorescu, D. RhoGDI2 is an invasion and metastasis suppressor gene in human cancer. Cancer Res. 2002, 62, 6418–6423. [Google Scholar]
- Theodorescu, D.; Sapinoso, L.M.; Conaway, M.R.; Oxford, G.; Hampton, G.M.; Frierson, H.F., Jr. Reduced expression of metastasis suppressor RhoGDI2 is associated with decreased survival for patients with bladder cancer. Clin. Cancer Res. 2004, 10, 3800–3806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapper, J.; Kettunen, E.; El-Rifai, W.; Seppälä, M.; Andersson, L.C.; Knuutila, S. Changes in gene expression during progression of ovarian carcinoma. Cancer Genet. Cytogenet. 2001, 128, 1–6. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, B. D4-GDI, a Rho GTPase regulator, promotes breast cancer cell invasiveness. Cancer Res. 2006, 66, 5592–5598. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Baek, K.E.; Yoo, J. RhoGDI2 as a therapeutic target in cancer. Expert Opin. Ther. Targets 2010, 14, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Baek, K.E.; Park, S.M.; Kim, I.K.; Choi, Y.L.; Cho, H.J.; Nam, I.K.; Hwang, E.M.; Park, J.Y.; Han, J.Y.; et al. RhoGDI2 expression is associated with tumor growth and malignant progression of gastric cancer. Clin. Cancer Res. 2009, 15, 2612–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.J.; Kim, I.K.; Park, S.M.; Baek, K.E.; Nam, I.K.; Park, S.H.; Ryu, K.J.; Choi, J.; Ryu, J.; Hong, S.C.; et al. VEGF-C mediates RhoGDI2-induced gastric cancer cell metastasis and cisplatin resistance. Int. J. Cancer 2014, 135, 1553–1563. [Google Scholar]
- Scheffzek, K.; Stephan, I.; Jensen, O.N.; Illenberger, D.; Gierschik, P. The Rac-RhoGDI complex and the structural basis for the regulation of Rho proteins by RhoGDI. Nat. Struct. Biol. 2000, 7, 122–126. [Google Scholar] [CrossRef]
- Vetter, I.R.; Wittinghofer, A. The guanine nucleotide-binding switch in three dimensions. Science 2000, 294, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Bellanger, J.M.; Astier, C.; Sardet, C.; Ohta, Y.; Stossel, T.P.; Debant, A. The Rac1- and RhoG-specific GEF domain of Trio targets filamin to remodel cytoskeletal actin. Nat. Cell Biol. 2000, 2, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Worthylake, D.K.; Rossman, K.L.; Sondek, J. Crystal structure of Rac1 in complex with the guanine nucleotide exchange region of Tiam1. Nature 2000, 408, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xu, G.; Sotnikova, A.; Szczepanowski, M.; Giefing, M.; Krause, K.; Krams, M.; Siebert, R.; Jin, J.; Klapper, W. Loss of expression of LyGDI (ARHGDIB), a rho GDP-dissociation inhibitor, in Hodgkin lymphoma. Br. J. Haematol. 2007, 139, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Seraj, M.J.; Harding, M.A.; Gildea, J.J.; Welch, D.R.; Theodorescu, D. The relationship of BRMS1 and RhoGDI2 gene expression to metastatic potential in lineage related human bladder cancer cell lines. Clin. Exp. Metastasis 2000, 18, 519–525. [Google Scholar] [CrossRef]
- Kishi, K.; Sasaki, T.; Kuroda, S.; Itoh, T.; Takai, Y. Regulation of cytoplasmic division of Xenopus embryo by rho p21 and its inhibitory GDP/GTP exchange protein (rho GDI). J. Cell Biol. 1993, 120, 1187–1195. [Google Scholar] [CrossRef] [Green Version]
- Bromberg, Y.; Shani, E.; Joseph, G.; Gorzalczany, Y.; Sperling, E.; Pick, E. The GDP bound form of the small G protein Rac1p21is a potent activator of the superoxide-forming NADPH oxidase of macrophages. J. Biol. Chem. 1994, 269, 7055–7058. [Google Scholar] [CrossRef]
- Lin, Q.; Fuji, R.N.; Yang, W.; Cerione, R.A. RhoGDI is required for Cdc42-mediated cellular transformation. Curr. Biol. 2003, 13, 1469–1479. [Google Scholar] [CrossRef] [Green Version]
- Parri, M.; Chiarugi, P. Rac and Rho GTPases in cancer cell motility control. Cell Commun. Signal. 2010, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Cavallin, L.E.; Yan, B.; Zhu, S.; Duran, E.M.; Wang, H.; Hale, L.P.; Dong, C.; Cesarman, E.; Mesri, E.A.; et al. Antitumorigenesis of antioxidants in a transgenic Rac1 model of Kaposi’s sarcoma. Proc. Natl. Acad. Sci. USA 2009, 106, 8683–8688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz-Moreno, V.; Gadea, G.; Ahn, J.; Paterson, H.; Marra, P.; Pinner, S.; Sahai, E.; Marshall, C.J. Rac activation and inactivation control plasticity of tumor cell movement. Cell 2008, 135, 510–523. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.X.; Hartwig, J.H.; Akyurek, L.M. Filamins in cell signaling, transcription and organ development. Trends Cell Biol. 2010, 20, 113–123. [Google Scholar] [CrossRef]
- Nakamura, F.; Stossel, T.P.; Hartwig, J.H. The filamins: Organizers of cell structure and function. Cell Adhes. Migr. 2011, 5, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Flier, A.; Sonnenberg, A. Structural and functional aspects of filamins. Biochim. Biophys. Acta (BBA)- Mol. Cell Res. 2001, 1538, 99–117. [Google Scholar] [CrossRef] [Green Version]
- Himmel, M.; Van Der Ven, P.F.; Stocklein, W.; Furst, D.O. The limits of promiscuity: Isoform-specific dimerization of filamins. Biochemistry 2003, 42, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.D.; Seok, S.H.; Im, H.; Kwon, A.R.; Lee, S.J.; Kim, H.R.; Cho, Y.; Park, D.; Lee, B.J. Crystal structure of the dimerization domain of human filamin A. Proteins 2009, 75, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Lian, G.; Lu, J.; Hu, J.; Zhang, J.; Cross, S.H.; Ferland, R.J.; Sheen, V.L. Filamin a regulates neural progenitor proliferation and cortical size through Wee1-dependent Cdk1 phosphorylation. J. Neurosci. 2012, 32, 7672–7684. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Walsh, C.A. The many faces of filamin: A versatile molecular scaffold for cell motility and signalling. Nat. Cell Biol. 2004, 6, 1034–1038. [Google Scholar] [CrossRef]
- Ohta, Y.; Suzuki, N.; Nakamura, S.; Hartwig, J.H.; Stossel, T.P. The small GTPase RalA targets filamin to induce filopodia. Proc. Natl. Acad. Sci. USA 1999, 96, 2122–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, G.S.; Grundl, M.; Allen, J.S., 3rd; Matte, M.L. R-Ras interacts with filamin a to maintain endothelial barrier function. J. Cell. Physiol. 2011, 226, 2287–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; He, Q.; Berk, B.A.; Hartwig, J.H.; Stossel, T.P.; Nakamura, F. An adventitious interaction of filamin A with RhoGDI2(Tyr153Glu). Biochem. Biophys. Res. Commun. 2016, 469, 659–664. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Ryu, K.-J.; Kim, M.; Kim, T.; Kim, S.-H.; Han, H.; Kim, H.; Hong, K.-S.; Song, C.Y.; Choi, Y.; et al. RhoGDI2-Mediated Rac1 Recruitment to Filamin A Enhances Rac1 Activity and Promotes Invasive Abilities of Gastric Cancer Cells. Cancers 2022, 14, 255. https://doi.org/10.3390/cancers14010255
Kim H-J, Ryu K-J, Kim M, Kim T, Kim S-H, Han H, Kim H, Hong K-S, Song CY, Choi Y, et al. RhoGDI2-Mediated Rac1 Recruitment to Filamin A Enhances Rac1 Activity and Promotes Invasive Abilities of Gastric Cancer Cells. Cancers. 2022; 14(1):255. https://doi.org/10.3390/cancers14010255
Chicago/Turabian StyleKim, Hyo-Jin, Ki-Jun Ryu, Minju Kim, Taeyoung Kim, Seon-Hee Kim, Hyeontak Han, Hyemin Kim, Keun-Seok Hong, Chae Yeong Song, Yeonga Choi, and et al. 2022. "RhoGDI2-Mediated Rac1 Recruitment to Filamin A Enhances Rac1 Activity and Promotes Invasive Abilities of Gastric Cancer Cells" Cancers 14, no. 1: 255. https://doi.org/10.3390/cancers14010255
APA StyleKim, H. -J., Ryu, K. -J., Kim, M., Kim, T., Kim, S. -H., Han, H., Kim, H., Hong, K. -S., Song, C. Y., Choi, Y., Hwangbo, C., Kim, K. D., & Yoo, J. (2022). RhoGDI2-Mediated Rac1 Recruitment to Filamin A Enhances Rac1 Activity and Promotes Invasive Abilities of Gastric Cancer Cells. Cancers, 14(1), 255. https://doi.org/10.3390/cancers14010255