
Polypeptide-GalNAc-Transferase-13 Shows Prognostic Impact in Breast Cancer


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Anti-GalNAc-T13 Monoclonal Antibody (MAb) Production
2.2. Analysis of MAbs Specificity by Surface Plasmon Resonance
2.3. Western Blot
2.4. Clinical Samples and Immunohistochemistry Staining
2.5. Cell Lines
2.6. Reverse Transcription-Polymerase Chain Reaction (RT-PCR and qRT-PCR)
2.7. Immunocytochemistry and Immunofluorescence
2.8. Statistics
3. Results and Discussion
3.1. Production of a MAb Specific for GalNAc-T13
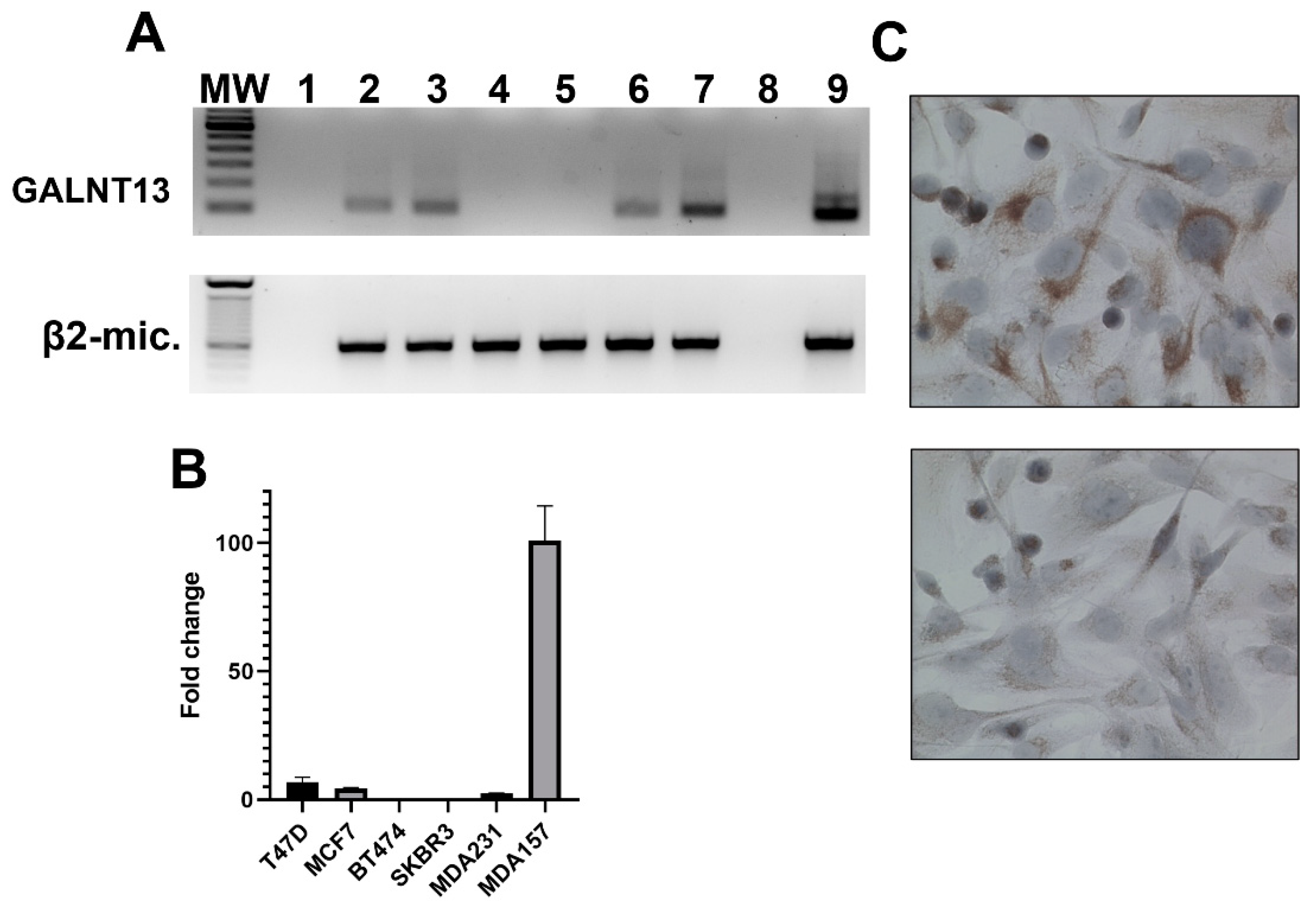
3.2. GalNAc-T13 Is Expressed in Breast Cancer
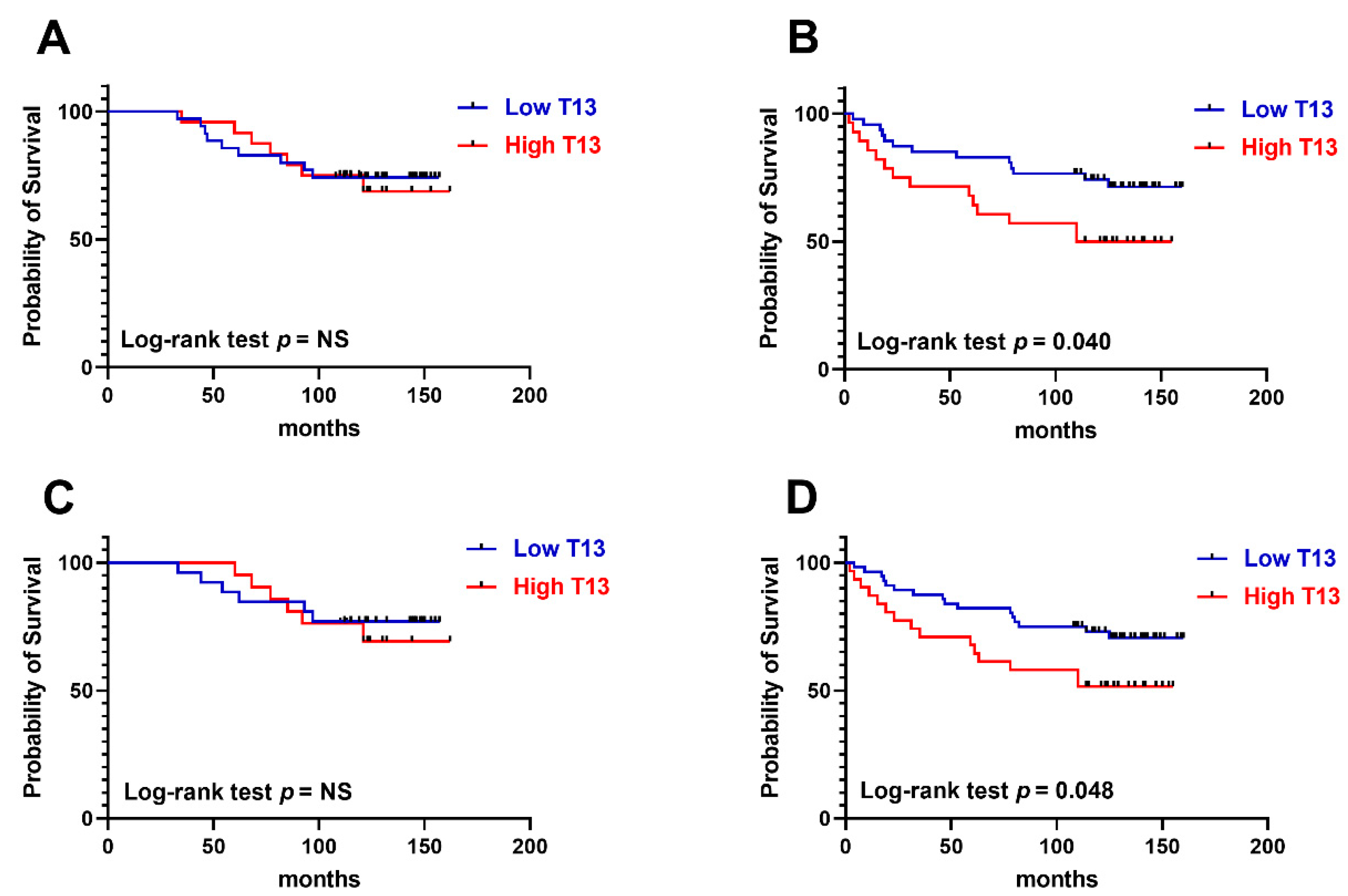
3.3. GalNAc-T13 Is an Aggressiveness Marker in Breast Cancer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Wang, S.; Ali, N. Advanced approaches to breast cancer classification and diagnosis. Front. Pharmacol. 2021, 11, 632079. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, A.; Pagliuca, F.; Zito Marino, F.; Accardo, M.; Cozzolino, I.; Franco, R. Current and potential immunohistochemical biomarkers for prognosis and therapeutic stratification of breast carcinoma. Semin. Cancer Biol. 2021, 72, 114–122. [Google Scholar] [CrossRef]
- Lüönd, F.; Tiede, S.; Christofori, G. Breast cancer as an example of tumour heterogeneity and tumour cell plasticity during malignant progression. Br. J. Cancer 2021, 125, 164–175. [Google Scholar] [CrossRef]
- Rodrigues, J.G.; Balmaña, M.; Macedo, J.A.; Poças, J.; Fernandes, Â.; de-Freitas-Junior, J.C.M.; Pinho, S.S.; Gomes, J.; Magalhães, A.; Gomes, C.; et al. Glycosylation in cancer: Selected roles in tumour progression, immune modulation and metastasis. Cell Immunol. 2018, 333, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.F.; Campos, D.; Reis, C.A.; Gomes, C. Targeting glycosylation: A new road for cancer drug discovery. Trends Cancer 2020, 6, 757–766. [Google Scholar] [CrossRef]
- Scott, E.; Elliott, D.J.; Munkley, J. Tumour associated glycans: A route to boost immunotherapy? Clin. Chim. Acta 2020, 502, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Kudelka, M.R.; Ju, T.; Heimburg-Molinaro, J.; Cummings, R.D. Simple sugars to complex disease-mucin-type O-glycans in cancer. Adv. Cancer Res. 2015, 126, 53–135. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.A.; Drake, R.R. Glycosylation and its implications in breast cancer. Expert Rev. Proteom. 2019, 16, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.R.; Hoessli, D.C.; Fang, M. N-acetylgalactosaminyltransferases in cancer. Oncotarget 2016, 7, 54067–54081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potapenko, I.O.; Lüders, T.; Russnes, H.G.; Helland, Å.; Sørlie, T.; Kristensen, V.N.; Nord, S.; Lingjærde, O.C.; Børresen-Dale, A.L.; Haakensen, V.D. Glycan-related gene expression signatures in breast cancer subtypes; relation to survival. Mol. Oncol. 2015, 9, 861–876. [Google Scholar] [CrossRef]
- Berois, N.; Mazal, D.; Ubillos, L.; Trajtenberg, F.; Nicolas, A.; Sastre-Garau, X.; Magdelenat, H.; Osinaga, E. UDP-N-acetyl-D-galactosamine: Polypeptide N-acetylgalactosaminyltransferase-6 as a new immunohistochemical breast cancer marker. J. Histochem. Cytochem. 2006, 54, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andergassen, U.; Liesche, F.; Kölbl, A.C.; Ilmer, M.; Hutter, S.; Friese, K.; Jeschke, U. Glycosyltransferases as markers for early tumorigenesis. BioMed Res. Int. 2015, 2015, 792672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liesche, F.; Kölbl, A.C.; Ilmer, M.; Hutter, S.; Jeschke, U.; Andergassen, U. Role of N-acetylgalactosaminyltransferase 6 in early tumorigenesis and formation of metastasis. Mol. Med. Rep. 2016, 13, 4309–4314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Nishidate, T.; Kijima, K.; Ohashi, T.; Takegawa, K.; Fujikane, T.; Hirata, K.; Nakamura, Y.; Katagiri, T. Critical roles of mucin 1 glycosylation by transactivated polypeptide N-acetylgalactosaminyltransferase 6 in mammary carcinogenesis. Cancer Res. 2010, 70, 2759–2769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Zhang, Y.; Fan, S.; Chen, L.; Tang, L.; Chen, X.; Lyu, J. GALNT6 promotes tumorigenicity and metastasis of breast cancer cell via beta-catenin/MUC1-C signaling pathway. Int. J. Biol. Sci. 2019, 15, 169–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Gallup, M.; Zlock, L.; Chen, Y.T.; Finkbeiner, W.E.; McNamara, N.A. Pivotal role of MUC1 glycosylation by cigarette smoke in modulating disruption of airway adherens junctions in vitro. J. Pathol. 2014, 234, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Yamada, S.; Wu, Y.; Wang, K.Y.; Liu, Y.P.; Uramoto, H.; Kohno, K.; Sasaguri, Y. Polypeptide N-acetylgalactosaminyltransferase-6 expression independently predicts poor overall survival in patients with lung adenocarcinoma after curative resection. Oncotarget 2016, 7, 54463–54473. [Google Scholar] [CrossRef] [Green Version]
- Freire, T.; Berois, N.; Sóñora, C.; Varangot, M.; Barrios, E.; Osinaga, E. UDP-N-acetyl-D-galactosamine:polypeptide N-acetylgalactosaminyltransferase 6 (ppGalNAc-T6) mRNA as a potential new marker for detection of bone marrow-disseminated breast cancer cells. Int. J. Cancer 2006, 119, 1383–1388. [Google Scholar] [CrossRef]
- Lin, W.R.; Yeh, C.T. GALNT14: An emerging marker capable of predicting therapeutic outcomes in multiple cancers. Int. J. Mol. Sci. 2020, 21, 1491. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Guo, X.; Wang, W.; Wang, Y.; Shan, Y.; Zhang, B.; Song, W.; Ma, S.; Ge, J.; Deng, H.; et al. N-Acetylgalactosaminyltransferase-14 as a potential biomarker for breast cancer by immunohistochemistry. BMC Cancer 2010, 10, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, K.H.; Park, M.S.; Nandu, T.S.; Gadad, S.; Kim, S.C.; Kim, M.Y. GALNT14 promotes lung-specific breast cancer metastasis by modulating self-renewal and interaction with the lung microenvironment. Nat. Commun. 2016, 7, 13796. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Iwasaki, H.; Wang, H.; Kudo, T.; Kalka, T.B.; Hennet, T.; Kubota, T.; Cheng, L.; Inaba, N.; Gotoh, M.; et al. Cloning and characterization of a new human UDP-N-acetyl-alpha-D-galactosamine:polypeptide N-acetylgalactosaminyltransferase, designated pp-GalNAc-T13, that is specifically expressed in neurons and synthesizes GalNAc alpha-serine/threonine antigen. J. Biol. Chem. 2003, 278, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Bennett, E.P.; Mandel, U.; Clausen, H.; Gerken, T.A.; Fritz, T.A.; Tabak, L.A. Control of mucin-type O-glycosylation: A classification of the polypeptide GalNAc-transferase gene family. Glycobiology 2012, 22, 736–756. [Google Scholar] [CrossRef] [Green Version]
- Berois, N.; Blanc, E.; Ripoche, H.; Mergui, X.; Trajtenberg, F.; Cantais, S.; Barrois, M.; Dessen, P.; Kågedal, B.; Bénard, J.; et al. ppGalNAc-T13: A new molecular marker of bone marrow involvement in neuroblastoma. Clin. Chem. 2006, 52, 1701–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, Y.; Zhang, Q.; Akita, K.; Nakada, H.; Hamamura, K.; Tokuda, N.; Tsuchida, A.; Matsubara, T.; Hori, T.; Okajima, T.; et al. pp-GalNAc-T13 induces high metastatic potential of murine Lewis lung cancer by generating trimeric Tn antigen. Biochem. Biophys. Res. Commun. 2012, 419, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Nogimori, K.; Hori, T.; Kawaguchi, K.; Fukui, T.; Mii, S.; Nakada, H.; Matsumoto, Y.; Yamauchi, Y.; Takahashi, M.; Furukawa, K.; et al. Increased expression levels of ppGalNAc-T13 in lung cancers: Significance in the prognostic diagnosis. Int. J. Oncol. 2016, 49, 1369–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festari, M.F.; Trajtenberg, F.; Berois, N.; Pantano, S.; Revoredo, L.; Kong, Y.; Solari-Saquieres, P.; Narimatsu, Y.; Freire, T.; Bay, S.; et al. Revisiting the human polypeptide GalNAc-T1 and T13 paralogs. Glycobiology 2017, 27, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Steentoft, C.; Yang, Z.; Wang, S.; Ju, T.; Vester-Christensen, M.B.; Festari, M.F.; King, S.L.; Moremen, K.; Larsen, I.S.B.; Goth, C.K.; et al. A validated collection of mouse monoclonal antibodies to human glycosyltransferases functioning in mucin-type O-glycosylation. Glycobiology 2019, 29, 645–656. [Google Scholar] [CrossRef]
- Gomes, J.; Marcos, N.T.; Berois, N.; Osinaga, E.; Magalhães, A.; Pinto-de-Sousa, J.; Almeida, R.; Gärtner, F.; Reis, C.A. Expression of UDP-N-acetyl-D-galactosamine: Polypeptide N-acetylgalactosaminyltransferase-6 in gastric mucosa, intestinal metaplasia, and gastric carcinoma. J. Histochem. Cytochem. 2009, 57, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ubillos, L.; Berriel, E.; Mazal, D.; Victoria, S.; Barrios, E.; Osinaga, E.; Berois, N. Polypeptide-GalNAc-T6 expression predicts better overall survival in patients with colon cancer. Oncol. Lett. 2018, 16, 225–234. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast cancer cell line classification and its relevance with breast tumor subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srour, M.K.; Qu, Y.; Deng, N.; Carlson, K.; Mirocha, J.; Gao, B.; Dadmanesh, F.; Cui, X.; Giuliano, A.E. Gene expression comparison between primary estrogen receptor-positive and triple-negative breast cancer with paired axillary lymph node metastasis. Breast J. 2021, 27, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.E.; Kim, N.I.; Lee, J.S.; Park, M.H.; Kang, K. Differentially expressed genes in matched normal, cancer, and lymph node metastases predict clinical outcomes in patients with breast cancer. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 111–122. [Google Scholar] [CrossRef]
- Lowe, J.B. Glycan-dependent leukocyte adhesion and recruitment in inflammation. Curr. Opin. Cell Biol. 2003, 15, 531–538. [Google Scholar] [CrossRef]
- Tenno, M.; Ohtsubo, K.; Hagen, F.K.; Ditto, D.; Zarbock, A.; Schaerli, P.; von Andrian, U.H.; Ley, K.; Le, D.; Tabak, L.A.; et al. Initiation of protein O glycosylation by the polypeptide GalNAcT-1 in vascular biology and humoral immunity. Mol. Cell Biol. 2007, 27, 8783–8796. [Google Scholar] [CrossRef] [Green Version]
- Chefetz, I.; Kohno, K.; Izumi, H.; Uitto, J.; Richard, G.; Sprecher, E. GALNT3, a gene associated with hyperphosphatemic familial tumoral calcinosis, is transcriptionally regulated by extracellular phosphate and modulates matrix metalloproteinase activity. Biochim. Biophys. Acta 2009, 1792, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Zhang, Q.; Akita, K.; Nakada, H.; Hamamura, K.; Tsuchida, A.; Okajima, T.; Furukawa, K.; Urano, T.; Furukawa, K. Trimeric Tn antigen on syndecan 1 produced by ppGalNAc-T13 enhances cancer metastasis via a complex formation with integrin α5β1 and matrix metalloproteinase 9. J. Biol. Chem. 2013, 288, 24264–24276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kind, S.; Jaretzke, A.; Büscheck, F.; Möller, K.; Dum, D.; Höflmayer, D.; Hinsch, A.; Weidemann, S.; Fraune, C.; Möller-Koop, C.; et al. A shift from membranous and stromal syndecan-1 (CD138) expression to cytoplasmic CD138 expression is associated with poor prognosis in breast cancer. Mol. Carcinog. 2019, 58, 2306–2315. [Google Scholar] [CrossRef]
- Sayyad, M.R.; Puchalapalli, M.; Vergara, N.G.; Wangensteen, S.M.; Moore, M.; Mu, L.; Edwards, C.; Anderson, A.; Kall, S.; Sullivan, M.; et al. Syndecan-1 facilitates breast cancer metastasis to the brain. Breast Cancer Res. Treat. 2019, 178, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Sobral-Leite, M.; Wesseling, J.; Smit, V.T.; Nevanlinna, H.; van Miltenburg, M.H.; Sanders, J.; Hofland, I.; Blows, F.M.; Coulson, P.; Patrycja, G.; et al. Annexin A1 expression in a pooled breast cancer series: Association with tumor subtypes and prognosis. BMC Med. 2015, 13, 156. [Google Scholar] [CrossRef] [Green Version]
- Silva-Oliveira, R.; Pereira, F.F.; Petronilho, S.; Martins, A.T.; Lameirinhas, A.; Constâncio, V.; Caldas-Ribeiro, I.; Salta, S.; Lopes, P.; Antunes, L.; et al. Clinical Significance of ARID1A and ANXA1 in HER-2 Positive Breast Cancer. J. Clin. Med. 2020, 9, 3911. [Google Scholar] [CrossRef]
- Bai, F.; Zhang, P.; Fu, Y.; Chen, H.; Zhang, M.; Huang, Q.; Li, D.; Li, B.; Wu, K. Targeting ANXA1 abrogates Treg-mediated immune suppression in triple-negative breast cancer. J. Immunother. Cancer 2020, 8, e000169. [Google Scholar] [CrossRef]
- Ren, L.; Yi, J.; Li, W.; Zheng, X.; Liu, J.; Wang, J.; Du, G. Apolipoproteins and cancer. Cancer Med. 2019, 8, 7032–7043. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Luo, G. Apolipoproteins, as the carrier proteins for lipids, are involved in the development of breast cancer. Clin. Transl. Oncol. 2020, 22, 1952–1962. [Google Scholar] [CrossRef]
- Flowers, S.A.; Grant, O.C.; Woods, R.J.; Rebeck, G.W. O-glycosylation on cerebrospinal fluid and plasma apolipoprotein E differs in the lipid-binding domain. Glycobiology 2020, 30, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Kacperczyk, M.; Kmieciak, A.; Kratz, E.M. The Role of ApoE expression and variability of its glycosylation in human reproductive health in the light of current information. Int. J. Mol. Sci. 2021, 22, 7197. [Google Scholar] [CrossRef]
- Liu, S.; Li, R.; Zuo, S.; Luo, R.; Fang, W.; Xie, Y. GRP94 overexpression as an indicator of unfavorable outcomes in breast cancer patients. Int. J. Clin. Exp. Pathol. 2018, 11, 3061–3067. [Google Scholar] [PubMed]
- Calderon, B.P.; Sennesael, A.L.; Glorieux, C. Glucose-regulated protein of 94 kDa contributes to the development of an aggressive phenotype in breast cancer cells. Biomed. Pharmacother. 2018, 105, 115–120. [Google Scholar] [CrossRef]
- Li, X.; Sun, L.; Hou, J.; Gui, M.; Ying, J.; Zhao, H.; Lv, N.; Meng, S. Cell membrane gp96 facilitates HER2 dimerization and serves as a novel target in breast cancer. Int. J. Cancer 2015, 137, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Cho, Y.B.; Lee, S. Cell surface GRP94 as a novel emerging therapeutic target for monoclonal antibody cancer therapy. Cells 2021, 10, 670. [Google Scholar] [CrossRef]
- Ma, C.; Liu, D.; Li, D.; Zhang, J.; Xu, X.Q.; Zhu, H.; Wan, X.F.; Miao, C.H.; Konkle, B.A.; Onigman, P.; et al. Comprehensive N- and O-glycosylation mapping of human coagulation factor V. J. Thromb. Haemost. 2020, 18, 1884–1892. [Google Scholar] [CrossRef]
- Tinholt, M.; Garred, Ø.; Borgen, E.; Beraki, E.; Schlichting, E.; Kristensen, V.; Sahlberg, K.K.; Iversen, N. Subtype-specific clinical and prognostic relevance of tumor-expressed F5 and regulatory F5 variants in breast cancer: The CoCaV study. J. Thromb. Haemost. 2018, 16, 1347–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinholt, M.; Stavik, B.; Tekpli, X.; Garred, Ø.; Borgen, E.; Kristensen, V.; Sahlberg, K.K.; Sandset, P.M.; Iversen, N. Coagulation factor V is a marker of tumor-infiltrating immune cells in breast cancer. Oncoimmunology 2020, 9, 1824644. [Google Scholar] [CrossRef]
- Corda, G.; Sala, G.; Lattanzio, R.; Iezzi, M.; Sallese, M.; Fragassi, G.; Lamolinara, A.; Mirza, H.; Barcaroli, D.; Ermler, S.; et al. Functional and prognostic significance of the genomic amplification of frizzled 6 (FZD6) in breast cancer. J. Pathol. 2017, 241, 350–361. [Google Scholar] [CrossRef]
- Zeng, C.M.; Chen, Z.; Fu, L. Frizzled receptors as potential therapeutic targets in human cancers. Int. J. Mol. Sci. 2018, 19, 1543. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, T.M.; Thomas, L.N.; Gupta, A.; Barnes, P.J.; Too, C.K. Prolactin and androgen R1881 induce pro-survival carboxypeptidase-D and EDD E3 ligase in triple-negative and HER2+ breast cancer. Am. J. Cancer Res. 2020, 10, 1321–1343. [Google Scholar]
- Goettig, P. Effects of glycosylation on the enzymatic activity and mechanisms of proteases. Int. J. Mol. Sci. 2016, 17, 1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijazi, M.M.; Young, P.E.; Dougherty, M.K.; Bressette, D.S.; Cao, T.T.; Pierce, J.H.; Wong, L.M.; Alimandi, M.; King, C.R. NRG-3 in human breast cancers: Activation of multiple erbB family proteins. Int. J. Oncol. 1998, 13, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Hayes, N.V.; Gullick, W.J. The neuregulin family of genes and their multiple splice variants in breast cancer. J. Mammary Gland. Biol. Neoplasia 2008, 13, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Ocaña, A.; Díez-González, L.; Esparís-Ogando, A.; Montero, J.C.; Amir, E.; Pandiella, A. Neuregulin expression in solid tumors: Prognostic value and predictive role to anti-HER3 therapies. Oncotarget 2016, 7, 45042–45051. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | Low GalNAc-T13 1 | High GalNAc-T13 | ||
|---|---|---|---|---|
| n | n (%) | n (%) | p2 | |
| Total | 338 | 190 (56.2) | 148 (43.8) | |
| Mean age 51.3 (range: 27–83) | ||||
| Histological type | NS 3 | |||
| Invasive ductal carcinoma | 322 | 178 (55.3) | 144 (44.7) | |
| Others | 16 | 12 (75) | 4 (25) | |
| Histological Grade | NS | |||
| 1 | 17 | 12 (70.6) | 5 (29.4) | |
| 2 | 211 | 105 (49.8) | 106 (50.2) | |
| 3 | 51 | 31 (60.8) | 20 (39.2) | |
| T | NS | |||
| T1 | 46 | 32 (69.6) | 14 (30.4) | |
| T2 | 226 | 125 (55.3) | 101 (44.7) | |
| T3 | 43 | 19 (44.2) | 24 (55.8) | |
| T4 | 20 | 11 (55) | 9 (45) | |
| N | NS | |||
| N0 | 153 | 86 (56.2) | 67 (43.8) | |
| N+ | 182 | 101 (55.5) | 81 (44.5) | |
| Hormonal Receptors | ||||
| ER 4 (-) | 139 | 75 (54) | 64 (46) | NS |
| (+) | 189 | 112 (59.3) | 77 (40.7) | |
| PR 5 (-) | 182 | 97 (53.3) | 85 (46.7) | NS |
| (+) | 144 | 89 (61.8) | 55 (38.2) | |
| Her2 (-) | 230 | 142 (61.7) | 88 (38.3) | 0.002 |
| (+) | 87 | 37 (42.5) | 50 (57.5) | |
| Molecular types | 0.022 | |||
| Luminal A | 106 | 64 (60.4) | 42 (39.6) | |
| Luminal B | 92 | 55 (59.8) | 37 (40.2) | |
| Her2 | 41 | 14 (34) | 27 (66) | |
| TNBC 6 | 78 | 46 (59) | 32 (41) | |
| Stage | NS | |||
| I | 25 | 15 (60) | 10 (40) | |
| II | 209 | 121 (57.9) | 88 (42.1) | |
| III | 101 | 51 (50.5) | 50 (49.5)17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, E.; Ubillos, L.; Elgul, N.; Festari, M.F.; Mazal, D.; Pritsch, O.; Alonso, I.; Osinaga, E.; Berois, N. Polypeptide-GalNAc-Transferase-13 Shows Prognostic Impact in Breast Cancer. Cancers 2021, 13, 5616. https://doi.org/10.3390/cancers13225616
Fernandez E, Ubillos L, Elgul N, Festari MF, Mazal D, Pritsch O, Alonso I, Osinaga E, Berois N. Polypeptide-GalNAc-Transferase-13 Shows Prognostic Impact in Breast Cancer. Cancers. 2021; 13(22):5616. https://doi.org/10.3390/cancers13225616
Chicago/Turabian StyleFernandez, Eugenia, Luis Ubillos, Nabila Elgul, María Florencia Festari, Daniel Mazal, Otto Pritsch, Isabel Alonso, Eduardo Osinaga, and Nora Berois. 2021. "Polypeptide-GalNAc-Transferase-13 Shows Prognostic Impact in Breast Cancer" Cancers 13, no. 22: 5616. https://doi.org/10.3390/cancers13225616
APA StyleFernandez, E., Ubillos, L., Elgul, N., Festari, M. F., Mazal, D., Pritsch, O., Alonso, I., Osinaga, E., & Berois, N. (2021). Polypeptide-GalNAc-Transferase-13 Shows Prognostic Impact in Breast Cancer. Cancers, 13(22), 5616. https://doi.org/10.3390/cancers13225616