
General Features and Novel Gene Signatures That Identify Epstein-Barr Virus-Associated Epithelial Cancers

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. DNA Microarray
2.3. Collection and Processing of Publicly Available Data
2.4. Data Analysis
2.5. Verification of Candidate Genes Expression in Cell Lines and Tumor Tissues
2.6. Statistical Analysis
3. Results
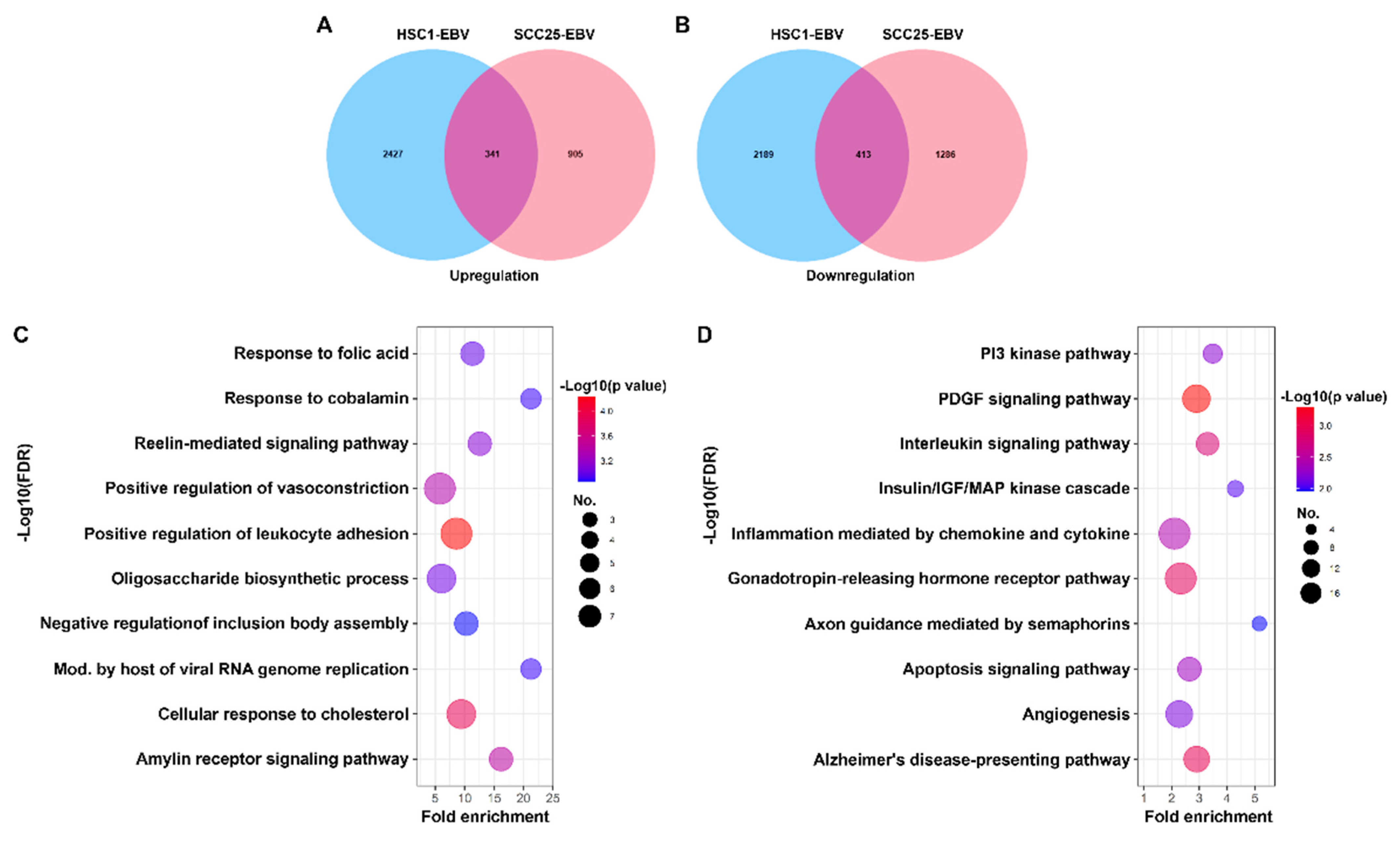
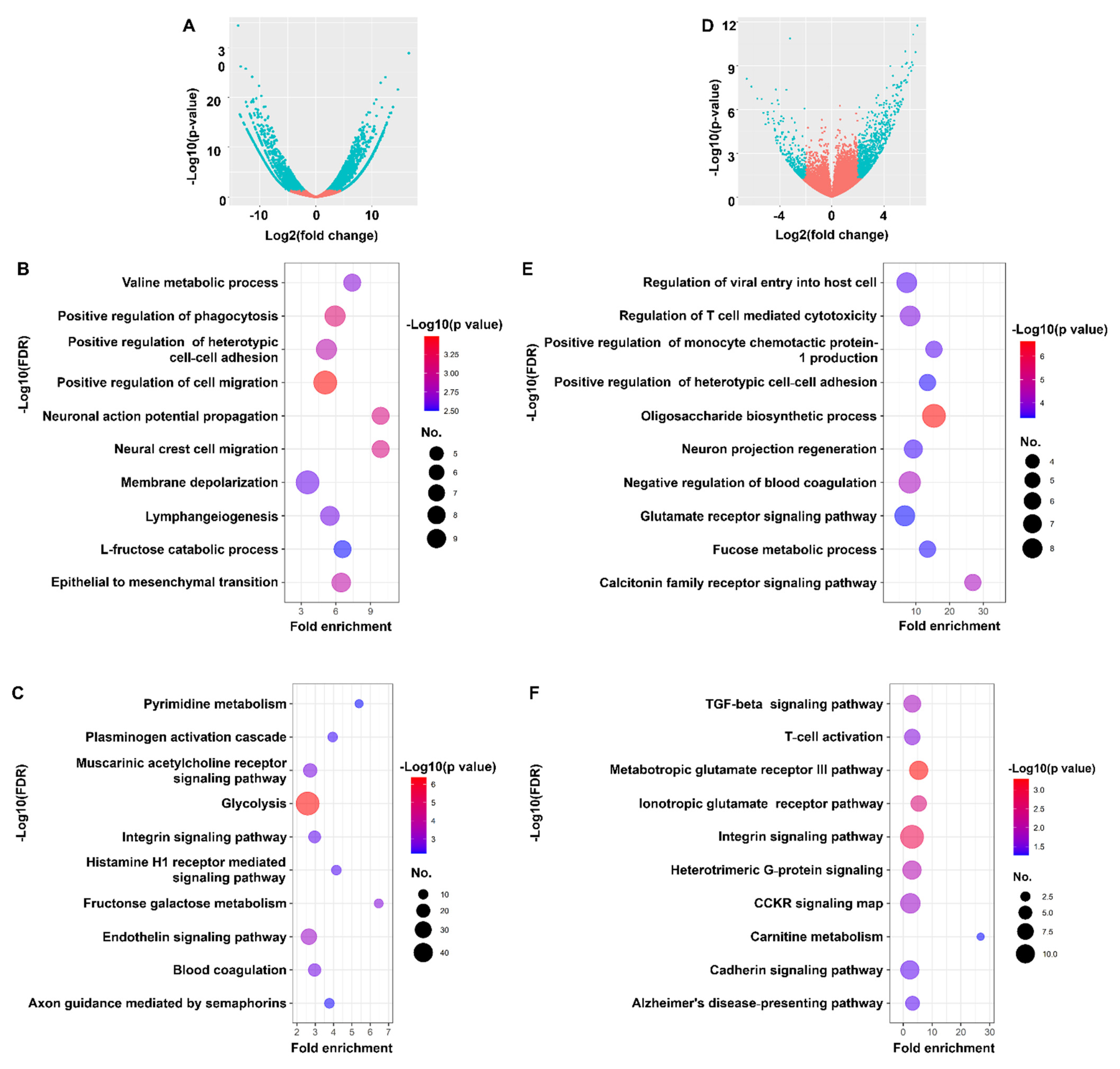
3.1. Identification of Differentially Expressed Genes in EBVaCAs
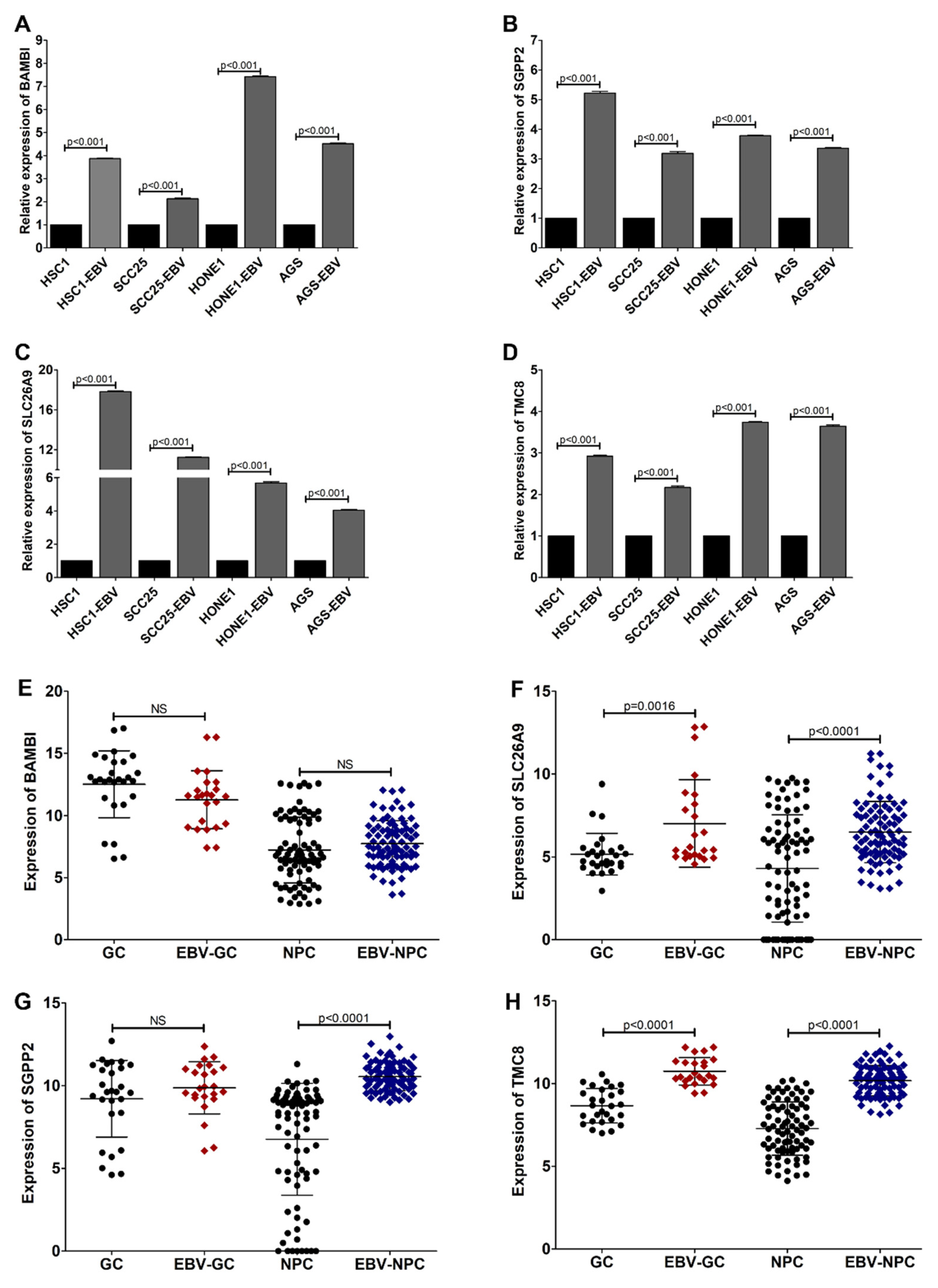
3.2. SLC26A9 and TMC8 as Promising Gene Signatures for EBVaCAs
3.3. TMC8 Is a Potential Gene Signature for EBV-Associated Malignancies
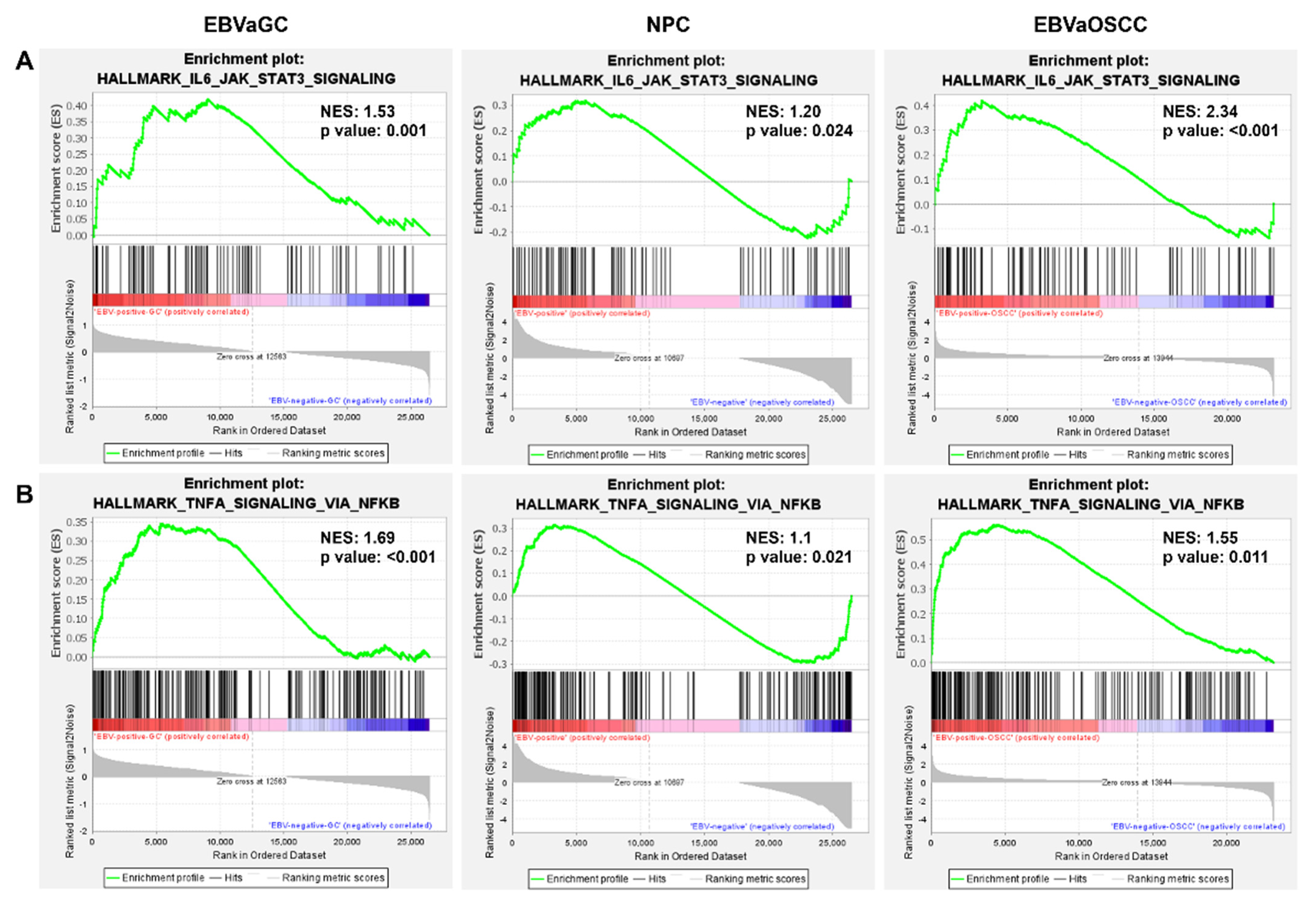
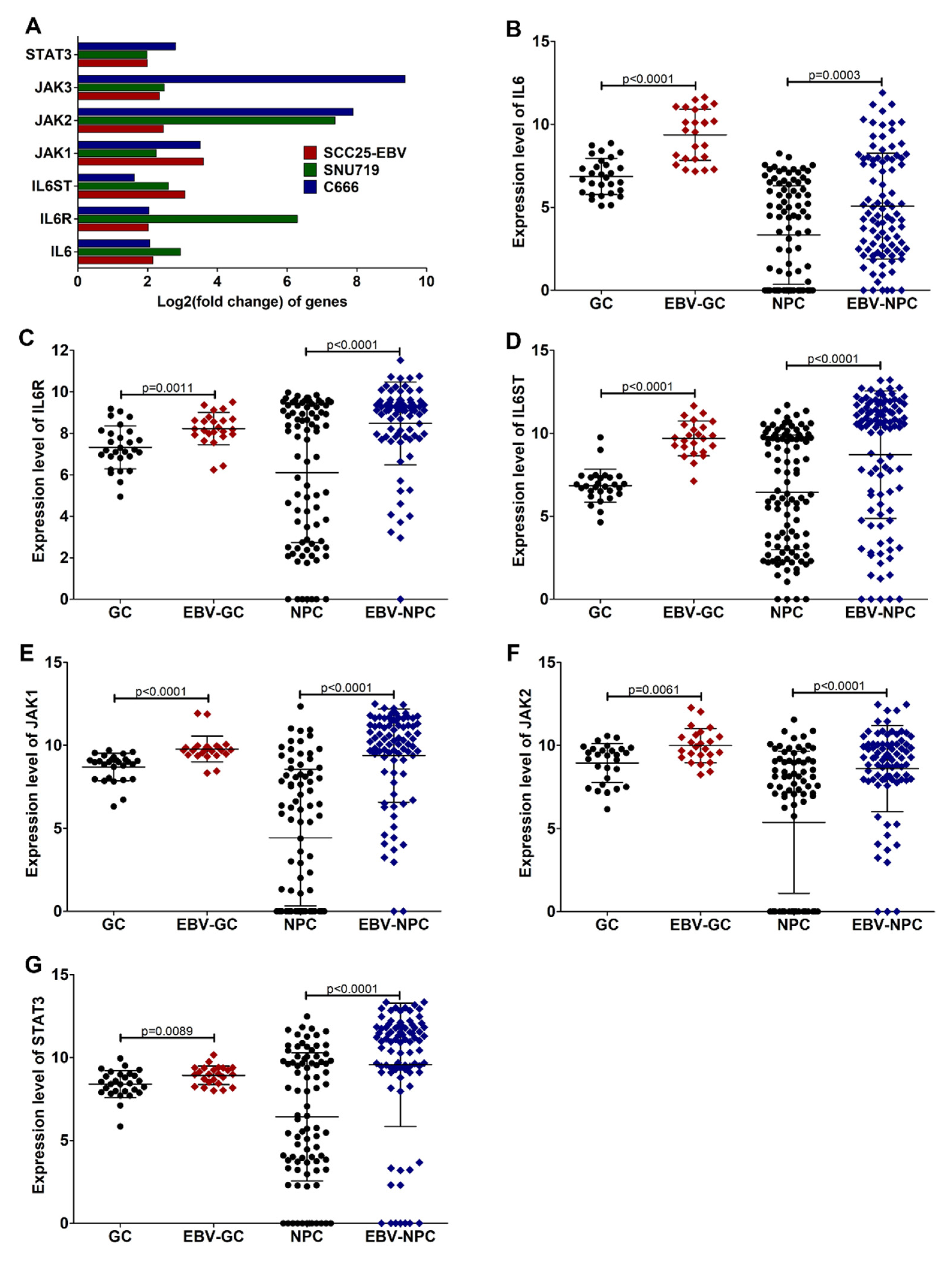
3.4. EBV Induces IL6/JAK/STAT3 and TNF-α/NF-KB Signaling in EBVaCAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsao, S.W.; Tsang, C.M.; To, K.F.; Lo, K.W. The role of Epstein–Barr virus in epithelial malignancies. J. Pathol. 2015, 235, 323–333. [Google Scholar] [CrossRef]
- Thompson, M.P.; Kurzrock, R. Epstein-Barr virus and cancer. Clin. Cancer Res. 2004, 10, 803–821. [Google Scholar] [CrossRef] [Green Version]
- Young, L.S.; Rickinson, A.B. Epstein–Barr virus: 40 years on. Nat. Rev. Cancer 2004, 4, 757–768. [Google Scholar] [CrossRef]
- Yen, C.Y.; Lu, M.C.; Tzeng, C.C.; Huang, J.Y.; Chang, H.W.; Chen, R.S.; Liu, S.Y.; Liu, S.T.; Shieh, B.; Li, C. Detection of EBV infection and gene expression in oral cancer from patients in Taiwan by microarray analysis. J. Biomed. Biotechnol. 2009, 2009, 904589. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Noguchi, Y.; de Rivera, N.; Hoshino, M.; Sakashita, H.; Yamada, T.; Inoue, H.; Miyazaki, Y.; Nozaki, T.; González-López, B.S.; et al. Detection of Epstein-Barr virus genome and latent infection gene expression in normal epithelia, epithelial dysplasia, and squamous cell carcinoma of the oral cavity. Tumor Biol. 2016, 37, 3389–3404. [Google Scholar] [CrossRef]
- She, Y.; Nong, X.; Zhang, M.; Wang, M. Epstein-Barr virus infection and oral squamous cell carcinoma risk: A meta-analysis. PLoS ONE 2017, 12, e0186860. [Google Scholar] [CrossRef] [Green Version]
- Heawchaiyaphum, C.; Iizasa, H.; Ekalaksananan, T.; Burassakarn, A.; Kiyono, T.; Kanehiro, Y.; Yoshiyama, H.; Pientong, C. Epstein–Barr virus infection of oral squamous cells. Microorganisms 2020, 8, 419. [Google Scholar] [CrossRef] [Green Version]
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein–Barr virus infection and nasopharyngeal carcinoma. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160270. [Google Scholar] [CrossRef] [PubMed]
- Warnakulasuriya, S. Global epidemiology of oral and oropharyngeal cancer. Oral Oncol. 2009, 45, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Hajj, N.; Sittler, S.; Lammert, N.; Barnes, R.; Meloni-Ehrig, A. Gastric cancer: Classification, histology and application of molecular pathology. J. Gastrointest. Oncol. 2012, 3, 251–261. [Google Scholar]
- Han, B.L.; Xu, X.Y.; Zhang, C.Z.; Wu, J.J.; Han, C.F.; Wang, H.; Wang, X.; Wang, G.S.; Yang, S.J.; Xie, Y. Systematic review on Epstein-Barr virus (EBV) DNA in diagnosis of nasopharyngeal carcinoma in Asian populations. Asian Pac. J. Cancer Prev. 2012, 13, 2577–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, H.; Huang, Y.; Zhao, S.; Zhou, Y.; Lv, W. Prognostic value of Epstein–Barr virus DNA level for nasopharyngeal carcinoma: A meta-analysis of 8128 cases. Eur. Arch. Otorhinolaryngol. 2020, 277, 9–18. [Google Scholar] [CrossRef]
- Lo, Y.D.; Chan, W.Y.; Ng, E.K.; Chan, L.Y.; Lai, P.B.; Tam, J.S.; Chung, S.C. Circulating Epstein-Barr virus DNA in the serum of patients with gastric carcinoma. Clin. Cancer Res. 2001, 7, 1856–1859. [Google Scholar]
- Su, X.; Ye, Z.; Wang, Z.; Long, Y.; Qiu, M.; He, C. Epstein-Barr virus infection associated with pepsinogens and helicobacter pylori infection in patients with gastric cancer. Virus Res. 2018, 256, 1–5. [Google Scholar] [CrossRef]
- To, K.Y. Identification of differential gene expression by high throughput analysis. Comb. Chem. High Throughput Screen. 2000, 3, 235–241. [Google Scholar] [CrossRef]
- Roessler, S.; Jia, H.L.; Budhu, A.; Forgues, M.; Ye, Q.H.; Lee, J.S.; Thorgeirsson, S.S.; Sun, Z.; Tang, Z.Y.; Qin, L.Q.; et al. A unique metastasis gene signature enables prediction of tumor relapse in early-stage hepatocellular carcinoma patients. Cancer Res. 2010, 70, 10202–10212. [Google Scholar] [CrossRef] [Green Version]
- Heemers, H.V.; Schmidt, L.J.; Sun, Z.; Regan, K.M.; Anderson, S.K.; Duncan, K.; Wang, D.; Liu, S.; Ballman, K.V.; Tindall, D.J. Identification of a clinically relevant androgen-dependent gene signature in prostate cancer. Cancer Res. 2011, 71, 1978–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, F.; Qin, X.; Wang, B.; Li, Q.; Hu, J.; Cheng, X.; Guo, D.; Cheng, F.; Fang, C.; Tan, Y.; et al. ALKBH5 facilitates hypoxia-induced paraspeckle assembly and IL8 secretion to generate an immunosuppressive tumor microenvironment. Cancer Res. 2021, 81, 5876–5888. [Google Scholar] [CrossRef]
- Park, S.; Rong, L.; Owczarek, T.B.; Bernardo, M.D.; Shoulson, R.L.; Chua, C.W.; Kim, J.Y.; Lankarani, A.; Chakrapani, P.; Syed, T.; et al. Novel mouse models of bladder cancer identify a prognostic signature associated with risk of disease progression. Cancer Res. 2021, 81, 5161–5175. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Kui-Sheng Yang, K.S.; Chen, J.L.; Wang, Z.B. Identification and validation of an immune-associated RNA-binding proteins signature to predict clinical outcomes and therapeutic responses in colon cancer patients. World J. Surg. Oncol. 2021, 19, 314. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Song, M.; Li, W.; Li, M.; Chen, S.; Chen, B. Identification, verification and pathway enrichment analysis of prognosis-related immune genes in patients with hepatocellular carcinoma. Front. Oncol. 2021, 11, 695001. [Google Scholar] [CrossRef]
- Zhang, D.; Li, Y.; Yang, S.; Wang, M.; Yao, J.; Zheng, Y.; Deng, Y.; Li, N.; Wei, B.; Wu, Y.; et al. Identification of a glycolysis-related gene signature for survival prediction of ovarian cancer patients. Cancer Med. 2021, 10, 8222–8237. [Google Scholar] [CrossRef] [PubMed]
- Di Chen, D.; Wang, M.; Xu, Y.; Jiang, X.; Xiong, L.; Zhang, L.; Yu, H.; Xiong, Z. A novel autophagy-related lncRNA prognostic signature associated with immune microenvironment and survival outcomes of gastric cancer patients. Int. J. Gen. Med. 2021, 14, 6935–6950. [Google Scholar] [CrossRef]
- Li, D.; Lin, X.; Chen, B.; Ma, Z.; Zeng, Y.; Wang, H. Identification and validation of EMT-related lncRNA prognostic signature for colorectal cancer. Front. Genet. 2021, 12, 723802. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, H.; Lu, L. Identification and validation of hypoxia-related lncRNA signature as a prognostic model for hepatocellular carcinoma. Front. Genet. 2021, 12, 744113. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Park, C.; Kim, H.J.; Park, J.; Hwang, J.; Kim, J.I.; Choi, M.G.; Kim, S.; Kim, K.M.; Kang, M.S. Deregulation of immune response genes in patients with Epstein-Barr virus-associated gastric cancer and outcomes. Gastroenterology 2015, 148, 137–147.e9. [Google Scholar] [CrossRef] [PubMed]
- Szeto, C.Y.; Lin, C.H.; Choi, S.C.; Yip, T.T.; Ngan, R.K.; Tsao, G.S. Integrated mRNA and microRNA transcriptome sequencing characterizes sequence variants and mRNA-microRNA regulatory network in nasopharyngeal carcinoma model systems. FEBS Open Bio 2014, 4, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; MacIsaac, K.D.; Zhou, T.; Huang, P.Y.; Xin, C.; Dobson, J.R.; Yu, K.; Chiang, D.Y.; Fan, Y.; Pelletier, M.; et al. Genomic Analysis of Nasopharyngeal Carcinoma Reveals TME-Based Subtypes. Mol. Cancer Res. 2017, 15, 1722–1732. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Koneva, L.A.; Virani, S.; Arthur, A.E.; Virani, A.; Hall, P.B.; Warden, C.D.; Carey, T.E.; Chepeha, D.B.; Prince, M.E.; et al. Subtypes of HPV-Positive Head and Neck Cancers Are Associated with HPV Characteristics, Copy Number Alterations, PIK3CA Mutation, and Pathway Signatures. Clin. Cancer Res. 2016, 22, 4735–4745. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Zong, J.; Lin, W.; Wang, M.; Xu, Y.; Zhou, R.; Lin, S.; Guo, Q.; Chen, H.; Ye, Y.; et al. EBV-miR-BART8-3p induces epithelial-mesenchymal transition and promotes metastasis of nasopharyngeal carcinoma cells through activating NF-κB and Erk1/2 pathways. J. Exp. Clin. Cancer Res. 2018, 37, 283. [Google Scholar] [CrossRef]
- Kheir, F.; Zhao, M.; Strong, M.J.; Yu, Y.; Nanbo, A.; Flemington, E.K.; Morris, G.F.; Reiss, K.; Li, L.; Lin, Z. Detection of Epstein-Barr virus infection in non-small cell lung cancer. Cancers 2019, 11, 759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, C.W.; Port, R.J.; Young, L.S. The role of the EBV-encoded latent membrane proteins LMP1 and LMP2 in the pathogenesis of nasopharyngeal carcinoma (NPC). Semin. Cancer Biol. 2012, 22, 144–153. [Google Scholar] [CrossRef]
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein–Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef]
- Pan, X.; Liu, J.H. Identification of four key biomarkers and small molecule drugs in nasopharyngeal carcinoma by weighted gene co-expression network analysis. Bioengineered 2021, 12, 3647–3661. [Google Scholar] [CrossRef]
- Chen, T.; Yang, C.; Dou, R.; Xiong, B. Identification of a novel 10 immune-related genes signature as a prognostic biomarker panel for gastric cancer. Cancer Med. 2021, 10, 6546–6560. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, X.; Wang, N.; Ma, L.; Xie, X.; Zhang, H.; Kang, H.; Zhou, Z. Identification of novel antioxidant gene signature to predict the prognosis of patients with gastric cancer. World J. Surg. Oncol. 2021, 19, 219. [Google Scholar] [CrossRef]
- Lu, X.Q.; Zhang, J.Q.; Zhang, S.X.; Qiao, J.; Qiu, M.T.; Liu, X.R.; Chen, X.X.; Gao, C.; Zhang, H.H. Identification of novel hub genes associated with gastric cancer using integrated bioinformatics analysis. BMC Cancer 2021, 21, 697. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Zhu, D.; Zhang, H.; Zhang, S.; Zhou, X.; He, X.; Zhu, J.; Zhu, W. MicroRNA expression profiling analysis in serum for nasopharyngeal carcinoma diagnosis. Gene 2020, 727, 144243. [Google Scholar] [CrossRef]
- Zhang, H.; Zou, X.; Wu, L.; Zhang, S.; Wang, T.; Liu, P.; Zhu, W.; Zhu, J. Identification of a 7-microRNA signature in plasma as promising biomarker for nasopharyngeal carcinoma detection. Cancer Med. 2020, 9, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Liu, W.; Lu, Y.; Luo, H. A five-gene signature for predicting overall survival of esophagus adenocarcinoma. Medicine 2021, 100, e25305. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Z.; Shi, Q.; Wang, K.; Zhang, Z.; Liu, Z.; Cui, X.; Li, F. The TP53-related signature predicts immune cell infiltration, therapeutic response and prognosis in patients with oesophageal carcinoma. Front. Genet. 2021, 12, 607238. [Google Scholar] [CrossRef] [PubMed]
- El Khouri, E.; Touré, A. Functional interaction of the cystic fibrosis transmembrane conductance regulator with members of the SLC26 family of anion transporters (SLC26A8 and SLC26A9): Physiological and pathophysiological relevance. Int. J. Biochem. Cell Biol. 2014, 52, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Balázs, A.; Mall, M.A. Role of the SLC26A9 chloride channel as disease modifier and potential therapeutic target in cystic fibrosis. Front. Pharmacol. 2018, 9, 1112. [Google Scholar] [CrossRef]
- Alper, S.L.; Sharma, A.K. The SLC26 gene family of anion transporters and channels. Mol. Aspects Med. 2013, 34, 494–515. [Google Scholar] [CrossRef] [Green Version]
- Kurima, K.; Yang, Y.; Sorber, K.; Griffith, A.J. Characterization of the transmembrane channel-like (TMC) gene family: Functional clues from hearing loss and epidermodysplasia verruciformis. Genomics 2003, 82, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.S.; Stokes, A.J. The transmembrane channel-like protein family and human papillomaviruses: Insights into epidermodysplasia verruciformis and progression to squamous cell carcinoma. Oncoimmunology 2014, 3, e28288. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Wang, S.; Yao, Y.; Shen, Y.; Yang, H. Comprehensive co-expression analysis reveals TMC8 as a prognostic immune-associated gene in head and neck squamous cancer. Oncol. Lett. 2021, 22, 498. [Google Scholar] [CrossRef]
- Guo, W.; Chen, X.; Zhu, L.; Wang, Q. A six-mRNA signature model for the prognosis of head and neck squamous cell carcinoma. Oncotarget 2017, 8, 94528–94538. [Google Scholar] [CrossRef] [Green Version]
- Frecha, C.; Chevalier, S.A.; van Uden, P.; Rubio, I.; Siouda, M.; Saidj, D.; Cohen, C.; Lomonte, P.; Accardi, R.; Tommasino, M. Expression of the epidermodysplasia verruciformis-associated genes EVER1 and EVER2 is activated by exogenous DNA and inhibited by LMP1 oncoprotein from Epstein-Barr virus. J. Virol. 2015, 89, 1461–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef]
- Lui, V.W.Y.; Wong, E.Y.L.; Ho, Y.; Hong, B.; Wong, S.C.C.; Tao, Q.; Choi, G.C.; Au, T.C.; Ho, K.; Yau, D.M.; et al. STAT3 activation contributes directly to Epstein-Barr virus–mediated invasiveness of nasopharyngeal cancer cells in vitro. Int. J. Cancer 2009, 125, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Tung, K.L.; Wu, Y.T.; Liu, C.; Lin, S.C.; Wu, C.H.; Wu, S.Y.; Chang, Y.; Lan, Y.Y. EBV rta-induced IL-6 promotes migration of bystander tumor cells through IL-6R/JAK/STAT3 pathway in vitro. Anticancer Res. 2020, 40, 3255–3264. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal. Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.D.; Zhou, B. TNF-α/NF-κ B/Snail pathway in cancer cell migration and invasion. Br. J. Cancer 2010, 102, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF family cytokines. Semin Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Pozniak, P.D.; White, M.K.; Khalili, K. TNF-α/NF-κB signaling in the CNS: Possible connection to EPHB2. J. Neuroimmune Pharmacol. 2014, 9, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luftig, M.; Yasui, T.; Soni, V.; Kang, M.S.; Jacobson, N.; Cahir-McFarland, E.; Seed, B.; Kieff, E. Epstein–Barr virus latent infection membrane protein 1 TRAF-binding site induces NIK/IKKα-dependent noncanonical NF-κB activation. Proc. Natl. Acad. Sci. USA 2004, 101, 141–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takada, H.; Imadome, K.I.; Shibayama, H.; Yoshimori, M.; Wang, L.; Saitoh, Y.; Uota, S.; Yamaoka, S.; Koyama, T.; Shimizu, N.; et al. EBV induces persistent NF-κB activation and contributes to survival of EBV-positive neoplastic T-or NK-cells. PLoS ONE 2017, 12, e0174136. [Google Scholar]
- Kieser, A. Pursuing different ‘TRADDes’: TRADD signaling induced by TNF-receptor 1 and the Epstein-Barr virus oncoprotein LMP1. Biol. Chem. 2008, 389, 1261–1271. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| BAMBI | CCAAGGGAGCTGGAATTGAGT | ACTGTCTCACGTTTCCCAGTTA |
| GAPDH | TCATCAGCAATGCCTCCTGCA | TGGGTAGCAGTGATGGCA |
| SGPP2 | ATACGGTCCTGGATGTGCTG | ATGACACACACGGGGAAGAG |
| SLC26A9 | TTGCAAAAACCTCCCCCACA | TCTTGTGCATGTAGCGAGCA |
| TMC8 | AGGAGTCGTCTGAGAAGGGG | GAGAGAATCCTGCTGCGGTC |
| Hallmark | NES | NOM p Value | FDR q Value |
|---|---|---|---|
| EBVaOSCC | |||
| HALLMARK_TNFA_SIGNALING_VIA_NFKB | 2.34 | <0.001 | <0.001 |
| HALLMARK_HYPOXIA | 1.85 | <0.001 | 0.002 |
| HALLMARK_UV_RESPONSE_UP | 1.81 | <0.001 | 0.003 |
| HALLMARK_P53_PATHWAY | 1.73 | <0.001 | 0.009 |
| HALLMARK_REACTIVE_OXYGEN_SPECIES_PATHWAY | 1.7 | 0.01 | 0.008 |
| NPC | |||
| HALLMARK_NOTCH_SIGNALING | 1.27 | 0.12 | 0.615 |
| HALLMARK_KRAS_SIGNALING_DN | 1.22 | 0.02 | 0.451 |
| HALLMARK_TNFA_SIGNALING_VIA_NFKB | 1.2 | 0.024 | 0.37 |
| HALLMARK_UV_RESPONSE_UP | 1.19 | 0.037 | 0.299 |
| HALLMARK_IL6_JAK_STAT3_SIGNALING | 1.1 | 0.021 | 0.485 |
| EBVaGC | |||
| HALLMARK_INTERFERON_ALPHA_RESPONSE | 2.57 | <0.001 | <0.001 |
| HALLMARK_INTERFERON_GAMMA_RESPONSE | 2.33 | <0.001 | <0.001 |
| HALLMARK_XENOBIOTIC_METABOLISM | 1.79 | <0.001 | 0.003 |
| HALLMARK_INFLAMMATORY_RESPONSE | 1.76 | <0.001 | 0.004 |
| HALLMARK_IL6_JAK_STAT3_SIGNALING | 1.69 | <0.001 | 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heawchaiyaphum, C.; Pientong, C.; Yoshiyama, H.; Iizasa, H.; Panthong, W.; Ekalaksananan, T. General Features and Novel Gene Signatures That Identify Epstein-Barr Virus-Associated Epithelial Cancers. Cancers 2022, 14, 31. https://doi.org/10.3390/cancers14010031
Heawchaiyaphum C, Pientong C, Yoshiyama H, Iizasa H, Panthong W, Ekalaksananan T. General Features and Novel Gene Signatures That Identify Epstein-Barr Virus-Associated Epithelial Cancers. Cancers. 2022; 14(1):31. https://doi.org/10.3390/cancers14010031
Chicago/Turabian StyleHeawchaiyaphum, Chukkris, Chamsai Pientong, Hironori Yoshiyama, Hisashi Iizasa, Watcharapong Panthong, and Tipaya Ekalaksananan. 2022. "General Features and Novel Gene Signatures That Identify Epstein-Barr Virus-Associated Epithelial Cancers" Cancers 14, no. 1: 31. https://doi.org/10.3390/cancers14010031
APA StyleHeawchaiyaphum, C., Pientong, C., Yoshiyama, H., Iizasa, H., Panthong, W., & Ekalaksananan, T. (2022). General Features and Novel Gene Signatures That Identify Epstein-Barr Virus-Associated Epithelial Cancers. Cancers, 14(1), 31. https://doi.org/10.3390/cancers14010031