
CK2-Mediated Phosphorylation Upregulates the Stability of USP13 and Promotes Ovarian Cancer Cell Proliferation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
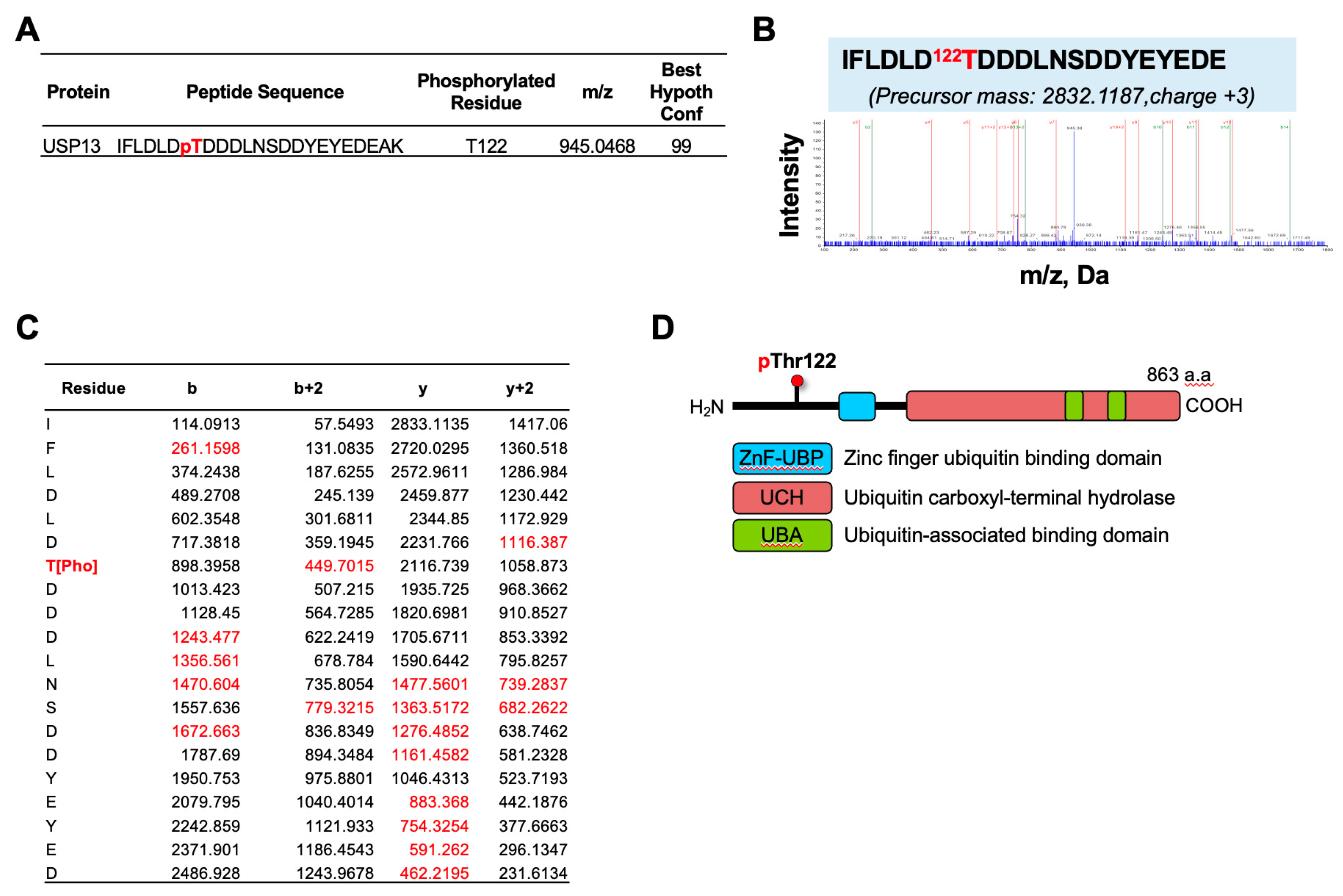
2.1. Identification of USP13 Phosphorylation in Ovarian Cancer Cells by LC-MS-MS Analysis
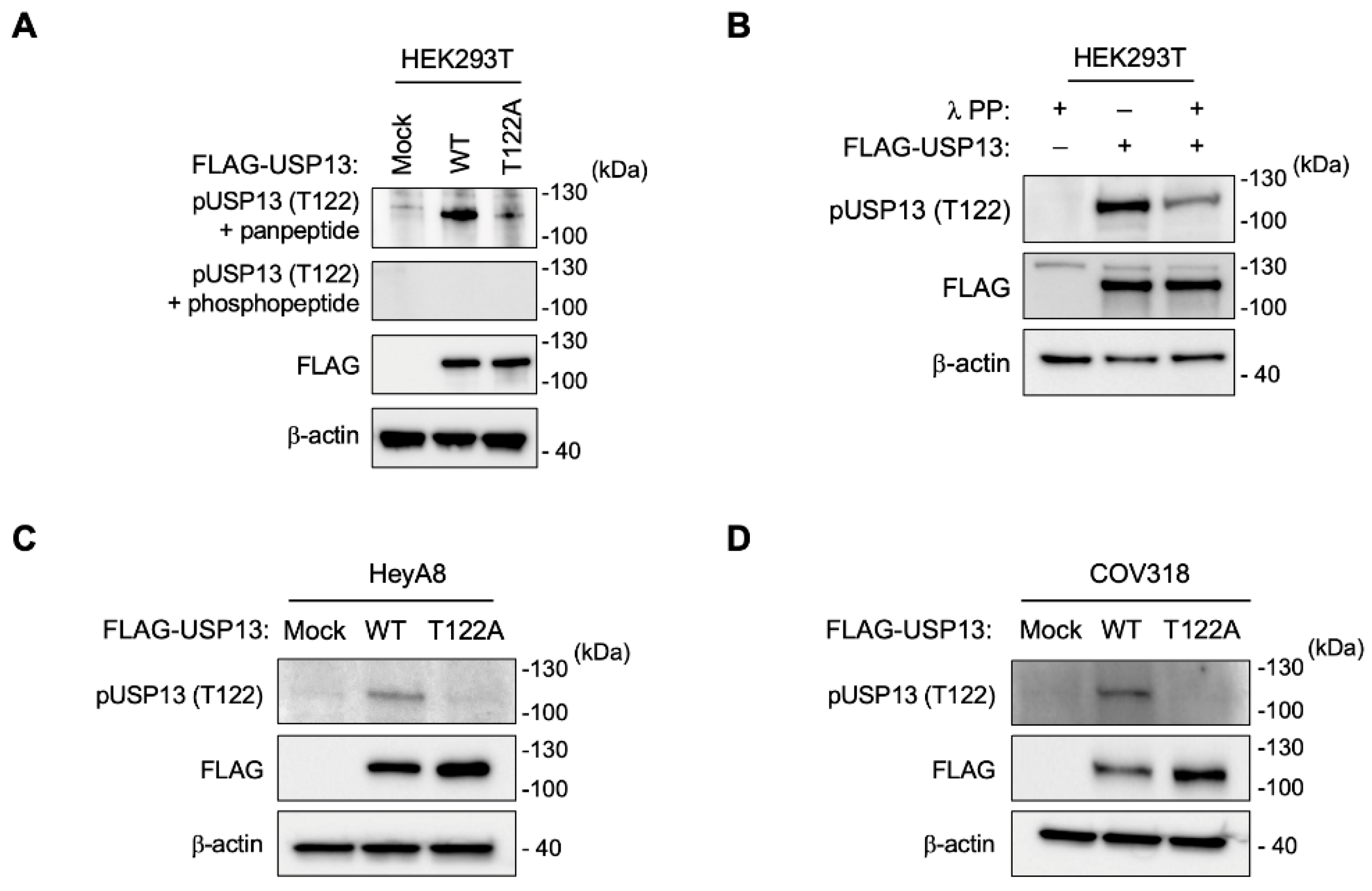
2.2. Generation of a Thr122 Phospho-Specific USP13 Antibody
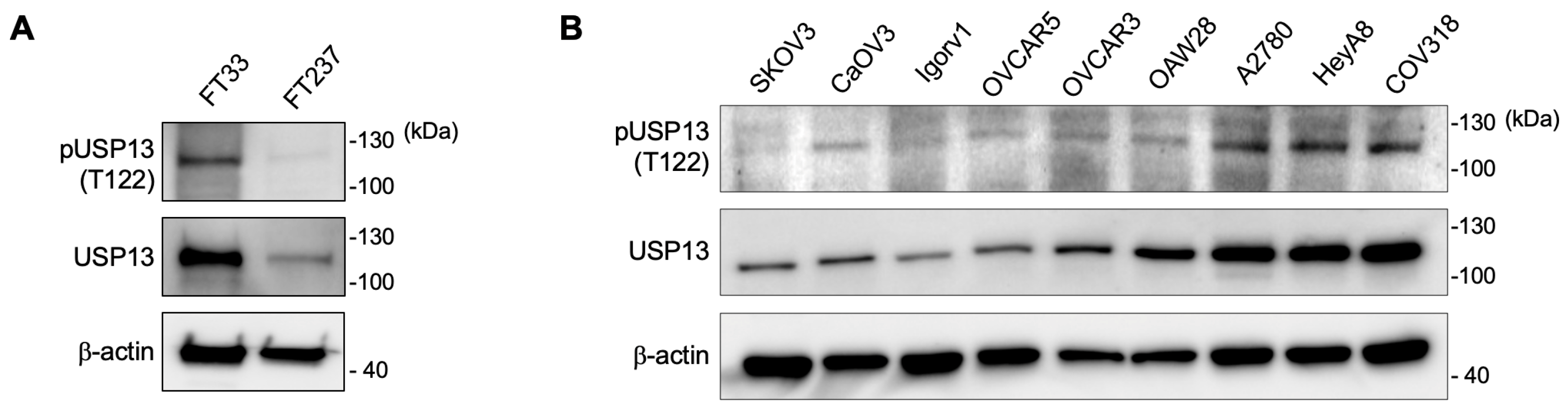
2.3. Phosphorylation of USP13 on Thr122 in Ovarian Cancer Cells
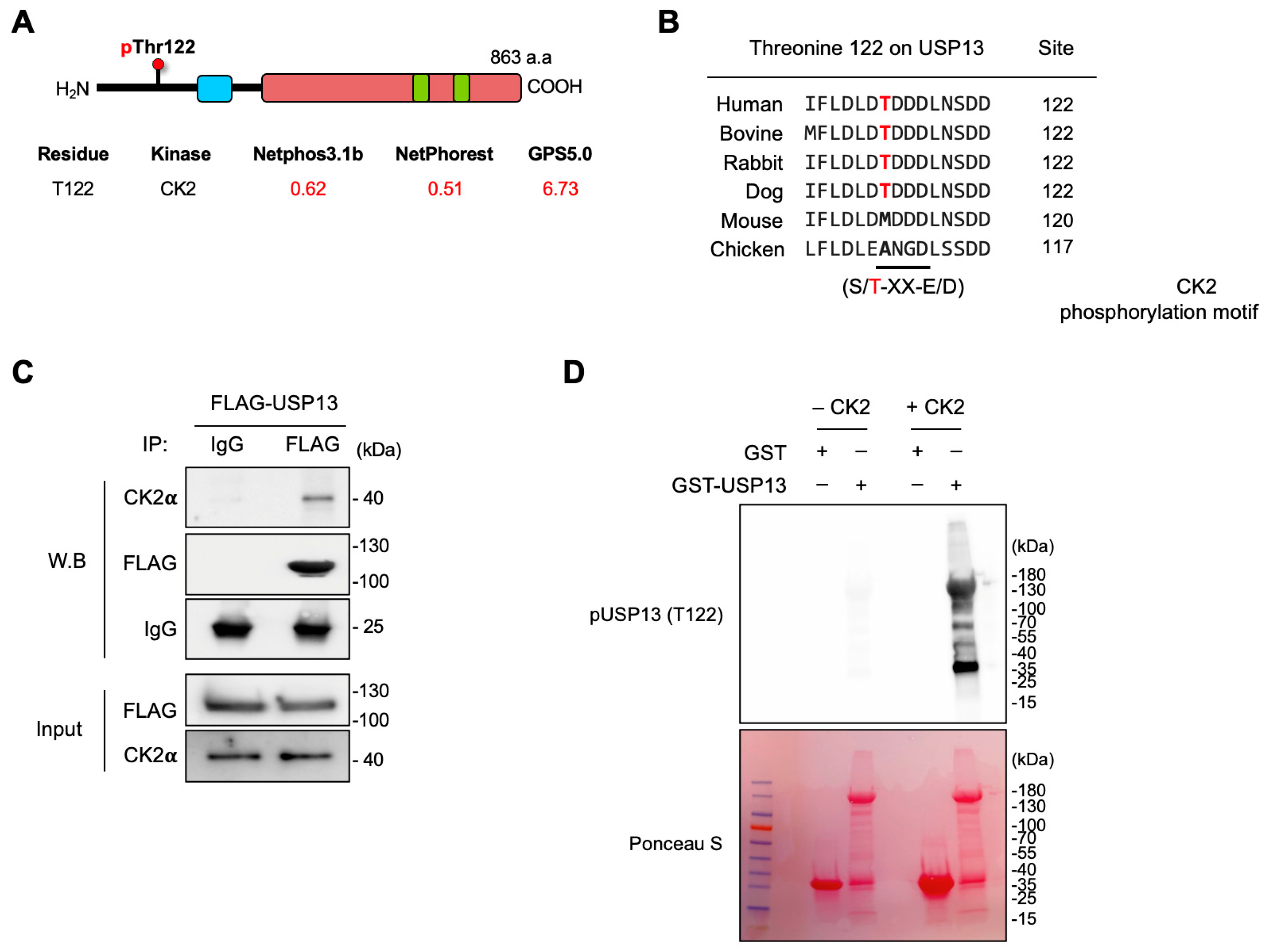
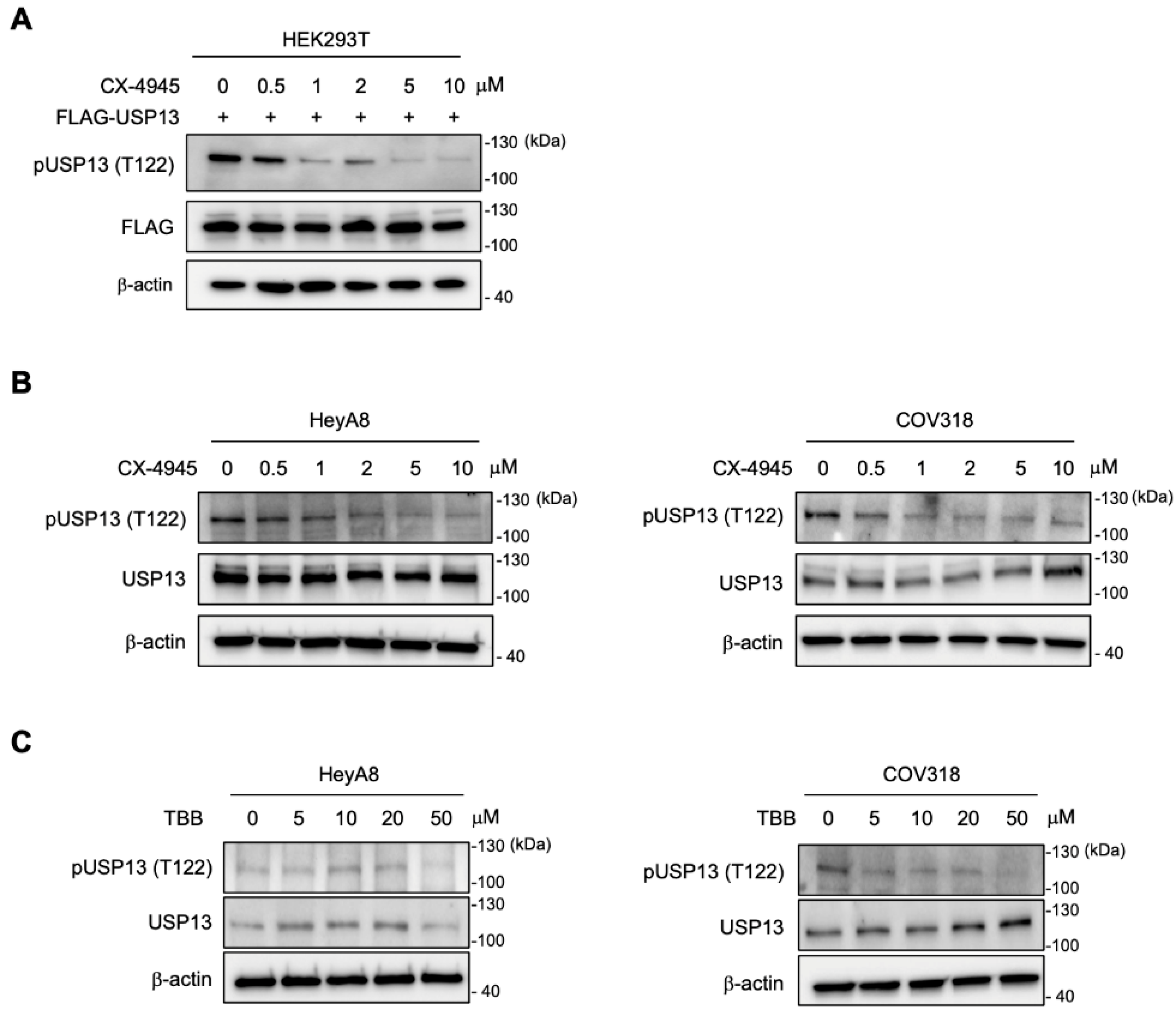
2.4. Casein Kinase 2 Is Responsible for Thr122 Phosphorylation on USP13
2.5. Inhibition of CK2 Decreased Thr122 Phosphorylation on USP13
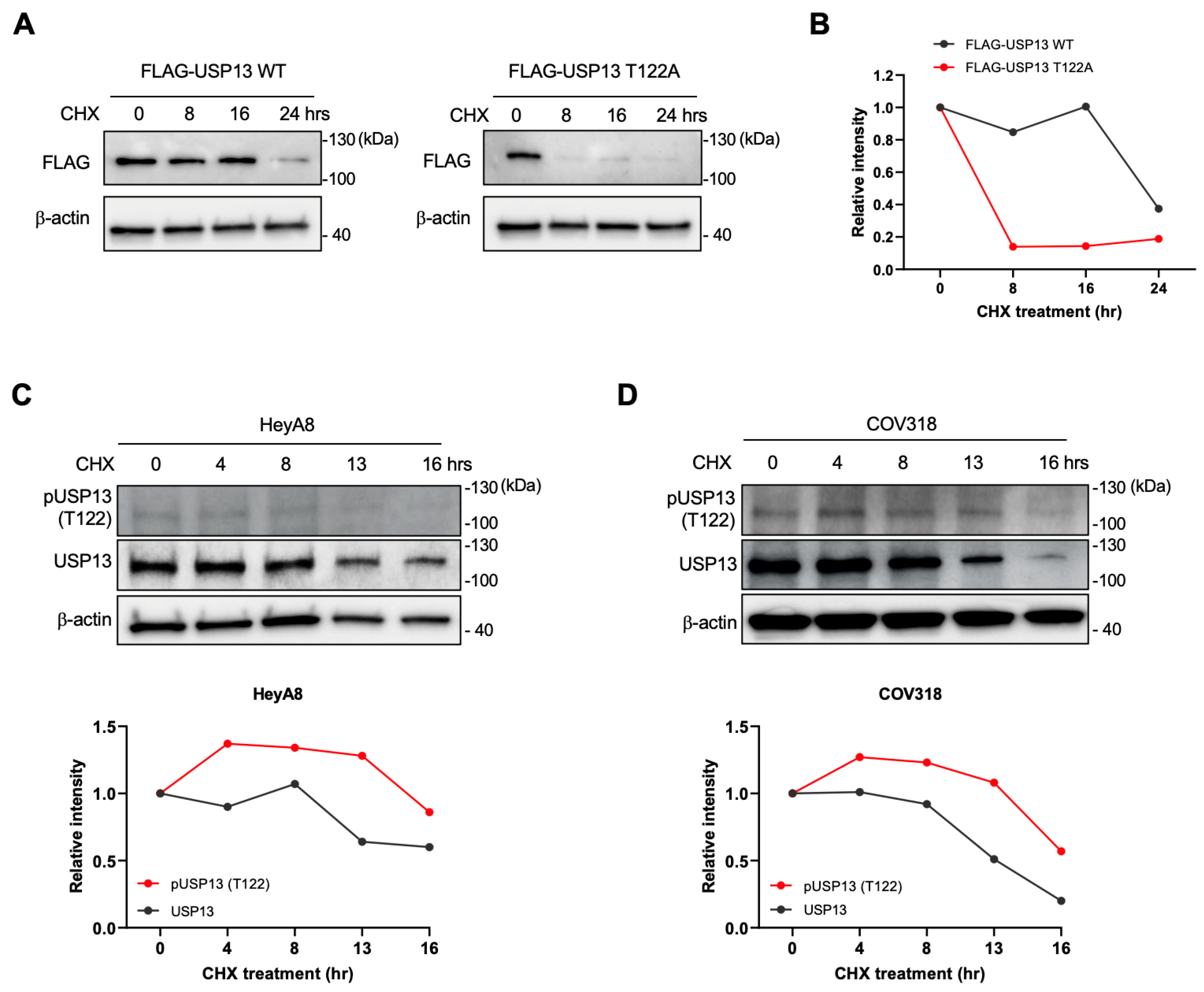
2.6. Thr122 Phosphorylation Upregulated USP13 Stability
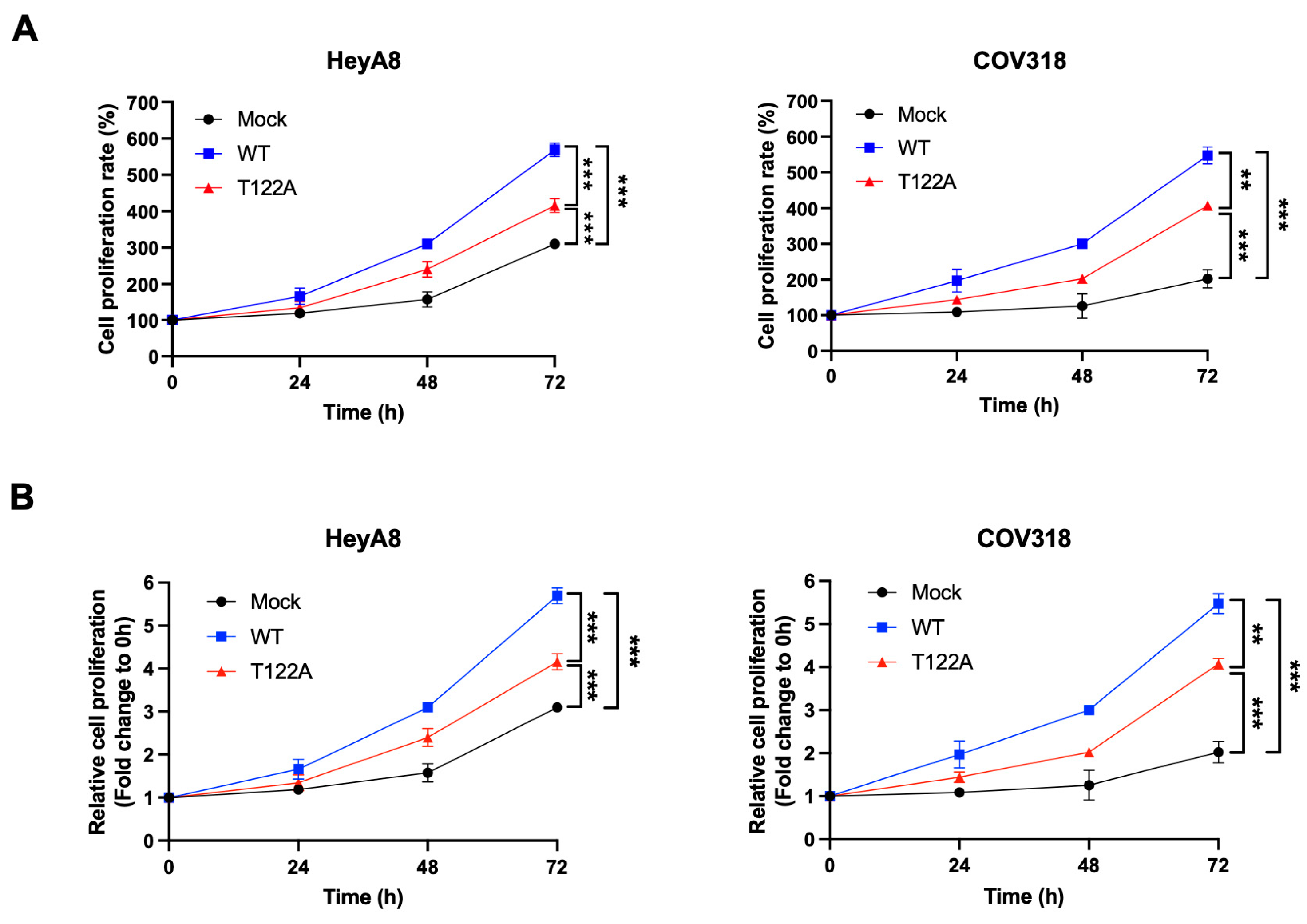
2.7. Thr122 Phosphorylation Is Important for the Proliferation of Ovarian Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Drug Treatment
4.2. Western Blot
4.3. Immunoprecipitation
4.4. Mass-Spectrometry Analysis and Database Search
4.5. Generation of Anti-Phospho-Thr122 USP13 Antibody
4.6. Protein Purification and CK2 Kinase Assay
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Feature article: Treating the untreatable: A breakthrough in understanding the biology and treatment of ovarian cancer. Exp. Biol. Med. 2012, 237, viii. [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Macintyre, G.; Goranova, T.E.; De Silva, D.; Ennis, D.; Piskorz, A.M.; Eldridge, M.; Sie, D.; Lewsley, L.A.; Hanif, A.; Wilson, C.; et al. Copy number signatures and mutational processes in ovarian carcinoma. Nat. Genet 2018, 50, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- Patch, A.M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; Bailey, P.J.; et al. Whole-genome characterization of chemoresistant ovarian cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef]
- Clague, M.J.; Urbe, S.; Komander, D. Breaking the chains: Deubiquitylating enzyme specificity begets function. Nat. Rev. Mol. Cell Biol. 2019, 20, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Mevissen, T.E.T.; Komander, D. Mechanisms of Deubiquitinase Specificity and Regulation. Annu. Rev. Biochem. 2017, 86, 159–192. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Turcu, F.E.; Ventii, K.H.; Wilkinson, K.D. Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu. Rev. Biochem. 2009, 78, 363–397. [Google Scholar] [CrossRef] [Green Version]
- Sowa, M.E.; Bennett, E.J.; Gygi, S.P.; Harper, J.W. Defining the human deubiquitinating enzyme interaction landscape. Cell 2009, 138, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Gutierrez, G.J.; Ronai, Z.A. Ubiquitin-recognition protein Ufd1 couples the endoplasmic reticulum (ER) stress response to cell cycle control. Proc. Natl. Acad. Sci. USA 2011, 108, 9119–9124. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhou, W.; Wu, Q.; Huang, Z.; Shi, Y.; Yang, K.; Chen, C.; Xie, Q.; Mack, S.C.; Wang, X.; et al. Deubiquitinase USP13 maintains glioblastoma stem cells by antagonizing FBXL14-mediated Myc ubiquitination. J. Exp. Med. 2017, 214, 245–267. [Google Scholar] [CrossRef]
- Geng, J.; Huang, X.; Li, Y.; Xu, X.; Li, S.; Jiang, D.; Liang, J.; Jiang, D.; Wang, C.; Dai, H. Down-regulation of USP13 mediates phenotype transformation of fibroblasts in idiopathic pulmonary fibrosis. Respir. Res. 2015, 16, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.; Yang, L.; Choi, H.H.; Baddour, J.; Achreja, A.; Liu, Y.; Li, Y.; Li, J.; Wan, G.; Huang, C.; et al. Amplification of USP13 drives ovarian cancer metabolism. Nat. Commun. 2016, 7, 13525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Kim, J.S.; Diaz-Martinez, L.A.; Han, C.; Lane, W.S.; Budnik, B.; Waldman, T. USP13 interacts with cohesin and regulates its ubiquitination in human cells. J. Biol. Chem. 2021, 296, 100194. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Choi, H.; Ware, A.D.; Morillo, B.C.; Wang, H.; Bouker, K.B.; Lu, X.; Waldman, T.; Han, C. USP13 promotes development and metastasis of high-grade serous ovarian carcinoma in a novel mouse model. Oncogene 2022, 41, 1974–1985. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Luo, K.; Yin, Y.; Wu, C.; Deng, M.; Li, L.; Chen, Y.; Nowsheen, S.; Lou, Z.; Yuan, J. USP13 regulates the RAP80-BRCA1 complex dependent DNA damage response. Nat. Commun. 2017, 8, 15752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xia, H.; Kim, M.; Xu, L.; Li, Y.; Zhang, L.; Cai, Y.; Norberg, H.V.; Zhang, T.; Furuya, T.; et al. Beclin1 controls the levels of p53 by regulating the deubiquitination activity of USP10 and USP13. Cell 2011, 147, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Man, X.; Piao, C.; Lin, X.; Kong, C.; Cui, X.; Jiang, Y. USP13 functions as a tumor suppressor by blocking the NF-kB-mediated PTEN downregulation in human bladder cancer. J. Exp. Clin. Cancer Res. 2019, 38, 259. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Zhang, R.; Su, M.; Liu, W. USP13 serves as a tumor suppressor via the PTEN/AKT pathway in oral squamous cell carcinoma. Cancer Manag. Res. 2019, 11, 9175–9183. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Zhang, Q.; Jing, Y.Y.; Zhang, M.; Wang, H.Y.; Cai, Z.; Liuyu, T.; Zhang, Z.D.; Xiong, T.C.; Wu, Y.; et al. USP13 negatively regulates antiviral responses by deubiquitinating STING. Nat. Commun. 2017, 8, 15534. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.; Fang, J.; Wang, S.; Deng, B.; Zhu, L. MicroRNA135b regulates the stability of PTEN and promotes glycolysis by targeting USP13 in human colorectal cancers. Oncol. Rep. 2015, 33, 1342–1348. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, P.; Wei, Y.; Piao, H.L.; Wang, W.; Maddika, S.; Wang, M.; Chen, D.; Sun, Y.; Hung, M.C.; et al. Deubiquitylation and stabilization of PTEN by USP13. Nat. Cell Biol. 2013, 15, 1486–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhang, M.; Jing, Y.; Yin, X.; Ma, P.; Zhang, Z.; Wang, X.; Di, W.; Zhuang, G. Deubiquitinase USP13 dictates MCL1 stability and sensitivity to BH3 mimetic inhibitors. Nat. Commun. 2018, 9, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Fiske, B.; Kawakami, A.; Li, J.; Fisher, D.E. Regulation of MITF stability by the USP13 deubiquitinase. Nat. Commun. 2011, 2, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhang, Y.; Liu, C.; Zhang, Y.; Wang, D.; Wang, S.; Wu, Y.; Liu, F.; Li, Q.; Liu, X.; et al. Amplification of USP13 drives non-small cell lung cancer progression mediated by AKT/MAPK signaling. Biomed. Pharm. 2019, 114, 108831. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Shin, S.C.; Song, E.J.; Kim, E.E. Regulation of Deubiquitinating Enzymes by Post-Translational Modifications. Int. J. Mol. Sci. 2020, 21, 4028. [Google Scholar] [CrossRef]
- Estavoyer, B.; Messmer, C.; Echbicheb, M.; Rudd, C.E.; Milot, E.; Affar, E.B. Mechanisms orchestrating the enzymatic activity and cellular functions of deubiquitinases. J. Biol. Chem. 2022, 298, 102198. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, F. Post-Translational Modifications of Deubiquitinating Enzymes: Expanding the Ubiquitin Code. Front. Pharm. 2021, 12, 685011. [Google Scholar] [CrossRef]
- Marin, O.; Meggio, F.; Sarno, S.; Cesaro, L.; Pagano, M.A.; Pinna, L.A. Tyrosine versus serine/threonine phosphorylation by protein kinase casein kinase-2. A study with peptide substrates derived from immunophilin Fpr3. J. Biol. Chem. 1999, 274, 29260–29265. [Google Scholar] [CrossRef] [Green Version]
- Silva-Pavez, E.; Tapia, J.C. Protein Kinase CK2 in Cancer Energetics. Front. Oncol. 2020, 10, 893. [Google Scholar] [CrossRef]
- Zeng, K.W.; Wang, J.K.; Wang, L.C.; Guo, Q.; Liu, T.T.; Wang, F.J.; Feng, N.; Zhang, X.W.; Liao, L.X.; Zhao, M.M.; et al. Small molecule induces mitochondrial fusion for neuroprotection via targeting CK2 without affecting its conventional kinase activity. Signal. Transduct. Target. 2021, 6, 71. [Google Scholar] [CrossRef]
- Litchfield, D.W. Protein kinase CK2: Structure, regulation and role in cellular decisions of life and death. Biochem. J. 2003, 369, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Borgo, C.; D’Amore, C.; Sarno, S.; Salvi, M.; Ruzzene, M. Protein kinase CK2: A potential therapeutic target for diverse human diseases. Signal. Transduct Target. 2021, 6, 183. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Lin, M.; Feng, X.; Ma, F.; Zhu, Y.; Liu, X.; Qu, C.; Sui, H.; Sun, B.; Zhu, A.; et al. Targeting CLK3 inhibits the progression of cholangiocarcinoma by reprogramming nucleotide metabolism. J. Exp. Med. 2020, 217, e20191779. [Google Scholar] [CrossRef] [PubMed]
- Esposito, M.; Akman, H.B.; Giron, P.; Ceregido, M.A.; Schepers, R.; Ramos Paez, L.C.; La Monaca, E.; De Greve, J.; Coux, O.; De Trez, C.; et al. USP13 controls the stability of Aurora B impacting progression through the cell cycle. Oncogene 2020, 39, 6009–6023. [Google Scholar] [CrossRef] [PubMed]
- D’Amore, C.; Borgo, C.; Sarno, S.; Salvi, M. Role of CK2 inhibitor CX-4945 in anti-cancer combination therapy—potential clinical relevance. Cell. Oncol. Dordr. 2020, 43, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, G.; Zhang, W.; Qin, B.; Ye, Z.; Shi, H.; Zhao, X.; Chen, Y.; Song, B.; Mei, Z.; et al. USP13: Multiple Functions and Target Inhibition. Front. Cell Dev. Biol. 2022, 10, 875124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Zhou, C.J.; Zhou, Z.R.; Song, A.X.; Hu, H.Y. Domain analysis reveals that a deubiquitinating enzyme USP13 performs non-activating catalysis for Lys63-linked polyubiquitin. PLoS ONE 2011, 6, e29362. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Wang, X.; He, J.; Xia, J.; Li, Y. Increased expression of protein kinase CK2alpha correlates with poor patient prognosis in epithelial ovarian cancer. PLoS ONE 2017, 12, e0174037. [Google Scholar] [CrossRef] [Green Version]
- Salvi, M.; Borgo, C.; Pinna, L.A.; Ruzzene, M. Targeting CK2 in cancer: A valuable strategy or a waste of time? Cell. Death. Discov. 2021, 7, 325. [Google Scholar] [CrossRef]
- Karst, A.M.; Drapkin, R. Primary culture and immortalization of human fallopian tube secretory epithelial cells. Nat. Protoc. 2012, 7, 1755–1764. [Google Scholar] [CrossRef]
- Karst, A.M.; Jones, P.M.; Vena, N.; Ligon, A.H.; Liu, J.F.; Hirsch, M.S.; Etemadmoghadam, D.; Bowtell, D.D.; Drapkin, R. Cyclin E1 deregulation occurs early in secretory cell transformation to promote formation of fallopian tube-derived high-grade serous ovarian cancers. Cancer Res. 2014, 74, 1141–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perets, R.; Wyant, G.A.; Muto, K.W.; Bijron, J.G.; Poole, B.B.; Chin, K.T.; Chen, J.Y.; Ohman, A.W.; Stepule, C.D.; Kwak, S.; et al. Transformation of the fallopian tube secretory epithelium leads to high-grade serous ovarian cancer in Brca;Tp53;Pten models. Cancer Cell 2013, 24, 751–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaspari, M.; Cuda, G. Nano LC-MS/MS: A robust setup for proteomic analysis. Methods Mol. Biol. 2011, 790, 115–126. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, J.; Zhang, J.; Mok, B.; Han, C. CK2-Mediated Phosphorylation Upregulates the Stability of USP13 and Promotes Ovarian Cancer Cell Proliferation. Cancers 2023, 15, 200. https://doi.org/10.3390/cancers15010200
Kwon J, Zhang J, Mok B, Han C. CK2-Mediated Phosphorylation Upregulates the Stability of USP13 and Promotes Ovarian Cancer Cell Proliferation. Cancers. 2023; 15(1):200. https://doi.org/10.3390/cancers15010200
Chicago/Turabian StyleKwon, Juntae, Jinmin Zhang, Boram Mok, and Cecil Han. 2023. "CK2-Mediated Phosphorylation Upregulates the Stability of USP13 and Promotes Ovarian Cancer Cell Proliferation" Cancers 15, no. 1: 200. https://doi.org/10.3390/cancers15010200
APA StyleKwon, J., Zhang, J., Mok, B., & Han, C. (2023). CK2-Mediated Phosphorylation Upregulates the Stability of USP13 and Promotes Ovarian Cancer Cell Proliferation. Cancers, 15(1), 200. https://doi.org/10.3390/cancers15010200