
Interactions between Platelets and Tumor Microenvironment Components in Ovarian Cancer and Their Implications for Treatment and Clinical Outcomes


Abstract
:Simple Summary
Abstract
1. Introduction

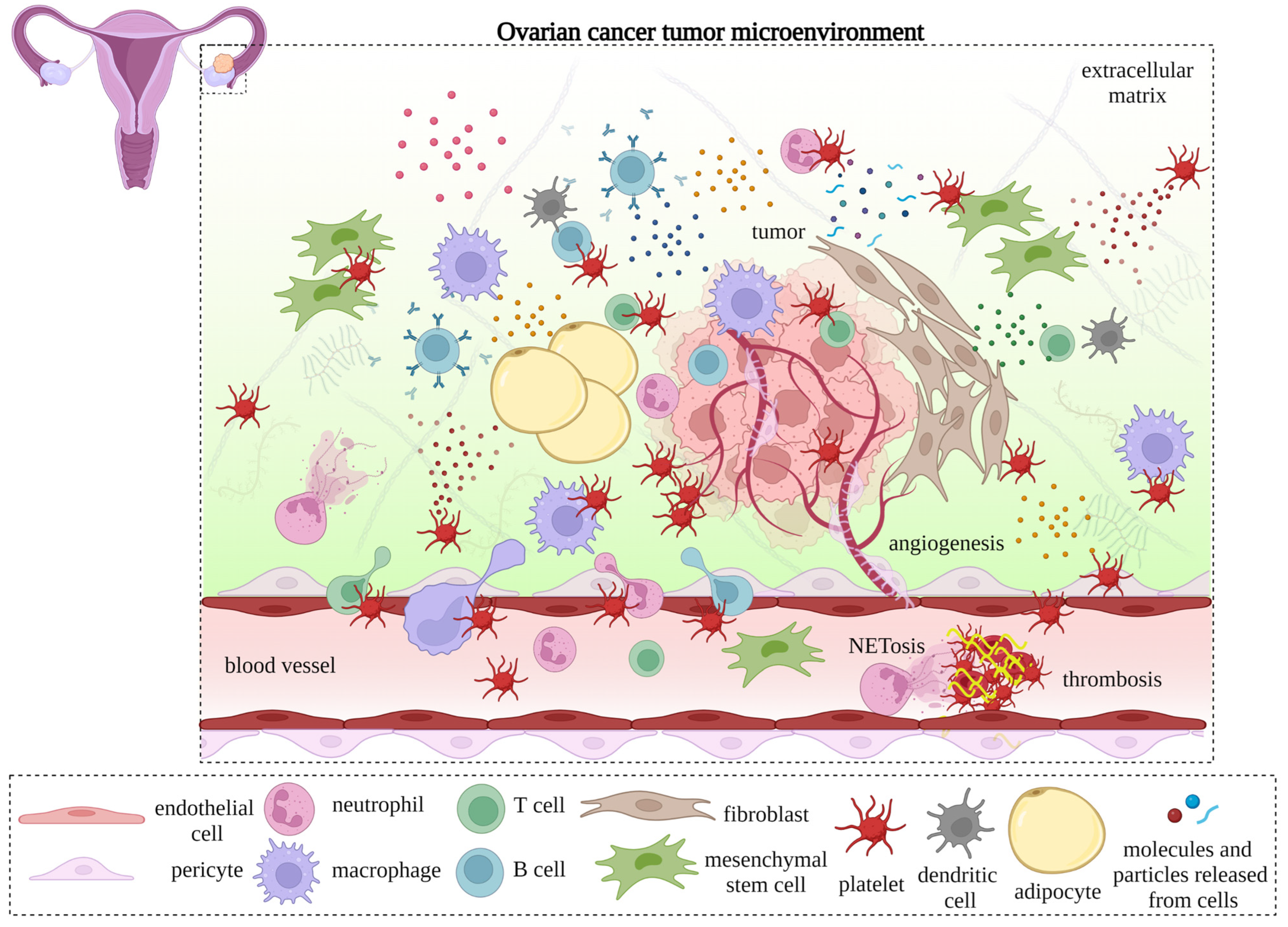{kind=link}
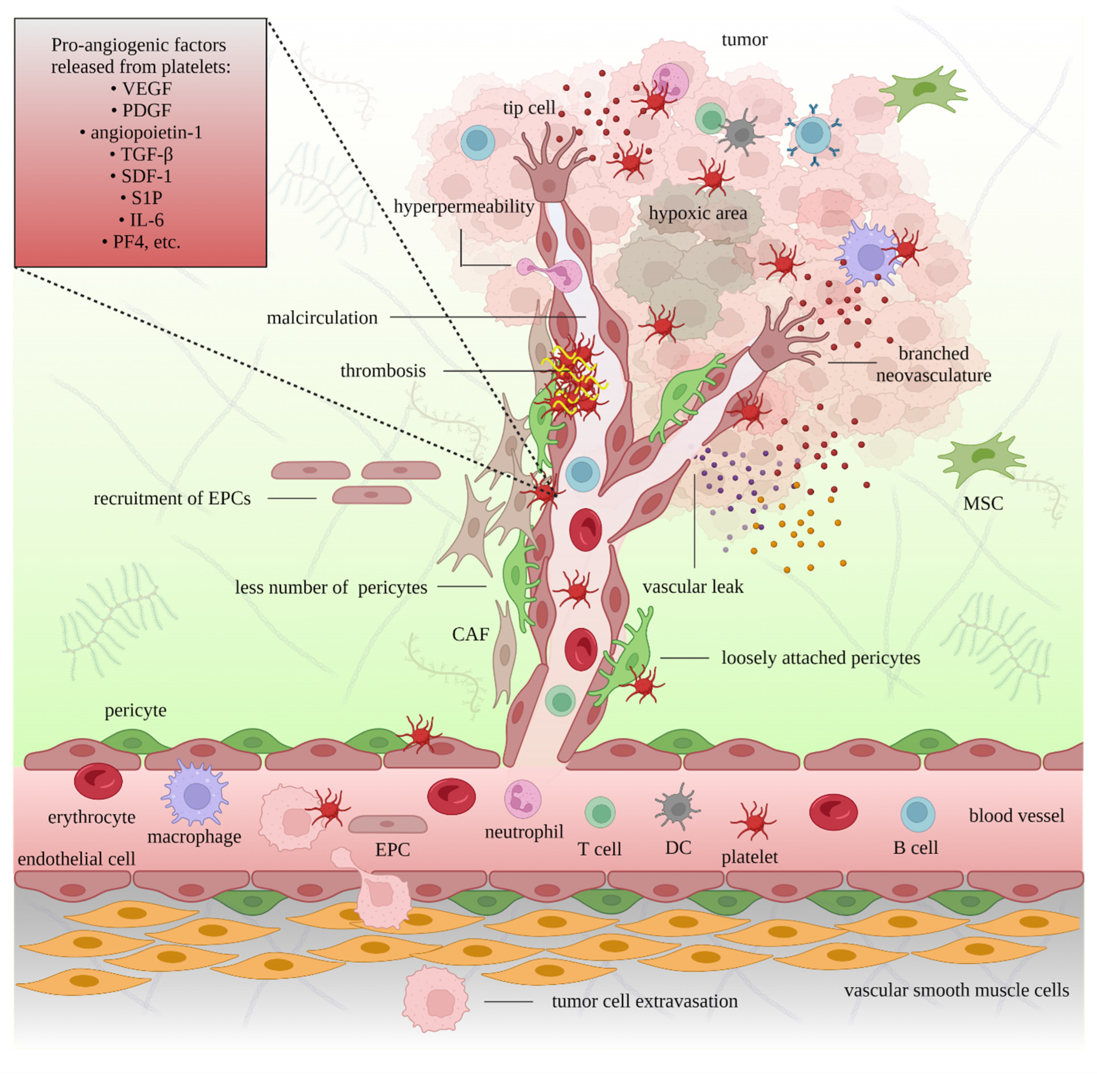{kind=link}
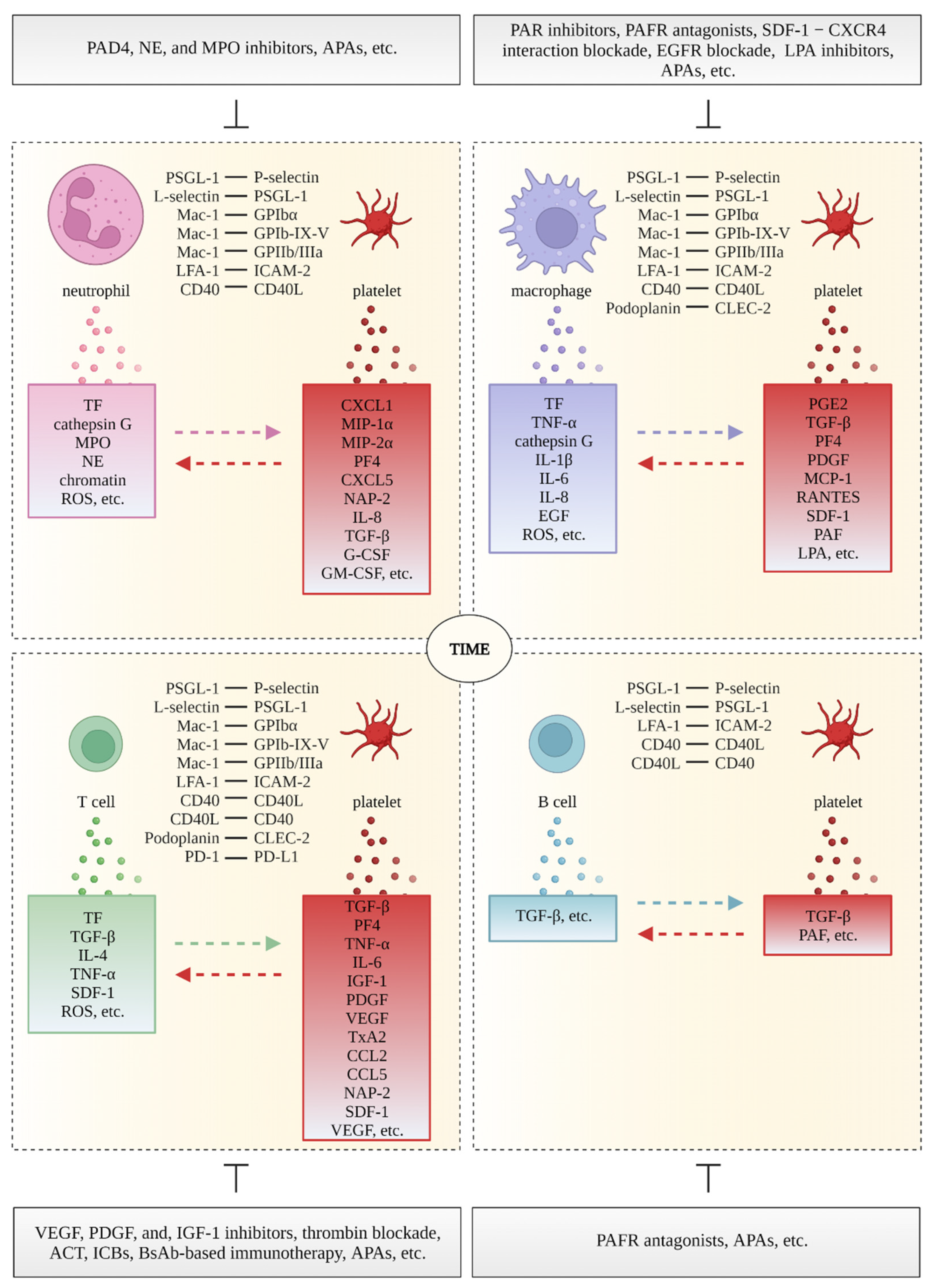{kind=link}
| Location | General Function | Examples | References |
|---|---|---|---|
| Surface molecules | Integrins | α2β1 (GPIa/IIa), α5β1, α6β1, αLβ2 (ICAM-2), αIIbβ3 (GPIIb/IIIa), αVβ3 | [36] |
| Selectins | P-selectin (CD62P), CLEC-2 | [37,38] | |
| Leucine-rich repeat receptors | GPIb-IX-V, TLR1, TLR2, TLR4, TLR6, MMPs | [36,39,40] | |
| ADP receptors | P2Y1, P2Y12 | [41] | |
| Thrombin receptors | PAR1, PAR4, GPIbα | [42,43] | |
| Tetraspanins | CD63, CD9, CD53 | [36] | |
| Prostaglandin receptors | PGD2 and PGE2 receptors | [44] | |
| Prostacyclin receptors | PGI2 receptors | [44] | |
| Thromboxane receptors | TxA2 receptors | [45] | |
| Lipid receptors | PAF and LPA receptors | [46,47] | |
| Ig receptors | GPVI, FcγRIIA (CD32), FcεRI (CD23) | [48,49] | |
| JAMs | JAM-1, JAM-2, JAM-3, PECAM-1 (CD31) | [50,51] | |
| Tyrosine kinase receptors | Thrombopoietin, leptin, insulin, PDGF receptors | [36] | |
| Immune checkpoints | PD-L1, GITRL, OX40L | [52,53,54] | |
| Other receptors | Serotonin receptors, GPIV (CD36), IAP (CD47) complement receptors, CD40, CD40L (CD154) | [55,56,57,58,59] | |
| α-granules | Adhesion molecules | vWF, αIIbβ3 (GPIIb/IIIa), αVβ3, P-selectin (CD62P), fibrinogen, fibronectin, thrombospondin | [36,60,61] |
| Proangiogenic factors | VEGF, angiopoietin-1, SDF-1 (CXCL12), S1P, TGF-β, IL-6, PF4 (CXCL4) | [61] | |
| Angiostatic factors | Endostatin, angiostatin, thrombospondin-1 | [56,61] | |
| Growth factors | VEGF, PDGF, EGF, FGF, HGF, IGF-1, CTGF, TGF-β | [61,62,63,64] | |
| Coagulation-associated components | Prothrombin, fibrinogen, factor V, factor VIII, factor XI, protein S | [61,65,66] | |
| Fibrinolytic factors | α2-macroglobulin, uPA, PAI-1 | [61] | |
| MMPs | MMP-1, MMP-2, MMP-3, MMP-9 | [67,68] | |
| Metalloproteinases | ADAM-10, ADAM-17, ADAMTS-13 | [69] | |
| TIMPs | TIMP-1, TIMP-2, TIMP-4 | [69] | |
| Inflammamodulatory molecules | CXCL1, PF4 (CXCL4), CXCL5, CXCL7 (NAP-2), IL-1β, IL-6, IL-8 (CXCL8), SDF-1 (CXCL12), CCL2 (MCP-1), CCL3 (MIP-1α), CCL5 (RANTES), CCL7, PAF, LPA, TGF-β, TNF-α, GM-CSF | [61,70,71,72,73,74,75] | |
| Immunologic molecules | Complement factors, IgG, IgA, IgM, thymosin-β4 | [61,76,77,78] | |
| Other components | Albumin, α1-antitrypsin, HMWK | [61] | |
| δ-granules | Nucleotides | ADP, ATP, GDP, GTP | [79] |
| Bioactive amines | Serotonin, histamine, epinephrine | [60] | |
| Ions | Calcium, magnesium, phosphate, pyrophosphate | [61] | |
| Polyphosphates | Polyphosphate (polyP) | [60] | |
| Lysosomes | Proteases | Cathepsin D/E, carboxypeptidase A/B, glycohydrolases, collagenase, elastase | [79,80] |
| Phosphatases | Acid phosphatase | [79] | |
| Phospholipases | Phospholipase A | [61] |
2. Interactions of Platelets with TME Compartments: Endothelial Cells, Pericytes, and Cancer-Associated Fibroblasts
2.1. Interactions with Endothelial Cells
2.1.1. In Angiogenesis
2.1.2. In Lymphangiogenesis
2.2. Interactions with Pericytes
2.3. Interactions with Cancer-Associated Fibroblasts
3. Interplay of Platelets with the Tumor Immune Microenvironment
3.1. Interplay with Tumor-Associated Neutrophils
3.2. Interplay with Tumor-Associated Macrophages
3.3. Interplay with T Cells
3.3.1. CD4+ Helper T Cells
3.3.2. Regulatory T Cells
3.3.3. CD8+ T Cells
3.4. Interplay with B Cells
| Hallmark | Platelet Constituents | Prognostic Role | References |
|---|---|---|---|
| Pro- tumorigenic | Endostatin, angiostatin, TSP-1, angiopoietin-1, VEGF, PDFG, HGF, FGF, SDF-1, S1P, TGF-β, IL-6 | Stimulation of angiogenesis | [61,90] |
| TGF-β, angiopoietin, PDGF, CLEC-2, IL-6 | Proliferation, differentiation, and irregularity of pericytes | [64,125,126,128,138] | |
| Platelet microparticles | Overexpression of MMPs on endothelial cells | [100] | |
| P-selectin, GPIIb | Transendothelial migration of cancer cells | [108,109] | |
| VEGF, angiopoietin-1, PDGF, IGF-1, CLEC-2 | Stimulation of lymphangiogenesis | [61,111,113] | |
| TGF-β, SDF-1, PDGF, CLEC-2 | Stimulation and differentiation, proliferation, and migration of CAFs | [15,147,148,149] | |
| TGF-β, G-CSF | Differentiation of neutrophils into N2-like phenotype | [173] | |
| IL-8, G-CSF, GM-CSF, CXCL1, CXCL2, CXCL5, MIP-1α | Recruitment of neutrophils into TME | [192,197] | |
| factor XII, P-selectin, GPIIb/IIIa | Platelet interaction with NETs and subsequent platelet aggregation | [202,204] | |
| mtDNA, P-selectin | NETosis | [202,205] | |
| P-selectin, TGF-β, COX-2, PGE2, PF4, PAF, PDGF | Activation of TAMs | [220,221,222] | |
| CCL2, CCL5, RANTES, SDF-1 | Migration of TAMs into TME | [238,241] | |
| CD40, CCL5, TxA2 | Apoptosis and inhibition of CD4+ T helper cells | [264,298] | |
| PF4, TGF-β, P-selectin | Suppression of Th1 and Th17 differentiation | [288,297,309,310] | |
| P-selectin | Stimulation of T cell exhaustion | [267] | |
| PD-L1, TGF-β | Inhibition of CD4+ and CD8+ T cell antitumor functions | [52,319,320,321,326] | |
| CD40L, P-selectin, TNF-α, IL-6, IGF-1, PDGF, SDF-1 | Activation, proliferation, and migration of Tregs | [16,62,73,74,263,285,293,302,303] | |
| PF4 | Differentiation of monocytes into MDSCs | [21] | |
| CD40L | Expansion of Bregs | [344] | |
| Platelet microparticles | Overexpression of MMPs | [61,100] | |
| CXCL5, CXCL7, TGF-β, TSP-1, P2Y12, COX-1, TxA2 | Formation of pre-metastatic niches | [352] | |
| Anti- tumorigenic | Endostatin, angiostatin, TSP-1 | Inhibition of angiogenesis | [56,61] |
| TRAIL, CCL2, MCP-1, IL-8 | Activation of tumoricidal macrophages | [229] | |
| Platelet microparticles | Enhancement of the phagocytic capacity of macrophages | [228] | |
| TGF-β, EGF | Upregulation of PD-1 in cancer cells and increase in the effectiveness of immunotherapy | [14,270] | |
| TGF-β, CD40L | Activation and infiltration of CD8+ T cells into the tumor | [327,353] | |
| CD40L | Differentiation of resting B cells into plasma cells and generation of antibodies | [342] | |
| OX40L | Infiltration of immune cells into TME | [54] |
4. Discussion and Perspectives
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Jelovac, D.; Armstrong, D.K. Recent progress in the diagnosis and treatment of ovarian cancer. CA Cancer J. Clin. 2011, 61, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Etemadmoghadam, D.; Defazio, A.; Beroukhim, R.; Mermel, C.; George, J.; Getz, G.; Tothill, R.; Okamoto, A.; Raeder, M.B.; Harnett, P.; et al. Integrated Genome-Wide DNA Copy Number and Expression Analysis Identifies Distinct Mechanisms of Primary Chemoresistance in Ovarian Carcinomas. Clin. Cancer Res. 2009, 15, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patch, A.-M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; Bailey, P.J.; et al. Whole–genome characterization of chemoresistant ovarian cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef]
- Jiménez-Sánchez, A.; Cybulska, P.; Mager, K.L.; Koplev, S.; Cast, O.; Couturier, D.-L.; Memon, D.; Selenica, P.; Nikolovski, I.; Mazaheri, Y.; et al. Unraveling tumor–immune heterogeneity in advanced ovarian cancer uncovers immunogenic effect of chemotherapy. Nat. Genet. 2020, 52, 582–593. [Google Scholar] [CrossRef]
- Jin, M.-Z.; Jin, W.-L. The updated landscape of tumor microenvironment and drug repurposing. Signal Transduct. Target. Ther. 2020, 5, 166. [Google Scholar] [CrossRef]
- Jordan, K.R.; Sikora, M.J.; Slansky, J.E.; Minic, A.; Richer, J.K.; Moroney, M.R.; Hu, J.; Wolsky, R.J.; Watson, Z.L.; Yamamoto, T.M.; et al. The Capacity of the Ovarian Cancer Tumor Microenvironment to Integrate Inflammation Signaling Conveys a Shorter Disease-free Interval. Clin. Cancer Res. 2020, 26, 6362–6373. [Google Scholar] [CrossRef]
- CChen, F.; Zhuang, X.; Lin, L.; Yu, P.; Wang, Y.; Shi, Y.; Hu, G.; Sun, Y. New horizons in tumor microenvironment biology: Challenges and opportunities. BMC Med. 2015, 13, 2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H.; et al. The clinical role of the TME in solid cancer. Br. J. Cancer 2019, 120, 45–53. [Google Scholar] [CrossRef]
- Corvigno, S.; Burks, J.K.; Hu, W.; Zhong, Y.; Jennings, N.B.; Fleming, N.D.; Westin, S.N.; Fellman, B.; Liu, J.; Sood, A.K. Immune microenvironment composition in high-grade serous ovarian cancers based on BRCA mutational status. J. Cancer Res. Clin. Oncol. 2021, 147, 3545–3555. [Google Scholar] [CrossRef]
- Kreuzinger, C.; Geroldinger, A.; Smeets, D.; Braicu, E.I.; Sehouli, J.; Koller, J.; Wolf, A.; Darb-Esfahani, S.; Joehrens, K.; Vergote, I.; et al. A Complex Network of Tumor Microenvironment in Human High-Grade Serous Ovarian Cancer. Clin. Cancer Res. 2017, 23, 7621–7632. [Google Scholar] [CrossRef] [Green Version]
- Ukita, M.; Hamanishi, J.; Yoshitomi, H.; Yamanoi, K.; Takamatsu, S.; Ueda, A.; Suzuki, H.; Hosoe, Y.; Furutake, Y.; Taki, M.; et al. CXCL13-producing CD4+ T cells accumulate in the early phase of tertiary lymphoid structures in ovarian cancer. J. Clin. Investig. 2022, 7, e157215. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Chan, D.K.; Yang, D.; Bruno, T.; Lieberman, R.; Siddiqui, J.; Soong, T.R.; Coffman, L.; Buckanovich, R.J. Shifting the Soil: Metformin Treatment Decreases the Protumorigenic Tumor Microenvironment in Epithelial Ovarian Cancer. Cancers 2022, 14, 2298. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.S.; Lee, H.; Gonzalez-Delgado, R.; Li, D.; Sasano, T.; Carlos-Alcalde, W.; Ma, Q.; Liu, J.; Sood, A.K.; Afshar-Kharghan, V. Platelets Increase the Expression of PD-L1 in Ovarian Cancer. Cancers 2022, 14, 2498. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Tajima, H.; Gabata, R.; Okazaki, M.; Shimbashi, H.; Ohbatake, Y.; Okamoto, K.; Nakanuma, S.; Sakai, S.; Makino, I.; et al. Impact of Extravasated Platelet Activation and Podoplanin-positive Cancer-associated Fibroblasts in Pancreatic Cancer Stroma. Anticancer. Res. 2019, 39, 5565–5572. [Google Scholar] [CrossRef] [PubMed]
- Nasti, T.H.; Bullard, D.C.; Yusuf, N. P-selectin enhances growth and metastasis of mouse mammary tumors by promoting regulatory T cell infiltration into the tumors. Life Sci. 2015, 131, 11–18. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Fushida, S.; Kinoshita, J.; Okazaki, M.; Ishikawa, S.; Ohbatake, Y.; Terai, S.; Okamoto, K.; Nakanuma, S.; Makino, I.; et al. Extravasated platelet aggregation contributes to tumor progression via the accumulation of myeloid-derived suppressor cells in gastric cancer with peritoneal metastasis. Oncol. Lett. 2020, 20, 1879–1887. [Google Scholar] [CrossRef]
- Lonsdorf, A.S.; Krämer, B.F.; Fahrleitner, M.; Schönberger, T.; Gnerlich, S.; Ring, S.; Gehring, S.; Schneider, S.W.; Kruhlak, M.J.; Meuth, S.G.; et al. Engagement of αIIbβ3 (GPIIb/IIIa) with ανβ3 Integrin Mediates Interaction of Melanoma Cells with Platelets. J. Biol. Chem. 2012, 287, 2168–2178. [Google Scholar] [CrossRef] [Green Version]
- Shirai, T.; Inoue, O.; Tamura, S.; Tsukiji, N.; Sasaki, T.; Endo, H.; Satoh, K.; Osada, M.; Sato-Uchida, H.; Fujii, H.; et al. C-type lectin-like receptor 2 promotes hematogenous tumor metastasis and prothrombotic state in tumor-bearing mice. J. Thromb. Haemost. 2016, 15, 513–525. [Google Scholar] [CrossRef] [Green Version]
- Haemmerle, M.; Bottsford-Miller, J.; Pradeep, S.; Taylor, M.L.; Choi, H.-J.; Hansen, J.M.; Dalton, H.J.; Stone, R.L.; Cho, M.S.; Nick, A.M.; et al. FAK regulates platelet extravasation and tumor growth after antiangiogenic therapy withdrawal. J. Clin. Investig. 2016, 126, 1885–1896. [Google Scholar] [CrossRef] [Green Version]
- Joseph, R.; Soundararajan, R.; Vasaikar, S.; Yang, F.; Allton, K.L.; Tian, L.; Hollander, P.D.; Isgandarova, S.; Haemmerle, M.; Mino, B.; et al. CD8+ T cells inhibit metastasis and CXCL4 regulates its function. Br. J. Cancer 2021, 125, 176–189. [Google Scholar] [CrossRef]
- Plantureux, L.; Mège, D.; Crescence, L.; Carminita, E.; Robert, S.; Cointe, S.; Brouilly, N.; Ezzedine, W.; Dignat-George, F.; Dubois, C.; et al. The Interaction of Platelets with Colorectal Cancer Cells Inhibits Tumor Growth but Promotes Metastasis. Cancer Res. 2020, 80, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, N.; Rani, B.; Gerwins, P.; Heindryckx, F. Platelets as Key Factors in Hepatocellular Carcinoma. Cancers 2019, 11, 1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levoux, J.; Prola, A.; Lafuste, P.; Gervais, M.; Chevallier, N.; Koumaiha, Z.; Kefi, K.; Braud, L.; Schmitt, A.; Yacia, A.; et al. Platelets Facilitate the Wound-Healing Capability of Mesenchymal Stem Cells by Mitochondrial Transfer and Metabolic Reprogramming. Cell Metab. 2021, 33, 283–299. [Google Scholar] [CrossRef]
- Sibilano, M.; Tullio, V.; Adorno, G.; Savini, I.; Gasperi, V.; Catani, M.V. Platelet-Derived miR-126-3p Directly Targets AKT2 and Exerts Anti-Tumor Effects in Breast Cancer Cells: Further Insights in Platelet-Cancer Interplay. Int. J. Mol. Sci. 2022, 23, 5484. [Google Scholar] [CrossRef] [PubMed]
- Ye, B.; Li, F.; Chen, M.; Weng, Y.; Qi, C.; Xie, Y.; Zhang, Q.; Ding, H.; Zhang, J.; Gao, X. A panel of platelet-associated circulating long non-coding RNAs as potential biomarkers for colorectal cancer. Genomics 2021, 114, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.L.; Nick, A.M.; McNeish, I.A.; Balkwill, F.; Han, H.D.; Bottsford-Miller, J.; Rupaimoole, R.; Armaiz-Pena, G.N.; Pecot, C.V.; Coward, J.; et al. Paraneoplastic Thrombocytosis in Ovarian Cancer. N. Engl. J. Med. 2012, 366, 610–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allensworth, S.; Langstraat, C.; Martin, J.; Lemens, M.; McGree, M.; Weaver, A.; Dowdy, S.; Podratz, K.; Bakkum-Gamez, J. Evaluating the prognostic significance of preoperative thrombocytosis in epithelial ovarian cancer. Gynecol. Oncol. 2013, 130, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.G.; Tran, A.-Q.; Rimel, B.; Cass, I.; Walsh, C.S.; Karlan, B.Y.; Li, A.J. Thrombocytosis at secondary cytoreduction for recurrent ovarian cancer predicts suboptimal resection and poor survival. Gynecol. Oncol. 2014, 132, 556–559. [Google Scholar] [CrossRef]
- Bottsford-Miller, J.; Choi, H.-J.; Dalton, H.J.; Stone, R.L.; Cho, M.S.; Haemmerle, M.; Nick, A.M.; Pradeep, S.; Zand, B.; Previs, R.A.; et al. Differential Platelet Levels Affect Response to Taxane-Based Therapy in Ovarian Cancer. Clin. Cancer Res. 2015, 21, 602–610. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.S.; Bottsford-Miller, J.; Vasquez, H.G.; Stone, R.; Zand, B.; Kroll, M.H.; Sood, A.K.; Afshar-Kharghan, V. Platelets increase the proliferation of ovarian cancer cells. Blood 2012, 120, 4869–4872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Cui, W.; Pei, Y.; Xu, D. Platelets promote invasion and induce epithelial to mesenchymal transition in ovarian cancer cells by TGF-β signaling pathway. Gynecol. Oncol. 2019, 153, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Haemmerle, M.; Taylor, M.L.; Gutschner, T.; Pradeep, S.; Cho, M.S.; Sheng, J.; Lyons, Y.M.; Nagaraja, A.S.; Dood, R.L.; Wen, Y.; et al. Platelets reduce anoikis and promote metastasis by activating YAP1 signaling. Nat. Commun. 2017, 8, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, A.N.; Afshar-Kharghan, V.; Sood, A.K. Platelet Effects on Ovarian Cancer. Semin. Oncol. 2014, 41, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.S.; Gonzalez-Pagan, O.; Pinto, K.C.; Sood, A.; Afshar-Kharghan, V. The Inhibition of Platelets Restore Anti-Tumor Immune Response to Ovarian Cancer and Its Therapeutic Implication. Blood 2018, 132, 3698. [Google Scholar] [CrossRef]
- Saboor, M.; Ayub, Q.; Ilyas, S. Moinuddin Platelet receptors; an instrumental of platelet physiology. Pak. J. Med. Sci. 2013, 29, 891–896. [Google Scholar] [CrossRef]
- McEver, R.P.; Beckstead, J.H.; Moore, K.L.; Marshall-Carlson, L.; Bainton, D.F. GMP-140, a platelet alpha-granule membrane protein, is also synthesized by vascular endothelial cells and is localized in Weibel-Palade bodies. J. Clin. Investig. 1989, 84, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Rayes, J.; Lax, S.; Wichaiyo, S.; Watson, S.K.; Di, Y.; Lombard, S.; Grygielska, B.; Smith, S.W.; Skordilis, K.; Watson, S.P. The podoplanin-CLEC-2 axis inhibits inflammation in sepsis. Nat. Commun. 2017, 8, 2239. [Google Scholar] [CrossRef] [Green Version]
- Cognasse, F.; Nguyen, K.A.; Damien, P.; McNicol, A.; Pozzetto, B.; Hamzeh-Cognasse, H.; Garraud, O. The inflammatory role of platelets via their TLRs and Siglec receptors. Front. Immunol. 2015, 6, 83. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef]
- Cho, M.S.; Noh, K.; Haemmerle, M.; Li, D.; Park, H.; Hu, Q.; Hisamatsu, T.; Mitamura, T.; Mak, S.L.C.; Kunapuli, S.; et al. Role of ADP receptors on platelets in the growth of ovarian cancer. Blood 2017, 130, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Camerer, E.; Qazi, A.A.; Duong, D.N.; Cornelissen, I.; Advincula, R.; Coughlin, S.R. Platelets, protease-activated receptors, and fibrinogen in hematogenous metastasis. Blood 2004, 104, 397–401. [Google Scholar] [CrossRef]
- Jain, S.; Zuka, M.; Liu, J.; Russell, S.; Dent, J.; Guerrero, J.A.; Forsyth, J.; Maruszak, B.; Gartner, T.K.; Felding-Habermann, B.; et al. Platelet glycoprotein Ibα supports experimental lung metastasis. Proc. Natl. Acad. Sci. USA 2007, 104, 9024–9028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tynan, S.S.; Andersen, N.H.; Wills, M.T.; Harker, L.A.; Hanson, S.R. On the multiplicity of platelet prostaglandin receptors. Prostaglandins 1984, 27, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.Z.S.; Jin, J.; Kunapuli, S.P. Molecular Mechanism of Thromboxane A2-induced Platelet Aggregation. J. Biol. Chem. 1999, 274, 29108–29114. [Google Scholar] [CrossRef] [Green Version]
- Burgers, J.A.; Akkerman, J.W.N. Regulation of the receptor for platelet-activating factor on human platelets. Biochem. J. 1993, 291, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, K.; Shibata, S.; Ozaki, Y.; Yatomi, Y.; Igarashi, Y. Identification of lysophospholipid receptors in human platelets: The relation of two agonists, lysophosphatidic acid and sphingosine 1-phosphate. FEBS Lett. 2000, 468, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, M.; Ezumi, Y.; Arai, M.; Takayama, H. A Novel Association of Fc Receptor γ-Chain with Glycoprotein VI and Their Co-expression as a Collagen Receptor in Human Platelets. J. Biol. Chem. 1997, 272, 23528–23531. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, S.; Pawankar, R.; Suzuki, K.; Nakahata, T.; Furukawa, S.; Okumura, K.; Ra, C. Functional expression of the high affinity receptor for IgE (FcepsilonRI) in human platelets and its’ intracellular expression in human megakaryocytes. Blood 1999, 93, 2543–2551. [Google Scholar] [CrossRef]
- Santoso, S.; Sachs, U.J.H.; Kroll, H.; Linder, M.; Ruf, A.; Preissner, K.T.; Chavakis, T. The Junctional Adhesion Molecule 3 (JAM-3) on Human Platelets is a Counterreceptor for the Leukocyte Integrin Mac-1. J. Exp. Med. 2002, 196, 679–691. [Google Scholar] [CrossRef]
- Newman, D.K.; Hamilton, C.; Newman, P.J. Inhibition of antigen-receptor signaling by Platelet Endothelial Cell Adhesion Molecule-1 (CD31) requires functional ITIMs, SHP-2, and p56lck. Blood 2001, 97, 2351–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinterleitner, C.; Strähle, J.; Malenke, E.; Hinterleitner, M.; Henning, M.; Seehawer, M.; Bilich, T.; Heitmann, J.; Lutz, M.; Mattern, S.; et al. Platelet PD-L1 reflects collective intratumoral PD-L1 expression and predicts immunotherapy response in non-small cell lung cancer. Nat. Commun. 2021, 12, 7005. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Heitmann, J.S.; Clar, K.L.; Kropp, K.N.; Hinterleitner, M.; Engler, T.; Koch, A.; Hartkopf, A.D.; Zender, L.; Salih, H.R.; et al. Platelet-expressed immune checkpoint regulator GITRL in breast cancer. Cancer Immunol. Immunother. 2021, 70, 2483–2496. [Google Scholar] [CrossRef] [PubMed]
- Rittig, S.M.; Lutz, M.S.; Clar, K.L.; Zhou, Y.; Kropp, K.N.; Koch, A.; Hartkopf, A.D.; Hinterleitner, M.; Zender, L.; Salih, H.R.; et al. Controversial Role of the Immune Checkpoint OX40L Expression on Platelets in Breast Cancer Progression. Front. Oncol. 2022, 12, 917834. [Google Scholar] [CrossRef]
- Dale, G.L.; Friese, P.; Batar, P.; Hamilton, S.F.; Reed, G.L.; Jackson, K.W.; Clemetson, K.J.; Alberio, L. Stimulated platelets use serotonin to enhance their retention of procoagulant proteins on the cell surface. Nature 2002, 415, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, J.S.; Romeo, M.J.; Yu, C.; Nghiem, K.; Monsale, J.; Rick, M.E.; Wink, D.A.; Frazier, W.A.; Roberts, D.D. Thrombospondin-1 stimulates platelet aggregation by blocking the antithrombotic activity of nitric oxide/cGMP signaling. Blood 2008, 111, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Hamzeh-Cognasse, H.; Damien, P.; Chabert, A.; Pozzetto, B.; Cognasse, F.; Garraud, O. Platelets and Infections-Complex Interactions with Bacteria. Front. Immunol. 2015, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Inwald, D.P.; McDowall, A.; Peters, M.J.; Callard, R.E.; Klein, N.J. CD40 Is Constitutively Expressed on Platelets and Provides a Novel Mechanism for Platelet Activation. Circ. Res. 2003, 92, 1041–1048. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Jochems, C.; Talaie, T.; Anderson, A.; Jales, A.; Tsang, K.Y.; Madan, R.A.; Gulley, J.L.; Schlom, J. Elevated serum soluble CD40 ligand in cancer patients may play an immunosuppressive role. Blood 2012, 120, 3030–3038. [Google Scholar] [CrossRef] [Green Version]
- Fitch-Tewfik, J.L.; Flaumenhaft, R. Platelet Granule Exocytosis: A Comparison with Chromaffin Cells. Front. Endocrinol. 2013, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Wojtukiewicz, M.Z.; Sierko, E.; Hempel, D.; Tucker, S.C.; Honn, K.V. Platelets and cancer angiogenesis nexus. Cancer Metastasis Rev. 2017, 36, 249–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Garcia, A.; Jackson, S.P.; Kunapuli, S.P. Insulin-like growth factor-1 regulates platelet activation through PI3-Kα isoform. Blood 2007, 110, 4206–4213. [Google Scholar] [CrossRef] [PubMed]
- Kubota, S.; Kawata, K.; Yanagita, T.; Doi, H.; Kitoh, T.; Takigawa, M. Abundant Retention and Release of Connective Tissue Growth Factor (CTGF/CCN2) by Platelets. J. Biochem. 2004, 136, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Haemmerle, M.; Stone, R.L.; Menter, D.G.; Afshar-Kharghan, V.; Sood, A.K. The Platelet Lifeline to Cancer: Challenges and Opportunities. Cancer Cell 2018, 33, 965–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, P.N. Platelets and coagulation proteins. Fed. Proc. 1981, 40, 2086–2091. [Google Scholar] [PubMed]
- Stavenuiter, F.; Davis, N.F.; Duan, E.; Gale, A.J.; Heeb, M.J. Platelet protein S directly inhibits procoagulant activity on platelets and microparticles. Thromb. Haemost. 2013, 109, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, V.; Boire, A.; Tchernychev, B.; Kaneider, N.C.; Leger, A.J.; O’Callaghan, K.; Covic, L.; Kuliopulos, A. Platelet Matrix Metalloprotease-1 Mediates Thrombogenesis by Activating PAR1 at a Cryptic Ligand Site. Cell 2009, 137, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Sheu, J.R.; Fong, T.H.; Liu, C.M.; Shen, M.Y.; Chen, T.L.; Chang, Y.; Lu, M.S.; Hsiao, G. Expression of matrix metalloproteinase-9 in human platelets: Regulation of platelet activation in in vitro and in vivo studies. Br. J. Pharmacol. 2004, 143, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Santos-Martínez, M.J.; Medina, C.; Jurasz, P.; Radomski, M.W. Role of metalloproteinases in platelet function. Thromb. Res. 2008, 121, 535–542. [Google Scholar] [CrossRef]
- Lishko, V.K.; Yakubenko, V.P.; Ugarova, T.P.; Podolnikova, N.P. Leukocyte integrin Mac-1 (CD11b/CD18, αMβ2, CR3) acts as a functional receptor for platelet factor 4. J. Biol. Chem. 2018, 293, 6869–6882. [Google Scholar] [CrossRef] [Green Version]
- Walz, A.; Dewald, B.; von Tscharner, V.; Baggiolini, M. Effects of the neutrophil-activating peptide NAP-2, platelet basic protein, connective tissue-activating peptide III and platelet factor 4 on human neutrophils. J. Exp. Med. 1989, 170, 1745–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Jin, G.; Li, W.; McIntyre, T.M. Epidermal Growth Factor (EGF) Autocrine Activation of Human Platelets Promotes EGF Receptor–Dependent Oral Squamous Cell Carcinoma Invasion, Migration, and Epithelial Mesenchymal Transition. J. Immunol. 2018, 201, 2154–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhong, H.; Zhao, Y.; Luo, X.; Gao, W. Role of platelet biomarkers in inflammatory response. Biomark. Res. 2020, 8, 28. [Google Scholar] [CrossRef]
- Bock, M.; Bergmann, C.B.; Jung, S.; Kalbitz, M.; Relja, B.; Huber-Wagner, S.; Biberthaler, P.; van Griensven, M.; Hanschen, M. The posttraumatic activation of CD4+ T regulatory cells is modulated by TNFR2- and TLR4-dependent pathways, but not by IL-10. Cell. Immunol. 2018, 331, 137–145. [Google Scholar] [CrossRef]
- Raiden, S.; Schettini, J.; Salamone, G.; Trevani, A.; Vermeulen, M.; Gamberale, R.; Giordano, M.; Geffner, J. Human Platelets Produce Granulocyte-Macrophage Colony-Stimulating Factor and Delay Eosinophil Apoptosis. Lab. Investig. 2003, 83, 589–598. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Conway, E.M. Platelets and Complement Crosstalk in Early Atherogenesis. Front. Cardiovasc. Med. 2019, 6, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, J.N.; Saucerman, S. Platelet Igg, Iga, Igm, and Albumin: Correlation of Platelet and Plasma Concentrations in Normal Subjects and in Patients with Itp or Dysproteinemia. Blood 1988, 72, 362–365. [Google Scholar] [CrossRef]
- Huff, T.; Otto, A.M.; Müller, C.S.G.; Meier, M.; Hannappel, E. Thymosin β4 is released from human blood platelets and attached by factor XIIIa (transglutaminase) to fibrin and collagen. FASEB J. 2002, 16, 691–696. [Google Scholar] [CrossRef] [Green Version]
- de Jong, J.S.S.G.; Dekker, L.R.C. Platelets and Cardiac Arrhythmia. Front. Physiol. 2010, 1, 166. [Google Scholar] [CrossRef] [Green Version]
- Chesney, C.M.; Harper, E.; Colman, R.W. Human Platelet Collagenase. J. Clin. Investig. 1974, 53, 1647–1654. [Google Scholar] [CrossRef] [Green Version]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J.A.; Chang, S.-H.; Dvorak, A.M.; Dvorak, H.F. Why are tumour blood vessels abnormal and why is it important to know. Br. J. Cancer 2009, 100, 865–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, D. The Tumor Vascular Endothelium as Decision Maker in Cancer Therapy. Front. Oncol. 2018, 8, 367. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.J.; Olive, P.L.; Durand, R.E. Intermittent blood flow in a murine tumor: Radiobiological effects. Cancer Res. 1987, 47, 597–601. [Google Scholar] [PubMed]
- Fitzgerald, G.; Soro-Arnaiz, I.; de Bock, K. The Warburg Effect in Endothelial Cells and its Potential as an Anti-angiogenic Target in Cancer. Front. Cell Dev. Biol. 2018, 6, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvorak, H.F.; Brown, L.F.; Detmar, M.; Dvorak, A.M. Vascular permeability factor/vascular endothelial growth factor, microvascular hyperpermeability, and angiogenesis. Am. J. Pathol. 1995, 146, 1029–1039. [Google Scholar] [PubMed]
- Perren, T.J.; Swart, A.M.; Pfisterer, J.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; Kurzeder, C.; et al. A Phase 3 Trial of Bevacizumab in Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2484–2496. [Google Scholar] [CrossRef] [Green Version]
- Burger, R.A.; Sill, M.W.; Monk, B.J.; Greer, B.E.; Sorosky, J.I. Phase II Trial of Bevacizumab in Persistent or Recurrent Epithelial Ovarian Cancer or Primary Peritoneal Cancer: A Gynecologic Oncology Group Study. J. Clin. Oncol. 2007, 25, 5165–5171. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.-J.; Pena, G.N.A.; Pradeep, S.; Cho, M.S.; Coleman, R.L.; Sood, A.K. Anti-vascular therapies in ovarian cancer: Moving beyond anti-VEGF approaches. Cancer Metastasis Rev. 2014, 34, 19–40. [Google Scholar] [CrossRef] [Green Version]
- Kisucka, J.; Butterfield, C.E.; Duda, D.G.; Eichenberger, S.C.; Saffaripour, S.; Ware, J.; Ruggeri, Z.M.; Jain, R.K.; Folkman, J.; Wagner, D.D. Platelets and platelet adhesion support angiogenesis while preventing excessive hemorrhage. Proc. Natl. Acad. Sci. USA 2006, 103, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Martins, P.D.C.; García-Vallejo, J.-J.; van Thienen, J.V.; Fernandez-Borja, M.; van Gils, J.M.; Beckers, C.; Horrevoets, A.J.; Hordijk, P.L.; Zwaginga, J.-J. P-Selectin Glycoprotein Ligand-1 Is Expressed on Endothelial Cells and Mediates Monocyte Adhesion to Activated Endothelium. Arter. Thromb. Vasc. Biol. 2007, 27, 1023–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, L.; Cai, Y.; Zhou, X.; Yang, H. Activated Platelets Interact with Lung Cancer Cells Through P-Selectin Glycoprotein Ligand-1. Pathol. Oncol. Res. 2012, 18, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Roest, M.; de Laat, B.; de Groot, P.G.; Huskens, D. Interplay between platelets and coagulation. Blood Rev. 2020, 46, 100733. [Google Scholar] [CrossRef]
- Yan, M.; Lesyk, G.; Radziwon-Balicka, A.; Jurasz, P. Pharmacological Regulation of Platelet Factors That Influence Tumor Angiogenesis. Semin. Oncol. 2014, 41, 370–377. [Google Scholar] [CrossRef]
- Raica, M.; Cimpean, A.M. Platelet-Derived Growth Factor (PDGF)/PDGF Receptors (PDGFR) Axis as Target for Antitumor and Antiangiogenic Therapy. Pharmaceuticals 2010, 3, 572–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, H.; May, A.E.; Daub, K.; Heinzmann, U.; Lang, P.; Schumm, M.; Vestweber, D.; Massberg, S.; Schönberger, T.; Pfisterer, I.; et al. Adherent Platelets Recruit and Induce Differentiation of Murine Embryonic Endothelial Progenitor Cells to Mature Endothelial Cells In Vitro. Circ. Res. 2006, 98, e2–e10. [Google Scholar] [CrossRef] [Green Version]
- Trikha, M.; Zhou, Z.; Timar, J.; Raso, E.; Kennel, M.; Emmell, E.; Nakada, M.T. Multiple roles for platelet GPIIb/IIIa and alphavbeta3 integrins in tumor growth, angiogenesis, and metastasis. Cancer Res. 2002, 62, 2824–2833. [Google Scholar]
- Ho-Tin-Noé, B.; Goerge, T.; Cifuni, S.M.; Duerschmied, D.; Wagner, D.D. Platelet Granule Secretion Continuously Prevents Intratumor Hemorrhage. Cancer Res. 2008, 68, 6851–6858. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, D.; Strilic, B.; Sivaraj, K.K.; Wettschureck, N.; Offermanns, S. Platelet-Derived Nucleotides Promote Tumor-Cell Transendothelial Migration and Metastasis via P2Y2 Receptor. Cancer Cell 2013, 24, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Feng, S.-B.; Cao, Z.-W.; Bei, J.-J.; Chen, Q.; Xu, X.-J.; Zhou, Z.; Yu, Z.-P.; Hu, H.-Y. Up-Regulated Expression of Matrix Metalloproteinases in Endothelial Cells Mediates Platelet Microvesicle-Induced Angiogenesis. Cell. Physiol. Biochem. 2017, 41, 2319–2332. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, X. Platelets are associated with xenograft tumor growth and the clinical malignancy of ovarian cancer through an angiogenesis-dependent mechanism. Mol. Med. Rep. 2014, 11, 2449–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artini, P.G.; Ruggiero, M.; Monteleone, P.; Carpi, A.; Cristello, F.; Cela, V.; Genazzani, A.R. Vascular endothelial growth factor and its soluble receptor in benign and malignant ovarian tumors. Biomed. Pharmacother. 2008, 62, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Byrne, A.T.; Ross, L.; Holash, J.; Nakanishi, M.; Hu, L.; Hofmann, J.I.; Yancopoulos, G.D.; Jaffe, R.B. Vascular endothelial growth factor-trap decreases tumor burden, inhibits ascites, and causes dramatic vascular remodeling in an ovarian cancer model. Clin. Cancer Res. 2003, 9, 5721–5728. [Google Scholar] [PubMed]
- Li, L.; Wang, L.; Zhang, W.; Tang, B.; Zhang, J.; Song, H.; Yao, D.; Tang, Y.; Chen, X.; Yang, Z.; et al. Correlation of serum VEGF levels with clinical stage, therapy efficacy, tumor metastasis and patient survival in ovarian cancer. Anticancer. Res. 2004, 24, 1973–1979. [Google Scholar] [PubMed]
- Matei, D.; Kelich, S.; Cao, L.; Menning, N.; Emerson, R.E.; Rao, J.; Jeng, M.H.; Sledge, G.W. PDGF BB induces VEGF secretion in ovarian cancer. Cancer Biol. Ther. 2007, 6, 1951–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silini, A.; Ghilardi, C.; Figini, S.; Sangalli, F.; Fruscio, R.; Dahse, R.; Pedley, R.B.; Giavazzi, R.; Bani, M. Regulator of G-protein signaling 5 (RGS5) protein: A novel marker of cancer vasculature elicited and sustained by the tumor’s proangiogenic microenvironment. Cell. Mol. Life Sci. 2011, 69, 1167–1178. [Google Scholar] [CrossRef] [Green Version]
- Bergmeier, W.; Piffath, C.L.; Goerge, T.; Cifuni, S.M.; Ruggeri, Z.M.; Ware, J.; Wagner, D.D. The role of platelet adhesion receptor GPIbα far exceeds that of its main ligand, von Willebrand factor, in arterial thrombosis. Proc. Natl. Acad. Sci. USA 2006, 103, 16900–16905. [Google Scholar] [CrossRef] [Green Version]
- Borsig, L.; Wong, R.; Feramisco, J.; Nadeau, D.R.; Varki, N.M.; Varki, A. Heparin and cancer revisited: Mechanistic connections involving platelets, P-selectin, carcinoma mucins, and tumor metastasis. Proc. Natl. Acad. Sci. USA 2001, 98, 3352–3357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Zhang, W.-J.; Cai, H.-Q.; Liu, H.-L.; Peng, L.; Li, C.-H.; Ye, L.-Y.; Xu, S.-Q.; Yang, Z.-H.; Lou, J.-N. Platelet adhesion and fusion to endothelial cell facilitate the metastasis of tumor cell in hypoxia-reoxygenation condition. Clin. Exp. Metastasis 2010, 28, 1–12. [Google Scholar] [CrossRef]
- Stacker, S.A.; Williams, S.P.; Karnezis, T.; Shayan, R.; Fox, S.B.; Achen, M.G. Lymphangiogenesis and lymphatic vessel remodelling in cancer. Nat. Rev. Cancer 2014, 14, 159–172. [Google Scholar] [CrossRef]
- Suzuki-Inoue, K. Essential in vivo roles of the platelet activation receptor CLEC-2 in tumour metastasis, lymphangiogenesis and thrombus formation. J. Biochem. 2011, 150, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.; Bui, H.; Farrelly, O.; Yang, J.; Li, L.; Enis, D.; Ma, W.; Chen, M.; Oliver, G.; Welsh, J.D.; et al. Hemostasis stimulates lymphangiogenesis through release and activation of VEGFC. Blood 2019, 134, 1764–1775. [Google Scholar] [CrossRef] [PubMed]
- Da, M.-X.; Wu, Z.; Tian, H.-W. Tumor Lymphangiogenesis and Lymphangiogenic Growth Factors. Arch. Med. Res. 2008, 39, 365–372. [Google Scholar] [CrossRef]
- Sopo, M.; Anttila, M.; Muukkonen, O.-T.; Ylä-Herttuala, S.; Kosma, V.-M.; Keski-Nisula, L.; Sallinen, H. Microvessels in Epithelial Ovarian Tumors: High Microvessel Density Is a Significant Feature of Malignant Ovarian Tumors. Anticancer. Res. 2020, 40, 6923–6931. [Google Scholar] [CrossRef]
- Kitano, H.; Kageyama, S.-I.; Hewitt, S.M.; Hayashi, R.; Doki, Y.; Ozaki, Y.; Fujino, S.; Takikita, M.; Kubo, H.; Fukuoka, J. Podoplanin Expression in Cancerous Stroma Induces Lymphangiogenesis and Predicts Lymphatic Spread and Patient Survival. Arch. Pathol. Lab. Med. 2010, 134, 1520–1527. [Google Scholar] [CrossRef]
- Bianchi, R.; Fischer, E.; Yuen, D.; Ernst, E.; Ochsenbein, A.M.; Chen, L.; Otto, V.I.; Detmar, M. Mutation of Threonine 34 in Mouse Podoplanin-Fc Reduces CLEC-2 Binding and Toxicity in Vivo While Retaining Anti-lymphangiogenic Activity. J. Biol. Chem. 2014, 289, 21016–21027. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.-W.; Yang, S.-H.; Cai, L.-Q.; Zhang, J.; Wang, J.; Wang, Z.-H. Roles of vascular endothelial growth factor and platelet-derived growth factor in lymphangiogenesis in epithelial ovarian carcinoma. Zhonghua Fu Chan Ke Za Zhi 2009, 44, 760–764. [Google Scholar]
- Krishnapriya, S.; Sidhanth, C.; Manasa, P.; Sneha, S.; Bindhya, S.; Nagare, R.P.; Ramachandran, B.; Vishwanathan, P.; Murhekar, K.; Shirley, S.; et al. Cancer stem cells contribute to angiogenesis and lymphangiogenesis in serous adenocarcinoma of the ovary. Angiogenesis 2019, 22, 441–455. [Google Scholar] [CrossRef]
- Liao, S.; Liu, J.; Lin, P.; Shi, T.; Jain, R.K.; Xu, L. TGF-β Blockade Controls Ascites by Preventing Abnormalization of Lymphatic Vessels in Orthotopic Human Ovarian Carcinoma Models. Clin. Cancer Res. 2011, 17, 1415–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, M.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the neurovascular unit: Key functions and signaling pathways. Nat. Neurosci. 2016, 19, 771–783. [Google Scholar] [CrossRef]
- Xian, X.; Håkansson, J.; Ståhlberg, A.; Lindblom, P.; Betsholtz, C.; Gerhardt, H.; Semb, H. Pericytes limit tumor cell metastasis. J. Clin. Investig. 2006, 116, 642–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, A.; Franklin, M.J.; Dudek, A.Z. Pericytes and vessel maturation during tumor angiogenesis and metastasis. Am. J. Hematol. 2010, 85, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Song, S. The role of pericytes in blood-vessel formation and maintenance. Neuro-Oncology 2005, 7, 452–464. [Google Scholar] [CrossRef] [Green Version]
- Sinha, D.; Chong, L.; George, J.; Schlueter, H.; Mönchgesang, S.; Mills, S.; Jason Australian Ovarian Cancer Study Group; Parish, C.R.; Bowtell, D.D.; Kaur, P. Pericytes Promote Malignant Ovarian Cancer Progression in Mice and Predict Poor Prognosis in Serous Ovarian Cancer Patients. Clin. Cancer Res. 2016, 22, 1813–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasano, T.; Gonzalez-Delgado, R.; Muñoz, N.M.; Carlos-Alcade, W.; Cho, M.S.; Sheth, R.A.; Sood, A.K.; Afshar-Kharghan, V. Podoplanin promotes tumor growth, platelet aggregation, and venous thrombosis in murine models of ovarian cancer. J. Thromb. Haemost. 2021, 20, 104–114. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: Developmental, Physiological, and Pathological Perspectives, Problems, and Promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [Green Version]
- Zonneville, J.; Safina, A.; Truskinovsky, A.M.; Arteaga, C.L.; Bakin, A.V. TGF-β signaling promotes tumor vasculature by enhancing the pericyte-endothelium association. BMC Cancer 2018, 18, 670. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yang, N.; Park, J.-W.; Katsaros, D.; Fracchioli, S.; Cao, G.; O’Brien-Jenkins, A.; Randall, T.C.; Rubin, S.C.; Coukos, G. Tumor-derived vascular endothelial growth factor up-regulates angiopoietin-2 in host endothelium and destabilizes host vasculature, supporting angiogenesis in ovarian cancer. Cancer Res. 2003, 63, 3403–3412. [Google Scholar]
- Sallinen, H.; Anttila, M.; Gröhn, O.; Koponen, J.; Hämäläinen, K.; Kholova, I.; Kosma, V.-M.; Heinonen, S.; Alitalo, K.; Ylä-Herttuala, S. Cotargeting of VEGFR-1 and -3 and angiopoietin receptor Tie2 reduces the growth of solid human ovarian cancer in mice. Cancer Gene Ther. 2010, 18, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Heldin, C.-H.; Westermark, B. Mechanism of Action and In Vivo Role of Platelet-Derived Growth Factor. Physiol. Rev. 1999, 79, 1283–1316. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Shahzad, M.M.; Moreno-Smith, M.; Lin, Y.; Jennings, N.B.; Allen, J.K.; Landen, C.N.; Mangala, L.S.; Armaiz-Pena, G.N.; Schmandt, R.; et al. Targeting pericytes with a PDGF-B aptamer in human ovarian carcinoma models. Cancer Biol. Ther. 2010, 9, 176–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taraboletti, G.; Roberts, D.; Liotta, L.A.; Giavazzi, R. Platelet thrombospondin modulates endothelial cell adhesion, motility, and growth: A potential angiogenesis regulatory factor. J. Cell Biol. 1990, 111, 765–772. [Google Scholar] [CrossRef]
- Bagavandoss, P.; Wilks, J. Specific inhibition of endothelial cell proliferation by thrombospondin. Biochem. Biophys. Res. Commun. 1990, 170, 867–872. [Google Scholar] [CrossRef]
- Matuszewska, K.; Kortenaar, S.T.; Pereira, M.; Santry, L.A.; Petrik, D.; Lo, K.-M.; Bridle, B.W.; Wootton, S.K.; Lawler, J.; Petrik, J. Addition of an Fc-IgG induces receptor clustering and increases the in vitro efficacy and in vivo anti-tumor properties of the thrombospondin-1 type I repeats (3TSR) in a mouse model of advanced stage ovarian cancer. Gynecol. Oncol. 2021, 164, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.; Duquette, M.; Liu, J.; Drapkin, R.; Lawler, J.; Petrik, J. Combined therapy with thrombospondin-1 type I repeats (3TSR) and chemotherapy induces regression and significandy improves survival in a preclinical model of advanced stage epithelial ovarian cancer. FASEB J. 2014, 29, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Matuszewska, K.; Santry, L.A.; van Vloten, J.P.; Auyeung, A.W.K.; Major, P.P.; Lawler, J.; Wootton, S.K.; Bridle, B.W.; Petrik, J. Combining Vascular Normalization with an Oncolytic Virus Enhances Immunotherapy in a Preclinical Model of Advanced-Stage Ovarian Cancer. Clin. Cancer Res. 2019, 25, 1624–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucharaba, A.; Serre, C.-M.; Grès, S.; Saulnier-Blache, J.S.; Bordet, J.-C.; Guglielmi, J.; Clézardin, P.; Peyruchaud, O. Platelet-derived lysophosphatidic acid supports the progression of osteolytic bone metastases in breast cancer. J. Clin. Investig. 2004, 114, 1714–1725. [Google Scholar] [CrossRef] [Green Version]
- Gopinathan, G.; Milagre, C.; Pearce, O.M.; Reynolds, L.E.; Hodivala-Dilke, K.; Leinster, D.A.; Zhong, H.; Hollingsworth, R.E.; Thompson, R.; Whiteford, J.R.; et al. Interleukin-6 Stimulates Defective Angiogenesis. Cancer Res. 2015, 75, 3098–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merritt, W.M.; Nick, A.M.; Carroll, A.R.; Lu, C.; Matsuo, K.; Dumble, M.; Jennings, N.; Zhang, S.; Lin, Y.G.; Spannuth, W.A.; et al. Bridging the Gap between Cytotoxic and Biologic Therapy with Metronomic Topotecan and Pazopanib in Ovarian Cancer. Mol. Cancer Ther. 2010, 9, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Tullemans, B.M.E.; Nagy, M.; Sabrkhany, S.; Griffioen, A.W.; Egbrink, M.G.A.O.; Aarts, M.; Heemskerk, J.W.M.; Kuijpers, M.J.E. Tyrosine Kinase Inhibitor Pazopanib Inhibits Platelet Procoagulant Activity in Renal Cell Carcinoma Patients. Front. Cardiovasc. Med. 2018, 5, 142. [Google Scholar] [CrossRef] [Green Version]
- Tracy, L.E.; Minasian, R.A.; Caterson, E. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Luga, V.; Zhang, L.; Viloria-Petit, A.M.; Ogunjimi, A.A.; Inanlou, M.R.; Chiu, E.; Buchanan, M.; Hosein, A.N.; Basik, M.; Wrana, J.L. Exosomes Mediate Stromal Mobilization of Autocrine Wnt-PCP Signaling in Breast Cancer Cell Migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Yan, C.; Mu, L.; Huang, K.; Li, X.; Tao, D.; Wu, Y.; Qin, J. Fibroblast-derived exosomes contribute to chemoresistance through priming cancer stem cells in colorectal cancer. PLoS ONE 2015, 10, e0125625. [Google Scholar] [CrossRef] [Green Version]
- Ronca, R.; van Ginderachter, J.; Turtoi, A. Paracrine interactions of cancer-associated fibroblasts, macrophages and endothelial cells: Tumor allies and foes. Curr. Opin. Oncol. 2018, 30, 45–53. [Google Scholar] [CrossRef]
- Kojima, Y.; Acar, A.; Eaton, E.N.; Mellody, K.T.; Scheel, C.; Ben-Porath, I.; Onder, T.T.; Wang, Z.C.; Richardson, A.L.; Weinberg, R.A.; et al. Autocrine TGF-β and stromal cell-derived factor-1 (SDF-1) signaling drives the evolution of tumor-promoting mammary stromal myofibroblasts. Proc. Natl. Acad. Sci. USA 2010, 107, 20009–20014. [Google Scholar] [CrossRef] [Green Version]
- Anderberg, C.; Li, H.; Fredriksson, L.; Andrae, J.; Betsholtz, C.; Li, X.; Eriksson, U.; Pietras, K. Paracrine Signaling by Platelet-Derived Growth Factor-CC Promotes Tumor Growth by Recruitment of Cancer-Associated Fibroblasts. Cancer Res. 2009, 69, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Aoto, K.; Ito, K.; Aoki, S. Complex formation between platelet-derived growth factor receptor β and transforming growth factor β receptor regulates the differentiation of mesenchymal stem cells into cancer-associated fibroblasts. Oncotarget 2018, 9, 34090–34102. [Google Scholar] [CrossRef] [Green Version]
- Zeltz, C.; Alam, J.; Liu, H.; Erusappan, P.M.; Hoschuetzky, H.; Molven, A.; Parajuli, H.; Cukierman, E.; Costea, D.-E.; Lu, N.; et al. α11β1 Integrin is Induced in a Subset of Cancer-Associated Fibroblasts in Desmoplastic Tumor Stroma and Mediates In Vitro Cell Migration. Cancers 2019, 11, 765. [Google Scholar] [CrossRef] [Green Version]
- Fukagawa, D.; Sugai, T.; Osakabe, M.; Suga, Y.; Nagasawa, T.; Itamochi, H.; Sugiyama, T. Protein expression patterns in cancer-associated fibroblasts and cells undergoing the epithelial-mesenchymal transition in ovarian cancers. Oncotarget 2018, 9, 27514–27524. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, R.; Ha, J.H.; Jayaraman, M.; Liu, J.; Moxley, K.M.; Isidoro, C.; Sood, A.K.; Song, Y.S.; Dhanasekaran, D.N. Ovarian cancer cell-derived lysophosphatidic acid induces glycolytic shift and cancer-associated fibroblast-phenotype in normal and peritumoral fibroblasts. Cancer Lett. 2018, 442, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Rother, E.; Brandl, R.; Baker, D.L.; Goyal, P.; Gebhard, H.; Tigyi, G.; Siess, W. Subtype-Selective Antagonists of Lysophosphatidic Acid Receptors Inhibit Platelet Activation Triggered by the Lipid Core of Atherosclerotic Plaques. Circulation 2003, 108, 741–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, E.S.; Heo, S.C.; Lee, I.H.; Choi, Y.J.; Park, J.H.; Choi, K.U.; Park, Y.; Suh, D.-S.; Yoon, M.-S.; Kim, J.H. Ovarian cancer-derived lysophosphatidic acid stimulates secretion of VEGF and stromal cell-derived factor-1α from human mesenchymal stem cells. Exp. Mol. Med. 2010, 42, 280–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chignard, M.; Le Couedic, J.P.; Tence, M.; Vargaftig, B.B.; Benveniste, J. The role of platelet-activating factor in platelet aggregation. Nature 1979, 279, 799–800. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.A.; Park, H.; Lim, E.H.; Kim, K.H.; Choi, J.S.; Lee, J.H.; Shin, J.W.; Lee, K.W. Exosomes from ovarian cancer cells induce adipose tissue-derived mesenchymal stem cells to acquire the physical and functional characteristics of tumor-supporting myofibroblasts. Gynecol. Oncol. 2011, 123, 379–386. [Google Scholar] [CrossRef] [PubMed]
- So, K.A.; Min, K.J.; Hong, J.H.; Lee, J.-K. Interleukin-6 expression by interactions between gynecologic cancer cells and human mesenchymal stem cells promotes epithelial-mesenchymal transition. Int. J. Oncol. 2015, 47, 1451–1459. [Google Scholar] [CrossRef] [Green Version]
- Gastl, G.; Plante, M.; Finstad, C.L.; Wong, G.Y.; Federici, M.G.; Bander, N.H.; Rubin, S.C. High IL-6 levels in ascitic fluid correlate with reactive thrombocytosis in patients with epithelial ovarian cancer. Br. J. Haematol. 1993, 83, 433–441. [Google Scholar] [CrossRef]
- Shield, K.; Ackland, M.L.; Ahmed, N.; Rice, G.E. Multicellular spheroids in ovarian cancer metastases: Biology and pathology. Gynecol. Oncol. 2009, 113, 143–148. [Google Scholar] [CrossRef]
- Rynne-Vidal, A.; Au-Yeung, C.L.; Jiménez-Heffernan, J.A.; Perez-Lozano, M.-L.; Cremades-Jimeno, L.; Bárcena, C.; Cristobal, I.; Fernández-Chacón, C.; Yeung, T.L.; Mok, S.C.; et al. Mesothelial-to-mesenchymal transition as a possible therapeutic target in peritoneal metastasis of ovarian cancer. J. Pathol. 2017, 242, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Matte, I.; Lane, D.; Laplante, C.; Rancourt, C.; Piché, A. Profiling of cytokines in human epithelial ovarian cancer ascites. Am. J. Cancer Res. 2012, 2, 566–580. [Google Scholar] [PubMed]
- Yan, D.; Liu, X.; Xu, H.; Guo, S.-W. Mesothelial Cells Participate in Endometriosis Fibrogenesis through Platelet-Induced Mesothelial-Mesenchymal Transition. J. Clin. Endocrinol. Metab. 2020, 105, e4124–e4147. [Google Scholar] [CrossRef] [PubMed]
- Steidel, C.; Ender, F.; Rody, A.; von Bubnoff, N.; Gieseler, F. Biologically Active Tissue Factor-Bearing Larger Ectosome-Like Extracellular Vesicles in Malignant Effusions from Ovarian Cancer Patients: Correlation with Incidence of Thrombosis. Int. J. Mol. Sci. 2021, 22, 790. [Google Scholar] [CrossRef]
- Orellana, R.; Kato, S.; Erices, R.; Bravo, M.L.; Gonzalez, P.; Oliva, B.; Cubillos, S.; Valdivia, A.; Ibañez, C.; Brañes, J.; et al. Platelets enhance tissue factor protein and metastasis initiating cell markers, and act as chemoattractants increasing the migration of ovarian cancer cells. BMC Cancer 2015, 15, 290. [Google Scholar] [CrossRef] [Green Version]
- Malacrida, B.; Nichols, S.; Maniati, E.; Jones, R.; Delanie-Smith, R.; Roozitalab, R.; Tyler, E.J.; Thomas, M.; Boot, G.; Mackerodt, J.; et al. A human multi-cellular model shows how platelets drive production of diseased extracellular matrix and tissue invasion. Iscience 2021, 24, 102676. [Google Scholar] [CrossRef] [PubMed]
- Motohara, T.; Masuda, K.; Morotti, M.; Zheng, Y.; El-Sahhar, S.; Chong, K.Y.; Wietek, N.; Alsaadi, A.; Carrami, E.M.; Hu, Z.; et al. An evolving story of the metastatic voyage of ovarian cancer cells: Cellular and molecular orchestration of the adipose-rich metastatic microenvironment. Oncogene 2018, 38, 2885–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryner, L.; Guan, Y.; Firestein, R.; Xiao, Y.; Choi, Y.; Rabe, C.; Lu, S.; Fuentes, E.; Huw, L.-Y.; Lackner, M.R.; et al. Upregulation of Periostin and Reactive Stroma Is Associated with Primary Chemoresistance and Predicts Clinical Outcomes in Epithelial Ovarian Cancer. Clin. Cancer Res. 2015, 21, 2941–2951. [Google Scholar] [CrossRef] [Green Version]
- Moran-Jones, K.; Gloss, B.S.; Murali, R.; Chang, D.K.; Colvin, E.K.; Jones, M.D.; Yuen, S.; Howell, V.M.; Brown, L.M.; Wong, C.W.; et al. Connective tissue growth factor as a novel therapeutic target in high grade serous ovarian cancer. Oncotarget 2015, 6, 44551–44562. [Google Scholar] [CrossRef]
- Yeung, T.-L.; Leung, C.S.; Wong, K.-K.; Samimi, G.; Thompson, M.S.; Liu, J.; Zaid, T.M.; Ghosh, S.; Birrer, M.J.; Mok, S.C. TGF-β Modulates Ovarian Cancer Invasion by Upregulating CAF-Derived Versican in the Tumor Microenvironment. Cancer Res. 2013, 73, 5016–5028. [Google Scholar] [CrossRef] [Green Version]
- Ghoneum, A.; Afify, H.; Salih, Z.; Kelly, M.; Said, N. Role of tumor microenvironment in ovarian cancer pathobiology. Oncotarget 2018, 9, 22832–22849. [Google Scholar] [CrossRef] [Green Version]
- Crawford, Y.; Kasman, I.; Yu, L.; Zhong, C.; Wu, X.; Modrusan, Z.; Kaminker, J.; Ferrara, N. PDGF-C Mediates the Angiogenic and Tumorigenic Properties of Fibroblasts Associated with Tumors Refractory to Anti-VEGF Treatment. Cancer Cell 2009, 15, 21–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wawro, M.E.; Sobierajska, K.; Ciszewski, W.M.; Niewiarowska, J. Nonsteroidal Anti-Inflammatory Drugs Prevent Vincristine-Dependent Cancer-Associated Fibroblasts Formation. Int. J. Mol. Sci. 2019, 20, 1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masucci, M.T.; Minopoli, M.; Carriero, M.V. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front. Oncol. 2019, 9, 1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmons, T.R.; Giridharan, T.; Singel, K.L.; Khan, A.N.H.; Ricciuti, J.; Howard, K.; Toro, S.L.S.-D.; Debreceni, I.L.; Aarts, C.E.; Brouwer, M.C.; et al. Mechanisms Driving Neutrophil-Induced T-cell Immunoparalysis in Ovarian Cancer. Cancer Immunol. Res. 2021, 9, 790–810. [Google Scholar] [CrossRef]
- Shang, A.; Wang, W.; Gu, C.; Chen, C.; Zeng, B.; Yang, Y.; Ji, P.; Sun, J.; Wu, J.; Lu, W.; et al. Long non-coding RNA HOTTIP enhances IL-6 expression to potentiate immune escape of ovarian cancer cells by upregulating the expression of PD-L1 in neutrophils. J. Exp. Clin. Cancer Res. 2019, 38, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanabuchi, S.; Watanabe, N.; Wang, Y.-H.; Wang, Y.-H.; Ito, T.; Shaw, J.; Cao, W.; Qin, F.X.-F.; Liu, Y.-J. Human plasmacytoid predendritic cells activate NK cells through glucocorticoid-induced tumor necrosis factor receptor-ligand (GITRL). Blood 2006, 107, 3617–3623. [Google Scholar] [CrossRef]
- Lines, J.L.; Pantazi, E.; Mak, J.; Sempere, L.F.; Wang, L.; O’Connell, S.; Ceeraz, S.; Suriawinata, A.A.; Yan, S.; Ernstoff, M.S.; et al. VISTA Is an Immune Checkpoint Molecule for Human T Cells. Cancer Res. 2014, 74, 1924–1932. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Li, L.; Wang, X.; Wang, C.; Wang, D. Neutrophil-to-Lymphocyte Ratio and Platelet Count Predict Long-Term Outcome of Stage IIIC Epithelial Ovarian Cancer. Cell. Physiol. Biochem. 2018, 46, 178–186. [Google Scholar] [CrossRef]
- Komura, N.; Mabuchi, S.; Yokoi, E.; Kozasa, K.; Kuroda, H.; Sasano, T.; Matsumoto, Y.; Kimura, T. Comparison of clinical utility between neutrophil count and neutrophil–lymphocyte ratio in patients with ovarian cancer: A single institutional experience and a literature review. Int. J. Clin. Oncol. 2017, 23, 104–113. [Google Scholar] [CrossRef]
- Williams, M.R.; Azcutia, V.; Newton, G.; Alcaide, P.; Luscinskas, F.W. Emerging mechanisms of neutrophil recruitment across endothelium. Trends Immunol. 2011, 32, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Larsen, E.; Celi, A.; Gilbert, G.E.; Furie, B.C.; Erban, J.K.; Bonfanti, R.; Wagner, D.D.; Furie, B. PADGEM protein: A receptor that mediates the interaction of activated platelets with neutrophils and monocytes. Cell 1989, 59, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.I.; Chen, Z.; Xu, H.; Li, C.Q.; Dong, J.-F.; McIntire, L.V.; Ballantyne, C.M.; Zhang, L.; Furman, M.I.; Berndt, M.C.; et al. Platelet Glycoprotein Ibα Is a Counterreceptor for the Leukocyte Integrin Mac-1 (Cd11b/Cd18). J. Exp. Med. 2000, 192, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuijper, P.H.M.; Torres, H.I.G.; Lammers, J.-W.J.; Sixma, J.J.; Koenderman, L.; Zwaginga, J.J. Platelet Associated Fibrinogen and ICAM-2 Induce Firm Adhesion of Neutrophils under Flow Conditions. Thromb. Haemost. 1998, 80, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, K.; Klink, M.; Wilczyński, J.R.; Szyłło, K.; Malinowski, A.; Sułowska, Z.; Nowak, M. Heterogeneity of the Mac-1 expression on peripheral blood neutrophils in patients with different types of epithelial ovarian cancer. Immunobiology 2016, 221, 323–332. [Google Scholar] [CrossRef]
- Klink, M.; Jastrzembska, K.; Nowak, M.; Bednarska, K.; Szpakowski, M.; Szyllo, K.; Sulowska, Z. Ovarian Cancer Cells Modulate Human Blood Neutrophils Response to Activation In Vitro. Scand. J. Immunol. 2008, 68, 328–336. [Google Scholar] [CrossRef]
- Stadtmann, A.; Germena, G.; Block, H.; Boras, M.; Rossaint, J.; Sundd, P.; Lefort, C.; Fisher, C.I.; Buscher, K.; Gelschefarth, B.; et al. The PSGL-1–L-selectin signaling complex regulates neutrophil adhesion under flow. J. Exp. Med. 2013, 210, 2171–2180. [Google Scholar] [CrossRef] [Green Version]
- Di Wu, Q.; Wang, J.H.; Condron, C.; Bouchier-Hayes, D.; Redmond, H.P. Human neutrophils facilitate tumor cell transendothelial migration. Am. J. Physiol. Physiol. 2001, 280, C814–C822. [Google Scholar] [CrossRef] [Green Version]
- Vanichakarn, P.; Blair, P.; Wu, C.; Freedman, J.E.; Chakrabarti, S. Neutrophil CD40 enhances platelet-mediated inflammation. Thromb. Res. 2008, 122, 346–358. [Google Scholar] [CrossRef]
- Frenette, P.S.; Denis, C.V.; Weiss, L.; Jurk, K.; Subbarao, S.; Kehrel, B.; Hartwig, J.H.; Vestweber, D.; Wagner, D.D. P-Selectin Glycoprotein Ligand 1 (Psgl-1) Is Expressed on Platelets and Can Mediate Platelet–Endothelial Interactions in Vivo. J. Exp. Med. 2000, 191, 1413–1422. [Google Scholar] [CrossRef] [Green Version]
- Weber, C.; Springer, A.T. Neutrophil accumulation on activated, surface-adherent platelets in flow is mediated by interaction of Mac-1 with fibrinogen bound to alphaIIbbeta3 and stimulated by platelet-activating factor. J. Clin. Investig. 1997, 100, 2085–2093. [Google Scholar] [CrossRef]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil pheno-type by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Zhang, G.; Wang, Y.; He, M.; Xu, Q.; Lu, J.; Liu, H.; Xu, C. Tumour-associated neutrophils orchestrate intratumoural IL-8-driven immune evasion through Jagged2 activation in ovarian cancer. Br. J. Cancer 2020, 123, 1404–1416. [Google Scholar] [CrossRef]
- Singh, A.; Bisht, P.; Bhattacharya, S.; Guchhait, P. Role of Platelet Cytokines in Dengue Virus Infection. Front. Cell. Infect. Microbiol. 2020, 10, 561366. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.E.; Ceglowski, J.R.; Roweth, H.G.; Forward, J.A.; Tippy, M.D.; El-Husayni, S.; Kulenthirarajan, R.; Malloy, M.W.; Machlus, K.R.; Chen, W.Y.; et al. Aspirin inhibits platelets from reprogramming breast tumor cells and promoting metastasis. Blood Adv. 2019, 3, 198–211. [Google Scholar] [CrossRef] [Green Version]
- Micheli, D.C.; Jammal, M.P.; Martins-Filho, A.; Côrtes, J.R.X.D.M.; de Souza, C.N.; Nomelini, R.S.; Murta, E.F.C.; Tavares-Murta, B.M. Serum cytokines and CXCR2: Potential tumour markers in ovarian neoplasms. Biomarkers 2020, 25, 474–482. [Google Scholar] [CrossRef]
- Rodrigues, I.S.S.; Martins-Filho, A.; Micheli, D.C.; de Lima, C.A.; Tavares-Murta, B.M.; Murta, E.F.C.; Nomelini, R.S. IL-6 and IL-8 as Prognostic Factors in Peritoneal Fluid of Ovarian Cancer. Immunol. Investig. 2019, 49, 510–521. [Google Scholar] [CrossRef]
- Coffelt, S.B.; Wellenstein, M.D.; de Visser, K.E. Neutrophils in cancer: Neutral no more. Nat. Rev. Cancer 2016, 16, 431–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metelli, A.; Wu, B.X.; Riesenberg, B.; Guglietta, S.; Huck, J.D.; Mills, C.; Li, A.; Rachidi, S.; Krieg, C.; Rubinstein, M.P.; et al. Thrombin contributes to cancer immune evasion via proteolysis of platelet-bound GARP to activate LTGF-β. Sci. Transl. Med. 2020, 12, eaay4860. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2017, 18, 134–147. [Google Scholar] [CrossRef]
- Elaskalani, O.; Abdol Razak, N.B.; Metharom, P. Neutrophil extracellular traps induce aggregation of washed human platelets independently of extracellular DNA and histones. Cell Commun. Signal. 2018, 16, 24. [Google Scholar] [CrossRef] [Green Version]
- Etulain, J.; Martinod, K.; Wong, S.L.; Cifuni, S.M.; Schattner, M.; Wagner, D.D. P-selectin promotes neutrophil extracellular trap formation in mice. Blood 2015, 126, 242–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, C.; Darb-Esfahani, S.; Meyer, A.-S.; Hübner, K.; Rom, J.; Sohn, C.; Braicu, I.; Sehouli, J.; Hänsch, G.M.; Gaida, M.M. Neutrophil Granulocytes in Ovarian Cancer—Induction of Epithelial-To-Mesenchymal-Transition and Tumor Cell Migration. J. Cancer 2016, 7, 546–554. [Google Scholar] [CrossRef]
- Si-Tahar, M.; Pidard, D.; Balloy, V.; Moniatte, M.; Kieffer, N.; van Dorsselaer, A.; Chignard, M. Human Neutrophil Elastase Proteolytically Activates the Platelet Integrin αIIbβ3 through Cleavage of the Carboxyl Terminus of the αIIb Subunit Heavy Chain. J. Biol. Chem. 1997, 272, 11636–11647. [Google Scholar] [CrossRef] [Green Version]
- Itagaki, K.; Kaczmarek, E.; Lee, Y.T.; Tang, I.T.; Isal, B.; Adibnia, Y.; Sandler, N.; Grimm, M.J.; Segal, B.H.; Otterbein, L.E.; et al. Mitochondrial DNA Released by Trauma Induces Neutrophil Extracellular Traps. PLoS ONE 2015, 10, e0120549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demers, M.; Krause, D.S.; Schatzberg, D.; Martinod, K.; Voorhees, J.R.; Fuchs, T.A.; Scadden, D.T.; Wagner, D.D. Cancers predispose neutrophils to release extracellular DNA traps that contribute to cancer-associated thrombosis. Proc. Natl. Acad. Sci. USA 2012, 109, 13076–13081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Ko, S.Y.; Mohamed, M.S.; Kenny, H.A.; Lengyel, E.; Naora, H. Neutrophils facilitate ovarian cancer premetastatic niche formation in the omentum. J. Exp. Med. 2018, 216, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Singel, K.; Grzankowski, K.S.; Khan, A.N.M.N.H.; Grimm, M.J.; D’Auria, A.C.; Morrell, K.; Eng, K.H.; Hylander, B.; Mayor, P.C.; Emmons, T.R.; et al. Mitochondrial DNA in the tumour microenvironment activates neutrophils and is associated with worse outcomes in patients with advanced epithelial ovarian cancer. Br. J. Cancer 2018, 120, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Ma, Z.; Wang, R.; Wang, Y.; Wang, S.; Cheng, Z.; Xu, H.; Jin, X.; Li, W.; Wang, X. Thrombin facilitates invasion of ovarian cancer along peritoneum by inducing monocyte differentiation toward tumor-associated macrophage-like cells. Cancer Immunol. Immunother. 2010, 59, 1097–1108. [Google Scholar] [CrossRef]
- Duan, Z.; Luo, Y. Targeting macrophages in cancer immunotherapy. Signal Transduct. Target. Ther. 2021, 6, 127. [Google Scholar] [CrossRef]
- Reinartz, S.; Schumann, T.; Finkernagel, F.; Wortmann, A.; Jansen, J.M.; Meissner, W.; Krause, M.; Schwörer, A.; Wagner, U.; Müller-Brüsselbach, S.; et al. Mixed-polarization phenotype of ascites-associated macrophages in human ovarian carcinoma: Correlation of CD163 expression, cytokine levels and early relapse. Int. J. Cancer 2013, 134, 32–42. [Google Scholar] [CrossRef]
- Kawamura, K.; Komohara, Y.; Takaishi, K.; Katabuchi, H.; Takeya, M. Detection of M2 macrophages and colony-stimulating factor 1 expression in serous and mucinous ovarian epithelial tumors. Pathol. Int. 2009, 59, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Yull, F.; Khabele, D. Bipolar Tumor-Associated Macrophages in Ovarian Cancer as Targets for Therapy. Cancers 2018, 10, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, M.; Shen, J.; Yu, S.; Fei, J.; Zhu, X.; Zhao, J.; Zhai, L.; Sadhukhan, A.; Zhou, J. Tumor-Associated Macrophages (TAMs): A Critical Activator in Ovarian Cancer Metastasis. OncoTargets Ther. 2019, 12, 8687–8699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burleson, K.M.; Casey, R.C.; Skubitz, K.M.; Pambuccian, S.E.; Oegema, T.R., Jr.; Skubitz, A.P.N. Ovarian carcinoma ascites spheroids adhere to extracellular matrix components and mesothelial cell monolayers. Gynecol. Oncol. 2004, 93, 170–181. [Google Scholar] [CrossRef]
- Long, L.; Hu, Y.; Long, T.; Lu, X.; Tuo, Y.; Li, Y.; Wang, M.; Ke, Z. Tumor-associated macrophages induced spheroid formation by CCL18-ZEB1-M-CSF feedback loop to promote transcoelomic metastasis of ovarian cancer. J. Immunother. Cancer 2021, 9, e003973. [Google Scholar] [CrossRef]
- Zhang, M.; He, Y.; Sun, X.; Li, Q.; Wang, W.; Zhao, A.; Di, W. A high M1/M2 ratio of tumor-associated macrophages is associated with extended survival in ovarian cancer patients. J. Ovarian Res. 2014, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Michelson, A.D.; Barnard, M.R.; Krueger, L.A.; Valeri, C.R.; Furman, M.I. Circulating Monocyte-Platelet Aggregates Are a More Sensitive Marker of In Vivo Platelet Activation Than Platelet Surface P-Selectin. Circulation 2001, 104, 1533–1537. [Google Scholar] [CrossRef] [Green Version]
- Duque, G.A.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [Green Version]
- Nagata, K.; Tsuji, T.; Todoroki, N.; Katagiri, Y.; Tanoue, K.; Yamazaki, H.; Hanai, N.; Irimura, T. Activated platelets induce superoxide anion release by monocytes and neutrophils through P-selectin (CD62). J. Immunol. 1993, 151, 3267–3273. [Google Scholar] [CrossRef]
- Griess, B.; Mir, S.; Datta, K.; Teoh-Fitzgerald, M. Scavenging reactive oxygen species selectively inhibits M2 macrophage polarization and their pro-tumorigenic function in part, via Stat3 suppression. Free Radic. Biol. Med. 2019, 147, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Yoon, B.R.; Kim, H.Y.; Yoo, S.-J.; Kang, S.W.; Lee, W.-W. Activated Platelets Convert CD14+CD16- Into CD14+CD16+ Monocytes with Enhanced FcγR-Mediated Phagocytosis and Skewed M2 Polarization. Front. Immunol. 2021, 11, 611133. [Google Scholar] [CrossRef]
- Reader, J.; Holt, D.; Fulton, A. Prostaglandin E2 EP receptors as therapeutic targets in breast cancer. Cancer Metastasis Rev. 2011, 30, 449–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimp, A.H.; Hollema, H.; Kempinga, C.; van der Zee, A.G.; de Vries, E.G.; Daemen, T. Expression of cyclooxygenase-2 and inducible nitric oxide synthase in human ovarian tumors and tumor-associated macrophages. Cancer Res. 2001, 61, 7305–7309. [Google Scholar] [PubMed]
- Dijkgraaf, E.M.; Heusinkveld, M.; Tummers, B.; Vogelpoel, L.T.; Goedemans, R.; Jha, V.; Nortier, J.W.; Welters, M.J.; Kroep, J.R.; van der Burg, S.H. Chemotherapy Alters Monocyte Differentiation to Favor Generation of Cancer-Supporting M2 Macrophages in the Tumor Microenvironment. Cancer Res. 2013, 73, 2480–2492. [Google Scholar] [CrossRef] [Green Version]
- Mussbacher, M.; Salzmann, M.; Brostjan, C.; Hoesel, B.; Schoergenhofer, C.; Datler, H.; Hohensinner, P.; Basílio, J.; Petzelbauer, P.; Assinger, A.; et al. Cell Type-Specific Roles of NF-κB Linking Inflammation and Thrombosis. Front. Immunol. 2019, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Suzuki-Inoue, K. Platelets and cancer-associated thrombosis: Focusing on the platelet activation receptor CLEC-2 and podoplanin. Blood 2019, 134, 1912–1918. [Google Scholar] [CrossRef]
- Laffont, B.; Corduan, A.; Rousseau, M.; Duchez, A.C.; Lee, C.H.C.; Boilard, E.; Provost, P. Platelet microparticles reprogram macrophage gene expression and function. Thromb. Haemost. 2016, 115, 311–323. [Google Scholar] [CrossRef]
- Olingy, C.E.; Dinh, H.Q.; Hedrick, C.C. Monocyte heterogeneity and functions in cancer. J. Leukoc. Biol. 2019, 106, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Deng, C.; Wu, S.; Zhang, L.; Yang, M.; Lin, Q.; Xie, Q.; Ding, H.; Lian, N.; Gao, S.; Huang, Y.; et al. Role of monocyte tissue factor on patients with non-small cell lung cancer. Clin. Respir. J. 2017, 12, 1125–1133. [Google Scholar] [CrossRef]
- Hell, L.; Däullary, T.; Burghart, V.; Mauracher, L.-M.; Grilz, E.; Moser, B.; Kramer, G.; Schmid, J.; Ay, C.; Pabinger, I.; et al. Extracellular Vesicle-Associated Tissue Factor Activity in Prostate Cancer Patients with Disseminated Intravascular Coagulation. Cancers 2021, 13, 1487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, Z.; Zhou, P.; Yu, M.; Li, B.; Liu, Y.; Jin, J.; Liu, W.; Jing, H.; Du, J.; et al. Phosphatidylserine-exposing tumor-derived microparticles exacerbate coagulation and cancer cell transendothelial migration in triple-negative breast cancer. Theranostics 2021, 11, 6445–6460. [Google Scholar] [CrossRef] [PubMed]
- Montoro-García, S.; Shantsila, E.; Hernández-Romero, D.; Jover, E.; Valdés, M.; Marin, F.; Lip, G.Y.H. Small-size platelet microparticles trigger platelet and monocyte functionality and modulate thrombogenesis via P-selectin. Br. J. Haematol. 2014, 166, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Vasina, E.M.; Cauwenberghs, S.; Feijge, M.A.H.; Heemskerk, J.W.M.; Weber, C.; Koenen, R.R. Microparticles from apoptotic platelets promote resident macrophage differentiation. Cell Death Dis. 2011, 2, e211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ádány, R.; Kappelmayer, J.; Berényi, E.; Szegedi, A.; Fábián, E.; Muszbek, L. Factors of the Extrinsic Pathway of Blood Coagulation in Tumour Associated Macrophages. Thromb. Haemost. 1989, 62, 850–855. [Google Scholar] [CrossRef]
- Ruf, W.; Mueller, B. Thrombin Generation and the Pathogenesis of Cancer. Semin. Thromb. Hemost. 2006, 32, 061–068. [Google Scholar] [CrossRef]
- Chanakira, A.; Westmark, P.R.; Ong, I.M.; Sheehan, J.P. Tissue factor-factor VIIa complex triggers protease activated receptor 2-dependent growth factor release and migration in ovarian cancer. Gynecol. Oncol. 2017, 145, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Gil-Bernabé, A.M.; Ferjančič, Š.; Tlalka, M.; Zhao, L.; Allen, P.D.; Im, J.H.; Watson, K.; Hill, S.A.; Amirkhosravi, A.; Francis, J.L.; et al. Recruitment of monocytes/macrophages by tissue factor-mediated coagulation is essential for metastatic cell survival and premetastatic niche establishment in mice. Blood 2012, 119, 3164–3175. [Google Scholar] [CrossRef] [Green Version]
- Läubli, H.; Spanaus, K.-S.; Borsig, L. Selectin-mediated activation of endothelial cells induces expression of CCL5 and promotes metastasis through recruitment of monocytes. Blood 2009, 114, 4583–4591. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, M.Z.; Zuba-Surma, E.; Kucia, M.; Reca, R.; Wojakowski, W.; Ratajczak, J. The pleiotropic effects of the SDF-1–CXCR4 axis in organogenesis, regeneration and tumorigenesis. Leukemia 2006, 20, 1915–1924. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Martín, L.; Estecha, A.; Samaniego, R.; Sánchez-Ramón, S.; Vega, M.Á.; Sánchez-Mateos, P. The chemokine CXCL12 regulates monocyte-macrophage differentiation and RUNX3 expression. Blood 2011, 117, 88–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Li, B.; Liang, Y.; Reeves, P.M.; Qu, X.; Ran, C.; Liu, Q.; Callahan, M.V.; Sluder, A.E.; Gelfand, J.A.; et al. Dual blockade of CXCL12-CXCR4 and PD-1–PD-L1 pathways prolongs survival of ovarian tumor–bearing mice by prevention of immunosuppression in the tumor microenvironment. FASEB J. 2019, 33, 6596–6608. [Google Scholar] [CrossRef]
- Junior, I.A.D.S.; Stone, S.C.; Rossetti, R.M.; Jancar, S.; Lepique, A.P. Modulation of Tumor-Associated Macrophages (TAM) Phenotype by Platelet-Activating Factor (PAF) Receptor. J. Immunol. Res. 2017, 2017, 5482768. [Google Scholar] [CrossRef] [Green Version]
- Deuster, E.; Hysenaj, I.; Kahaly, M.; Schmoeckel, E.; Mayr, D.; Beyer, S.; Kolben, T.; Hester, A.; Kraus, F.; Chelariu-Raicu, A.; et al. The Platelet-Activating Factor Receptor’s Association with the Outcome of Ovarian Cancer Patients and Its Experimental Inhibition by Rupatadine. Cells 2021, 10, 2337. [Google Scholar] [CrossRef]
- Spiel, A.; Bartko, J.; Schwameis, M.; Firbas, C.; Siller-Matula, J.M.; Schuetz, M.; Weigl, M.; Jilma, B. Increased platelet aggregation and in vivo platelet activation after granulocyte colony-stimulating factor administration. Thromb. Haemost. 2011, 105, 655–662. [Google Scholar] [CrossRef]
- Zeng, X.-Y.; Xie, H.; Yuan, J.; Jiang, X.-Y.; Yong, J.-H.; Zeng, D.; Dou, Y.-Y.; Xiao, S.-S. M2-like tumor-associated macrophages-secreted EGF promotes epithelial ovarian cancer metastasis via activating EGFR-ERK signaling and suppressing lncRNA LIMT expression. Cancer Biol. Ther. 2019, 20, 956–966. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Zhang, M.; Zhang, X.; Cai, Q.; Hong, S.; Jiang, W.; Xu, C. Synergistic effects of combined platelet-activating factor receptor and epidermal growth factor receptor targeting in ovarian cancer cells. J. Hematol. Oncol. 2014, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Reinartz, S.; Lieber, S.; Pesek, J.; Brandt, D.T.; Asafova, A.; Finkernagel, F.; Watzer, B.; Nockher, W.A.; Nist, A.; Stiewe, T.; et al. Cell type-selective pathways and clinical associations of lysophosphatidic acid biosynthesis and signaling in the ovarian cancer microenvironment. Mol. Oncol. 2018, 13, 185–201. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, R.; Lee, S.-C.; David, M.; Bordet, J.-C.; Norman, D.D.; Patil, R.; Miller, D.; Sahay, D.; Ribeiro, J.; Clézardin, P.; et al. Interaction of platelet-derived autotaxin with tumor integrin αVβ3 controls metastasis of breast cancer cells to bone. Blood 2014, 124, 3141–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Xiao, M.; Zhang, Z.; Cui, R.; Jiang, X.; Wang, S.; Bai, H.; Liu, C.; Zhang, Z. Potential interaction between lysophosphatidic acid and tumor-associated macrophages in ovarian carcinoma. J. Inflamm. 2020, 17, 23. [Google Scholar] [CrossRef]
- Onallah, H.; Catane, L.J.; Tropé, C.G.; Falkenthal, T.E.H.; Reich, R.; Davidson, B. Activity and clinical relevance of autotaxin and lysophosphatidic acid pathways in high-grade serous carcinoma. Virchows Arch. 2018, 473, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Khandoga, A.L.; Fujiwara, Y.; Goyal, P.; Pandey, D.; Tsukahara, R.; Bolen, A.; Guo, H.; Wilke, N.; Liu, J.; Valentine, W.J.; et al. Lysophosphatidic acid-induced platelet shape change revealed through LPA1–5 receptor-selective probes and albumin. Platelets 2008, 19, 415–427. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.C.; Davis, R.R.; Said, N.; Hollis, T.; Daniel, L.W. Blocking LPA-dependent signaling increases ovarian cancer cell death in response to chemotherapy. Redox Biol. 2018, 15, 380–386. [Google Scholar] [CrossRef]
- Wang, W.; Zou, W.; Liu, J.R. Tumor-infiltrating T cells in epithelial ovarian cancer: Predictors of prognosis and biological basis of immunotherapy. Gynecol. Oncol. 2018, 151, 1–3. [Google Scholar] [CrossRef]
- Zhang, L.; Conejo-Garcia, J.R.; Katsaros, D.; Gimotty, P.A.; Massobrio, M.; Regnani, G.; Makrigiannakis, A.; Gray, H.; Schlienger, K.; Liebman, M.N.; et al. Intratumoral T Cells, Recurrence, and Survival in Epithelial Ovarian Cancer. N. Engl. J. Med. 2003, 348, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.P.; Balmaceda, C.; Bravo, M.L.; Kato, S.; Villarroel, A.; Owen, G.I.; Roa, J.C.; Cuello, M.A.; Ibañez, C. Patient inflammatory status and CD4+/CD8+ intraepithelial tumor lymphocyte infiltration are predictors of outcomes in high-grade serous ovarian cancer. Gynecol. Oncol. 2018, 151, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Starossom, S.C.; Veremeyko, T.; Yung, A.W.; Dukhinova, M.; Au, C.; Lau, A.Y.; Weiner, H.L.; Ponomarev, E.D. Platelets Play Differential Role During the Initiation and Progression of Autoimmune Neuroinflammation. Circ. Res. 2015, 117, 779–792. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, N.; Zhu, L.; Ersoy, M.; Hermansson, A.; Hjemdahl, P.; Hu, H.; Hansson, G.K.; Li, N. Platelets regulate CD4+ T-cell differentiation via multiple chemokines in humans. Thromb. Haemost. 2011, 106, 353–362. [Google Scholar] [CrossRef]
- Eken, C.; Sadallah, S.; Amicarella, F.; Iezzi, G.; Schifferli, J.A. Ectosomes released by platelets induce differentiation of CD4+ T cells into T regulatory cells. Thromb. Haemost. 2014, 112, 1219–1229. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Hu, W.; Song, X.; Zhao, Y. Immune Modulation of Platelet-Derived Mitochondria on Memory CD4+ T Cells in Humans. Int. J. Mol. Sci. 2020, 21, 6295. [Google Scholar] [CrossRef]
- Vitkovsky, Y.; Brill, G.; Koltakov, A.; Farzam, N.; Varon, D.; Bank, I.; Solpov, A.; Shenkman, B.; Savion, N. Platelets enhance CD4+ lymphocyte adhesion to extracellular matrix under flow conditions: Role of platelet aggregation, integrins, and non-integrin receptors. Thromb. Haemost. 2006, 95, 815–821. [Google Scholar] [CrossRef]
- Shenkman, B.; Brill, G.; Solpov, A.; Vitkovsky, Y.; Kuznik, B.; Koltakov, A.; Kotev-Emeth, S.; Savion, N.; Bank, I. CD4+ lymphocytes require platelets for adhesion to immobilized fibronectin in flow: Role of β1 (CD29)-, β2 (CD18)-related integrins and non-integrin receptors. Cell. Immunol. 2006, 242, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; de la Motte, C.; Reyes, B.M.R.; Sans, M.; Levine, A.D.; Fiocchi, C. Cutting Edge: T Cells Trigger CD40-Dependent Platelet Activation and Granular RANTES Release: A Novel Pathway for Immune Response Amplification. J. Immunol. 2004, 172, 2011–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, P.J.; Riley, J.L.; Harlan, D.M.; Abe, R.; Tadaki, D.K.; Hoffmann, S.C.; White, L.; Francomano, T.; Perfetto, S.J.; Kirk, A.D.; et al. Cd40 Ligand (Cd154) Triggers a Short-Term Cd4+ T Cell Activation Response That Results in Secretion of Immunomodulatory Cytokines and Apoptosis. J. Exp. Med. 2000, 191, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Salas-Perdomo, A.; Miró-Mur, F.; Urra, X.; Justicia, C.; Gallizioli, M.; Zhao, Y.; Brait, V.H.; Laredo, C.; Tudela, R.; Hidalgo, A.; et al. T Cells Prevent Hemorrhagic Transformation in Ischemic Stroke by P-Selectin Binding. Arter. Thromb. Vasc. Biol. 2018, 38, 1761–1771. [Google Scholar] [CrossRef] [Green Version]
- Hirata, T.; Merrill-Skoloff, G.; Aab, M.; Yang, J.; Furie, B.C.; Furie, B. P-Selectin Glycoprotein Ligand 1 (Psgl-1) Is a Physiological Ligand for E-Selectin in Mediating T Helper 1 Lymphocyte Migration. J. Exp. Med. 2000, 192, 1669–1676. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, R.; Carrette, F.; Barraza, M.L.; Otero, D.C.; Magaña, J.; Bosenberg, M.W.; Swain, S.L.; Bradley, L.M. PSGL-1 Is an Immune Checkpoint Regulator that Promotes T Cell Exhaustion. Immunity 2016, 44, 1190–1203. [Google Scholar] [CrossRef] [Green Version]
- Zaslavsky, A.B.; Adams, M.P.; Cao, X.; Maj, T.; Choi, J.E.; Stangl-Kremser, J.; Patel, S.; Putelo, A.; Lee, S.K.; Nallandhighal, S.; et al. Platelet PD-L1 suppresses anti-cancer immune cell activity in PD-L1 negative tumors. Sci. Rep. 2020, 10, 19296. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, B.; Zai, W.; Kang, L.; Yuan, A.; Hu, Y.; Wu, J. Perfluorocarbon nanoparticle-mediated platelet inhibition promotes intratumoral infiltration of T cells and boosts immunotherapy. Proc. Natl. Acad. Sci. USA 2019, 116, 11972–11977. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Malloy, M.W.; Roweth, H.G.; McAllister, S.S.; Italiano, J.J.E.; Battinelli, E.M. Platelets upregulate tumor cell programmed death ligand 1 in an epidermal growth factor receptor-dependent manner in vitro. Blood Adv. 2022, 6, 5668–5675. [Google Scholar] [CrossRef]
- Lutz, M.S.; Klimovich, B.; Maurer, S.; Heitmann, J.S.; Märklin, M.; Zekri, L.; Jung, G.; Salih, H.R.; Hinterleitner, C. Platelets subvert antitumor efficacy of T cell-recruiting bispecific antibodies. J. Immunother. Cancer 2022, 10, e003655. [Google Scholar] [CrossRef] [PubMed]
- Crawford, A.; Haber, L.; Kelly, M.P.; Vazzana, K.; Canova, L.; Ram, P.; Pawashe, A.; Finney, J.; Jalal, S.; Chiu, D.; et al. A Mucin 16 bispecific T cell–engaging antibody for the treatment of ovarian cancer. Sci. Transl. Med. 2019, 11, eaau7534. [Google Scholar] [CrossRef] [PubMed]
- Yeku, O.O.; Rao, T.D.; Laster, I.; Kononenko, A.; Purdon, T.J.; Wang, P.; Cui, Z.; Liu, H.; Brentjens, R.J.; Spriggs, D. Bispecific T-Cell Engaging Antibodies Against MUC16 Demonstrate Efficacy Against Ovarian Cancer in Monotherapy and in Combination With PD-1 and VEGF Inhibition. Front. Immunol. 2021, 12, 663379. [Google Scholar] [CrossRef] [PubMed]
- Raffin, C.; Vo, L.T.; Bluestone, J.A. Treg cell-based therapies: Challenges and perspectives. Nat. Rev. Immunol. 2019, 20, 158–172. [Google Scholar] [CrossRef]
- Fialová, A.; Partlová, S.; Sojka, L.; Hromádková, H.; Brtnický, T.; Fučíková, J.; Kocián, P.; Rob, L.; Bartůňková, J.; Špíšek, R. Dynamics of T-cell infiltration during the course of ovarian cancer: The gradual shift from a Th17 effector cell response to a predominant infiltration by regulatory T-cells. Int. J. Cancer 2012, 132, 1070–1079. [Google Scholar] [CrossRef]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.-P.; Gimotty, P.A.; Gilks, C.B.; Lal, P.; Zhang, L.; et al. Tumour hypoxia promotes tolerance and angiogenesis via CCL28 and Treg cells. Nature 2011, 475, 226–230. [Google Scholar] [CrossRef]
- Giuntoli, R.L., 2nd; Webb, T.J.; Zoso, A.; Rogers, O.; Diaz-Montes, T.P.; Bristow, R.E.; Oelke, M. Ovarian Cancer-Associated Ascites Demonstrates Altered Immune Environment: Implications for Antitumor Immunity. Anticancer. Res. 2009, 29, 2875–2884. [Google Scholar]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar] [CrossRef]
- Knutson, K.L.; Maurer, M.J.; Preston, C.C.; Moysich, K.B.; Goergen, K.; Hawthorne, K.M.; Cunningham, J.M.; Odunsi, K.; Hartmann, L.C.; Kalli, K.R.; et al. Regulatory T cells, inherited variation, and clinical outcome in epithelial ovarian cancer. Cancer Immunol. Immunother. 2015, 64, 1495–1504. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Yan, C.; Xu, D.; Zhang, Z.; Li, K.; Li, X.; Zhou, M.; Hao, D. Immuno-genomic characterisation of high-grade serous ovarian cancer reveals immune evasion mechanisms and identifies an immunological subtype with a favourable prognosis and improved therapeutic efficacy. Br. J. Cancer 2022, 126, 1570–1580. [Google Scholar] [CrossRef]
- Angiari, S.; Rossi, B.; Piccio, L.; Zinselmeyer, B.H.; Budui, S.; Zenaro, E.; della Bianca, V.; Bach, S.D.; Scarpini, E.; Bolomini-Vittori, M.; et al. Regulatory T Cells Suppress the Late Phase of the Immune Response in Lymph Nodes through P-Selectin Glycoprotein Ligand-1. J. Immunol. 2013, 191, 5489–5500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohm, A.P.; Miller, S.D. Role of ICAM-1 and P-selectin expression in the development and effector function of CD4+CD25+regulatory T cells. J. Autoimmun. 2003, 21, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Urzainqui, A.; Serrador, J.M.; Viedma, F.; Yáñez-Mó, M.; Rodríguez, A.; Corbí, A.L.; Alonso-Lebrero, J.L.; Luque, A.; Deckert, M.; Vázquez, J.; et al. ITAM-Based Interaction of ERM Proteins with Syk Mediates Signaling by the Leukocyte Adhesion Receptor PSGL-1. Immunity 2002, 17, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Rupp, M.-C.; Bergmann, C.B.; Jung, S.; Bock, M.; Biberthaler, P.; Heimann, L.; Hanschen, M. The posttraumatic response of CD4+ regulatory T cells is modulated by direct cell-cell contact via CD40L- and P-selectin-dependent pathways. Central Eur. J. Immunol. 2021, 46, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Guiducci, C.; Valzasina, B.; Dislich, H.; Colombo, M.P. CD40/CD40L interaction regulates CD4+CD25+ T reg homeostasis through dendritic cell-produced IL-2. Eur. J. Immunol. 2005, 35, 557–567. [Google Scholar] [CrossRef]
- Renshaw, B.R.; Fanslow, W.C.; Armitage, R.J.; Campbell, K.A.; Liggitt, D.; Wright, B.; Davison, B.L.; Maliszewski, C.R. Humoral immune responses in CD40 ligand-deficient mice. J. Exp. Med. 1994, 180, 1889–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; Pan, J.; Setiadi, H.; Pate, K.D.; McEver, R.P. Interleukin 4 or oncostatin M induces a prolonged increase in P-selectin mRNA and protein in human endothelial cells. J. Exp. Med. 1996, 184, 81–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legein, B.; Temmerman, L.; Biessen, E.A.L.; Lutgens, E. Inflammation and immune system interactions in atherosclerosis. Cell. Mol. Life Sci. 2013, 70, 3847–3869. [Google Scholar] [CrossRef]
- Kampan, N.C.; Madondo, M.T.; McNally, O.M.; Stephens, A.N.; Quinn, M.A.; Plebanski, M. Interleukin 6 Present in Inflammatory Ascites from Advanced Epithelial Ovarian Cancer Patients Promotes Tumor Necrosis Factor Receptor 2-Expressing Regulatory T Cells. Front. Immunol. 2017, 8, 1482. [Google Scholar] [CrossRef]
- Torrey, H.; Butterworth, J.; Mera, T.; Okubo, Y.; Wang, L.; Baum, D.; Defusco, A.; Plager, S.; Warden, S.; Huang, D.; et al. Targeting TNFR2 with antagonistic antibodies inhibits proliferation of ovarian cancer cells and tumor-associated T regs. Sci. Signal. 2017, 10, eaaf8608. [Google Scholar] [CrossRef]
- Haribhai, D.; Luo, X.; Chen, J.; Jia, S.; Shi, L.; Schroeder, J.A.; Weiler, H.; Aster, R.H.; Hessner, M.J.; Hu, J.; et al. TGF-β1 along with other platelet contents augments Treg cells to suppress anti-FVIII immune responses in hemophilia A mice. Blood Adv. 2016, 1, 139–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, N.; Krebs, F.K.; Zimmer, S.; Mitzel-Rink, H.; Kumm, E.J.; Jurk, K.; Grabbe, S.; Loquai, C.; Tuettenberg, A. Platelet-Derived GARP Induces Peripheral Regulatory T Cells—Potential Impact on T Cell Suppression in Patients with Melanoma-Associated Thrombocytosis. Cancers 2020, 12, 3653. [Google Scholar] [CrossRef] [PubMed]
- Re, S.L.; Lecocq, M.; Uwambayinema, F.; Yakoub, Y.; Delos, M.; Demoulin, J.-B.; Lucas, S.; Sparwasser, T.; Renauld, J.-C.; Lison, D.; et al. Platelet-Derived Growth Factor–Producing CD4+ Foxp3+ Regulatory T Lymphocytes Promote Lung Fibrosis. Am. J. Respir. Crit. Care Med. 2011, 184, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Roane, B.M.; Meza-Perez, S.; Katre, A.A.; Goldsberry, W.N.; Randall, T.D.; Norian, L.A.; Birrer, M.J.; Arend, R.C. Neutralization of TGFβ Improves Tumor Immunity and Reduces Tumor Progression in Ovarian Carcinoma. Mol. Cancer Ther. 2021, 20, 602–611. [Google Scholar] [CrossRef]
- Yahya, M.A.; Sharon, S.M.; Hantisteanu, S.; Hallak, M.; Bruchim, I. The Role of the Insulin-Like Growth Factor 1 Pathway in Immune Tumor Microenvironment and Its Clinical Ramifications in Gynecologic Malignancies. Front. Endocrinol. 2018, 9, 297. [Google Scholar] [CrossRef]
- Camorani, S.; Passariello, M.; Agnello, L.; Esposito, S.; Collina, F.; Cantile, M.; di Bonito, M.; Ulasov, I.V.; Fedele, M.; Zannetti, A.; et al. Aptamer targeted therapy potentiates immune checkpoint blockade in triple-negative breast cancer. J. Exp. Clin. Cancer Res. 2020, 39, 180. [Google Scholar] [CrossRef]
- Liu, C.Y.; Battaglia, M.; Lee, S.H.; Sun, Q.-H.; Aster, R.H.; Visentin, G.P. Platelet Factor 4 Differentially Modulates CD4+CD25+ (Regulatory) versus CD4+CD25− (Nonregulatory) T Cells. J. Immunol. 2005, 174, 2680–2686. [Google Scholar] [CrossRef] [Green Version]
- Kabashima, K.; Murata, T.; Tanaka, H.; Matsuoka, T.; Sakata, D.; Yoshida, N.; Katagiri, K.; Kinashi, T.; Tanaka, T.; Miyasaka, M.; et al. Thromboxane A2 modulates interaction of dendritic cells and T cells and regulates acquired immunity. Nat. Immunol. 2003, 4, 694–701. [Google Scholar] [CrossRef]
- von Hundelshausen, P.; Agten, S.M.; Eckardt, V.; Blanchet, X.; Schmitt, M.M.; Ippel, H.; Neideck, C.; Bidzhekov, K.; Leberzammer, J.; Wichapong, K.; et al. Chemokine interactome mapping enables tailored intervention in acute and chronic inflammation. Sci. Transl. Med. 2017, 9, eaah6650. [Google Scholar] [CrossRef]
- Tan, S.; Zhang, J.; Sun, Y.; Gisterå, A.; Sheng, Z.; Malmström, R.E.; Hou, M.; Peng, J.; Ma, C.; Liao, W.; et al. Platelets enhance CD4+ central memory T cell responses via platelet factor 4-dependent mitochondrial biogenesis and cell proliferation. Platelets 2021, 33, 360–370. [Google Scholar] [CrossRef]
- Figueras, A.; Alsina-Sanchís, E.; Lahiguera, Á.; Abreu, M.; Muinelo-Romay, L.; Moreno-Bueno, G.; Casanovas, O.; Graupera, M.; Matias-Guiu, X.; Vidal, A.; et al. A Role for CXCR4 in Peritoneal and Hematogenous Ovarian Cancer Dissemination. Mol. Cancer Ther. 2018, 17, 532–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, M.; Rath, D.; Gawaz, M. Role of chemokine receptors CXCR4 and CXCR7 for platelet function. Biochem. Soc. Trans. 2015, 43, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; I von Ungern-Sternberg, S.N.; Seizer, P.; Schlegel, F.; Büttcher, M.; Sindhu, N.A.; Müller, S.; Mack, A.; Gawaz, M. Platelet-derived CXCL12 regulates monocyte function, survival, differentiation into macrophages and foam cells through differential involvement of CXCR4–CXCR7. Cell Death Dis. 2015, 6, e1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, M.; Jene, N.; Byrne, D.; Millar, E.K.A.; O’Toole, S.A.; McNeil, C.M.; Bates, G.J.; Harris, A.L.; Banham, A.H.; Sutherland, R.L.; et al. Recruitment of regulatory T cells is correlated with hypoxia-induced CXCR4 expression, and is associated with poor prognosis in basal-like breast cancers. Breast Cancer Res. 2011, 13, R47. [Google Scholar] [CrossRef] [Green Version]
- Righi, E.; Kashiwagi, S.; Yuan, J.; Santosuosso, M.; Leblanc, P.; Ingraham, R.; Forbes, B.; Edelblute, B.; Collette, B.; Xing, D.; et al. CXCL12/CXCR4 Blockade Induces Multimodal Antitumor Effects That Prolong Survival in an Immunocompetent Mouse Model of Ovarian Cancer. Cancer Res. 2011, 71, 5522–5534. [Google Scholar] [CrossRef] [Green Version]
- Toker, A.; Nguyen, L.T.; Stone, S.C.; Yang, S.C.; Katz, S.R.; Shaw, P.A.; Clarke, B.A.; Ghazarian, D.; Al-Habeeb, A.; Easson, A.; et al. Regulatory T Cells in Ovarian Cancer Are Characterized by a Highly Activated Phenotype Distinct from that in Melanoma. Clin. Cancer Res. 2018, 24, 5685–5696. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, A.; Piermattei, A.; Buzzonetti, A.; Pasciuto, T.; Zampetti, N.; Fossati, M.; Angelico, G.; Iacobelli, V.; Nero, C.; Iannucci, V.; et al. PD-L1 Expression on Circulating Tumour-Derived Microvesicles as a Complementary Tool for Stratification of High-Grade Serous Ovarian Cancer Patients. Cancers 2021, 13, 5200. [Google Scholar] [CrossRef]
- Kamada, T.; Togashi, Y.; Tay, C.; Ha, D.; Sasaki, A.; Nakamura, Y.; Sato, E.; Fukuoka, S.; Tada, Y.; Tanaka, A.; et al. PD-1+ regulatory T cells amplified by PD-1 blockade promote hyperprogression of cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 9999–10008. [Google Scholar] [CrossRef] [Green Version]
- Dinkla, S.; van Cranenbroek, B.; van der Heijden, W.A.; He, X.; Wallbrecher, R.; Dumitriu, I.E.; van der Ven, A.J.; Bosman, G.J.C.G.M.; Koenen, H.J.P.M.; Joosten, I. Platelet microparticles inhibit IL-17 production by regulatory T cells through P-selectin. Blood 2016, 127, 1976–1986. [Google Scholar] [CrossRef] [Green Version]
- Ayyoub, M.; Deknuydt, F.; Raimbaud, I.; Dousset, C.; Leveque, L.; Bioley, G.; Valmori, D. Human memory FOXP3 + Tregs secrete IL-17 ex vivo and constitutively express the T H 17 lineage-specific transcription factor RORγt. Proc. Natl. Acad. Sci. USA 2009, 106, 8635–8640. [Google Scholar] [CrossRef] [Green Version]
- Fleischer, J.; Grage-Griebenow, E.; Kasper, B.; Heine, H.; Ernst, M.; Brandt, E.; Flad, H.-D.; Petersen, F. Platelet Factor 4 Inhibits Proliferation and Cytokine Release of Activated Human T Cells. J. Immunol. 2002, 169, 770–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forlow, S.B.; McEver, R.P.; Nollert, M.U. Leukocyte-leukocyte interactions mediated by platelet microparticles under flow. Blood 2000, 95, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.J.; Comps-Agrar, L.; Hackney, J.; Yu, X.; Huseni, M.; Yang, Y.; Park, S.; Javinal, V.; Chiu, H.; Irving, B.; et al. The Immunoreceptor TIGIT Regulates Antitumor and Antiviral CD8 + T Cell Effector Function. Cancer Cell 2014, 26, 923–937. [Google Scholar] [CrossRef] [Green Version]
- Osorio Ovarian Tumor Tissue Analysis (OTTA) Consortium; Goode, E.L.; Block, M.S.; Kalli, K.R.; Vierkant, R.A.; Chen, W.; Fogarty, Z.C.; Gentry-Maharaj, A.; Toloczko, A.; Hein, A.; et al. Dose-Response Association of CD8+ Tumor-Infiltrating Lymphocytes and Survival Time in High-Grade Serous Ovarian Cancer. JAMA Oncol. 2017, 3, e173290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieber, S.; Reinartz, S.; Raifer, H.; Finkernagel, F.; Dreyer, T.; Bronger, H.; Jansen, J.M.; Wagner, U.; Worzfeld, T.; Müller, R.; et al. Prognosis of ovarian cancer is associated with effector memory CD8+ T cell accumulation in ascites, CXCL9 levels and activation-triggered signal transduction in T cells. Oncoimmunology 2018, 7, e1424672. [Google Scholar] [CrossRef] [Green Version]
- Meikle, C.K.; Meisler, A.J.; Bird, C.M.; Jeffries, J.A.; Azeem, N.; Garg, P.; Crawford, E.L.; Kelly, C.A.; Gao, T.Z.; Wuescher, L.M.; et al. Platelet-T cell aggregates in lung cancer patients: Implications for thrombosis. PLoS ONE 2020, 15, e0236966. [Google Scholar] [CrossRef]
- Veerman, K.M.; Williams, M.J.; Uchimura, K.; Singer, M.S.; Merzaban, J.S.; Naus, S.; Carlow, D.A.; Owen, P.; Rivera-Nieves, J.; Rosen, S.D.; et al. Interaction of the selectin ligand PSGL-1 with chemokines CCL21 and CCL19 facilitates efficient homing of T cells to secondary lymphoid organs. Nat. Immunol. 2007, 8, 532–539. [Google Scholar] [CrossRef]
- Tinoco, R.; Neubert, E.N.; Stairiker, C.J.; Henriquez, M.L.; Bradley, L.M. PSGL-1 Is a T Cell Intrinsic Inhibitor That Regulates Effector and Memory Differentiation and Responses During Viral Infection. Front. Immunol. 2021, 12, 677824. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, X.J.; Kundu, S.D.; Pins, M.; Javonovic, B.; Meyer, R.; Kim, S.-J.; Greenberg, N.M.; Kuzel, T.; Meagher, R.; et al. Blockade of transforming growth factor-β signaling in tumor-reactive CD8+ T cells activates the antitumor immune response cycle. Mol. Cancer Ther. 2006, 5, 1733–1743. [Google Scholar] [CrossRef] [Green Version]
- Marcheteau, E.; Farge, T.; Pérès, M.; Labrousse, G.; Tenet, J.; Delmas, S.; Chusseau, M.; Duprez-Paumier, R.; Franchet, C.; Dalenc, F.; et al. Thrombospondin-1 Silencing Improves Lymphocyte Infiltration in Tumors and Response to Anti-PD-1 in Triple-Negative Breast Cancer. Cancers 2021, 13, 4059. [Google Scholar] [CrossRef]
- Rachidi, S.; Metelli, A.; Riesenberg, B.; Wu, B.X.; Nelson, M.H.; Wallace, C.; Paulos, C.M.; Rubinstein, M.P.; Garrett-Mayer, E.; Hennig, M.; et al. Platelets subvert T cell immunity against cancer via GARP-TGFβ axis. Sci. Immunol. 2017, 2, eaai7911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riesenberg, B.P.; Ansa-Addo, E.A.; Gutierrez, J.; Timmers, C.D.; Liu, B.; Li, Z. Cutting Edge: Targeting Thrombocytes to Rewire Anticancer Immunity in the Tumor Microenvironment and Potentiate Efficacy of PD-1 Blockade. J. Immunol. 2019, 203, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, J.; Chen, Z.; Hu, Q.; Wang, C.; Yan, J.; Dotti, G.; Huang, P.; Gu, Z. Engineering PD-1-Presenting Platelets for Cancer Immunotherapy. Nano Lett. 2018, 18, 5716–5725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ke, X.; Zeng, S.; Wu, M.; Lou, J.; Wu, L.; Huang, P.; Huang, L.; Wang, F.; Pan, S. Analysis of CD8+ Treg cells in patients with ovarian cancer: A possible mechanism for immune impairment. Cell. Mol. Immunol. 2015, 12, 580–591. [Google Scholar] [CrossRef] [Green Version]
- Napoletano, C.; Bellati, F.; Landi, R.; Pauselli, S.; Marchetti, C.; Visconti, V.; Sale, P.; Liberati, M.; Rughetti, A.; Frati, L.; et al. Ovarian cancer cytoreduction induces changes in T cell population subsets reducing immunosuppression. J. Cell. Mol. Med. 2010, 14, 2748–2759. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Chen, X.; Lou, J.; Zhang, S.; Zhang, X.; Huang, L.; Sun, R.; Huang, P.; Wang, F.; Pan, S. TGF-β1 contributes to CD8+ Treg induction through p38 MAPK signaling in ovarian cancer microenvironment. Oncotarget 2016, 7, 44534–44544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, J.R.; Wick, D.A.; Nielsen, J.S.; Tran, E.; Milne, K.; McMurtrie, E.; Nelson, B.H. Profound elevation of CD8+ T cells expressing the intraepithelial lymphocyte marker CD103 (αE/β7 Integrin) in high-grade serous ovarian cancer. Gynecol. Oncol. 2010, 118, 228–236. [Google Scholar] [CrossRef]
- Nierodzik, M.L.; Karpatkin, S. Thrombin induces tumor growth, metastasis, and angiogenesis: Evidence for a thrombin-regulated dormant tumor phenotype. Cancer Cell 2006, 10, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Kopp, H.-G.; Placke, T.; Salih, H.R. Platelet-Derived Transforming Growth Factor-β Down-Regulates NKG2D Thereby Inhibiting Natural Killer Cell Antitumor Reactivity. Cancer Res. 2009, 69, 7775–7783. [Google Scholar] [CrossRef] [Green Version]
- Alexander, E.T.; Minton, A.R.; Peters, M.C.; van Ryn, J.; Gilmour, S.K. Thrombin inhibition and cisplatin block tumor progression in ovarian cancer by alleviating the immunosuppressive microenvironment. Oncotarget 2016, 7, 85291–85305. [Google Scholar] [CrossRef] [Green Version]
- Motz, G.T.; Santoro, S.P.; Wang, L.-P.; Garrabrant, T.; Lastra, R.R.; Hagemann, I.S.; Lal, P.; Feldman, M.D.; Benencia, F.; Coukos, G. Tumor endothelium FasL establishes a selective immune barrier promoting tolerance in tumors. Nat. Med. 2014, 20, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Zelenay, S.; van der Veen, A.G.; Böttcher, J.P.; Snelgrove, K.J.; Rogers, N.; Acton, S.E.; Chakravarty, P.; Girotti, M.R.; Marais, R.; Quezada, S.A.; et al. Cyclooxygenase-Dependent Tumor Growth through Evasion of Immunity. Cell 2015, 162, 1257–1270. [Google Scholar] [CrossRef] [Green Version]
- Melchers, F. Checkpoints that control B cell development. J. Clin. Investig. 2015, 125, 2203–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petitprez, F.; de Reyniès, A.; Keung, E.Z.; Chen, T.W.-W.; Sun, C.-M.; Calderaro, J.; Jeng, Y.-M.; Hsiao, L.-P.; Lacroix, L.; Bougoüin, A.; et al. B cells are associated with survival and immunotherapy response in sarcoma. Nature 2020, 577, 556–560. [Google Scholar] [CrossRef]
- Wei, X.; Jin, Y.; Tian, Y.; Zhang, H.; Wu, J.; Lu, W.; Lu, X. Regulatory B cells contribute to the impaired antitumor immunity in ovarian cancer patients. Tumor Biol. 2015, 37, 6581–6588. [Google Scholar] [CrossRef]
- Capello, M.; Vykoukal, J.V.; Katayama, H.; Bantis, L.E.; Wang, H.; Kundnani, D.L.; Aguilar-Bonavides, C.; Aguilar, M.; Tripathi, S.C.; Dhillon, D.S.; et al. Exosomes harbor B cell targets in pancreatic adenocarcinoma and exert decoy function against complement-mediated cytotoxicity. Nat. Commun. 2019, 10, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Visser, K.E.; Korets, L.V.; Coussens, L.M. De novo carcinogenesis promoted by chronic inflammation is B lymphocyte dependent. Cancer Cell 2005, 7, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Miao, S.; Shu, D.; Zhu, Y.; Lu, M.; Zhang, Q.; Pei, Y.; He, A.-D.; Ma, R.; Zhang, B.; Ming, Z.-Y. Cancer cell-derived immunoglobulin G activates platelets by binding to platelet FcγRIIa. Cell Death Dis. 2019, 10, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Veen, W.; Globinska, A.; Jansen, K.; Straumann, A.; Kubo, T.; Verschoor, D.; Wirz, O.F.; Castro-Giner, F.; Tan, G.; Rückert, B.; et al. A novel proangiogenic B cell subset is increased in cancer and chronic inflammation. Sci. Adv. 2020, 6, eaaz3559. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.P.; Elstrand, M.B.; Holth, A.; Silins, I.; Berner, A.; Trope, C.G.; Davidson, B.; Risberg, B. Nk- and B-Cell Infiltration Correlates with Worse Outcome in Metastatic Ovarian Carcinoma. Am. J. Clin. Pathol. 2006, 125, 451–458. [Google Scholar] [CrossRef]
- Nielsen, J.S.; Sahota, R.A.; Milne, K.; Kost, S.E.; Nesslinger, N.J.; Watson, P.H.; Nelson, B.H. CD20+ Tumor-Infiltrating Lymphocytes Have an Atypical CD27− Memory Phenotype and Together with CD8+ T Cells Promote Favorable Prognosis in Ovarian Cancer. Clin. Cancer Res. 2012, 18, 3281–3292. [Google Scholar] [CrossRef] [Green Version]
- Elzey, B.D.; Grant, J.F.; Sinn, H.W.; Nieswandt, B.; Waldschmidt, T.J.; Ratliff, T.L. Cooperation between platelet-derived CD154 and CD4+ T cells for enhanced germinal center formation. J. Leukoc. Biol. 2005, 78, 80–84. [Google Scholar] [CrossRef]
- Morva, A.; Lemoine, S.; Achour, A.; Pers, J.-O.; Youinou, P.; Jamin, C. Maturation and function of human dendritic cells are regulated by B lymphocytes. Blood 2012, 119, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Lo, C.M.; Ling, C.C.; Liu, X.B.; Ng, K.T.-P.; Chu, A.C.Y.; Ma, Y.Y.; Li, C.X.; Fan, S.T.; Man, K. Regulatory B cells accelerate hepatocellular carcinoma progression via CD40/CD154 signaling pathway. Cancer Lett. 2014, 355, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Wallace, C.H.; Wu, B.X.; Salem, M.; Ansa-Addo, E.A.; Metelli, A.; Sun, S.; Gilkeson, G.; Shlomchik, M.J.; Liu, B.; Li, Z. B lymphocytes confer immune tolerance via cell surface GARP-TGF-β complex. J. Clin. Investig. 2018, 3, e99863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrl, J.H.; Thevenin, C.; Rieckmann, P.; Fauci, A.S. Transforming growth factor-beta suppresses human B lymphocyte Ig production by inhibiting synthesis and the switch from the membrane form to the secreted form of Ig mRNA. J. Immunol. 1991, 146, 4016–4023. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A. The regulation of IgA class switching. Nat. Rev. Immunol. 2008, 8, 421–434. [Google Scholar] [CrossRef]
- Bulk, J.V.D.; de Miranda, N.F.; Dijke, P.T. Therapeutic targeting of TGF-β in cancer: Hacking a master switch of immune suppression. Clin. Sci. 2021, 135, 35–52. [Google Scholar] [CrossRef]
- Olkhanud, P.B.; Damdinsuren, B.; Bodogai, M.; Gress, R.E.; Sen, R.; Wejksza, K.; Malchinkhuu, E.; Wersto, R.P.; Biragyn, A. Tumor-Evoked Regulatory B Cells Promote Breast Cancer Metastasis by Converting Resting CD4+ T Cells to T-Regulatory Cells. Cancer Res. 2011, 71, 3505–3515. [Google Scholar] [CrossRef] [Green Version]
- Gill, P.; Jindal, N.L.; Jagdis, A.; Vadas, P. Platelets in the immune response: Revisiting platelet-activating factor in anaphylaxis. J. Allergy Clin. Immunol. 2015, 135, 1424–1432. [Google Scholar] [CrossRef] [Green Version]
- Denizot, Y.; Guglielmi, L.; Donnard, M.; Trimoreau, F. Platelet-activating Factor and Normal or Leukaemic Haematopoiesis. Leuk. Lymphoma 2003, 44, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Braun, A.; Anders, H.-J.; Gudermann, T.; Mammadova-Bach, E. Platelet-Cancer Interplay: Molecular Mechanisms and New Therapeutic Avenues. Front. Oncol. 2021, 11, 665534. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Fu, Q.; Diggs, L.P.; McVey, J.C.; McCallen, J.; Wabitsch, S.; Ruf, B.; Brown, Z.; Heinrich, B.; Zhang, Q.; et al. Platelets control liver tumor growth through P2Y12-dependent CD40L release in NAFLD. Cancer Cell 2022, 40, 986–998.e5. [Google Scholar] [CrossRef] [PubMed]
- Menter, D.G.; Tucker, S.C.; Kopetz, S.; Sood, A.K.; Crissman, J.D.; Honn, K.V. Platelets and cancer: A casual or causal relationship: Revisited. Cancer Metastasis Rev. 2014, 33, 231–269. [Google Scholar] [CrossRef] [Green Version]
- Gawaz, M.; Neumann, F.-J.; Dickfeld, T.; Reininger, A.; Adelsberger, H.; Gebhardt, A.; Schömig, A. Vitronectin Receptor (α v β 3) Mediates Platelet Adhesion to the Luminal Aspect of Endothelial Cells. Circulation 1997, 96, 1809–1818. [Google Scholar] [CrossRef]
- Bombeli, T.; Schwartz, B.R.; Harlan, J.M. Adhesion of Activated Platelets to Endothelial Cells: Evidence for a GPIIbIIIa-dependent Bridging Mechanism and Novel Roles for Endothelial Intercellular Adhesion Molecule 1 (ICAM-1), αvβ3 Integrin, and GPIbα. J. Exp. Med. 1998, 187, 329–339. [Google Scholar] [CrossRef]
- Langer, H.; May, A.E.; Bültmann, A.; Gawaz, M. ADAM15 is an adhesion receptor for platelet GPIIb-IIIa and induces platelet activation. Thromb. Haemost. 2005, 94, 555–561. [Google Scholar] [CrossRef]
- Tan, D.S.; Agarwal, R.; Kaye, S.B. Mechanisms of transcoelomic metastasis in ovarian cancer. Lancet Oncol. 2006, 7, 925–934. [Google Scholar] [CrossRef]
- Schmied, L.; Höglund, P.; Meinke, S. Platelet-Mediated Protection of Cancer Cells from Immune Surveillance—Possible Implications for Cancer Immunotherapy. Front. Immunol. 2021, 12, 640578. [Google Scholar] [CrossRef]
- Schlesinger, M. Role of platelets and platelet receptors in cancer metastasis. J. Hematol. Oncol. 2018, 11, 125. [Google Scholar] [CrossRef] [Green Version]
- Olsson, A.-K.; Cedervall, J. NETosis in Cancer—Platelet–Neutrophil Crosstalk Promotes Tumor-Associated Pathology. Front. Immunol. 2016, 7, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linke, B.; Schreiber, Y.; Picard-Willems, B.; Slattery, P.; Nüsing, R.M.; Harder, S.; Geisslinger, G.; Scholich, K. Activated Platelets Induce an Anti-Inflammatory Response of Monocytes/Macrophages through Cross-Regulation of PGE2 and Cytokines. Mediat. Inflamm. 2017, 2017, 1463216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhong, H.; Bao, W.; Boulad, N.; Evangelista, J.; Haider, M.A.; Bussel, J.; Yazdanbakhsh, K. Defective regulatory B-cell compartment in patients with immune thrombocytopenia. Blood 2012, 120, 3318–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfes, V.; Idel, C.; Pries, R.; Plötze-Martin, K.; Habermann, J.; Gemoll, T.; Bohnet, S.; Latz, E.; Ribbat-Idel, J.; Franklin, B.S.; et al. PD-L1 is expressed on human platelets and is affected by immune checkpoint therapy. Oncotarget 2018, 9, 27460–27470. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Cheng, J.; Ye, M.; Liu, D.; Zhang, Y. Association of pretreatment thrombocytosis with prognosis in ovarian cancer: A systematic review and meta-analysis. J. Gynecol. Oncol. 2019, 30, e5. [Google Scholar] [CrossRef]
- Canzler, U.; Lück, H.-J.; Neuser, P.; Sehouli, J.; Burges, A.; Harter, P.; Schmalfeldt, B.; Aminossadati, B.; Mahner, S.; Kommoss, S.; et al. Prognostic role of thrombocytosis in recurrent ovarian cancer: A pooled analysis of the AGO Study Group. Arch. Gynecol. Obstet. 2020, 301, 1267–1274. [Google Scholar] [CrossRef]
- Nakao, S.; Minaguchi, T.; Itagaki, H.; Hosokawa, Y.; Shikama, A.; Tasaka, N.; Akiyama, A.; Ochi, H.; Matsumoto, K.; Satoh, T. Pretreatment thrombocytosis as an independent predictive factor for chemoresistance and poor survival in epithelial ovarian cancer. J. Ovarian Res. 2020, 13, 55–59. [Google Scholar] [CrossRef]
- Watrowski, R.; Heinze, G.; Jäger, C.; Forster, J.; Zeillinger, R. Usefulness of the preoperative platelet count in the diagnosis of adnexal tumors. Tumor Biol. 2016, 37, 12079–12087. [Google Scholar] [CrossRef]
- Hufnagel, D.H.; Cozzi, G.D.; Crispens, M.A.; Beeghly-Fadiel, A. Platelets, Thrombocytosis, and Ovarian Cancer Prognosis: Surveying the Landscape of the Literature. Int. J. Mol. Sci. 2020, 21, 8169. [Google Scholar] [CrossRef]
- Naito, R.; Miyauchi, K.; Yasuda, S.; Kaikita, K.; Akao, M.; Ako, J.; Matoba, T.; Nakamura, M.; Hagiwara, N.; Kimura, K.; et al. Rivaroxaban Monotherapy vs Combination Therapy with Antiplatelets on Total Thrombotic and Bleeding Events in Atrial Fibrillation with Stable Coronary Artery Disease. JAMA Cardiol. 2022, 7, 787–794. [Google Scholar] [CrossRef]
- Anadani, M.; de Havenon, A.; Henninger, N.; Kuohn, L.; Mac Grory, B.; Furie, K.L.; Kim, A.S.; Easton, J.D.; Johnston, S.C.; Yaghi, S. Antiplatelet Use and Ischemic Stroke Risk in Minor Stroke or Transient Ischemic Attack: A Post Hoc Analysis of the POINT Trial. Stroke 2021, 52, e773–e776. [Google Scholar] [CrossRef] [PubMed]
- Cooke, N.M.; Spillane, C.D.; Sheils, O.; O’Leary, J.; Kenny, D. Aspirin and P2Y12 inhibition attenuate platelet-induced ovarian cancer cell invasion. BMC Cancer 2015, 15, 627. [Google Scholar] [CrossRef] [Green Version]
- Denslow, A.; Świtalska, M.; Jarosz, J.; Papiernik, D.; Porshneva, K.; Nowak, M.; Wietrzyk, J. Clopidogrel in a combined therapy with anticancer drugs—Effect on tumor growth, metastasis, and treatment toxicity: Studies in animal models. PLoS ONE 2017, 12, e0188740. [Google Scholar] [CrossRef]
- Wright, J.R.; Chauhan, M.; Shah, C.; Ring, A.; Thomas, A.L.; Goodall, A.H.; Adlam, D. The TICONC (Ticagrelor-Oncology) Study. JACC CardioOncol. 2020, 2, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Cuzick, J.; Thorat, M.A.; Bosetti, C.; Brown, P.H.; Burn, J.; Cook, N.R.; Ford, L.G.; Jacobs, E.J.; Jankowski, J.A.; La Vecchia, C.; et al. Estimates of benefits and harms of prophylactic use of aspirin in the general population. Ann. Oncol. 2014, 26, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.-O.; Park, S.-Y.; Cho, E.S.; Zhang, X.; Lee, S.K.; Ahn, H.-J.; Chun, K.-S.; Chung, W.-Y.; Song, N.-Y. Platelet CLEC2-Podoplanin Axis as a Promising Target for Oral Cancer Treatment. Front. Immunol. 2021, 12, 807600. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.E.; Navarro-Núñez, L.; Finney, B.A.; Mourão-Sá, D.; Pollitt, A.Y.; Watson, S.P. CLEC-2 is not required for platelet aggregation at arteriolar shear. J. Thromb. Haemost. 2010, 8, 2328–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, X.; Nilsson, M.; Goldman, J.; Reck, M.; Nakagawa, K.; Kato, T.; Ares, L.P.; Frimodt-Moller, B.; Wolff, K.; Visseren-Grul, C.; et al. Dual EGFR-VEGF Pathway Inhibition: A Promising Strategy for Patients With EGFR-Mutant NSCLC. J. Thorac. Oncol. 2020, 16, 205–215. [Google Scholar] [CrossRef]
- Subbiah, V.; Dumbrava, E.I.; Jiang, Y.; Thein, K.Z.; Naing, A.; Hong, D.S.; Fu, S.; Piha-Paul, S.A.; Tsimberidou, A.M.; Janku, F.; et al. Dual EGFR blockade with cetuximab and erlotinib combined with anti-VEGF antibody bevacizumab in advanced solid tumors: A phase 1 dose escalation triplet combination trial. Exp. Hematol. Oncol. 2020, 9, 7–8. [Google Scholar] [CrossRef]
- Hu, Q.; Hada, A.; Han, L. Platelet count as a biomarker for monitoring treatment response and disease recurrence in recurrent epithelial ovarian cancer. J. Ovarian Res. 2020, 13, 78. [Google Scholar] [CrossRef]
- Soff, G.A.; Ray-Coquard, I.; Rivera, L.J.M.; Fryzek, J.; Mullins, M.; Bylsma, L.C.; Park, J.K. Systematic literature review and meta-analysis on use of Thrombopoietic agents for chemotherapy-induced thrombocytopenia. PLoS ONE 2022, 17, e0257673. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, P.; Han, Y.; Zhang, H. Platelet transfusion for cancer secondary thrombocytopenia: Platelet and cancer cell interaction. Transl. Oncol. 2021, 14, 101022. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oncul, S.; Cho, M.S. Interactions between Platelets and Tumor Microenvironment Components in Ovarian Cancer and Their Implications for Treatment and Clinical Outcomes. Cancers 2023, 15, 1282. https://doi.org/10.3390/cancers15041282
Oncul S, Cho MS. Interactions between Platelets and Tumor Microenvironment Components in Ovarian Cancer and Their Implications for Treatment and Clinical Outcomes. Cancers. 2023; 15(4):1282. https://doi.org/10.3390/cancers15041282
Chicago/Turabian StyleOncul, Selin, and Min Soon Cho. 2023. "Interactions between Platelets and Tumor Microenvironment Components in Ovarian Cancer and Their Implications for Treatment and Clinical Outcomes" Cancers 15, no. 4: 1282. https://doi.org/10.3390/cancers15041282
APA StyleOncul, S., & Cho, M. S. (2023). Interactions between Platelets and Tumor Microenvironment Components in Ovarian Cancer and Their Implications for Treatment and Clinical Outcomes. Cancers, 15(4), 1282. https://doi.org/10.3390/cancers15041282