
Characterisation of Aberrant Metabolic Pathways in Hepatoblastoma Using Liquid Chromatography and Tandem Mass Spectrometry (LC-MS/MS)

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Samples
2.2. Untargeted Metabolomics
2.3. Targeted Metabolomics
2.4. Gene Set Enrichment Analysis
2.5. Gene Expression Data Analysis
2.6. Immunohistochemistry (IHC)
3. Results
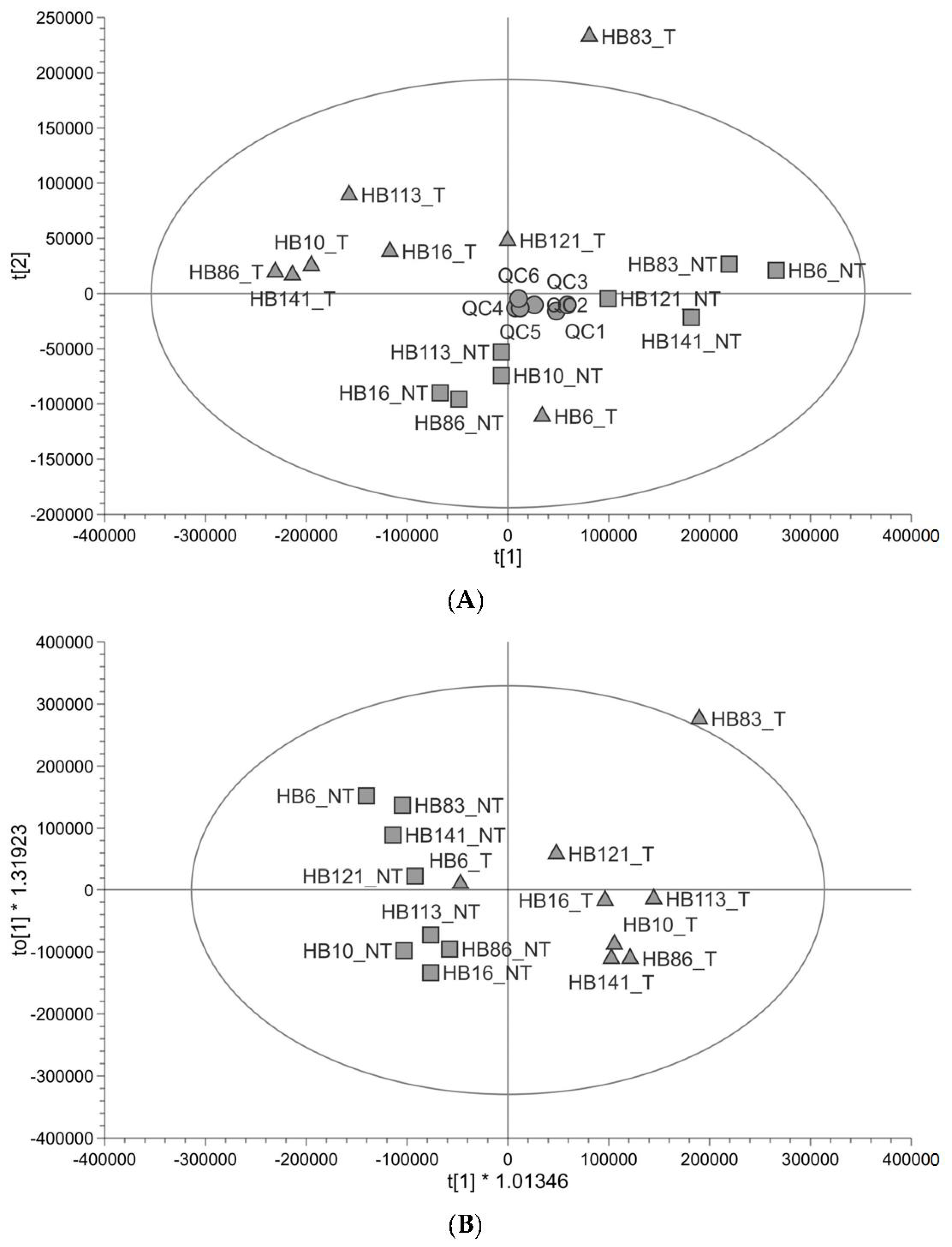
3.1. The Metabolomic Profile of Hepatoblastoma Is Different to That of Non-Tumour Tissue in Paired Samples
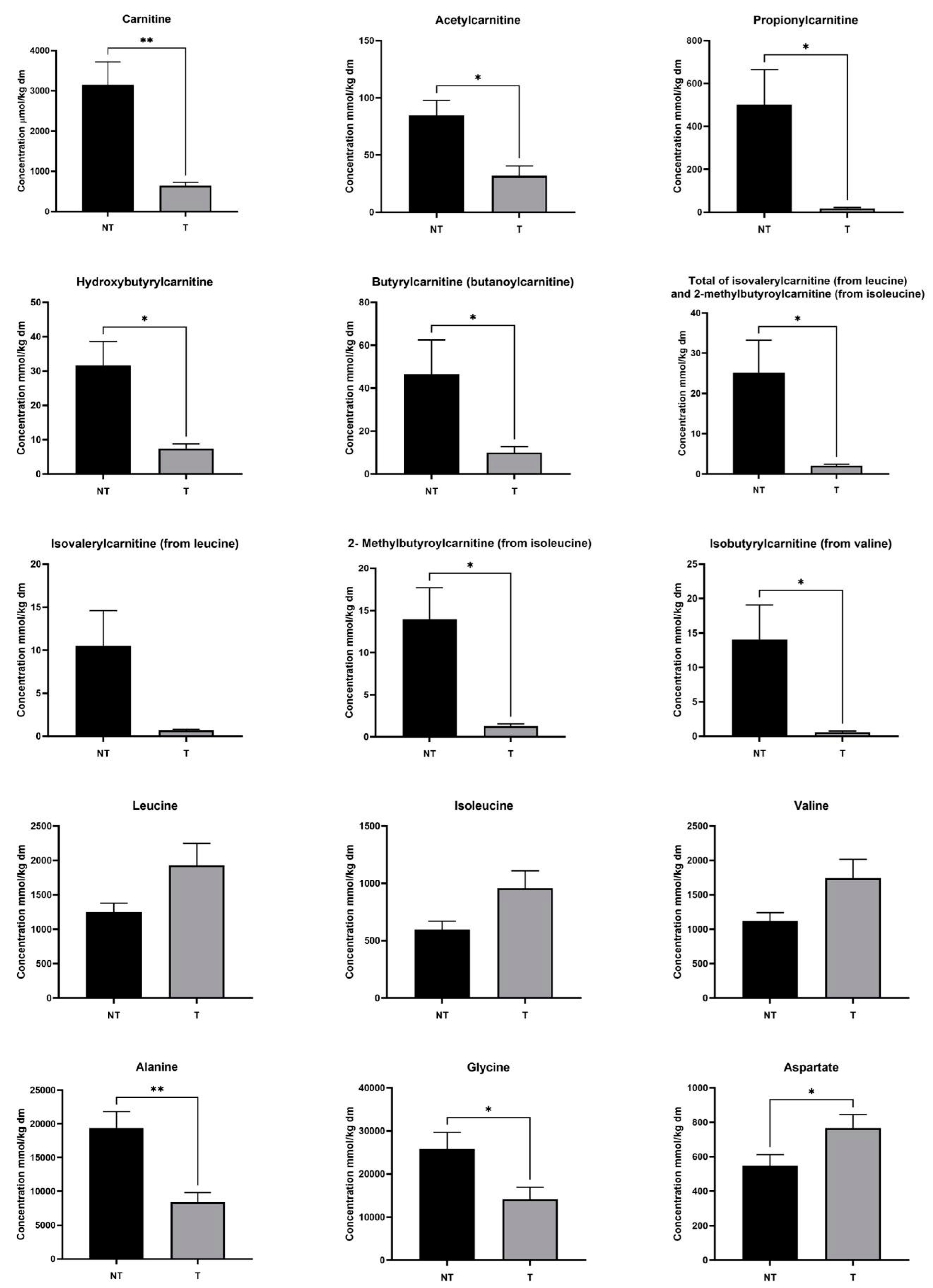
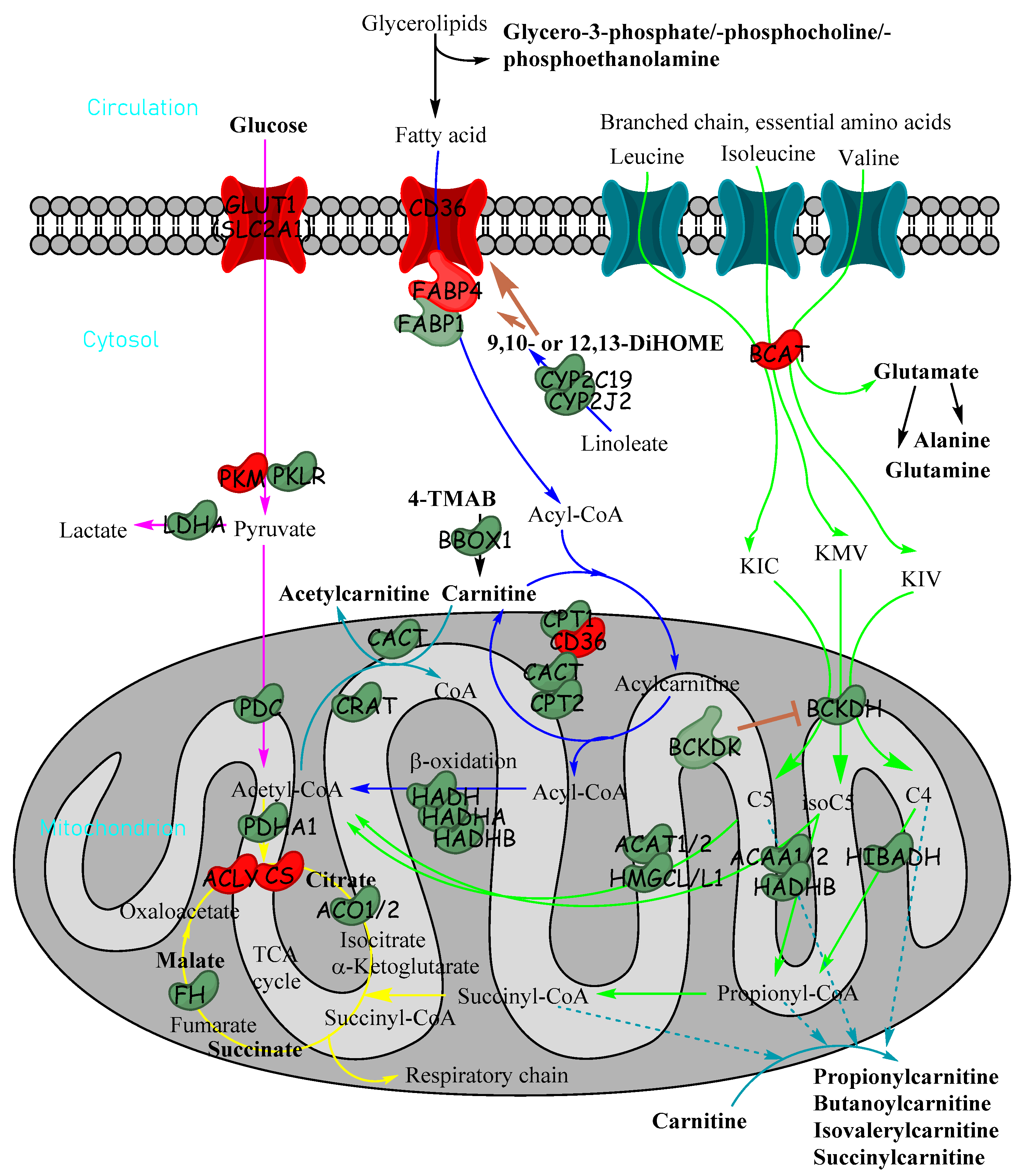
3.2. Targeted Metabolomics Demonstrates a Reduction in Short-Chain Acylcarnitine Levels in Hepatoblastoma Tissue
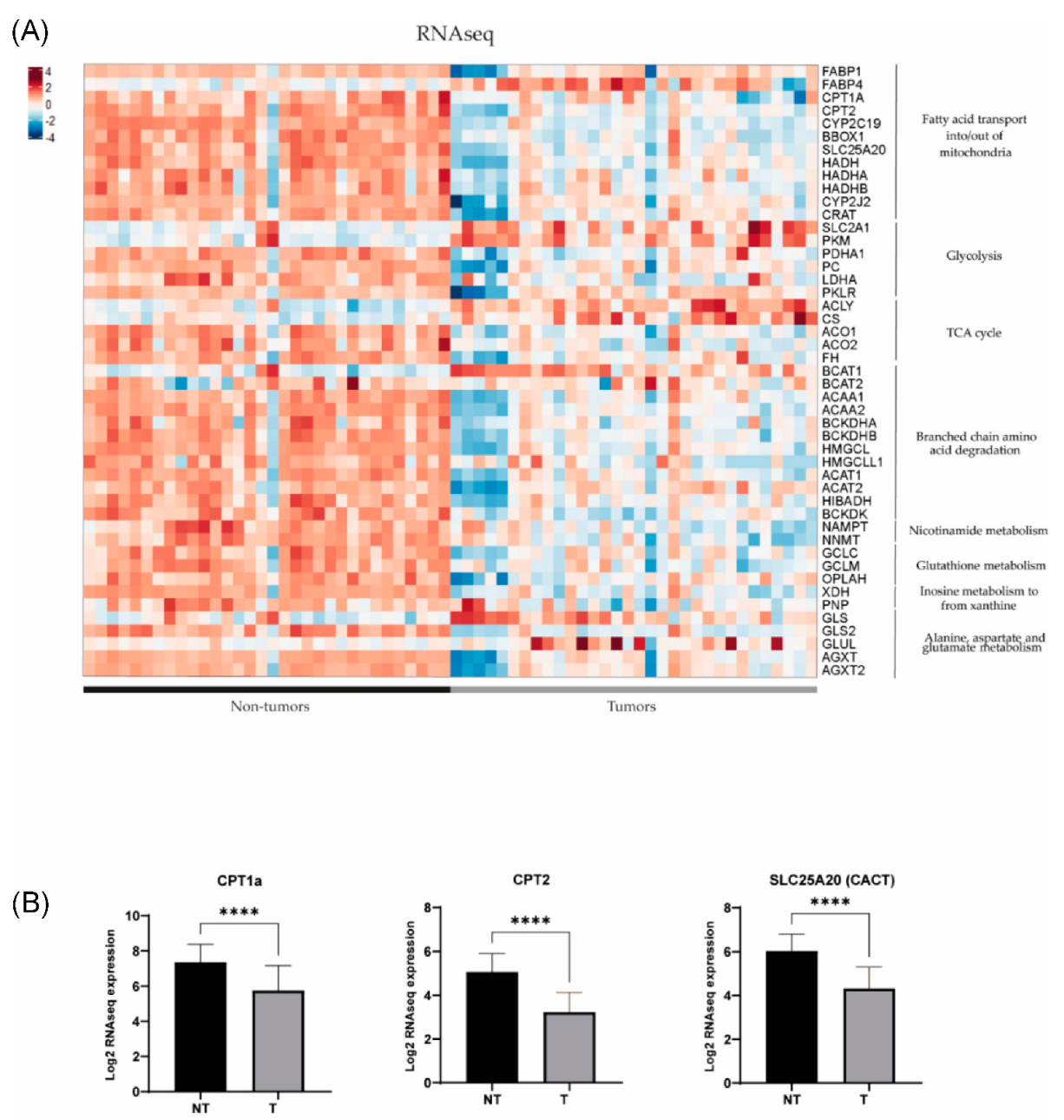
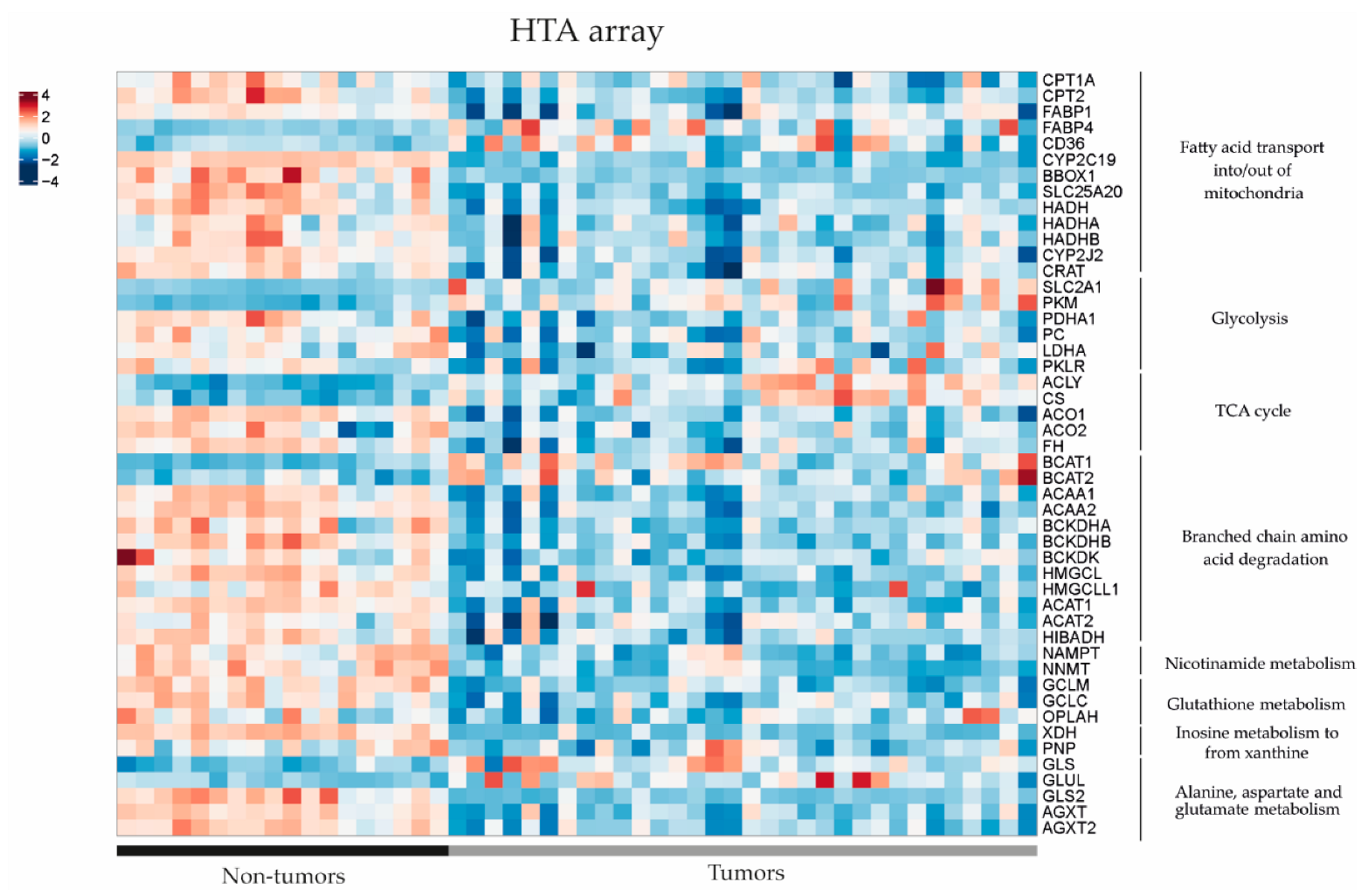
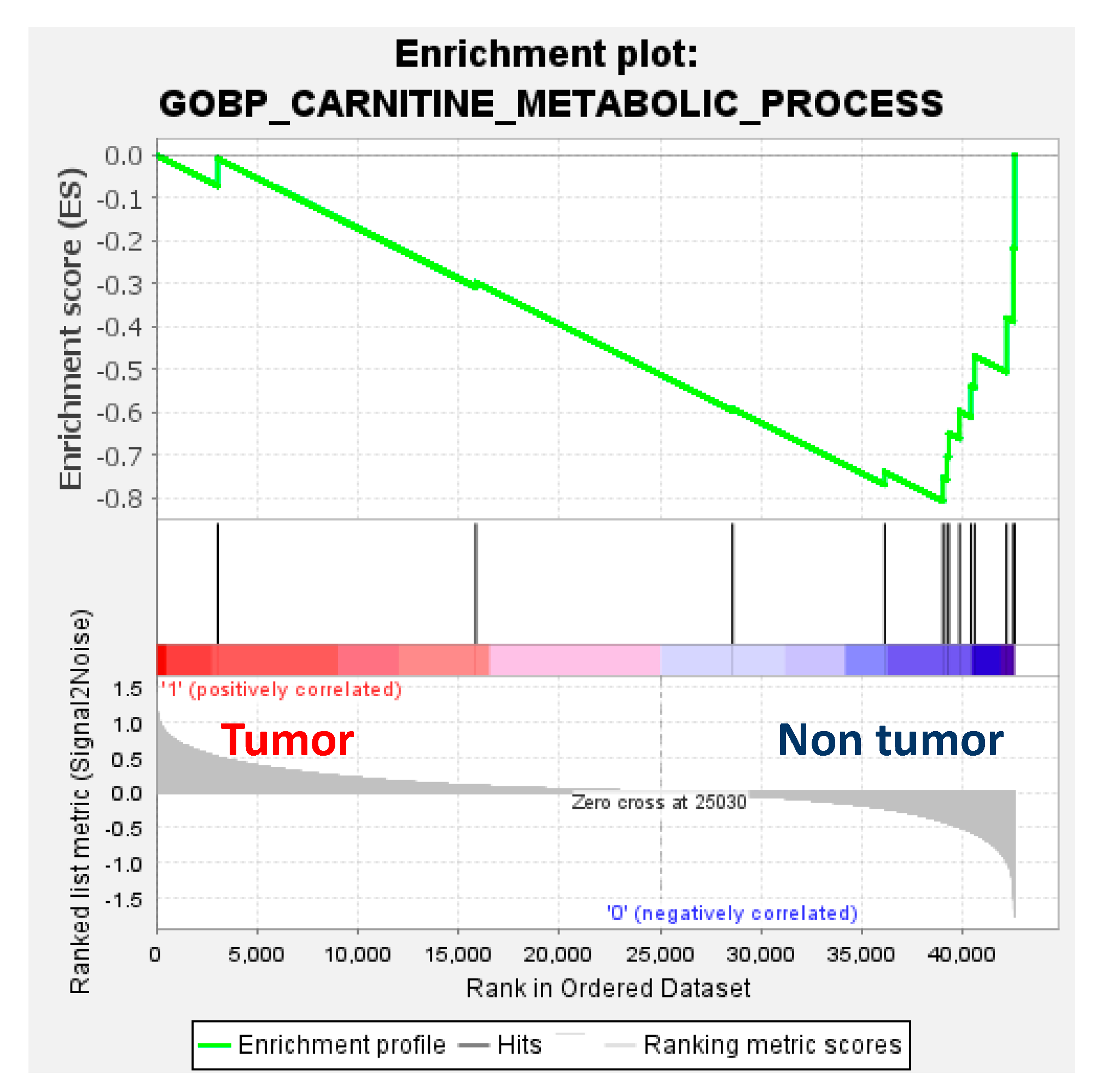
3.3. Metabolomic Profile in Hepatoblastoma Correlates with Transcriptomic Profile
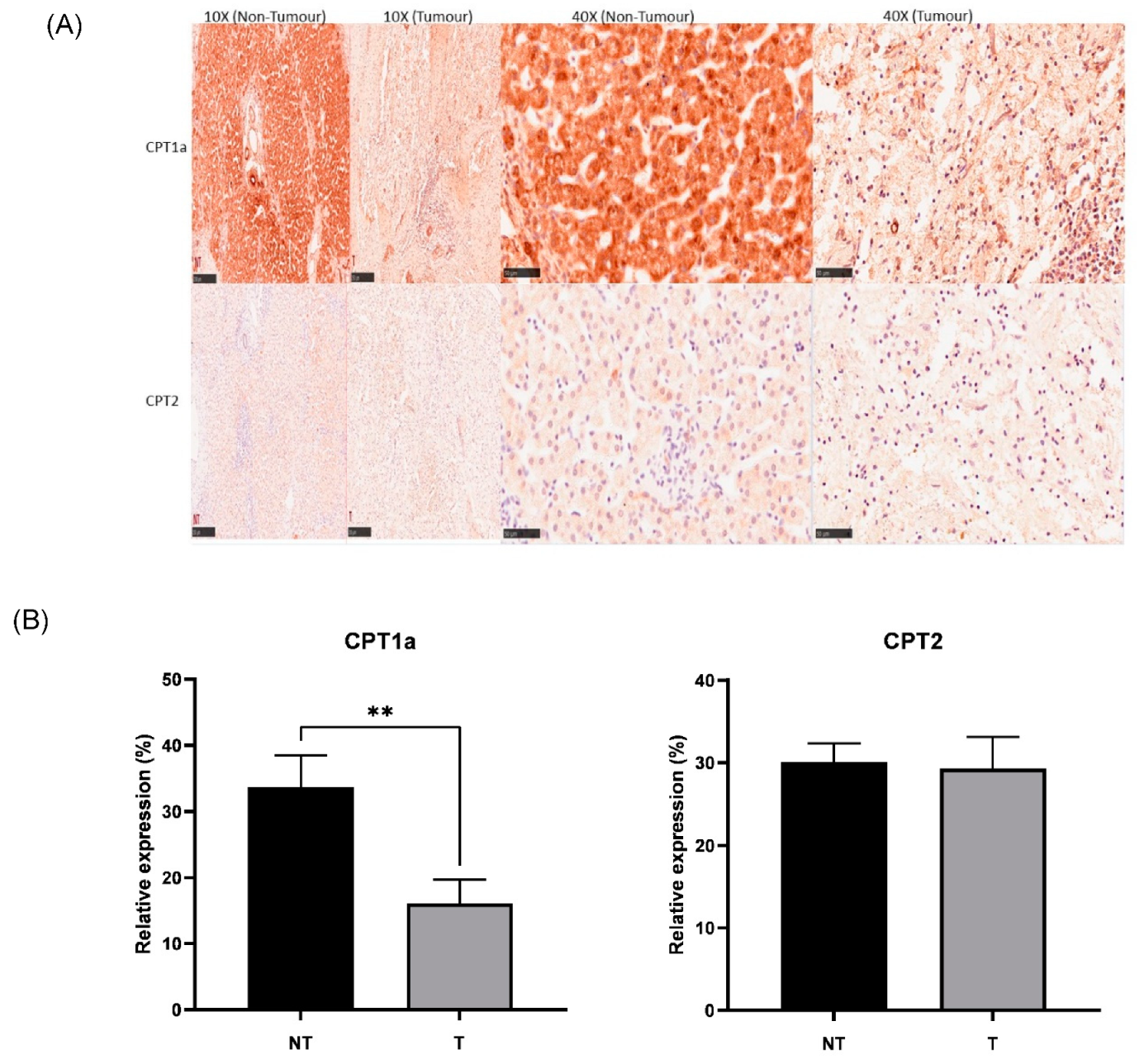
3.4. CPT1a Is Downregulated in Hepatoblastoma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Feng, J.; Polychronidis, G.; Heger, U.; Frongia, G.; Mehrabi, A.; Hoffmann, K. Incidence trends and survival prediction of hepatoblastoma in children: A population-based study. Cancer Commun. 2019, 39, 62. [Google Scholar] [CrossRef] [PubMed]
- Kahla, J.A.; Siegel, D.A.; Dai, S.; Lupo, P.J.; Foster, J.H.; Scheurer, M.E.; Heczey, A.A. Incidence and 5-year survival of children and adolescents with hepatoblastoma in the United States. Pediatr. Blood Cancer 2022, 69, e29763. [Google Scholar] [CrossRef] [PubMed]
- Aronson, D.C.; Czauderna, P.; Maibach, R.; Perilongo, G.; Morland, B. The treatment of hepatoblastoma: Its evolution and the current status as per the SIOPEL trials. J. Indian Assoc. Pediatr. Surg. 2014, 19, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Khanna, R.; Verma, S.K. Pediatric hepatocellular carcinoma. World J. Gastroenterol. 2018, 24, 3980–3999. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, S.; Lopez-Terrada, D.; Alaggio, R. Hepatoblastoma and Pediatric Hepatocellular Carcinoma: An Update. Pediatr. Dev. Pathol. 2020, 23, 79–95. [Google Scholar] [CrossRef]
- Meyers, R.L.; Maibach, R.; Hiyama, E.; Häberle, B.; Krailo, M.; Rangaswami, A.; Aronson, D.C.; Malogolowkin, M.H.; Perilongo, G.; von Schweinitz, D.; et al. Risk-stratified staging in paediatric hepatoblastoma: A unified analysis from the Children’s Hepatic tumors International Collaboration. Lancet Oncol. 2017, 18, 122–131. [Google Scholar] [CrossRef]
- Buendia, M.A. Genetic alterations in hepatoblastoma and hepatocellular carcinoma: Common and distinctive aspects. Med. Pediatr. Oncol. 2002, 39, 530–535. [Google Scholar] [CrossRef]
- Buendia, M.A.; Armengol, C.; Cairo, S. Molecular classification of hepatoblastoma and prognostic value of the HB 16-gene signature. Hepatology 2017, 66, 1351–1352. [Google Scholar] [CrossRef]
- Cairo, S.; Armengol, C.; De Reyniès, A.; Wei, Y.; Thomas, E.; Renard, C.A.; Goga, A.; Balakrishnan, A.; Semeraro, M.; Gresh, L.; et al. Hepatic stem-like phenotype and interplay of Wnt/beta-catenin and Myc signaling in aggressive childhood liver cancer. Cancer Cell 2008, 14, 471–484. [Google Scholar] [CrossRef]
- Cairo, S.; Armengol, C.; Maibach, R.; Häberle, B.; Becker, K.; Carrillo-Reixach, J.; Guettier, C.; Vokuhl, C.; Schmid, I.; Buendia, M.A.; et al. A combined clinical and biological risk classification improves prediction of outcome in hepatoblastoma patients. Eur. J. Cancer 2020, 141, 30–39. [Google Scholar] [CrossRef]
- Carrillo-Reixach, J.; Torrens, L.; Simon-Coma, M.; Royo, L.; Domingo-Sàbat, M.; Abril-Fornaguera, J.; Akers, N.; Sala, M.; Ragull, S.; Arnal, M.; et al. Epigenetic footprint enables molecular risk stratification of hepatoblastoma with clinical implications. J. Hepatol. 2020, 73, 328–341. [Google Scholar] [CrossRef]
- Nagae, G.; Yamamoto, S.; Fujita, M.; Fujita, T.; Nonaka, A.; Umeda, T.; Fukuda, S.; Tatsuno, K.; Maejima, K.; Hayashi, A.; et al. Genetic and epigenetic basis of hepatoblastoma diversity. Nat. Commun. 2021, 12, 5423. [Google Scholar] [CrossRef] [PubMed]
- Abril-Fornaguera, J.; Torrens, L.; Andreu-Oller, C.; Carrillo-Reixach, J.; Rialdi, A.; Balaseviciute, U.; Pinyol, R.; Montironi, C.; Haber, P.K.; Del Río-Álvarez, Á.; et al. Identification of IGF2 as Genomic Driver and Actionable Therapeutic Target in Hepatoblastoma. Mol. Cancer Ther. 2023, 22, 485–498. [Google Scholar] [CrossRef]
- Chokkathukalam, A.; Kim, D.H.; Barrett, M.P.; Breitling, R.; Creek, D.J. Stable isotope-labeling studies in metabolomics: New insights into structure and dynamics of metabolic networks. Bioanalysis 2014, 6, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.P.; Aguiar, T.F.M.; Maschietto, M.; Lemes, R.B.; Caires-Júnior, L.C.; Goulart, E.; Telles-Silva, K.A.; Novak, E.; Cristofani, L.M.; Odone, V.; et al. Hepatoblastomas exhibit marked NNMT downregulation driven by promoter DNA hypermethylation. Tumor Biol. 2020, 42, 1010428320977124. [Google Scholar] [CrossRef]
- Tasic, L.; Avramović, N.; Jadranin, M.; Quintero, M.; Stanisic, D.; Martins, L.G.; Costa, T.; Novak, E.; Odone, V.; Rivas, M.; et al. High-Resolution Magic-Angle-Spinning NMR in Revealing Hepatoblastoma Hallmarks. Biomedicines 2022, 10, 3091. [Google Scholar] [CrossRef]
- Abuawad, A.; Mbadugha, C.; Ghaemmaghami, A.M.; Kim, D.-H. Metabolic characterisation of THP-1 macrophage polarisation using LC–MS-based metabolite profiling. Metabolomics 2020, 16, 33. [Google Scholar] [CrossRef] [PubMed]
- Jaber, M.A.; Ghanim, B.Y.; Al-Natour, M.; Arqoub, D.A.; Abdallah, Q.; Abdelrazig, S.; Alkrad, J.A.; Kim, D.-H.; Qinna, N.A. Potential biomarkers and metabolomics of acetaminophen-induced liver injury during alcohol consumption: A preclinical investigation on C57/BL6 mice. Toxicol. Appl. Pharmacol. 2023, 465, 116451. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K.; Roessner, U.; Trengove, R. Proposed quantitative and alphanumeric metabolite identification metrics. Metabolomics 2014, 10, 1047–1049. [Google Scholar] [CrossRef]
- Sun, D.; Cree, M.G.; Zhang, X.-j.; Bøersheim, E.; Wolfe, R.R. Measurement of stable isotopic enrichment and concentration of long-chain fatty acyl-carnitines in tissue by HPLC-MS. J. Lipid Res. 2006, 47, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Sunny, N.E.; Kalavalapalli, S.; Bril, F.; Garrett, T.J.; Nautiyal, M.; Mathew, J.T.; Williams, C.M.; Cusi, K. Cross-talk between branched-chain amino acids and hepatic mitochondria is compromised in nonalcoholic fatty liver disease. Am. J. Physiol. Metab. 2015, 309, E311–E319. [Google Scholar] [CrossRef] [PubMed]
- Surrati, A.; Linforth, R.; Fisk, I.D.; Sottile, V.; Kim, D.-H. Non-destructive characterisation of mesenchymal stem cell differentiation using LC-MS-based metabolite footprinting. Analyst 2016, 141, 3776–3787. [Google Scholar] [CrossRef]
- Song, Z.; Tang, G.; Zhuang, C.; Wang, Y.; Wang, M.; Lv, D.; Lu, G.; Meng, J.; Xia, M.; Zhu, Z.; et al. Metabolomic profiling of cerebrospinal fluid reveals an early diagnostic model for central nervous system involvement in acute lymphoblastic leukaemia. Br. J. Haematol. 2022, 198, 994–1010. [Google Scholar] [CrossRef]
- McGarry, J.D.; Brown, N.F. The mitochondrial carnitine palmitoyltransferase system. From concept to molecular analysis. Eur. J. Biochem. 1997, 244, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.L.; Lennon, D.L.; Stratman, F.W. Increased acetyl carnitine in rat skeletal muscle as a result of high-intensity short-duration exercise. Implications in the control of pyruvate dehydrogenase activity. FEBS Lett. 1981, 126, 21–24. [Google Scholar] [CrossRef]
- Lo, E.K.K.; Felicianna; Xu, J.H.; Zhan, Q.; Zeng, Z.; El-Nezami, H. The Emerging Role of Branched-Chain Amino Acids in Liver Diseases. Biomedicines 2022, 10, 1444. [Google Scholar] [CrossRef]
- Buchard, B.; Teilhet, C.; Abeywickrama Samarakoon, N.; Massoulier, S.; Joubert-Zakeyh, J.; Blouin, C.; Reynes, C.; Sabatier, R.; Biesse-Martin, A.S.; Vasson, M.P.; et al. Two Metabolomics Phenotypes of Human Hepatocellular Carcinoma in Non-Alcoholic Fatty Liver Disease According to Fibrosis Severity. Metabolites 2021, 11, 54. [Google Scholar] [CrossRef]
- Ericksen, R.E.; Lim, S.L.; McDonnell, E.; Shuen, W.H.; Vadiveloo, M.; White, P.J.; Ding, Z.; Kwok, R.; Lee, P.; Radda, G.K.; et al. Loss of BCAA Catabolism during Carcinogenesis Enhances mTORC1 Activity and Promotes Tumor Development and Progression. Cell Metab. 2019, 29, 1151–1165. [Google Scholar] [CrossRef]
- Wang, H.; Lu, J.; Chen, X.; Schwalbe, M.; Gorka, J.E.; Mandel, J.A.; Wang, J.; Goetzman, E.S.; Ranganathan, S.; Dobrowolski, S.F.; et al. Acquired deficiency of peroxisomal dicarboxylic acid catabolism is a metabolic vulnerability in hepatoblastoma. J. Biol. Chem. 2021, 296, 100283. [Google Scholar] [CrossRef]
- Maschietto, M.; Rodrigues, T.C.; Kashiwabara, A.Y.; de Araujo É, S.S.; Marques Aguiar, T.F.; da Costa, C.M.L.; da Cunha, I.W.; Dos Reis Vasques, L.; Cypriano, M.; Brentani, H.; et al. DNA methylation landscape of hepatoblastomas reveals arrest at early stages of liver differentiation and cancer-related alterations. Oncotarget 2017, 8, 97871–97889. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotation | Pathway | Fold Change (Tumour)/(Non-Tumour) | FDR-Corrected p-Value | VIP Value | Level of Identification * |
|---|---|---|---|---|---|
| 4-Trimethylammoniobutanoate | Fatty acid transport into/out of mitochondria | 0.57 | 6.53 × 10−3 | 1.3 | 2 |
| L-Carnitine | Fatty acid transport into/out of mitochondria | 0.21 | 3.66 × 10−5 | 12.3 | 2 |
| O-Acetylcarnitine | Fatty acid transport into/out of mitochondria | 0.30 | 6.58 × 10−5 | 4.8 | 2 |
| Propionylcarnitine | Fatty acid transport into/out of mitochondria | 0.07 | 5.97 × 10−6 | 4.5 | 2 |
| O-Butanoylcarnitine | Fatty acid transport into/out of mitochondria | 0.54 | 1.48 × 10−3 | 1.4 | 2 |
| Isovalerylcarnitine | Fatty acid transport into/out of mitochondria | 0.14 | 6.58 × 10−5 | 1.1 | 3 |
| O-Succinylcarnitine | Fatty acid transport into/out of mitochondria | 0.07 | 3.70 × 10−7 | 1.2 | 4 |
| 9,10-DiHOME | Cell signalling/fatty acid beta-oxidation | 0.03 | 1.58 × 10−7 | 1.0 | 4 |
| 1-Palmitoylglycerophosphocholine | Glycerophospholipid metabolism | 0.34 | 3.89 × 10−5 | 1.6 | 4 |
| sn-Glycero-3-Phosphocholine | Glycerophospholipid metabolism | 0.07 | 3.63 × 10−9 | 6.2 | 2 |
| sn-Glycero-3-phosphoethanolamine | Glycerophospholipid metabolism | 0.04 | 2.66 × 10−7 | 1.9 | 4 |
| sn-Glycerol 3-phosphate | Glycerophospholipid metabolism | 0.18 | 7.38 × 10−6 | 1.5 | 2 |
| 4-Oxoproline | Unknown | 0.62 | 2.10 × 10−3 | 2.8 | 4 |
| 5-Oxoproline | Glutathione metabolism | 0.47 | 4.40 × 10−5 | 1.7 | 2 |
| L-Glutathione (reduced) | Glutathione metabolism (an antioxidant) | 0.05 | 3.01 × 10−6 | 1.8 | 3 |
| L-Glutamate | Glutathione metabolism; alanine, aspartate and glutamate metabolism | 0.89 | 3.39 × 10−2 | 1.2 | 2 |
| L-Glutamine | Alanine, aspartate and glutamate metabolism | 0.53 | 8.86 × 10−5 | 1.1 | 2 |
| L-Alanine | Alanine, aspartate and glutamate metabolism | 0.31 | 5.22 × 10−5 | 1.6 | 2 |
| Creatine | Facilitates recycling of ATP; arginine and proline metabolism | 0.29 | 2.48 × 10−6 | 11.5 | 2 |
| D-Glucose | Glycolysis | 0.47 | 1.66 × 10−4 | 2.4 | 2 |
| Citric acid | TCA cycle | 0.59 | 5.94 × 10−3 | 1.9 | 2 |
| Succinate | TCA cycle | 0.68 | 4.24 × 10−3 | 2.1 | 2 |
| (S)-Malate | TCA cycle | 0.28 | 2.66 × 10−4 | 3.0 | 2 |
| Nicotinamide | The main source of NAD + (which is a major oxidising agent) | 0.45 | 1.39 × 10−4 | 3.9 | 2 |
| Betaine | One-carbon metabolism | 0.36 | 1.21 × 10−4 | 11.4 | 2 |
| Inosine | Nucleotide metabolism (purine) | 0.67 | 4.37 × 10−4 | 1.6 | 2 |
| Xanthine | Nucleotide metabolism (purine) | 0.30 | 7.42 × 10−7 | 1.6 | 2 |
| Uridine | Nucleotide metabolism (pyrimidine) | 0.46 | 1.51 × 10−4 | 1.2 | 2 |
| L-2-Aminoadipate | Lysine degradation | 0.60 | 2.55 × 10−2 | 1.3 | 3 |
| Norecasantalic acid | Unknown | 0.02 | 9.72 × 10−8 | 1.2 | 4 |
| 3-Acetamidopropanal | Unknown | 0.34 | 9.38 × 10−5 | 1.3 | 4 |
| D-Erythrose | Carbohydrate | 0.25 | 1.18 × 10−5 | 1.1 | 4 |
| Diazoxide | A drug | 0.04 | 1.44 × 10−5 | 3.1 | 3 |
| Dimethyl maleate | Unknown | 0.41 | 9.08 × 10−5 | 1.0 | 4 |
| Dimethyl maleate | Unknown | 0.60 | 2.55 × 10−2 | 1.3 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whitby, A.; Pabla, P.; Shastri, B.; Amugi, L.; Del Río-Álvarez, Á.; Kim, D.-H.; Royo, L.; Armengol, C.; Dandapani, M. Characterisation of Aberrant Metabolic Pathways in Hepatoblastoma Using Liquid Chromatography and Tandem Mass Spectrometry (LC-MS/MS). Cancers 2023, 15, 5182. https://doi.org/10.3390/cancers15215182
Whitby A, Pabla P, Shastri B, Amugi L, Del Río-Álvarez Á, Kim D-H, Royo L, Armengol C, Dandapani M. Characterisation of Aberrant Metabolic Pathways in Hepatoblastoma Using Liquid Chromatography and Tandem Mass Spectrometry (LC-MS/MS). Cancers. 2023; 15(21):5182. https://doi.org/10.3390/cancers15215182
Chicago/Turabian StyleWhitby, Alison, Pardeep Pabla, Bhoomi Shastri, Laudina Amugi, Álvaro Del Río-Álvarez, Dong-Hyun Kim, Laura Royo, Carolina Armengol, and Madhumita Dandapani. 2023. "Characterisation of Aberrant Metabolic Pathways in Hepatoblastoma Using Liquid Chromatography and Tandem Mass Spectrometry (LC-MS/MS)" Cancers 15, no. 21: 5182. https://doi.org/10.3390/cancers15215182
APA StyleWhitby, A., Pabla, P., Shastri, B., Amugi, L., Del Río-Álvarez, Á., Kim, D. -H., Royo, L., Armengol, C., & Dandapani, M. (2023). Characterisation of Aberrant Metabolic Pathways in Hepatoblastoma Using Liquid Chromatography and Tandem Mass Spectrometry (LC-MS/MS). Cancers, 15(21), 5182. https://doi.org/10.3390/cancers15215182