

Updated Clinical Evidence on the Role of Adipokines and Breast Cancer: A Review

 ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Adipose Tissue Microenvironment and Breast Cancer Cells
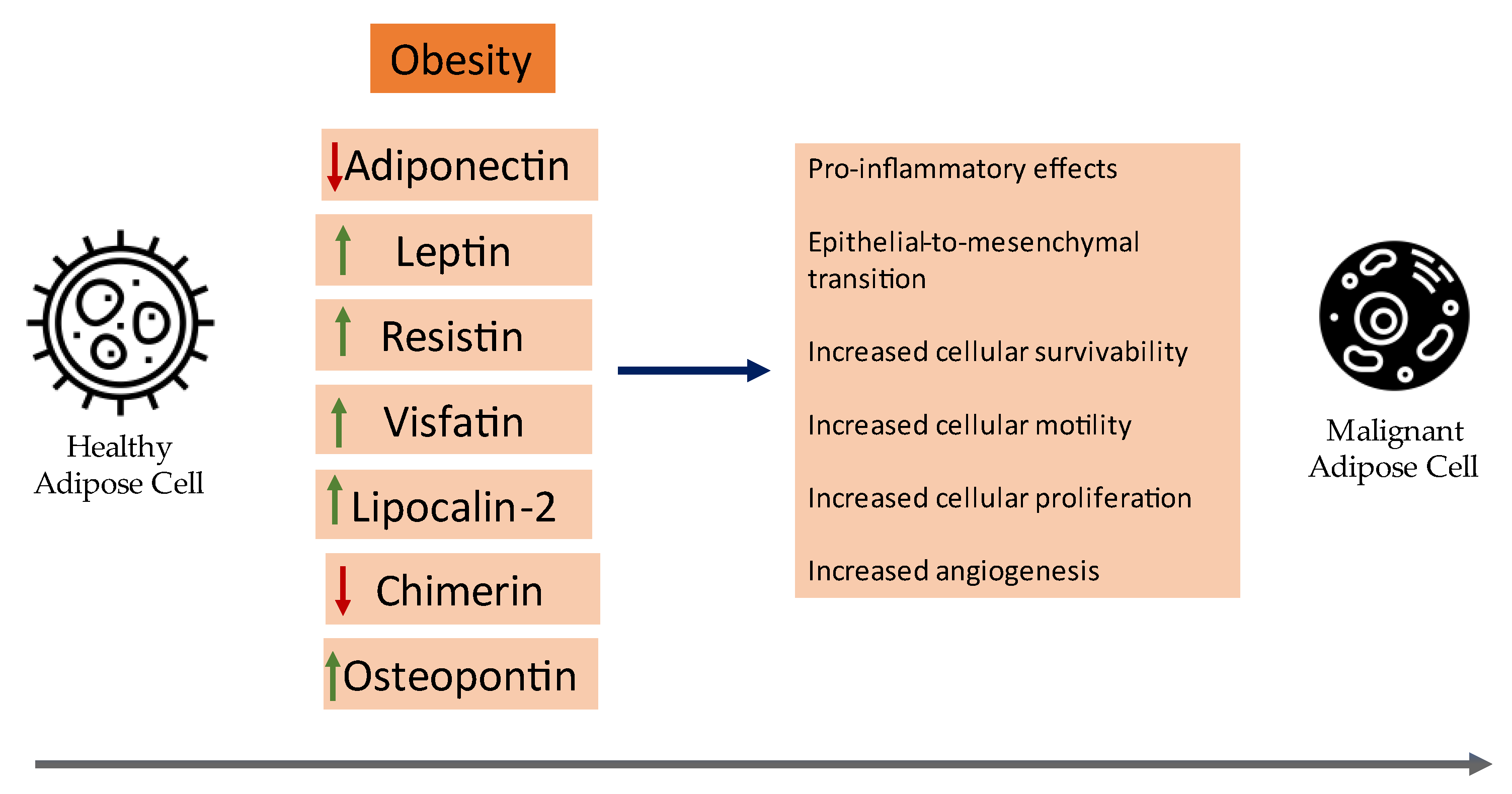
3. Adipokines
3.1. Adiponectin
3.2. Leptin
3.3. Resistin
3.4. Visfatin (eNampt)
3.5. Lipocalin-2
3.6. Chemerin
3.7. Osteopontin (OPN)

{kind=link}
| Meta-Analysis (Study) | Number of Studies | Number of Participants | Study Outcomes |
|---|---|---|---|
| Adiponectin | |||
| Gu L et al. Serum adiponectin in breast cancer: A meta-analysis. Medicine (Baltimore) 2018; 97:e11433 [30] | 31 eligible studies were included | Βreast cancer group (7388), Control Group (8491) | Lower serum adiponectin levels in BC cases, in pre-menopausal and post-menopausal women, especially among Asians but not in the Caucasian population. |
| Yu Z, Tang S, Ma H, Duan H, Zeng Y. Association of serum adiponectin with breast cancer: A meta-analysis of 27 case-control studies. Medicine (Baltimore). 2019 Feb;98(6):e14359 [31]. | 27 eligible studies were included | Βreast cancer group (7176), Control Group (8318) | Serum adiponectin was inversely associated with breast cancer. Decreased serum adiponectin levels in pre-menopausal women, post-menopausal status. In addition, low serum adiponectin levels in Asian women were more likely to be associated with breast cancer risk than in Caucasian women. |
| Yoon YS, Kwon AR, Lee YK, Oh SW. Circulating adipokines and risk of obesity related cancers: A systematic review and meta-analysis. Obes Res Clin Pract. 2019 Jul-Aug;13(4):329–339 [32] | 14 eligible studies were included | By cancer site and type, highest category of adiponectin was associated with decreased risk of breast cancer | |
| Leptin | |||
| Pan H, Deng LL, Cui JQ, Shi L, Yang YC, Luo JH, Qin D, Wang L. Association between serum leptin levels and breast cancer risk: An updated systematic review and meta-analysis. Medicine (Baltimore). 2018 Jul;97(27):e11345 [44]. | 35 eligible studies were included | Brest Cancer Group (6086) vs. Control Group (7158) | Serum leptin levels were related to breast cancer risk as demonstrated by calculations of the overall SMD = 0.46 (95% CI = 0.31–0.60, I2 = 93.5%). A subgroup analysis of BMI identified an association between breast cancer and serum leptin levels in patients who are overweight and obese (overweight: SMD = 0.35, 95% CI = 0.13–0.57, I2 = 88.1%; obesity: SMD = 1.38, 95% CI = 0.64–2.12, I2 = 89.6%). Additionally, menopausal status subgroup analysis revealed a significant association in post-menopausal women (SMD = 0.26, 95% CI = 0.12–0.40, I2 = 77.9%). Furthermore, we identified a significant association between breast cancer and serum leptin levels in Chinese women (SMD = 0.61, 95% CI = 0.44–0.79, I2 = 40.6%). |
| Yoon YS, Kwon AR, Lee YK, Oh SW. Circulating adipokines and risk of obesity related cancers: A systematic review and meta-analysis. Obes Res Clin Pract. 2019 Jul-Aug;13(4):329–339. [32] | 9 eligible studies were included | In the linear dose-response analysis by cancer type, each 5 ng/mL increase in leptin was not significantly associated with BC. Leptin was significantly associated with increased risk of cancer. | |
| Sayad S, Dastgheib SA, Farbod M, Asadian F, Karimi-Zarchi M, Salari S, Shaker SH, Sadeghizadeh-Yazdi J, Neamatzadeh H. Association of PON1, LEP and LEPR Polymorphisms with Susceptibility to Breast Cancer: A Meta-Analysis. Asian Pac J Cancer Prev. 2021 Aug 1;22(8):2323–2334. [43] | 12 studies on LEP rs7799039, and 14 studies on LEPR rs1137101 were selected. | 3444 cases and 3583 controls on LEP (leptin) rs7799039 polymorphism, 5330 cases and 6188 controls on LEPR(leptin receptor) rs1137101 polymorphism | LEP rs7799039 and LEPR rs1137101 polymorphisms were not associated with an increased risk for breast cancer. |
| Gui Y, Pan Q, Chen X, Xu S, Luo X, Chen L. The association between obesity related adipokines and risk of breast cancer: a meta-analysis. Oncotarget. 2017 May 13;8(43):75389–75399 [37]. | 46 eligible studies were included | Brest Cancer Group (6459) vs. Control Group (7155) | ER positive cases had significantly higher leptin levels than ER negative cases, Leptin and TNF-α levels were also significantly higher in lymph node metastasis (LNM) positive cases than in LNM negative cases, Leptin levels were significantly higher among post-menopausal cases than pre-menopausal cases |
| Resistin | |||
| Hashemi M, Bahari G, Tabasi F, Moazeni-Roodi A, Ghavami S. Association between rs1862513 and rs3745367 Genetic Polymorphisms of Resistin and Risk of Cancer: A Meta-Analysis. Asian Pac J Cancer Prev. 2018 Oct 26;19(10):2709–2716. [68] | 9 studies were included | 1951 cancer patients and 2295 healthy controls were included | The data revealed no correlation between the rs3745367 polymorphism and cancer risk. |
| Yoon YS, Kwon AR, Lee YK, Oh SW. Circulating adipokines and risk of obesity related cancers: A systematic review and meta-analysis. Obes Res Clin Pract. 2019 Jul-Aug;13(4):329–339 [32]. | 5 eligible studies were included | The highest vs. lowest meta-analysis showed no relationship between resistin and cancer risk, with high heterogeneity between studies. Exclusion of one study resulted in significance during sensitivity analysis. | |
| Gui Y, Pan Q, Chen X, Xu S, Luo X, Chen L. The association between obesity related adipokines and risk of breast cancer: a meta-analysis. Oncotarget. 2017 May 13;8(43):75389–75399. [37] | 6 studies were eligible | Brest Cancer Group (1236) vs. Control Group (1137) | Mean concentrations were higher than in the control group, but among Asian cases and not in non-Asian groups. |
| Visfatin (eNampt) | |||
| Gui Y, Pan Q, Chen X, Xu S, Luo X, Chen L. The association between obesity related adipokines and risk of breast cancer: a meta-analysis. Oncotarget. 2017 May 13;8(43):75389–75399. [37] | 3 studies were eligible | Brest Cancer Group (433) vs. Control Group (271) | Mean concentrations of visfatin were higher in cases than controls. There was no significant difference in the levels of visfatin between subjects with BMI > 25 kg/m2 and those with BMI < 25 kg/m2 |
| Ghaneialvar H, Shiri S, Kenarkoohi A, Fallah Vastani Z, Ahmadi A, Khorshidi A, Khooz R. Comparison of visfatin levels in patients with breast cancer and endometrial cancer with healthy individuals: A systematic review and meta-analysis. Health Sci Rep. 2022 Nov 18;5(6):e895. [51] | 10 studies were eligible | Breast Cancer group (260) Control Group (400) | Increased levels in breast cancer. |
| Mohammadi M, Mianabadi F, Mehrad-Majd H. Circulating visfatin levels and cancers risk: a systematic review and meta-analysis. J Cell Physiol. 2019;234(4):5011–5022. [69] | 27 studies were eligible | Breast Cancer group (2693) Control Group (3040) | Metaresults showed a significant higher level of visfatin in patients with cancer than in the controls |
| Osteopontin (OPN) | |||
| Hao C et al. Prognostic Value of Osteopontin Splice Variant-c Expression in Breast Cancers: A Meta-Analysis. Biomed Res Int 2016; 2016: 7310694 [67] | 10 time-to event studies were eligible | 1567 breast cancer patients | High level OPN indicated a poor outcome in the OS. High level OPN-c appeared to be more significantly associated with poor survival. Our analyses indicated that both OPN and OPN-c could be considered as prognostic markers for breast cancers |
| Lipocalin-2 (NGAL) | |||
| Wang Y et al. Neutrophil gelatinase-associated lipocalin protein as a biomarker in the diagnosis of breast cancer: A meta-analysis. Biomed Rep 2013;1:479–83 [70] | 4 studies were eligible | Breast Cancer Group (332). Control Group (142) | ROC curve (AUC) for Breast Cancer diagnosis was 0.90. Sensitivity: 64% (95% CI, 0.59–0.69). Specificity: 87% (95% CI, 0.81–0.92). |
| Adipokine | Proposed Implications in Carcinogenesis |
|---|---|
| Adiponectin | Decreased levels of adiponectin are associated with carcinogenesis. Protective effects through the AdipoR1 and AdipoR2 receptors. Downregulation of HIF-1. Activation of MPK. |
| Leptin | Activation of JAK/STAT and PI3k/AKT pathways that induce proliferation. Induction of angiogenesis through VEGF stimulation. Contributes to EMT that produces CAFs. |
| Resistin | Pro-glycolic adipokine that promotes inflammatory response via NF-κB and PI3K pathways. |
| Visfatin | Activates the ABL proto-oncogene 1 the STAT3 pathway and upregulates mRNA levels of cyclin D1 and CDK2. |
| Lipocalin-2 | Pro-inflammatory response, capable of inducing EMT. |
| Chemerin | Angiogenic and proliferative properties. Binds to the CMKLR-1 receptor. Increased levels seem to exert anti-oncogenic effects. |
| Osteopontin | Activation of integrins, pro-inflammatory adipokine. Associated with worse survival rates and mortality. |
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional Cloning of the Mouse Obese Gene and Its Human Homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, Regional, and National Prevalence of Overweight and Obesity in Children and Adults during 1980-2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.; LeRoith, D.; Gallagher, E.J. Diabetes, Obesity, and Breast Cancer. Endocrinology 2018, 159, 3801–3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Hajifathalian, K.; Ezzati, M.; Woodward, M.; Rimm, E.B.; Danaei, G.; Selmer, R.; Strand, B.H.; Dobson, A.; Hozawa, A.; et al. Metabolic Mediators of the Effects of Body-Mass Index, Overweight, and Obesity on Coronary Heart Disease and Stroke: A Pooled Analysis of 97 Prospective Cohorts with 1·8 Million Participants. Lancet 2014, 383, 970–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-Mass Index and Incidence of Cancer: A Systematic Review and Meta-Analysis of Prospective Observational Studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef]
- Goodwin, P.J.; Stambolic, V. Impact of the Obesity Epidemic on Cancer. Annu. Rev. Med. 2015, 66, 281–296. [Google Scholar] [CrossRef]
- Lyu, X.; Zhang, Q.; Fares, H.M.; Wang, Y.; Han, Y.; Sun, L. Contribution of Adipocytes in the Tumor Microenvironment to Breast Cancer Metabolism. Cancer Lett. 2022, 534, 215616. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Lu, P.; Beeraka, N.M.; Sukocheva, O.A.; Madhunapantula, S.R.V.; Liu, J.; Sinelnikov, M.Y.; Nikolenko, V.N.; Bulygin, K.V.; Mikhaleva, L.M.; et al. Mitochondrial Mutations and Mitoepigenetics: Focus on Regulation of Oxidative Stress-Induced Responses in Breast Cancers. Semin. Cancer Biol. 2022, 83, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, C.; Wang, X.; Sun, Y.; Zhang, J.; Chen, J.; Shi, Y. An Epigenetic Role of Mitochondria in Cancer. Cells 2022, 11, 2518. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2015, 16, 378. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Zhang, J.; Beeraka, N.M.; Tang, C.; Babayeva, Y.V.; Sinelnikov, M.Y.; Zhang, X.; Zhang, J.; Liu, J.; Reshetov, I.V.; et al. Advances in the Prevention and Treatment of Obesity-Driven Effects in Breast Cancers. Front. Oncol. 2022, 12, 2663. [Google Scholar] [CrossRef]
- Trayhurn, P. Endocrine and Signalling Role of Adipose Tissue: New Perspectives on Fat. Acta Physiol. Scand. 2005, 184, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, J.; Lv, W.; Zhao, C.; Xia, Y.; Wu, Y.; Zhang, Q. The Pleiotropic Roles of Adipocyte Secretome in Remodeling Breast Cancer. J. Exp. Clin. Cancer Res. 2022, 41, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hillers-Ziemer, L.E.; Kuziel, G.; Williams, A.E.; Moore, B.N.; Arendt, L.M. Breast Cancer Microenvironment and Obesity: Challenges for Therapy. Cancer Metastasis Rev. 2022, 41, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Bandini, E.; Rossi, T.; Gallerani, G.; Fabbri, F. Adipocytes and MicroRNAs Crosstalk: A Key Tile in the Mosaic of Breast Cancer Microenvironment. Cancers 2019, 11, 1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimta, A.A.; Tigu, A.B.; Muntean, M.; Cenariu, D.; Slaby, O.; Berindan-Neagoe, I. Molecular Links between Central Obesity and Breast Cancer. Int. J. Mol. Sci. 2019, 20, 5364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteside, T.L. The Tumor Microenvironment and Its Role in Promoting Tumor Growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef] [Green Version]
- Schäffler, A.; Schölmerich, J.; Buechler, C. Mechanisms of Disease: Adipokines and Breast Cancer—Endocrine and Paracrine Mechanisms That Connect Adiposity and Breast Cancer. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Crudele, L.; Piccinin, E.; Moschetta, A. Visceral Adiposity and Cancer: Role in Pathogenesis and Prognosis. Nutrients 2021, 13, 2101. [Google Scholar] [CrossRef]
- Jafari, N.; Kolla, M.; Meshulam, T.; Shafran, J.S.; Qiu, Y.; Casey, A.N.; Pompa, I.R.; Ennis, C.S.; Mazzeo, C.S.; Rabhi, N.; et al. Adipocyte-Derived Exosomes May Promote Breast Cancer Progression in Type 2 Diabetes. Sci. Signal. 2021, 14, 2467–2470. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, D.; Bae, S.J.; Ahn, S.G.; Jeong, J.; Park, W.C.; Ryu, Y.H.; Jeon, T.J.; Lee, Y.; Cha, Y.J.; et al. 18F-FDG Uptake of Visceral Adipose Tissue on Preoperative PET/CT as a Predictive Marker for Breast Cancer Recurrence. Sci. Rep. 2022, 12, 21109. [Google Scholar] [CrossRef]
- Lehr, S.; Hartwig, S.; Sell, H. Adipokines: A Treasure Trove for the Discovery of Biomarkers for Metabolic Disorders. Proteom. Clin. Appl. 2012, 6, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Nehme, R.; Diab-Assaf, M.; Decombat, C.; Delort, L.; Caldefie-Chezet, F. Targeting Adiponectin in Breast Cancer. Biomedicines 2022, 10, 2958. [Google Scholar] [CrossRef]
- Tsankof, A.; Tziomalos, K. Adiponectin: A Player in the Pathogenesis of Hormone-Dependent Cancers. Front. Endocrinol. 2022, 13, 2522. [Google Scholar] [CrossRef]
- Simone, V.; D’Avenia, M.; Argentiero, A.; Felici, C.; Rizzo, F.M.; De Pergola, G.; Silvestris, F. Obesity and Breast Cancer: Molecular Interconnections and Potential Clinical Applications. Oncologist 2016, 21, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelesidis, I.; Kelesidis, T.; Mantzoros, C.S. Adiponectin and Cancer: A Systematic Review. Br. J. Cancer 2006, 94, 1221–1225. [Google Scholar] [CrossRef]
- Georgiou, G.P.; Provatopoulou, X.; Kalogera, E.; Siasos, G.; Menenakos, E.; Zografos, G.C.; Gounaris, A. Serum Resistin Is Inversely Related to Breast Cancer Risk in Premenopausal Women. Breast 2016, 29, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Cao, C.; Fu, J.; Li, Q.; Li, D.H.; Chen, M.Y. Serum Adiponectin in Breast Cancer: A Meta-Analysis. Medicine 2018, 97, e11433. [Google Scholar] [CrossRef]
- Yu, Z.; Tang, S.; Ma, H.; Duan, H.; Zeng, Y.; Schaller, B. Association of Serum Adiponectin with Breast Cancer: A Meta-Analysis of 27 Case-Control Studies. Medicine 2019, 98, e14359. [Google Scholar] [CrossRef]
- Yoon, Y.S.; Kwon, A.R.; Lee, Y.K.; Oh, S.W. Circulating Adipokines and Risk of Obesity Related Cancers: A Systematic Review and Meta-Analysis. Obes. Res. Clin. Pract. 2019, 13, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Naimo, G.D.; Gelsomino, L.; Catalano, S.; Mauro, L.; Andò, S. Interfering Role of ERα on Adiponectin Action in Breast Cancer. Front. Endocrinol. 2020, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.Y.; Wang, M.; Ma, Z.B.; Yu, L.X.; Zhang, Q.; Gao, D.Z.; Wang, F.; Yu, Z.G. The Role of Adiponectin in Breast Cancer: A Meta-Analysis. PLoS ONE 2013, 8, e73183. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Cha, Y.J.; Koo, J.S. Adipocyte Biology in Breast Cancer: From Silent Bystander to Active Facilitator. Prog. Lipid Res. 2018, 69, 11–20. [Google Scholar] [CrossRef]
- Niu, J.; Jiang, L.; Guo, W.; Shao, L.; Liu, Y.; Wang, L. The Association between Leptin Level and Breast Cancer: A Meta-Analysis. PLoS ONE 2013, 8, e67349. [Google Scholar] [CrossRef] [Green Version]
- Gui, Y.; Pan, Q.; Chen, X.; Xu, S.; Luo, X.; Chen, L. The Association between Obesity Related Adipokines and Risk of Breast Cancer: A Meta-Analysis. Oncotarget 2017, 8, 75389–75399. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, C.; Tennekoon, K.H.; Karunanayake, E.H.; De Silva, K.; Amarasinghe, I.; Wijayasiri, A. Circulating Leptin, Soluble Leptin Receptor, Free Leptin Index, Visfatin and Selected Leptin and Leptin Receptor Gene Polymorphisms in Sporadic Breast Cancer. Endocr. J. 2017, 64, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Raut, P.K.; Choi, D.Y.; Kim, S.H.; Hong, J.T.; Kwon, T.K.; Jeong, J.H.; Park, P.H. Estrogen Receptor Signaling Mediates Leptin-Induced Growth of Breast Cancer Cells via Autophagy Induction. Oncotarget 2017, 8, 109417. [Google Scholar] [CrossRef]
- Catalano, S.; Mauro, L.; Marsico, S.; Giordano, C.; Rizza, P.; Rago, V.; Montanaro, D.; Maggiolini, M.; Panno, M.L.; Andó, S. Leptin Induces, via ERK1/ERK2 Signal, Functional Activation of Estrogen Receptor Alpha in MCF-7 Cells. J. Biol. Chem. 2004, 279, 19908–19915. [Google Scholar] [CrossRef] [Green Version]
- Tewari, S.; Vargas, R.; Reizes, O. The Impact of Obesity and Adipokines on Breast and Gynecologic Malignancies. Ann. N. Y. Acad. Sci. 2022, 1518, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Touvier, M.; Fezeu, L.; Ahluwalia, N.; Julia, C.; Charnaux, N.; Sutton, A.; Méjean, C.; Latino-Martel, P.; Hercberg, S.; Galan, P.; et al. Association between Prediagnostic Biomarkers of Inflammation and Endothelial Function and Cancer Risk: A Nested Case-Control Study. Am. J. Epidemiol. 2013, 177, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayad, S.; Dastgheib, S.A.; Farbod, M.; Asadian, F.; Karimi-Zarchi, M.; Salari, S.; Shaker, S.H.; Sadeghizadeh-Yazdi, J.; Neamatzadeh, H. Association of PON1, LEP and LEPR Polymorphisms with Susceptibility to Breast Cancer: A Meta-Analysis. Asian Pac. J. Cancer Prev. 2021, 22, 2323. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Deng, L.L.; Cui, J.Q.; Shi, L.; Yang, Y.C.; Luo, J.H.; Qin, D.; Wang, L. Association between Serum Leptin Levels and Breast Cancer Risk: An Updated Systematic Review and Meta-Analysis. Medicine 2018, 97, e11345. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, E.; Piperigkou, Z.; Karamanos, N.K.; Zolota, V. Altered Adipokine Expression in Tumor Microenvironment Promotes Development of Triple Negative Breast Cancer. Cancers 2022, 14, 4139. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Hung, A.C.; Wu, Y.C.; Lo, S.; Da Chen, H.; Chen, Y.K.; Hsieh, Y.C.; Hu, S.C.S.; Hou, M.F.; Yuan, S.S.F. ADSCs Stimulated by Resistin Promote Breast Cancer Cell Malignancy via CXCL5 in a Breast Cancer Coculture Model. Sci. Rep. 2022, 12, 15437. [Google Scholar] [CrossRef]
- Lee, J.O.; Kim, N.; Lee, H.J.; Lee, Y.W.; Kim, S.J.; Park, S.H.; Kim, H.S. Resistin, a Fat-Derived Secretory Factor, Promotes Metastasis of MDA-MB-231 Human Breast Cancer Cells through ERM Activation. Sci. Rep. 2016, 6, 18923. [Google Scholar] [CrossRef] [Green Version]
- Díez, J.J.; Iglesias, P.; Fernández-Reyes, M.J.; Aguilera, A.; Bajo, M.A.; Alvarez-Fidalgo, P.; Codoceo, R.; Selgas, R. Serum Concentrations of Leptin, Adiponectin and Resistin, and Their Relationship with Cardiovascular Disease in Patients with End-Stage Renal Disease. Clin. Endocrinol. 2005, 62, 242–249. [Google Scholar] [CrossRef]
- Luhn, P.; Dallal, C.M.; Weiss, J.M.; Black, A.; Huang, W.Y.; Lacey, J.V.; Hayes, R.B.; Stanczyk, F.Z.; Wentzensen, N.; Brinton, L.A. Circulating Adipokine Levels and Endometrial Cancer Risk in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1304–1312. [Google Scholar] [CrossRef] [Green Version]
- Dalamaga, M.; Karmaniolas, K.; Papadavid, E.; Pelekanos, N.; Sotiropoulos, G.; Lekka, A. Elevated Serum Visfatin/Nicotinamide Phosphoribosyl-Transferase Levels Are Associated with Risk of Postmenopausal Breast Cancer Independently from Adiponectin, Leptin, and Anthropometric and Metabolic Parameters. Menopause 2011, 18, 1198–1204. [Google Scholar] [CrossRef]
- Ghaneialvar, H.; Shiri, S.; Kenarkoohi, A.; Fallah Vastani, Z.; Ahmadi, A.; Khorshidi, A.; Khooz, R. Comparison of Visfatin Levels in Patients with Breast Cancer and Endometrial Cancer with Healthy Individuals: A Systematic Review and Meta-analysis. Health Sci. Rep. 2022, 5, e895. [Google Scholar] [CrossRef] [PubMed]
- Villodre, E.S.; Hu, X.; Larson, R.; Finetti, P.; Gomez, K.; Balema, W.; Stecklein, S.R.; Santiago-Sanchez, G.; Krishnamurthy, S.; Song, J.; et al. Lipocalin 2 Promotes Inflammatory Breast Cancer Tumorigenesis and Skin Invasion. Mol. Oncol. 2021, 15, 2752–2765. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bielenberg, D.R.; Rodig, S.J.; Doiron, R.; Clifton, M.C.; Kung, A.L.; Strong, R.K.; Zurakowski, D.; Moses, M.A. Lipocalin 2 Promotes Breast Cancer Progression. Proc. Natl. Acad. Sci. USA 2009, 106, 3913–3918. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.H.; Wu, M.M.; Chan, M.W.Y.; Pu, Y.S.; Chen, C.J.; Lee, T.C. Neutrophil Gelatinase-Associated Lipocalin (NGAL): A Promising Biomarker for Cancer Diagnosis and A Potential Target for Cancer Therapeutics. J. Cell Sci. Mol. Biol. 2014, 88, 1549–1559. [Google Scholar] [CrossRef]
- Abas, A.S.M.; Sherif, M.H.; Elmoneam Farag, S.A. Diagnostic and Prognostic Role of Serum Omentin and NGAL Levels in Egyptian Breast Cancer Patients. Int. J. Breast Cancer 2022, 2022, 5971981. [Google Scholar] [CrossRef]
- Wei, C.T.; Tsai, I.T.; Wu, C.C.; Hung, W.C.; Hsuan, C.F.; Yu, T.H.; Hsu, C.C.; Houng, J.Y.; Chung, F.M.; Lee, Y.J.; et al. Elevated Plasma Level of Neutrophil Gelatinase-Associated Lipocalin (NGAL) in Patients with Breast Cancer. Int. J. Med. Sci. 2021, 18, 2689–2696. [Google Scholar] [CrossRef]
- Kurozumi, S.; Alsaeed, S.; Orah, N.; Miligy, I.M.; Joseph, C.; Aljohani, A.; Toss, M.S.; Fujii, T.; Shirabe, K.; Green, A.R.; et al. Clinicopathological Significance of Lipocalin 2 Nuclear Expression in Invasive Breast Cancer. Breast Cancer Res. Treat. 2020, 179, 557–564. [Google Scholar] [CrossRef]
- Santiago-Sánchez, G.S.; Noriega-Rivera, R.; Hernández-O’farrill, E.; Valiyeva, F.; Quiñones-Diaz, B.; Villodre, E.S.; Debeb, B.G.; Rosado-Albacarys, A.; Vivas-Mejía, P.E. Targeting Lipocalin-2 in Inflammatory Breast Cancer Cells with Small Interference RNA and Small Molecule Inhibitors. Int. J. Mol. Sci. 2021, 22, 8581. [Google Scholar] [CrossRef]
- Wittamer, V.; Franssen, J.D.; Vulcano, M.; Mirjolet, J.F.; Le Poul, E.; Migeotte, I.; Brézillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific Recruitment of Antigen-Presenting Cells by Chemerin, a Novel Processed Ligand from Human Inflammatory Fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef]
- Lu, L.-L.; Sun, J.; Lai, J.J.; Jiang, Y.; Bai, L.H.; Zhang, L. Da Neuron-Glial Antigen 2 Overexpression in Hepatocellular Carcinoma Predicts Poor Prognosis. World J. Gastroenterol. 2015, 21, 6649–6659. [Google Scholar] [CrossRef]
- Zhao, S.; Li, C.; Ye, Y.-B.; Peng, F.; Chen, Q. Expression of Chemerin Correlates With a Favorable Prognosis in Patients with Non-Small Cell Lung Cancer. Lab. Med. 2011, 42, 553–557. [Google Scholar] [CrossRef]
- El-Sagheer, G.; Gayyed, M.; Ahmad, A.; El-Fattah, A.A.; Mohamed, M. Expression of Chemerin Correlates with a Poor Prognosis in Female Breast Cancer Patients. Breast Cancer 2018, 10, 169. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Zhu, X.; Lin, Z.; Luo, L.; Wen, D. The Potential Value of Serum Chemerin in Patients with Breast Cancer. Sci. Rep. 2021, 11, 6564. [Google Scholar] [CrossRef]
- Pachynski, R.K.; Wang, P.; Salazar, N.; Zheng, Y.; Nease, L.; Rosalez, J.; Leong, W.I.; Virdi, G.; Rennier, K.; Shin, W.J.; et al. Chemerin Suppresses Breast Cancer Growth by Recruiting Immune Effector Cells into the Tumor Microenvironment. Front. Immunol. 2019, 10, 983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Infantes, D.; White, U.A.; Elks, C.M.; Morrison, R.F.; Gimble, J.M.; Considine, R.V.; Ferrante, A.W.; Ravussin, E.; Stephens, J.M. Oncostatin m Is Produced in Adipose Tissue and Is Regulated in Conditions of Obesity and Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2014, 99, E217–E225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinetti, A.; Bajetta, E.; Ferrari, L.; Zilembo, N.; Seregni, E.; Del Vecchio, M.; Longarini, R.; La Torre, I.; Toffolatti, L.; Paleari, D.; et al. Osteoprotegerin and Osteopontin Serum Values in Postmenopausal Advanced Breast Cancer Patients Treated with Anastrozole. Endocr. Relat. Cancer 2004, 11, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, C.; Wang, Z.; Gu, Y.; Jiang, W.G.; Cheng, S. Prognostic Value of Osteopontin Splice Variant-c Expression in Breast Cancers: A Meta-Analysis. BioMed Res. Int. 2016, 2016, 7310694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemi, M.; Bahari, G.; Tabasi, F.; Moazeni-Roodi, A.; Ghavami, S. Association between rs1862513 and rs3745367 Genetic Polymorphisms of Resistin and Risk of Cancer: A Meta-Analysis. Asian Pac. J. Cancer Prev. 2018, 19, 2709–2716. [Google Scholar]
- Mohammadi, M.; Mianabadi, F.; Mehrad-Majd, H. Circulating visfatin levels and cancers risk: A systematic review and meta-analysis. J. Cell. Physiol. 2019, 234, 5011–5022. [Google Scholar] [CrossRef]
- Wang, Y.U.; Zeng, T. Neutrophil gelatinase-associated lipocalin protein as a biomarker in the diagnosis of breast cancer: A meta-analysis. Biomed. Rep. 2013, 1, 479–483. [Google Scholar] [CrossRef] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verras, G.-I.; Tchabashvili, L.; Chlorogiannis, D.-D.; Mulita, F.; Argentou, M.-I. Updated Clinical Evidence on the Role of Adipokines and Breast Cancer: A Review. Cancers 2023, 15, 1572. https://doi.org/10.3390/cancers15051572
Verras G-I, Tchabashvili L, Chlorogiannis D-D, Mulita F, Argentou M-I. Updated Clinical Evidence on the Role of Adipokines and Breast Cancer: A Review. Cancers. 2023; 15(5):1572. https://doi.org/10.3390/cancers15051572
Chicago/Turabian StyleVerras, Georgios-Ioannis, Levan Tchabashvili, David-Dimitris Chlorogiannis, Francesk Mulita, and Maria-Ioanna Argentou. 2023. "Updated Clinical Evidence on the Role of Adipokines and Breast Cancer: A Review" Cancers 15, no. 5: 1572. https://doi.org/10.3390/cancers15051572
APA StyleVerras, G. -I., Tchabashvili, L., Chlorogiannis, D. -D., Mulita, F., & Argentou, M. -I. (2023). Updated Clinical Evidence on the Role of Adipokines and Breast Cancer: A Review. Cancers, 15(5), 1572. https://doi.org/10.3390/cancers15051572