
Identifying the Spatial Architecture That Restricts the Proximity of CD8+ T Cells to Tumor Cells in Pancreatic Ductal Adenocarcinoma
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
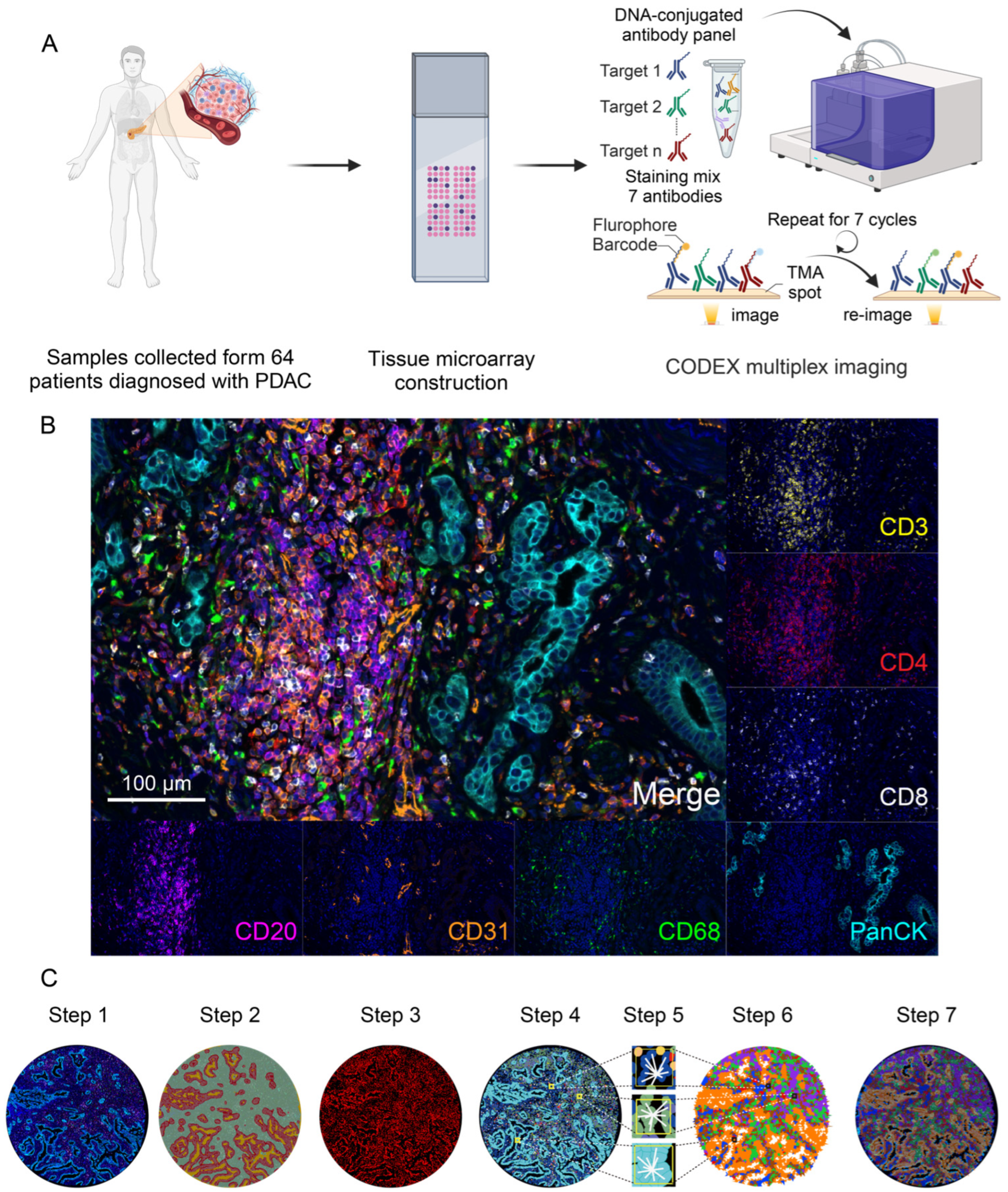
2.1. Patient Samples Collection and Tissue Microarray Preparation
2.2. Multiplex CODEX Staining and Image Acquisition
2.3. Image Analysis
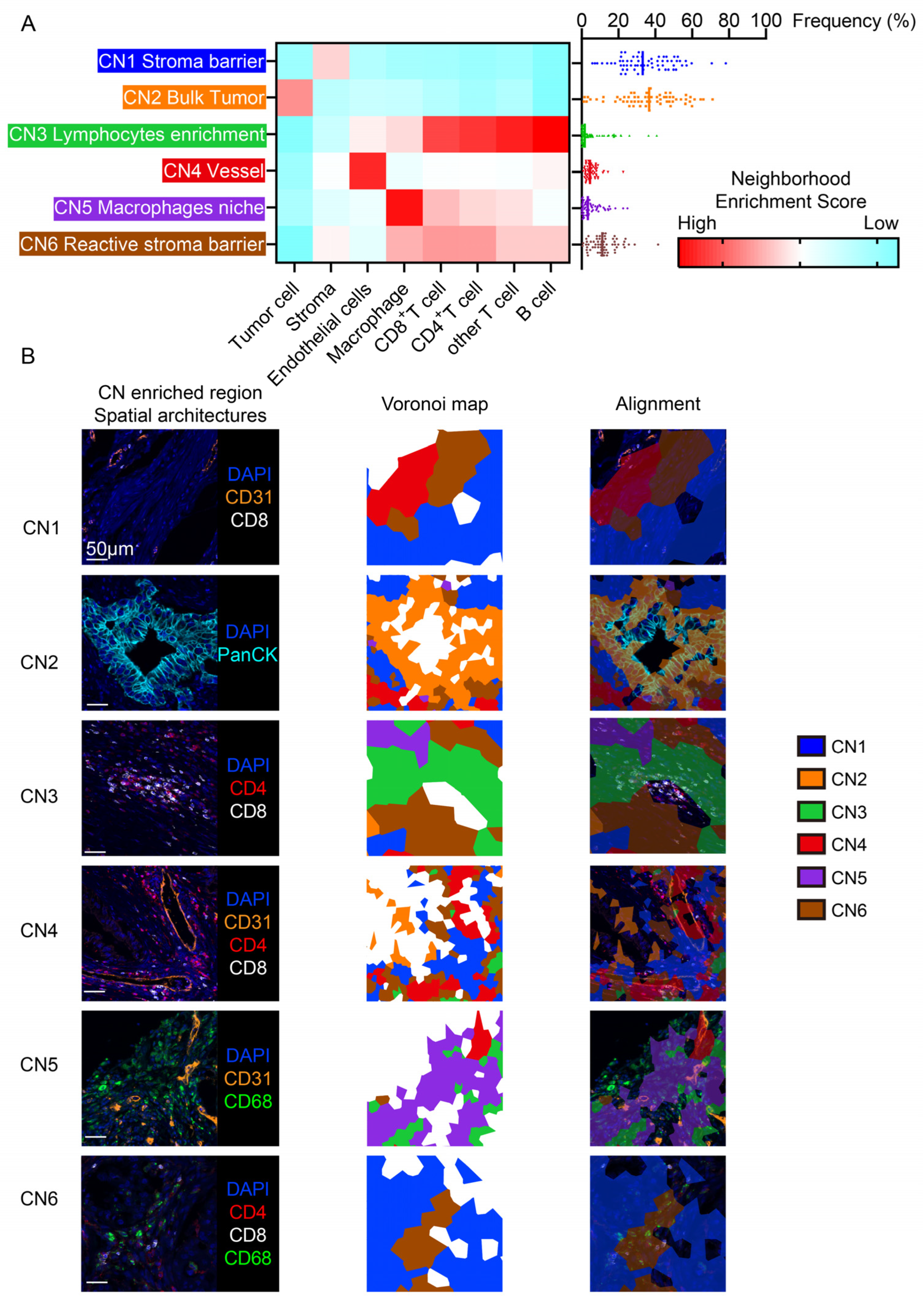
2.4. Cellular Neighborhood Analysis
2.5. Calculation of Spatial Distances and Spatial Score between Cell Types
its nearest cell with identity B)
2.6. Statistical Analysis
3. Results
3.1. CODEX Multiplex Imaging Characterized the Landscape of the PDAC Microenvironment
3.2. The Cellular Neighborhood Algorithm Revealed Distinct Functional Spatial Architectures
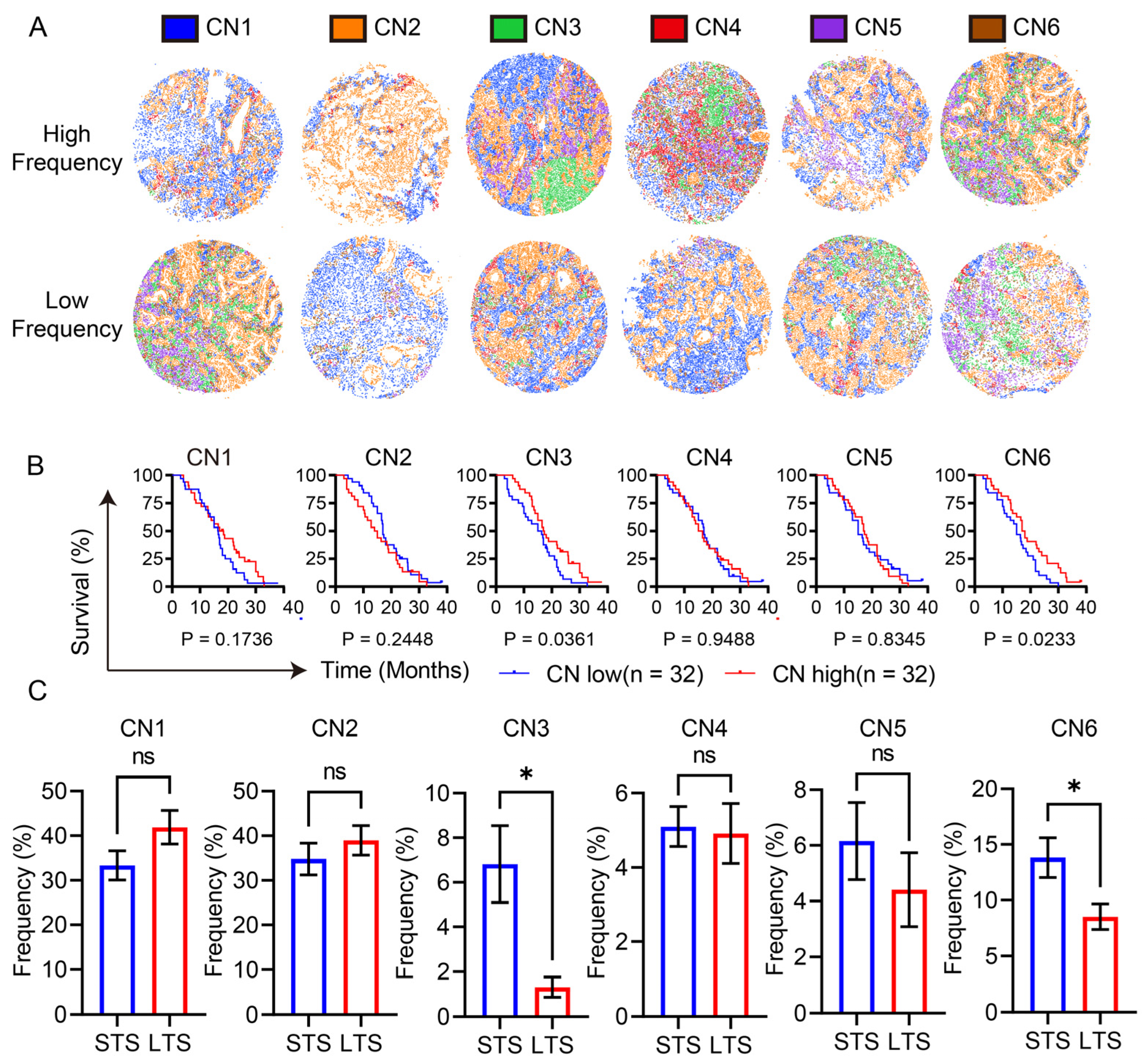
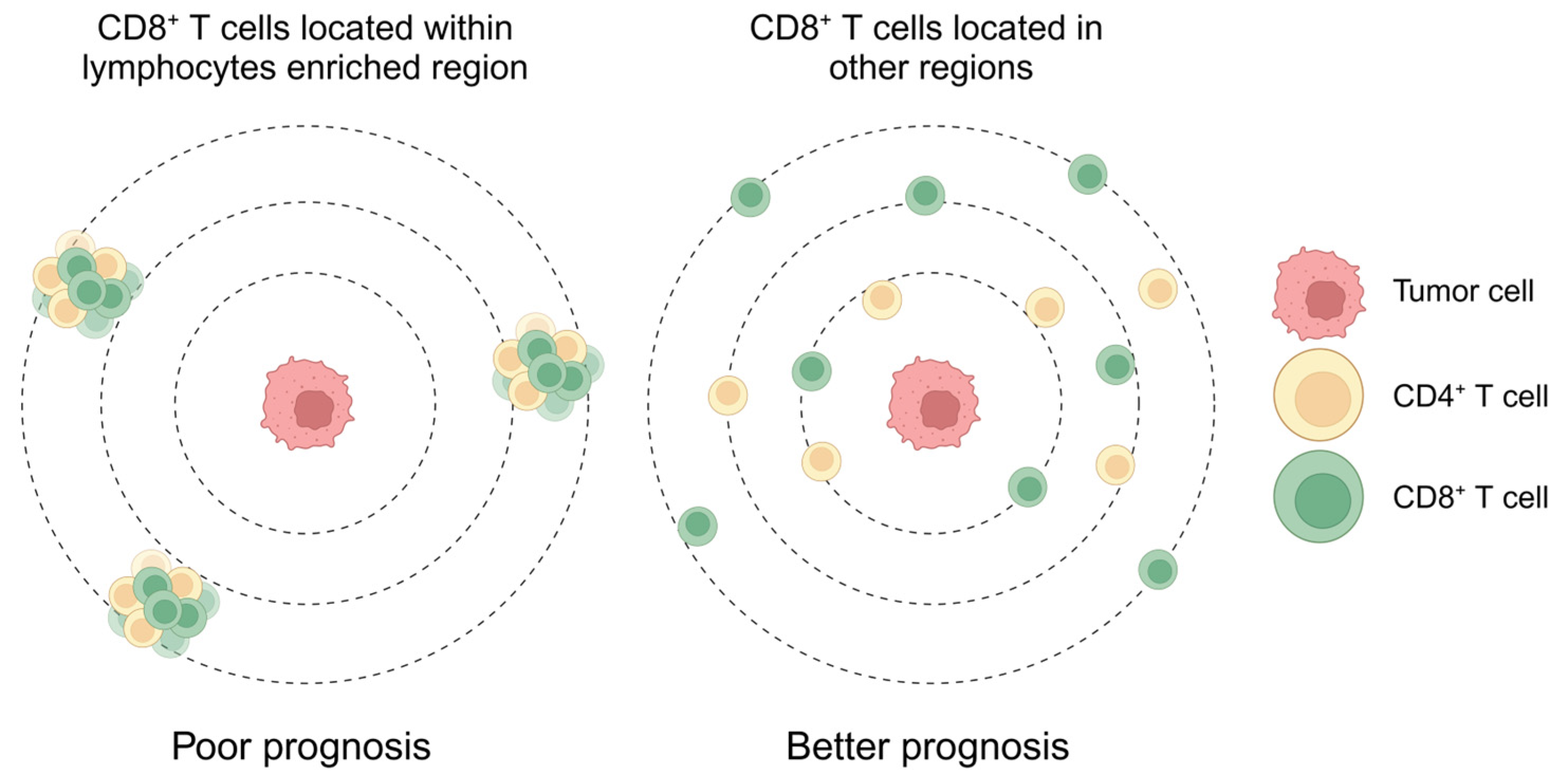
3.3. Lymphocyte Enrichment and the Reactive Stroma Barrier Heralded a Poor Prognosis
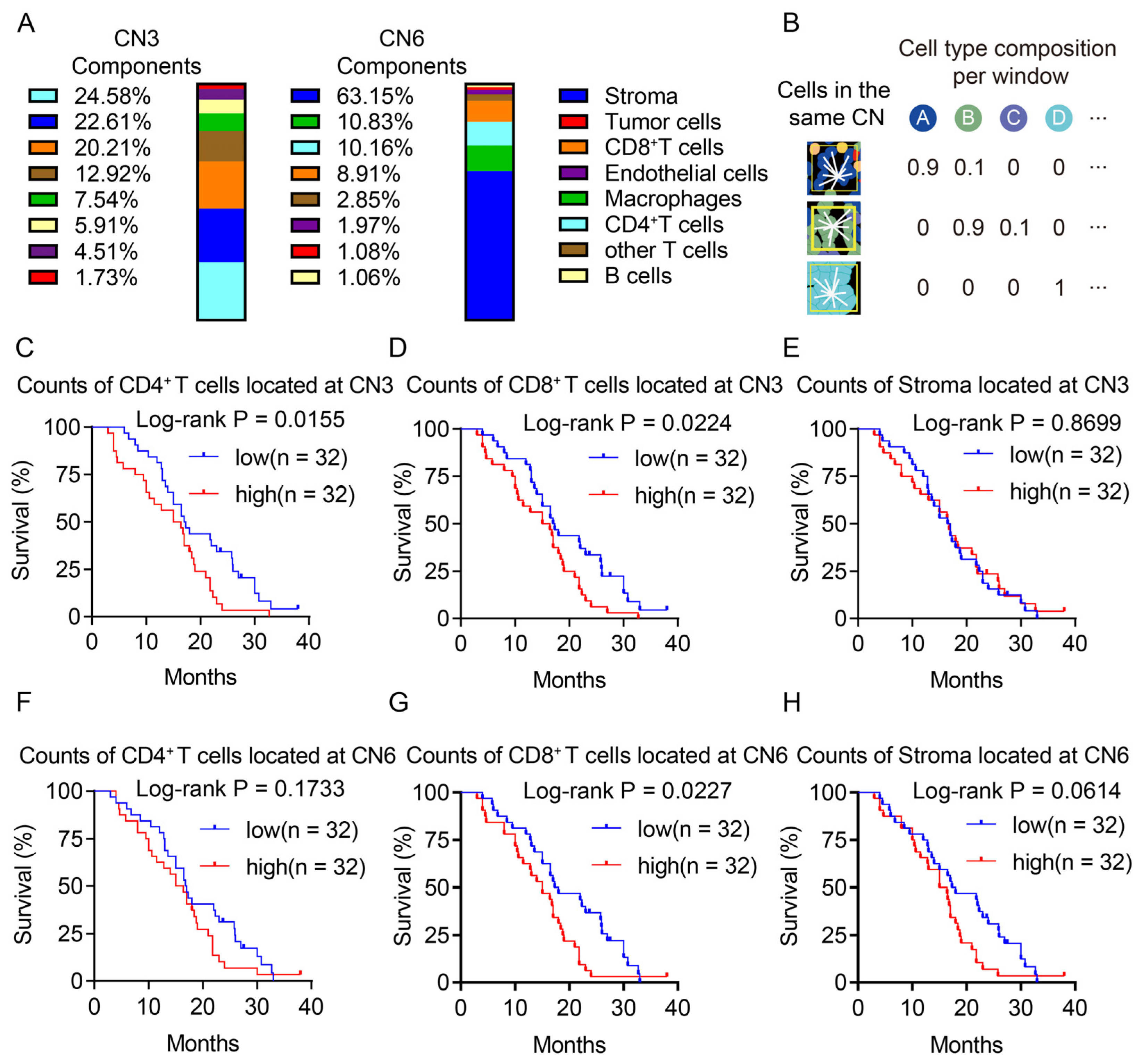
3.4. Both CD4+ T Cells and CD8+ T Cells in the Same Spatial Architectures was Associated with a Poor Prognosis
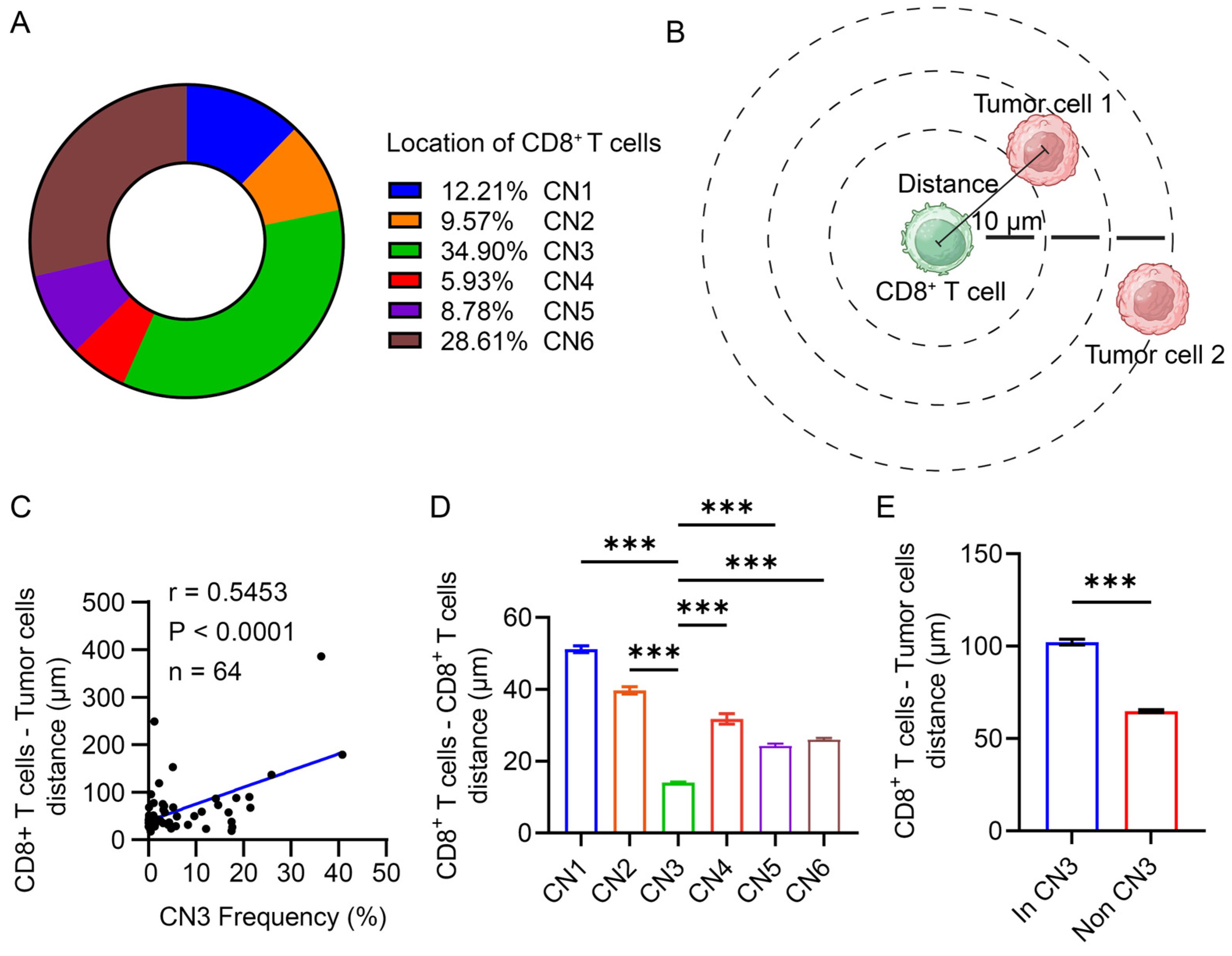
3.5. CD8+ T Cells Were Restricted in the Lymphocyte Enrichment Region
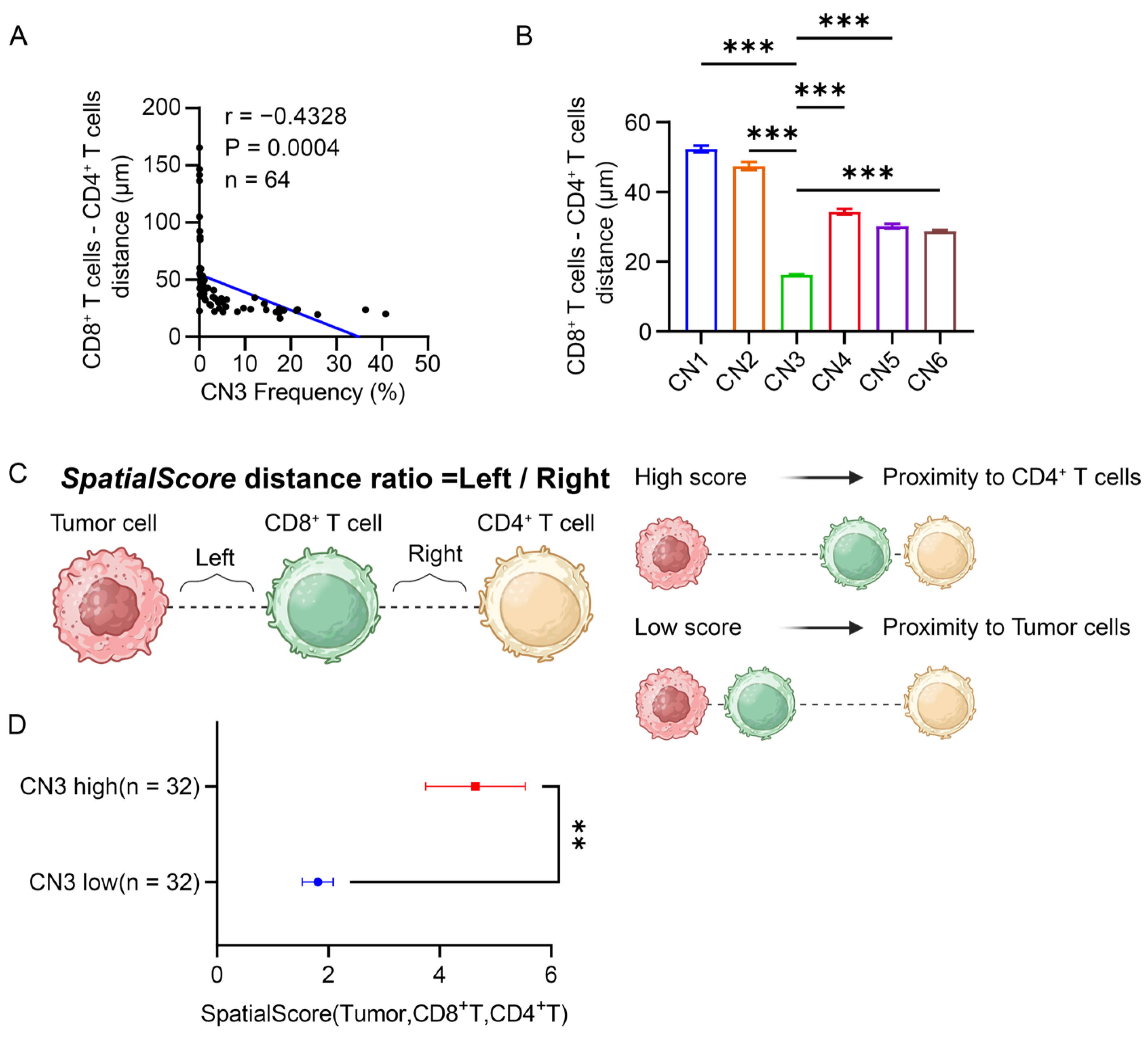
3.6. CD4+ T Cells Restricted the Proximity of CD8+ T Cells to the Tumor in the Lymphocyte Enrichment Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, D.S.; Mellman, I. Elements of Cancer Immunity and the Cancer-Immune Set Point. Nature 2017, 541, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Joyce, J.A.; Fearon, D.T. T Cell Exclusion, Immune Privilege, and the Tumor Microenvironment. Science 2015, 348, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Mariathasan, S.; Turley, S.J.; Nickles, D.; Castiglioni, A.; Yuen, K.; Wang, Y.; Kadel, E.E., III; Koeppen, H.; Astarita, J.L.; Cubas, R.; et al. Tgfbeta Attenuates Tumour Response to Pd-L1 Blockade by Contributing to Exclusion of T Cells. Nature 2018, 554, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Desbois, M.; Udyavar, A.R.; Ryner, L.; Kozlowski, C.; Guan, Y.; Durrbaum, M.; Lu, S.; Fortin, J.P.; Koeppen, H.; Ziai, J.; et al. Integrated Digital Pathology and Transcriptome Analysis Identifies Molecular Mediators of T-Cell Exclusion in Ovarian Cancer. Nat. Commun. 2020, 11, 5583. [Google Scholar] [CrossRef]
- Carstens, J.L.; Correa de Sampaio, P.; Yang, D.; Barua, S.; Wang, H.; Rao, A.; Allison, J.P.; LeBleu, V.S.; Kalluri, R. Spatial Computation of Intratumoral T Cells Correlates with Survival of Patients with Pancreatic Cancer. Nat. Commun. 2017, 8, 15095. [Google Scholar]
- Bockorny, B.; Grossman, J.E.; Hidalgo, M. Facts and Hopes in Immunotherapy of Pancreatic Cancer. Clin. Cancer Res. 2022, 28, 4606–4617. [Google Scholar] [CrossRef] [PubMed]
- Mashima, K.; Kimura, T.; Huang, W.; Yano, K.; Ashida, Y.; Yamagata, Y.; Miyazaki, K.; Yamashita, J.; Horio, T. Multiple Forms of Growth Inhibitors Secreted from Cultured Rat Liver Cells: Purification and Characterization. J. Biochem. 1988, 103, 1020–1026. [Google Scholar] [CrossRef] [PubMed]
- Black, S.; Phillips, D.; Hickey, J.W.; Kennedy-Darling, J.; Venkataraaman, V.G.; Samusik, N.; Goltsev, Y.; Schürch, C.M.; Nolan, G.P. Codex Multiplexed Tissue Imaging with DNA-Conjugated Antibodies. Nat. Protoc. 2021, 16, 3802–3835. [Google Scholar] [CrossRef]
- Schürch, C.M.; Bhate, S.S.; Barlow, G.L.; Phillips, D.J.; Noti, L.; Zlobec, I.; Chu, P.; Black, S.; Demeter, J.; McIlwain, D.R.; et al. Coordinated Cellular Neighborhoods Orchestrate Antitumoral Immunity at the Colorectal Cancer Invasive. Front. Cell 2020, 182, 1341–1359.e19. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, Y.; Samusik, N.; Kennedy-Darling, J.; Bhate, S.; Hale, M.; Vazquez, G.; Black, S.; Nolan, G.P. Deep Profiling of Mouse Splenic Architecture with Codex Multiplexed Imaging. Cell 2018, 174, 968–981.e15. [Google Scholar] [CrossRef]
- Walsh, L.A.; Quail, D.F. Decoding the Tumor Microenvironment with Spatial Technologies. Nat. Immunol. 2023, 24, 1982–1993. [Google Scholar] [CrossRef]
- Goltsev, Y.; Nolan, G. Codex Multiplexed Tissue Imaging. Nat. Rev. Immunol. 2023, 23, 613. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, M.; Deng, Y.; Su, G.; Enninful, A.; Guo, C.C.; Tebaldi, T.; Zhang, D.; Kim, D.; Bai, Z.; et al. High-Spatial-Resolution Multi-Omics Sequencing Via Deterministic Barcoding in Tissue. Cell 2020, 183, 1665–1681.e18. [Google Scholar] [CrossRef] [PubMed]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the Tumor Immune Microenvironment (Time) for Effective Therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.J.; Jaffee, E.M.; Zheng, L. The Tumour Microenvironment in Pancreatic Cancer—Clinical Challenges and Opportunities. Nat. Rev. Clin. Oncol. 2020, 17, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Nevala-Plagemann, C.; Hidalgo, M.; Garrido-Laguna, I. From State-of-the-Art Treatments to Novel Therapies for Advanced-Stage Pancreatic Cancer. Nat. Rev. Clin. Oncol. 2020, 17, 108–123. [Google Scholar] [CrossRef]
- Seo, Y.D.; Jiang, X.; Sullivan, K.M.; Jalikis, F.G.; Smythe, K.S.; Abbasi, A.; Vignali, M.; Park, J.O.; Daniel, S.K.; Pollack, S.M.; et al. Mobilization of Cd8(+) T Cells Via Cxcr4 Blockade Facilitates Pd-1 Checkpoint Therapy in Human Pancreatic Cancer. Clin. Cancer Res. 2019, 25, 3934–3945. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Todd, L.; Huang, L.; Noguera-Ortega, E.; Lu, Z.; Huang, L.; Kopp, M.; Li, Y.; Pattada, N.; Zhong, W.; et al. Desmoplastic Stroma Restricts T Cell Extravasation and Mediates Immune Exclusion and Immunosuppression in Solid Tumors. Nat. Commun. 2023, 14, 5110. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.; Wang, L.S.; Scholler, J.; Monslow, J.; Avery, D.; Newick, K.; O’Brien, S.; Evans, R.A.; Bajor, D.J.; Clendenin, C.; et al. Tumor-Promoting Desmoplasia Is Disrupted by Depleting Fap-Expressing Stromal Cells. Cancer Res. 2015, 75, 2800–2810. [Google Scholar] [CrossRef] [PubMed]
- Bandura, D.R.; Baranov, V.I.; Ornatsky, O.I.; Antonov, A.; Kinach, R.; Lou, X.; Pavlov, S.; Vorobiev, S.; Dick, J.E.; Tanner, S.D. Mass Cytometry: Technique for Real Time Single Cell Multitarget Immunoassay Based on Inductively Coupled Plasma Time-of-Flight Mass Spectrometry. Anal. Chem. 2009, 81, 6813–6822. [Google Scholar] [CrossRef]
- Agasti, S.S.; Wang, Y.; Schueder, F.; Sukumar, A.; Jungmann, R.; Yin, P. DNA-Barcoded Labeling Probes for Highly Multiplexed Exchange-Paint Imaging. Chem. Sci. 2017, 8, 3080–3091. [Google Scholar] [CrossRef]
- Tsujikawa, T.; Kumar, S.; Borkar, R.N.; Azimi, V.; Thibault, G.; Chang, Y.H.; Balter, A.; Kawashima, R.; Choe, G.; Sauer, D.; et al. Quantitative Multiplex Immunohistochemistry Reveals Myeloid-Inflamed Tumor-Immune Complexity Associated with Poor Prognosis. Cell Rep. 2017, 19, 203–217. [Google Scholar] [CrossRef]
- Stoltzfus, C.R.; Sivakumar, R.; Kunz, L.; Pope, B.E.O.; Menietti, E.; Speziale, D.; Adelfio, R.; Bacac, M.; Colombetti, S.; Perro, M.; et al. Multi-Parameter Quantitative Imaging of Tumor Microenvironments Reveals Perivascular Immune Niches Associated with Anti-Tumor Immunity. Front. Immunol. 2021, 12, 726492. [Google Scholar] [CrossRef] [PubMed]
- Abdulrahman, Z.; Santegoets, S.J.; Sturm, G.; Charoentong, P.; Ijsselsteijn, M.E.; Somarakis, A.; Hollt, T.; Finotello, F.; Trajanoski, Z.; van Egmond, S.L.; et al. Tumor-Specific T Cells Support Chemokine-Driven Spatial Organization of Intratumoral Immune Microaggregates Needed for Long Survival. J. Immunother. Cancer 2022, 10, e004346. [Google Scholar] [CrossRef] [PubMed]
- Becht, E.; McInnes, L.; Healy, J.; Dutertre, C.A.; Kwok, I.W.H.; Ng, L.G.; Ginhoux, F.; Newell, E.W. Dimensionality Reduction for Visualizing Single-Cell Data Using Umap. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Weed, D.T.; Zilio, S.; McGee, C.; Marnissi, B.; Sargi, Z.; Franzmann, E.; Thomas, G.; Leibowitz, J.; Nicolli, E.; Arnold, D.; et al. The Tumor Immune Microenvironment Architecture Correlates with Risk of Recurrence in Head and Neck Squamous Cell Carcinoma. Cancer Res. 2023, 83, 3886–3900. [Google Scholar] [CrossRef] [PubMed]
- Pitzalis, C.; Jones, G.W.; Bombardieri, M.; Jones, S.A. Ectopic Lymphoid-Like Structures in Infection, Cancer and Autoimmunity. Nat. Rev. Immunol. 2014, 14, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Drayton, D.L.; Ying, X.; Lee, J.; Lesslauer, W.; Ruddle, N.H. Ectopic Lt Alpha Beta Directs Lymphoid Organ Neogenesis with Concomitant Expression of Peripheral Node Addressin and a Hev-Restricted Sulfotransferase. J. Exp. Med. 2003, 197, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; Pitcher, L.A.; Sullivan, J.M.; Mitsdoerffer, M.; Acton, S.E.; Franz, B.; Wucherpfennig, K.; Turley, S.; Carroll, M.C.; Sobel, R.A.; et al. Th17 Cells Induce Ectopic Lymphoid Follicles in Central Nervous System Tissue Inflammation. Immunity 2011, 35, 986–996. [Google Scholar] [CrossRef] [PubMed]
- Link, A.; Hardie, D.L.; Favre, S.; Britschgi, M.R.; Adams, D.H.; Sixt, M.; Cyster, J.G.; Buckley, C.D.; Luther, S.A. Association of T-Zone Reticular Networks and Conduits with Ectopic Lymphoid Tissues in Mice and Humans. Am. J. Pathol. 2011, 178, 1662–1675. [Google Scholar] [CrossRef]
- Luther, S.A.; Bidgol, A.; Hargreaves, D.C.; Schmidt, A.; Xu, Y.; Paniyadi, J.; Matloubian, M.; Cyster, J.G. Differing Activities of Homeostatic Chemokines Ccl19, Ccl21, and Cxcl12 in Lymphocyte and Dendritic Cell Recruitment and Lymphoid Neogenesis. J. Immunol. 2002, 169, 424–433. [Google Scholar] [CrossRef]
- Yousuf, S.; Qiu, M.; von Voithenberg, L.V.; Hulkkonen, J.; Macinkovic, I.; Schulz, A.R.; Hartmann, D.; Mueller, F.; Mijatovic, M.; Ibberson, D.; et al. Spatially Resolved Multi-Omics Single-Cell Analyses Inform Mechanisms of Immune Dysfunction in Pancreatic Cancer. Gastroenterology 2023, 165, 891–908.e14. [Google Scholar] [CrossRef] [PubMed]
- Liudahl, S.M.; Betts, C.B.; Sivagnanam, S.; Morales-Oyarvide, V.; da Silva, A.; Yuan, C.; Hwang, S.; Grossblatt-Wait, A.; Leis, K.R.; Larson, W.; et al. Leukocyte Heterogeneity in Pancreatic Ductal Adenocarcinoma: Phenotypic and Spatial Features Associated with Clinical Outcome. Cancer Discov. 2021, 11, 2014–2031. [Google Scholar] [CrossRef] [PubMed]
- Vayrynen, S.A.; Zhang, J.; Yuan, C.; Vayrynen, J.P.; Costa, A.D.; Williams, H.; Morales-Oyarvide, V.; Lau, M.C.; Rubinson, D.A.; Dunne, R.F.; et al. Composition, Spatial Characteristics, and Prognostic Significance of Myeloid Cell Infiltration in Pancreatic Cancer. Clin. Cancer Res. 2021, 27, 1069–1081. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, Y.; Ma, J.; Yang, X.; Liu, D.; Zhu, Y.; Zhao, Y.; Fei, X.; Xu, D.; Dai, J. Identifying the Spatial Architecture That Restricts the Proximity of CD8+ T Cells to Tumor Cells in Pancreatic Ductal Adenocarcinoma. Cancers 2024, 16, 1434. https://doi.org/10.3390/cancers16071434
Xia Y, Ma J, Yang X, Liu D, Zhu Y, Zhao Y, Fei X, Xu D, Dai J. Identifying the Spatial Architecture That Restricts the Proximity of CD8+ T Cells to Tumor Cells in Pancreatic Ductal Adenocarcinoma. Cancers. 2024; 16(7):1434. https://doi.org/10.3390/cancers16071434
Chicago/Turabian StyleXia, Yihan, Junrui Ma, Xiaobao Yang, Danping Liu, Yujie Zhu, Yanan Zhao, Xuefeng Fei, Dakang Xu, and Jing Dai. 2024. "Identifying the Spatial Architecture That Restricts the Proximity of CD8+ T Cells to Tumor Cells in Pancreatic Ductal Adenocarcinoma" Cancers 16, no. 7: 1434. https://doi.org/10.3390/cancers16071434
APA StyleXia, Y., Ma, J., Yang, X., Liu, D., Zhu, Y., Zhao, Y., Fei, X., Xu, D., & Dai, J. (2024). Identifying the Spatial Architecture That Restricts the Proximity of CD8+ T Cells to Tumor Cells in Pancreatic Ductal Adenocarcinoma. Cancers, 16(7), 1434. https://doi.org/10.3390/cancers16071434