

Production of Trans-Cinnamic and p-Coumaric Acids in Engineered E. coli


Abstract
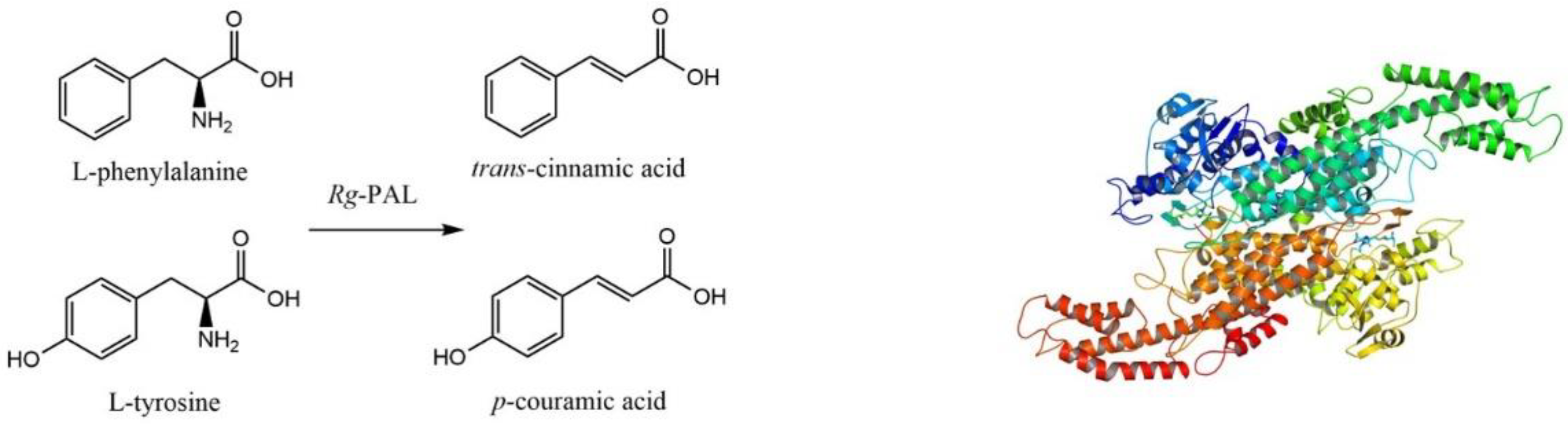
:1. Introduction
2. Results
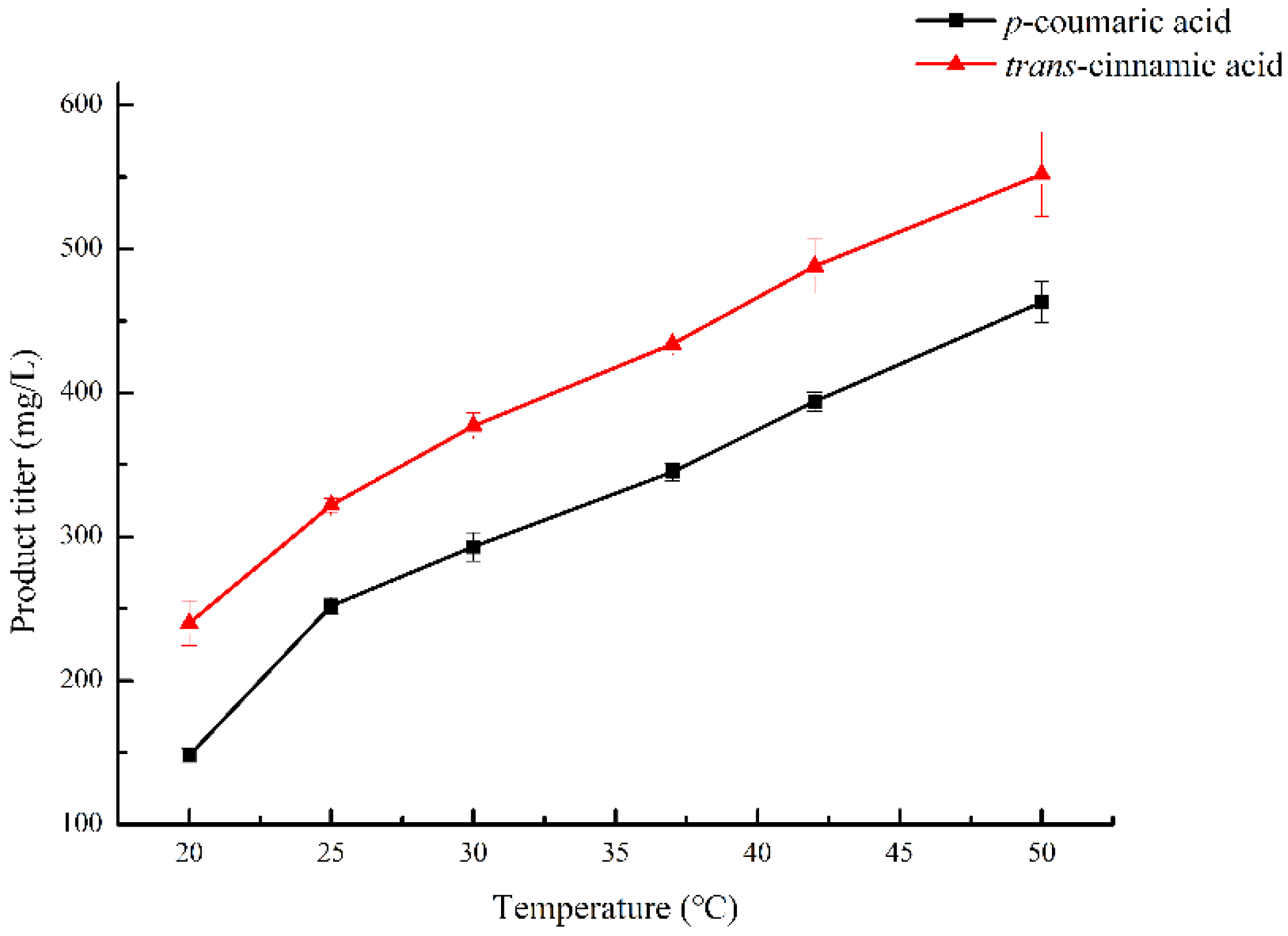
2.1. Optimization of Temperature
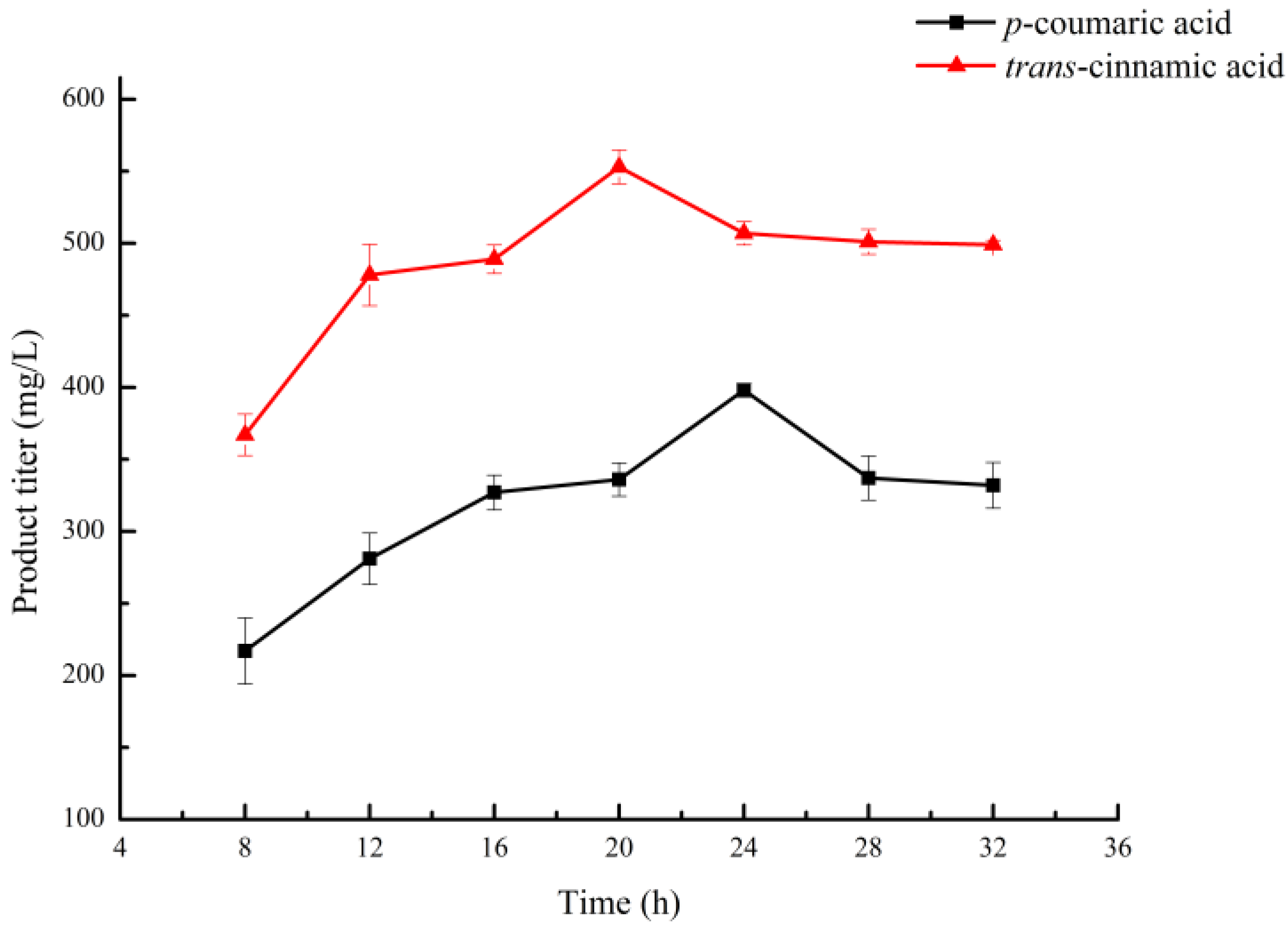
2.2. Optimization of Time
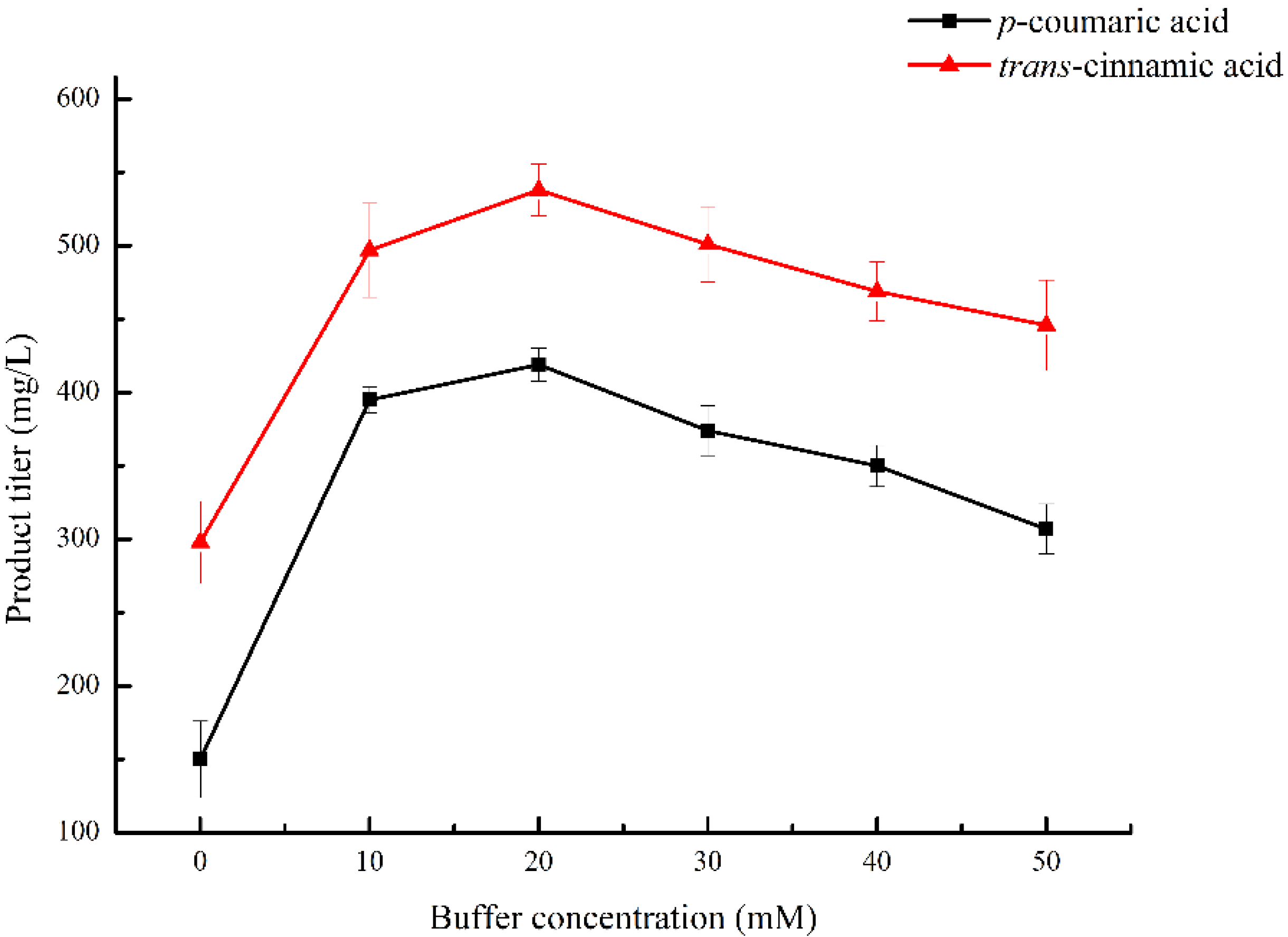
2.3. Optimization of Buffer Concentration
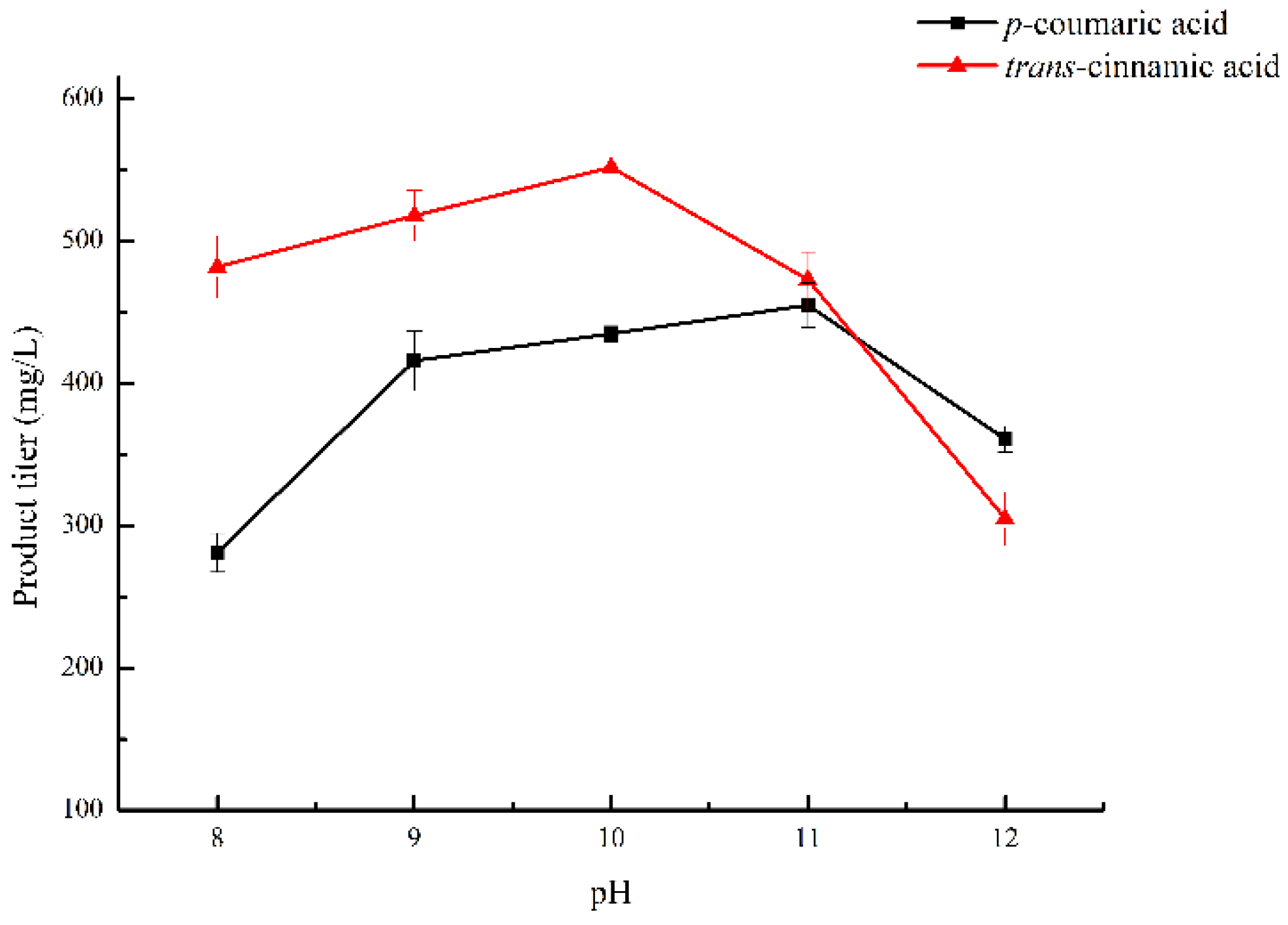
2.4. Optimization of pH
2.5. Optimization of Cell Amount
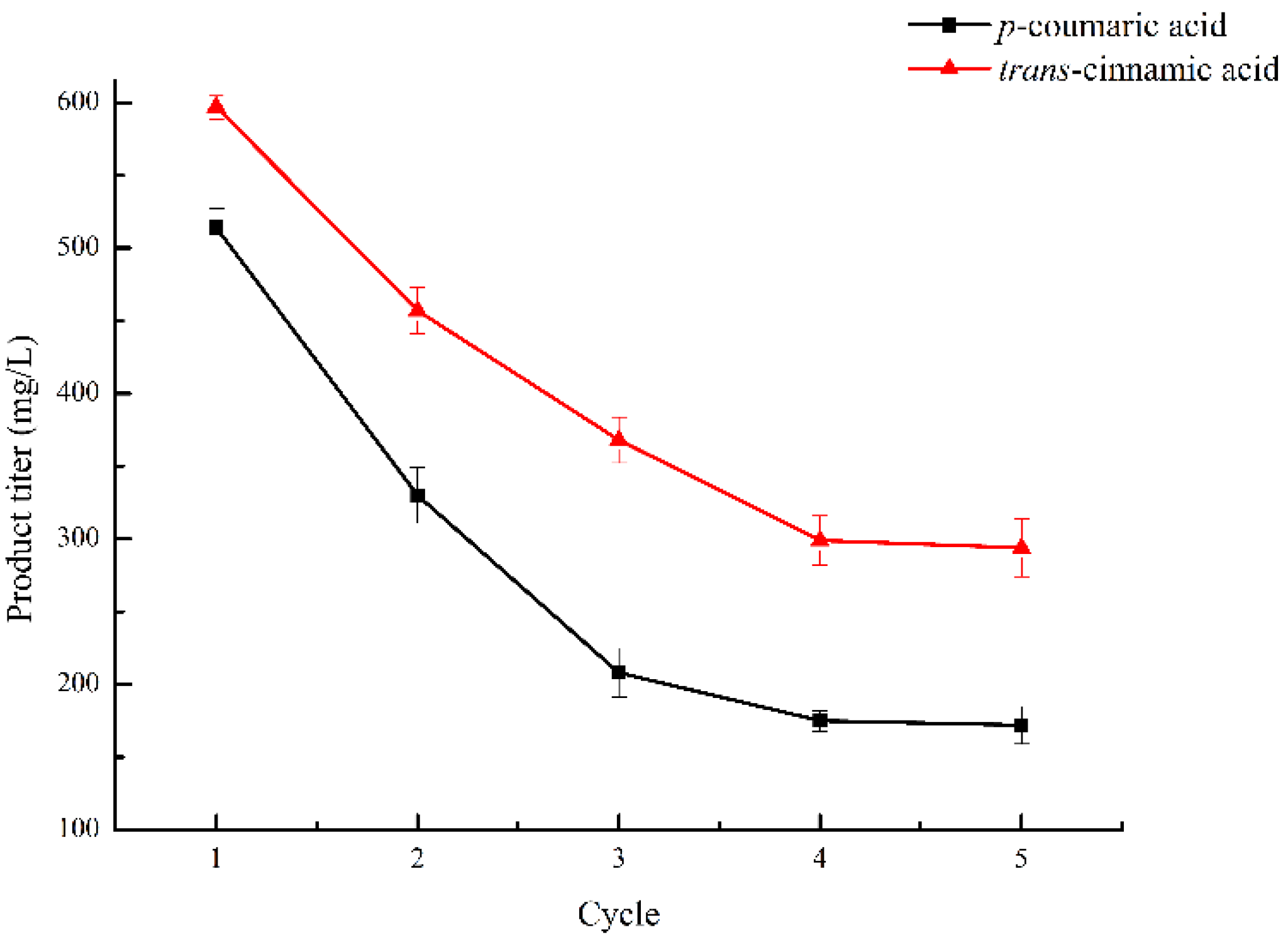
2.6. Reusability
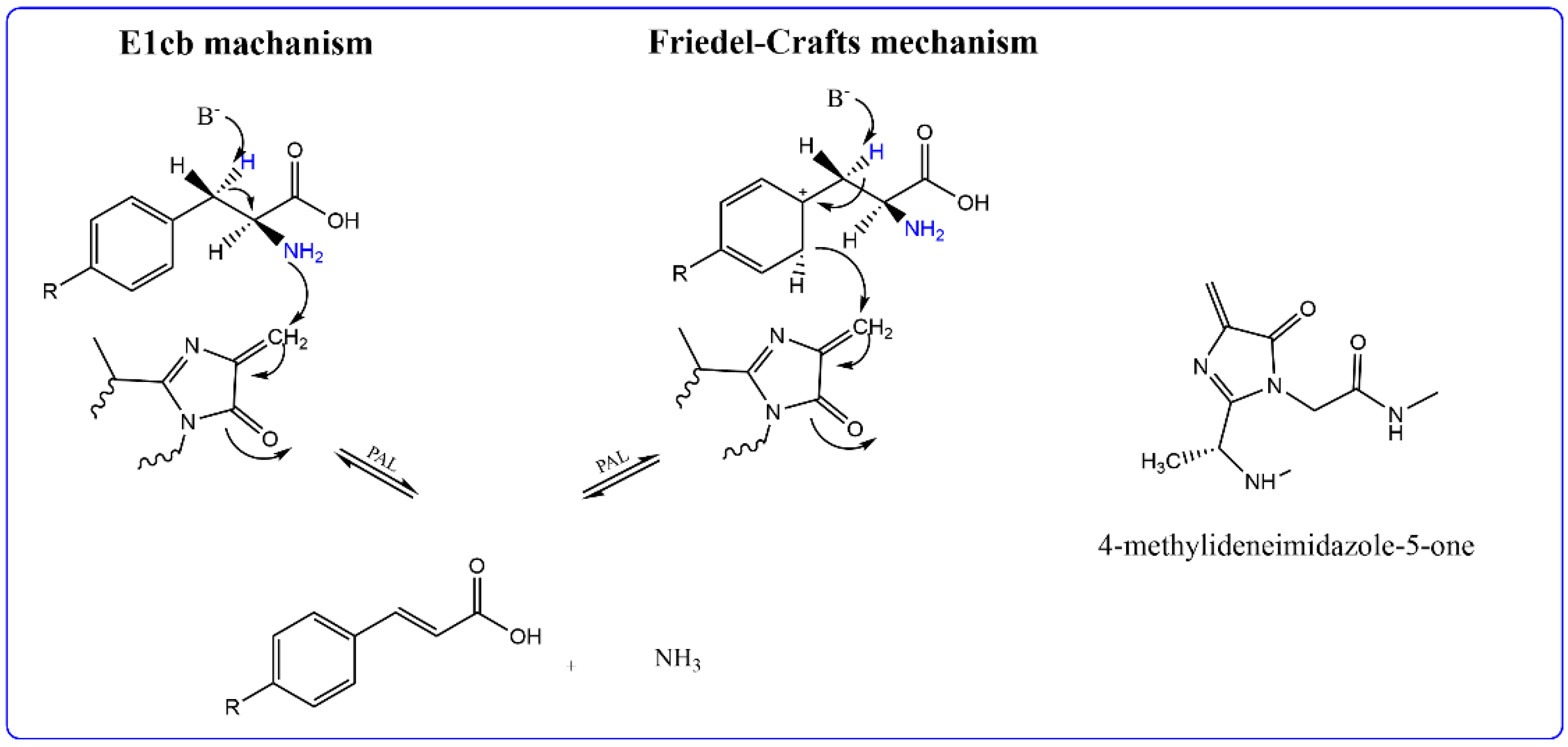
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
4.2. Media and Cultivations
4.3. Whole-Cell Biotransformation of L-Phe to Trans-Cinnamic Acid and L-Tyr to p-Coumaric Acid by E. coli BL21(DE3)/pETDuet-1-RgPAL
4.4. Analytical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hyun, M.W.; Yun, Y.H.; Kim, J.Y.; Kim, S.H. Fungal and Plant Phenylalanine Ammonia-Lyase. Mycobiology 2011, 39, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Whetten, R.W.; Sederoff, R.R. Phenylalanine Ammonia-Lyase from Loblolly Pine: Purification of the Enzyme and Isolation of Complementary DNA Clones. Plant Physiol. 1992, 98, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Koukol, J.; Conn, E.E. The Metabolism of Aromatic Compounds in Higher Plants. J. Biol. Chem. 1961, 236, 2692–2698. [Google Scholar] [CrossRef]
- Wang, X.H.; Gong, M.; Tang, L.; Zheng, S.; Lou, J.D.; Ou, L.; Gomes-Laranjo, J.; Zhang, C. Cloning, Bioinformatics and the Enzyme Activity Analyses of a Phenylalanine Ammonia-Lyase Gene Involved in Dragon’s Blood Biosynthesis in Dracaena cambodiana. Mol. Biol. Rep. 2013, 40, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Shao, F.; Ma, Y.; Lu, S. The Phenylalanine Ammonia-Lyase Gene Family in Salvia miltiorrhiza: Genome-Wide Characterization, Molecular Cloning and Expression Analysis. Mol. Biol. Rep. 2013, 40, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Young, M.R.; Towers, G.H.N.; Neish, A.C. Taxonomic Distribution of Ammonia-Lyases for L-Phenylalanine and L-Tyrosine in Relation To Lignification. Can. J. Bot. 1966, 44, 341–349. [Google Scholar] [CrossRef]
- Czichi, U.; Kindl, H. Formation of P-Coumaric Acid and o-Coumaric Acid from l-Phenylalanine by Microsomal Membrane Fractions from Potato: Evidence of Membrane-Bound Enzyme Complexes. Planta 1975, 125, 115–125. [Google Scholar] [CrossRef]
- Sikora, L.A.; Marzluf, G.A. Regulation of L-Phenylalanine Ammonia-Lyase by L-Phenylalanine and Nitrogen in Neurospora crassa. J. Bacteriol. 1982, 150, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Marusich, W.C.; Jensen, R.A.; Zamir, L.O. Induction of L-Phenylalanine Ammonia-Lyase during Utilization of Phenylalanine as a Carbon or Nitrogen Source in Rhodotorula glutinis. J. Bacteriol. 1981, 146, 1013–1019. [Google Scholar] [CrossRef]
- Orndorff, S.A.; Costantino, N.; Stewart, D.; Durham, D.R. Strain Improvement of Rhodotorula graminis for Production of a Novel l-Phenylalanine Ammonia-Lyase. Appl. Environ. Microbiol. 1988, 54, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Vance, C.P.; Bandoni, R.; Towers, G.H.N. Further Observations on Phenylalanine in Fungi. Phytochemistry 1975, 14, 1513–1514. [Google Scholar] [CrossRef]
- Bandoni, R.J.; Moore, K.; Subba Rao, P.V.; Towers, G.H.N. Phenylalanine and Tyrosine Ammonia-Lyase Activity in Some Basidiomycetes. Phytochemistry 1968, 7, 205–207. [Google Scholar] [CrossRef]
- Xiang, L.; Moore, B.S. Inactivation, Complementation, and Heterologous Expression of EncP, a Novel Bacterial Phenylalanine Ammonia-Lyase Gene. J. Biol. Chem. 2002, 277, 32505–32509. [Google Scholar] [CrossRef]
- Kawatra, A.; Dhankhar, R.; Mohanty, A.; Gulati, P. Biomedical Applications of Microbial Phenylalanine Ammonia Lyase: Current Status and Future Prospects. Biochimie 2020, 177, 142–152. [Google Scholar] [CrossRef]
- Alunni, S.; Cipiciani, A.; Fioroni, G.; Ottavi, L. Mechanisms of Inhibition of Phenylalanine Ammonia-Lyase by Phenol Inhibitors and Phenol/Glycine Synergistic Inhibitors. Arch. Biochem. Biophys. 2003, 412, 170–175. [Google Scholar] [CrossRef]
- Givot, I.L.; Smith, T.A.; Abeles, R.H. Studies on the Mechanism of Action and the Structure of the Electrophilic Center of Histidine Ammonia Lyase. J. Biol. Chem. 1969, 244, 6341–6353. [Google Scholar] [CrossRef]
- Poppe, L.; Rétey, J. Friedel-Crafts-Type Mechanism for the Enzymatic Elimination of Ammonia from Histidine and Phenylalanine. Angew. Chem.-Int. Ed. 2005, 44, 3668–3688. [Google Scholar] [CrossRef]
- Darvesh, A.S.; Carroll, R.T.; Bishayee, A.; Geldenhuys, W.J.; Van Der Schyf, C.J. Oxidative Stress and Alzheimer’s Disease: Dietary Polyphenols as Potential Therapeutic Agents. Expert Rev. Neurother. 2010, 10, 729–745. [Google Scholar] [CrossRef]
- Khazaei, M.R.; Bakhti, M.; Habibi-Rezaei, M. Nicotine Reduces the Cytotoxic Effect of Glycated Proteins on Microglial Cells. Neurochem. Res. 2010, 35, 548–558. [Google Scholar] [CrossRef]
- Schmidt, A.M.; Stern, D. Atherosclerosis and Diabetes: The RAGE Connection. Curr. Atheroscler. Rep. 2000, 2, 430–436. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Sompong, W.; Meeprom, A.; Ngamukote, S.; Yibchok-Anun, S. Cinnamic Acid and Its Derivatives Inhibit Fructose-Mediated Protein Glycation. Int. J. Mol. Sci. 2012, 13, 1778–1789. [Google Scholar] [CrossRef] [PubMed]
- Koopman, F.; Beekwilder, J.; Crimi, B.; van Houwelingen, A.; Hall, R.D.; Bosch, D.; van Maris, A.J.A.; Pronk, J.T.; Daran, J.M. De Novo Production of the Flavonoid Naringenin in Engineered Saccharomyces cerevisiae. Microb. Cell Fact. 2012, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, M.H.; El-Mahdy, M.A.; Abd-Ellah, M.F.; Helal, G.K.; Khalifa, F.; Hamada, F.M.A. Influence of P-Coumaric Acid on Doxorubicin-Induced Oxidative Stress in Rat’s Heart. Pharmacol. Res. 2003, 48, 461–465. [Google Scholar] [CrossRef]
- Liang, J.L.; Guo, L.; Sun, P.; Jiang, B.; Lin, J.; Guo, W.; Wan, H. A Novel Process for Obtaining Phenylpropanoic Acid Precursor Using Escherichia coli with a Constitutive Expression System. Food Sci. Biotechnol. 2016, 25, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Jiang, T.; Cong, Y.; Zheng, Z.; Ouyang, J. Molecular Characterization of a Recombinant Zea mays Phenylalanine Ammonia-Lyase (ZmPAL2) and Its Application in Trans-Cinnamic Acid Production from l-Phenylalanine. Appl. Biochem. Biotechnol. 2015, 176, 924–937. [Google Scholar] [CrossRef]
- Xue, Z.; McCluskey, M.; Cantera, K.; Ben-Bassat, A.; Sariaslani, F.S.; Huang, L. Improved Production of P-Hydroxycinnamic Acid from Tyrosine Using a Novel Thermostable Phenylalanine/Tyrosine Ammonia Lyase Enzyme. Enzyme Microb. Technol. 2007, 42, 58–64. [Google Scholar] [CrossRef]
- Abdullah, A.Z.; Sulaiman, N.S.; Kamaruddin, A.H. Biocatalytic Esterification of Citronellol with Lauric Acid by Immobilized Lipase on Aminopropyl-Grafted Mesoporous SBA-15. Biochem. Eng. J. 2009, 44, 263–270. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names | Characteristics | Source |
|---|---|---|
| Plasmid | ||
| pETDuet-1 | pBR322ori with PT7; AmpR | Novagen |
| pETDuet-1-Rg-PAL | pETDuet-1 with Rg-PAL | This study |
| Strain | ||
| E. coli DH5α | ΔLac U169 (Φ80 Lac Z ΔM15) | Invitrogen |
| E. coli BL21(DE3) | F-ompT hsdS (rB-mB-) gal dcm (DE3) | Invitrogen |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Xu, W.; Xu, W. Production of Trans-Cinnamic and p-Coumaric Acids in Engineered E. coli. Catalysts 2022, 12, 1144. https://doi.org/10.3390/catal12101144
Liu Y, Xu W, Xu W. Production of Trans-Cinnamic and p-Coumaric Acids in Engineered E. coli. Catalysts. 2022; 12(10):1144. https://doi.org/10.3390/catal12101144
Chicago/Turabian StyleLiu, Yuqi, Weizhuo Xu, and Wei Xu. 2022. "Production of Trans-Cinnamic and p-Coumaric Acids in Engineered E. coli" Catalysts 12, no. 10: 1144. https://doi.org/10.3390/catal12101144
APA StyleLiu, Y., Xu, W., & Xu, W. (2022). Production of Trans-Cinnamic and p-Coumaric Acids in Engineered E. coli. Catalysts, 12(10), 1144. https://doi.org/10.3390/catal12101144