

Enhancing Acetophenone Tolerance of Anti-Prelog Short-Chain Dehydrogenase/Reductase EbSDR8 Using a Whole-Cell Catalyst by Directed Evolution

Abstract
:1. Introduction
2. Results and Discussion
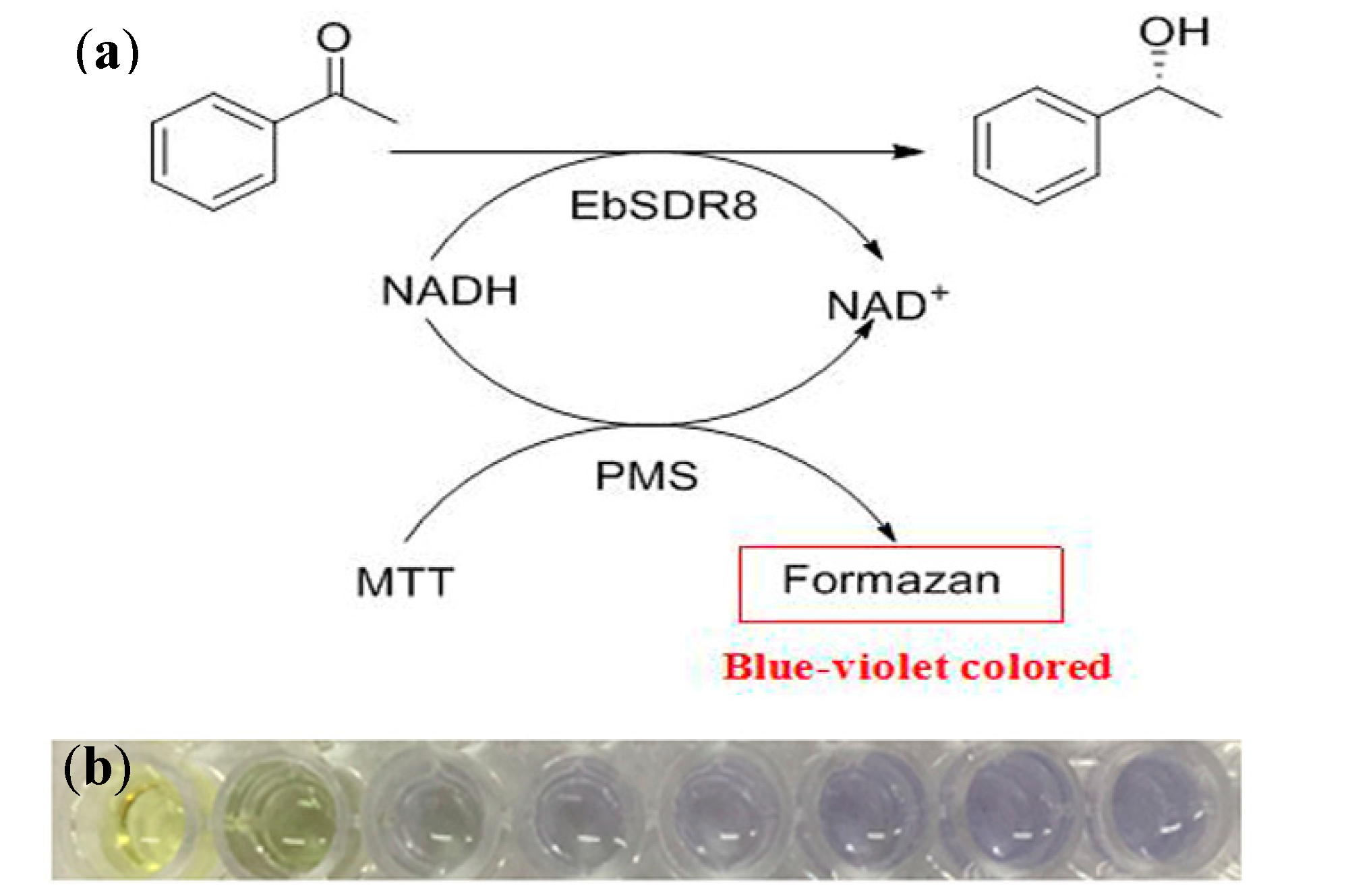
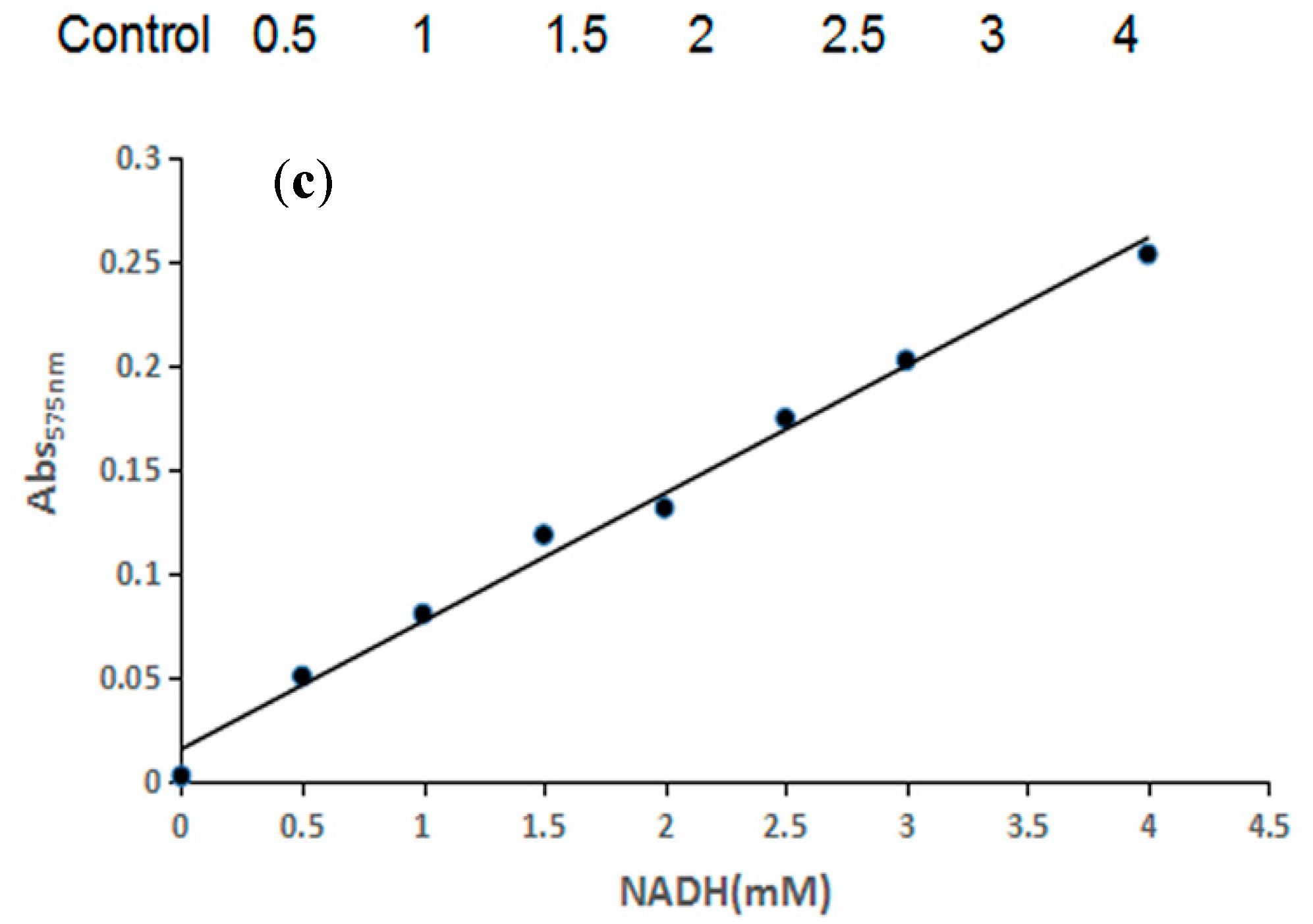
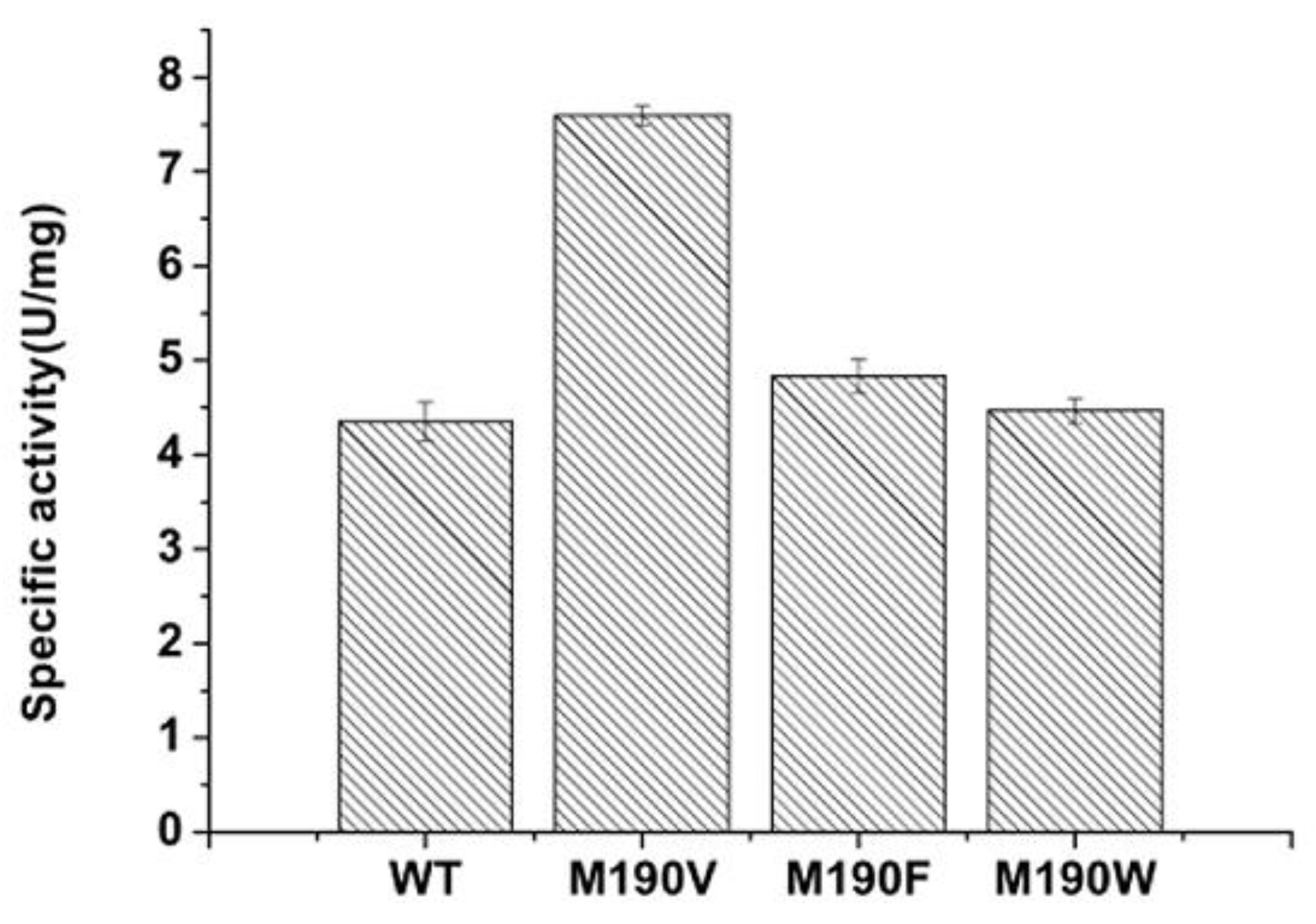
2.1. High-Throughput Screening Method and Activity Assay of the Mutants
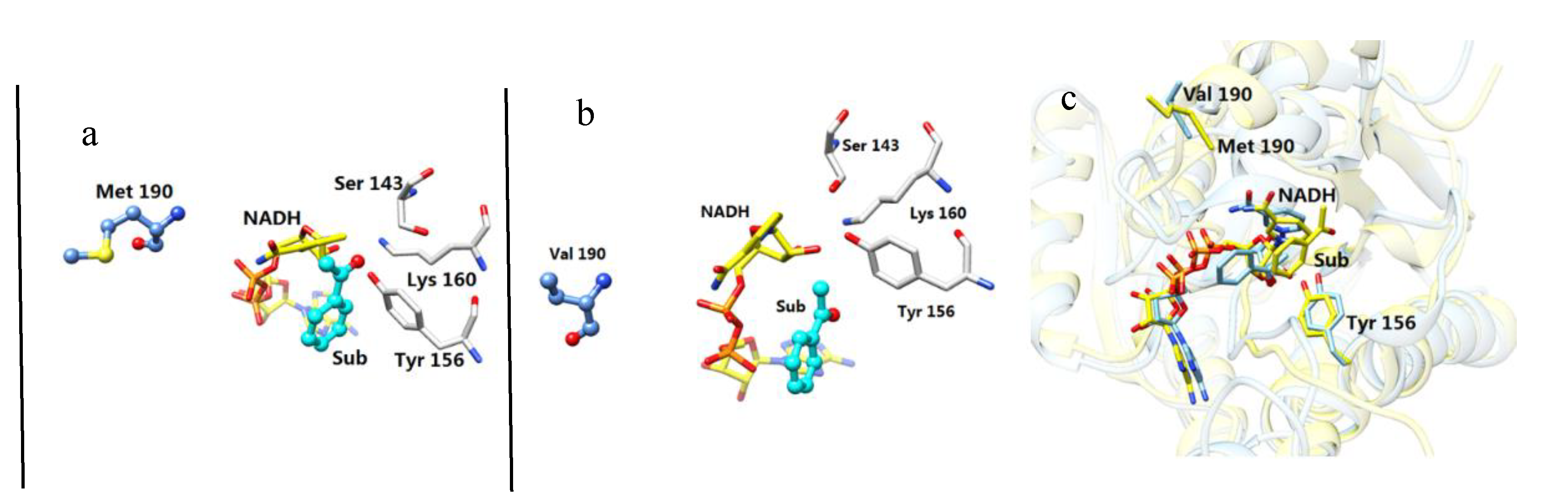
2.2. Comparative Structure Analysis of the Mutant M190V and the Wild-Type
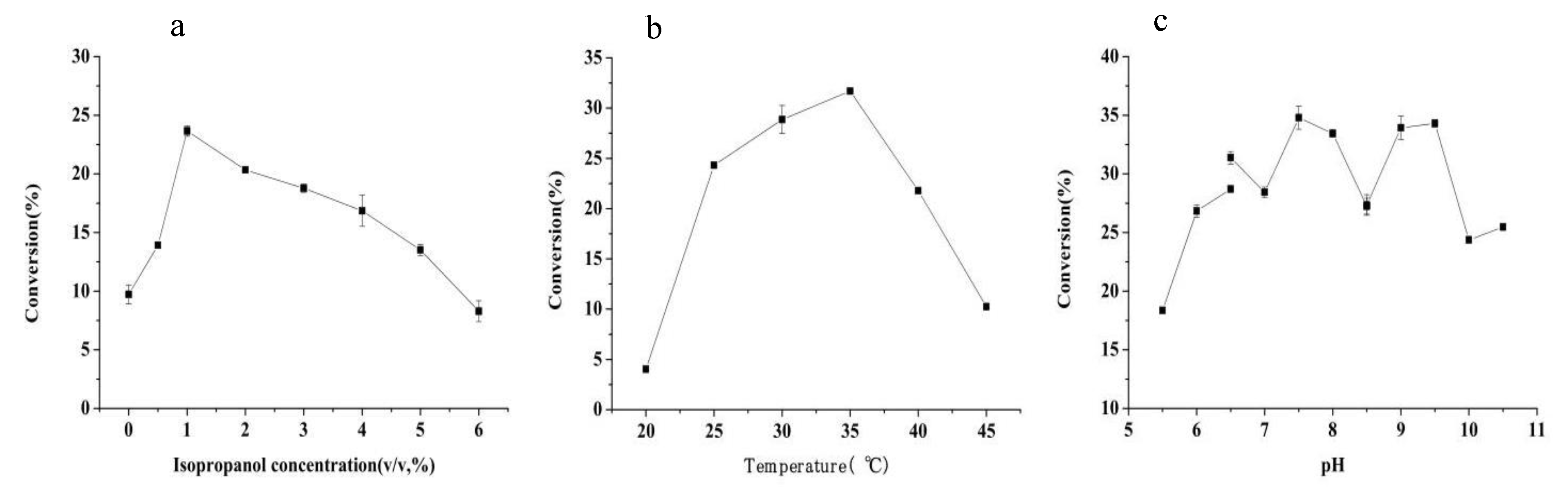
2.3. Process Optimization of the M190V Whole-Cell-Catalyzed Asymmetric Reduction of Acetophenone
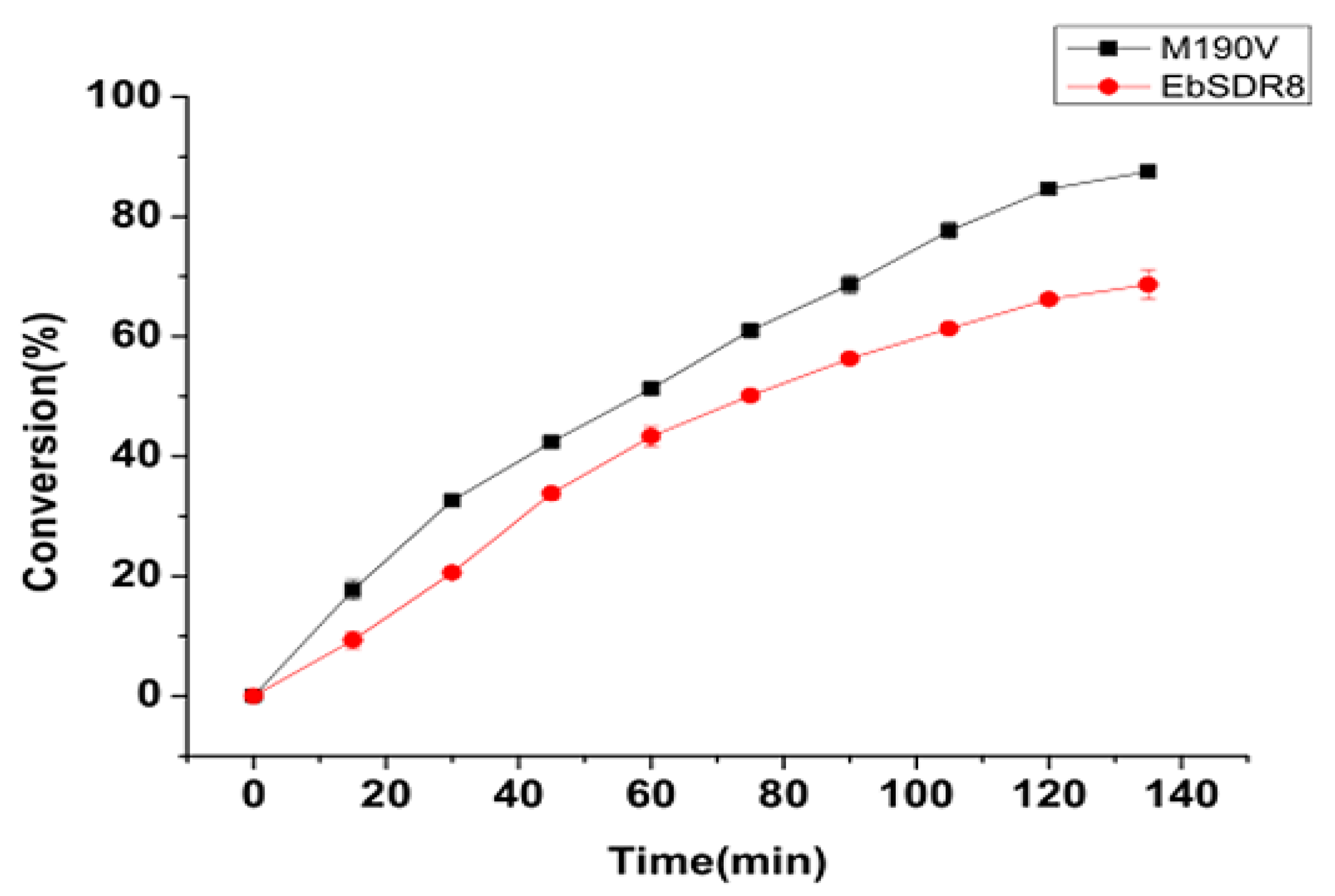
2.4. Time Course Study on the M190V Whole-Cell-Catalyzed Asymmetric Reduction of Acetophenone
3. Materials and Methods
3.1. Materials
3.2. Error-Prone PCR and Construction of Mutant Library
3.3. High-Throughput Screening
3.4. Enzyme Activity Assay and Kinetic Characterization
3.5. Characterization of EbSDR8 Mutant
3.6. Analytical Methods
3.7. Molecular Simulation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Approval and Consent to Participate
Abbreviations
References
- Li, A.P.; Ye, L.D.; Wu, H.P.; Yang, X.H.; Yu, H.W. Characterization of an excellent anti-Prelog short-chain dehydrogenase/reductase EbSDR8 from Empedobacter brevis ZJUY-1401. J. Mol. Catal. B Enzym. 2015, 122, 179–187. [Google Scholar] [CrossRef]
- Morris, R.E.; Bu, X. Induction of chiral porous solids containing only achiral building blocks. Nat. Chem. 2010, 2, 353–361. [Google Scholar] [CrossRef]
- Musa, M.M. Alcohol Dehydrogenases with anti-Prelog Stereopreference in Synthesis of Enantiopure Alcohols. ChemistryOpen 2022, 11, 4. [Google Scholar] [CrossRef]
- Han, M.N.; Wang, X.M.; Pei, C.H.; Zhang, C.; Xu, Z.; Zhang, H.L.; Li, W. Green and scalable synthesis of chiral aromatic alcohols through an efficient biocatalytic system. Microb. Biotechnol. 2021, 14, 444–452. [Google Scholar] [CrossRef]
- Goldberg, K.; Schroer, K.; Lutz, S.; Liese, A. Biocatalytic ketone reduction—A powerful tool for the production of chiral alcohols—Part I: Processes with isolated enzymes. Appl. Microbiol. Biotechnol. 2007, 76, 237–248. [Google Scholar] [CrossRef]
- Huisman, G.W.; Liang, J.; Krebber, A. Practical chiral alcohol manufacture using ketoreductases. Curr. Opin. Chem. Biol. 2010, 14, 122–129. [Google Scholar] [CrossRef]
- Garzón-Posse, F.; Becerra-Figueroa, L.; Hernández-Arias, J.; Gamba-Sánchez, D. Whole Cells as Biocatalysts in Organic Transformations. Molecules 2018, 23, 1265. [Google Scholar] [CrossRef]
- Asako, H.; Shimizu, M.; Makino, Y.; Itoh, N. Biocatalytic reduction system for the production of chiral methyl (R)/(S)-4-bromo-3-hydroxybutyrate. Tetrahedron Lett. 2010, 51, 2664–2666. [Google Scholar] [CrossRef]
- Hohne, M.; Schatzle, S.; Jochens, H.; Robins, K.; Bornscheuer, U.T. Rational assignment of key motifs for function guides in silico enzyme identification. Nat. Chem. Biol. 2010, 6, 807–813. [Google Scholar] [CrossRef]
- Zheng, G.W.; Xu, J.H. New opportunities for biocatalysis: Driving the synthesis of chiral chemicals. Curr. Opin. Biotechnol. 2011, 22, 784–792. [Google Scholar] [CrossRef]
- Alsafadi, D.; Paradisi, F. Effect of organic solvents on the activity and stability of halophilic alcohol dehydrogenase (ADH2) from Haloferax volcanii. Extremophiles 2013, 17, 115–122. [Google Scholar] [CrossRef]
- Kallberg, Y.; Oppermann, U.; Persson, B. Classification of the short-chain dehydrogenase/reductase superfamily using hidden Markov models. Febs J. 2010, 277, 2375–2386. [Google Scholar] [CrossRef]
- Li, Z.; Liu, W.; Chen, X.; Jia, S.; Wu, Q.; Zhu, D.; Ma, Y. Highly enantioselective double reduction of phenylglyoxal to (R)-1-phenyl-1,2-ethanediol by one NADPH-dependent yeast carbonyl reductase with a broad substrate profile. Tetrahedron 2013, 69, 3561–3564. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, S.C.; Lai, D.Y.; Zhang, S.L.; Chen, Z.M. Biocatalytic characterization of a short-chain alcohol dehydrogenase with broad substrate specificity from thermophilic Carboxydothermus hydrogenoformans. Biotechnol. Lett. 2013, 35, 359–365. [Google Scholar] [CrossRef]
- Shao, Z.H.; Su, B.M.; Yang, S.L.; Ye, L.D.; Yu, H.W. Rational design of the carbonyl reductase EbSDR8 for efficient biosynthesis of enantiopure (R)-3-chloro-1-phenyl-1-propanol. Appl. Microbiol. Biotechnol. 2020, 104, 9219–9228. [Google Scholar] [CrossRef]
- Nezhad, N.G.; Rahman, R.N.Z.R.A.; Normi, Y.M.; Oslan, S.N.; Shariff, F.M.; Leow, T.C. Thermostability engineering of industrial enzymes through structure modification. Appl. Microbiol. Biotechnol. 2022, 106, 4845–4866. [Google Scholar] [CrossRef]
- Basak, S.; Song, H.; Jiang, R.R. Error-prone PCR of global transcription factor cyclic AMP receptor protein for enhanced organic solvent (toluene) tolerance. Process Biochem. 2012, 47, 2152–2158. [Google Scholar] [CrossRef]
- Chong, H.Q.; Huang, L.; Yeow, J.W.; Wang, I.; Zhang, H.F.; Song, H.; Jiang, R.R. Improving ethanol tolerance of Escherichia coli by rewiring its global regulator cAMP receptor protein (CRP). PLoS ONE 2013, 8, e57628. [Google Scholar] [CrossRef]
- Chong, H.Q.; Yeow, J.W.; Wang, I.; Song, H.; Jiang, R.R. Improving acetate tolerance of Escherichia coli by rewiring its global regulator cAMP receptor protein (CRP). PLoS ONE 2013, 8, e77422. [Google Scholar]
- Yang, Y.; Liu, J.; Li, Z. Engineering of P450pyr Hydroxylase for the Highly Regio- and Enantioselective Subterminal Hydroxylation of Alkanes. Angew. Chem. Int. Ed. Engl. 2014, 53, 3120–3124. [Google Scholar] [CrossRef]
- Katz, M.; Frejd, T.; Hahn-Hagerdal, B.; Gorwa-Grauslund, M.F. Efficient anaerobic whole cell stereoselective bioreduction with recombinant Saccharomyces cerevisiae. Biotechnol. Bioeng. 2003, 84, 573–582. [Google Scholar] [CrossRef]
- Borg, A.J.E.; Beerens, K.; Pfeiffer, M.; Desmet, T.; Nidetzky, B. Stereo-electronic control of reaction selectivity in short-chain dehydrogenases: Decarboxylation, epimerization, and dehydration. Curr. Opin. Chem. Biol. 2021, 61, 43–52. [Google Scholar] [CrossRef]
- Bornscheuer, U.T.; Hauer, B.; Jaeger, K.E.; Schwaneberg, U. Directed Evolution Empowered Redesign of Natural Proteins for the Sustainable Production of Chemicals and Pharmaceuticals. Angew. Chem. Int. Ed. Engl. 2019, 58, 36–40. [Google Scholar] [CrossRef]
- Zheng, R.C.; Ge, Z.; Qiu, Z.K.; Wang, Y.S.; Zheng, Y.G. Asymmetric synthesis of (R)-1,3-butanediol from 4-hydroxy-2-butanone by a newly isolated strain Candida krusei ZJB-09162. Microbiol. Biotechnol. 2012, 94, 969–976. [Google Scholar] [CrossRef]
- Peters, J.; Minuth, T.; Kula, M.R. A novel NADH-dependent carbonyl reductase with an extremely broad substrate range from Candida parapsilosis: Purification and characterization. Enzyme Microb. Technol. 1993, 15, 950–958. [Google Scholar] [CrossRef]
- Rao, P.V.; Krishna, C.M.; Zigler, J.S. Identification and characterization of the enzymatic activity of zeta-crystallin from guinea pig lens. A novel NADPH: Quinone oxidoreductase. J. Biol. Chem. 1992, 267, 96–102. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Schneider, R.W. Effects of nonpathogenic strains of Fusarium oxysporum on celery root infection by Fusarium oxysporum f. sp. apii and a novel use of the Lineweaver-Burk double reciprocal plot technique. Phytopathology 1984, 74, 646–653. [Google Scholar] [CrossRef]
- Li, A.P.; Ye, L.D.; Yang, X.H.; Yang, C.C.; Gu, J.L.; Yu, H.W. Structure-guided stereoselectivity inversion of a short-chain dehydrogenase/reductase towards halogenated acetophenones. Chem. Commun. 2016, 52, 6284–6287. [Google Scholar] [CrossRef]
- Filling, C.; Berndt, K.D.; Benach, J.; Knapp, S.; Prozorovski, T.; Nordling, E.; Ladenstein, R.; Jornvall, H.; Oppermann, U. Critical residues for structure and catalysis in short-chain dehydrogenases/reductases. J. Biol. Chem. 2002, 277, 25677–25684. [Google Scholar] [CrossRef]
- Luo, X.; Wang, Y.J.; Shen, W.; Zheng, Y.G. Activity improvement of a Kluyveromyces lactis aldo-keto reductase KlAKR via rational design. J. Biotechnol. 2016, 224, 20–26. [Google Scholar] [CrossRef]
- Chen, W.W.; Ye, L.D.; Guo, F.; Lv, Y.; Yu, H.W. Enhanced activity of an alkaline phytase from Bacillus subtilis 168 in acidic and neutral environments by directed evolution. Biochem. Eng. J. 2015, 98, 137–143. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutants | Km (mM) | Kcat (s−1) | Kcat/Km (S−1M−1) |
|---|---|---|---|
| Wild-type | 4.95 | 2.53 | 511 |
| M190V | 4.57 | 3.98 | 870 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, B.; Yang, S.; Yu, H.; Ye, L. Enhancing Acetophenone Tolerance of Anti-Prelog Short-Chain Dehydrogenase/Reductase EbSDR8 Using a Whole-Cell Catalyst by Directed Evolution. Catalysts 2022, 12, 1071. https://doi.org/10.3390/catal12091071
Zhang H, Wang B, Yang S, Yu H, Ye L. Enhancing Acetophenone Tolerance of Anti-Prelog Short-Chain Dehydrogenase/Reductase EbSDR8 Using a Whole-Cell Catalyst by Directed Evolution. Catalysts. 2022; 12(9):1071. https://doi.org/10.3390/catal12091071
Chicago/Turabian StyleZhang, Hui, Bei Wang, Shengli Yang, Hongwei Yu, and Lidan Ye. 2022. "Enhancing Acetophenone Tolerance of Anti-Prelog Short-Chain Dehydrogenase/Reductase EbSDR8 Using a Whole-Cell Catalyst by Directed Evolution" Catalysts 12, no. 9: 1071. https://doi.org/10.3390/catal12091071
APA StyleZhang, H., Wang, B., Yang, S., Yu, H., & Ye, L. (2022). Enhancing Acetophenone Tolerance of Anti-Prelog Short-Chain Dehydrogenase/Reductase EbSDR8 Using a Whole-Cell Catalyst by Directed Evolution. Catalysts, 12(9), 1071. https://doi.org/10.3390/catal12091071