

Identification of New Amylolytic Enzymes from Marine Symbiotic Bacteria of Bacillus Species


Abstract
:1. Introduction
2. Results and Discussion
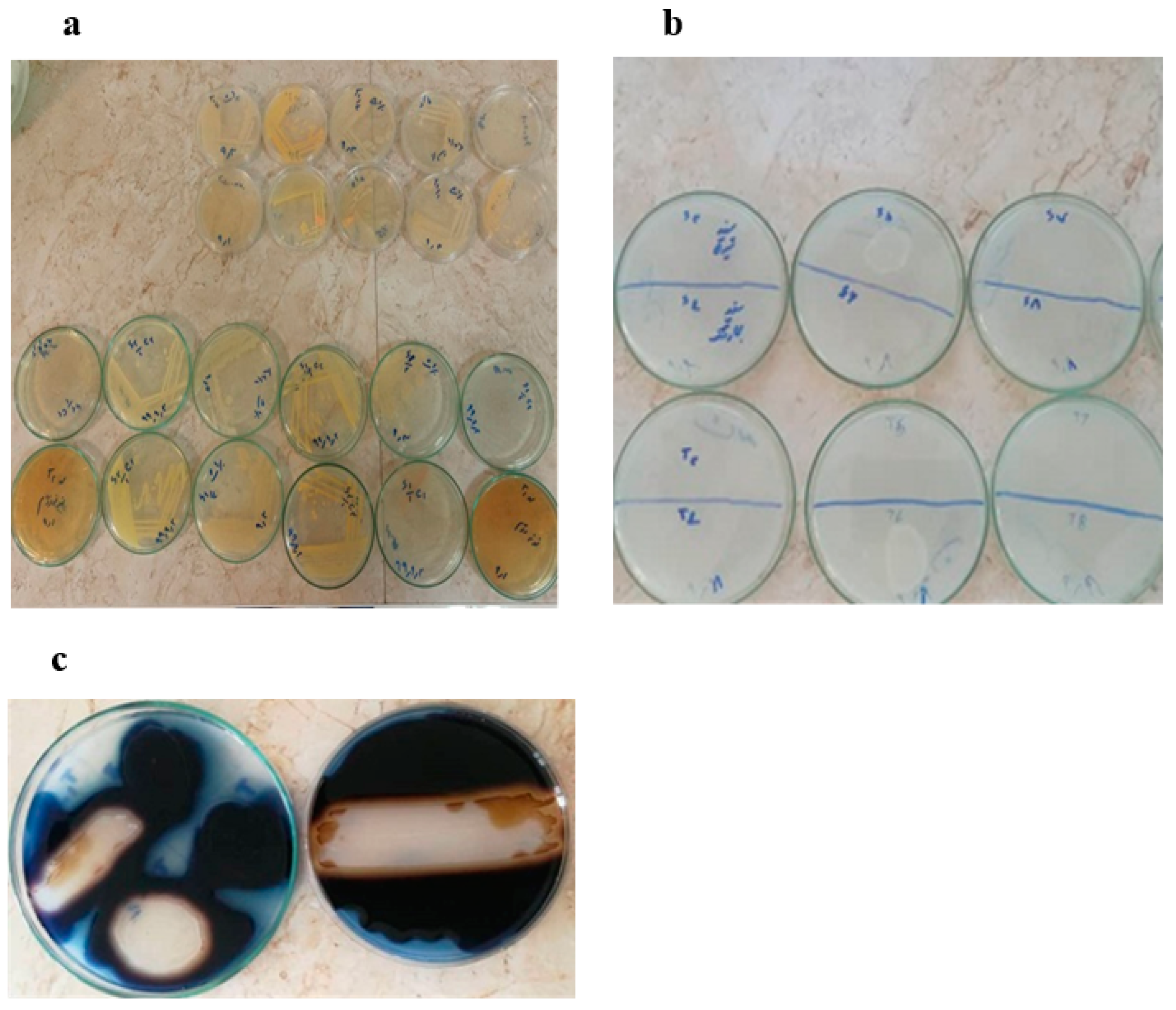
2.1. Primary and Secondary Screening of α-Amylase-Producing Bacteria
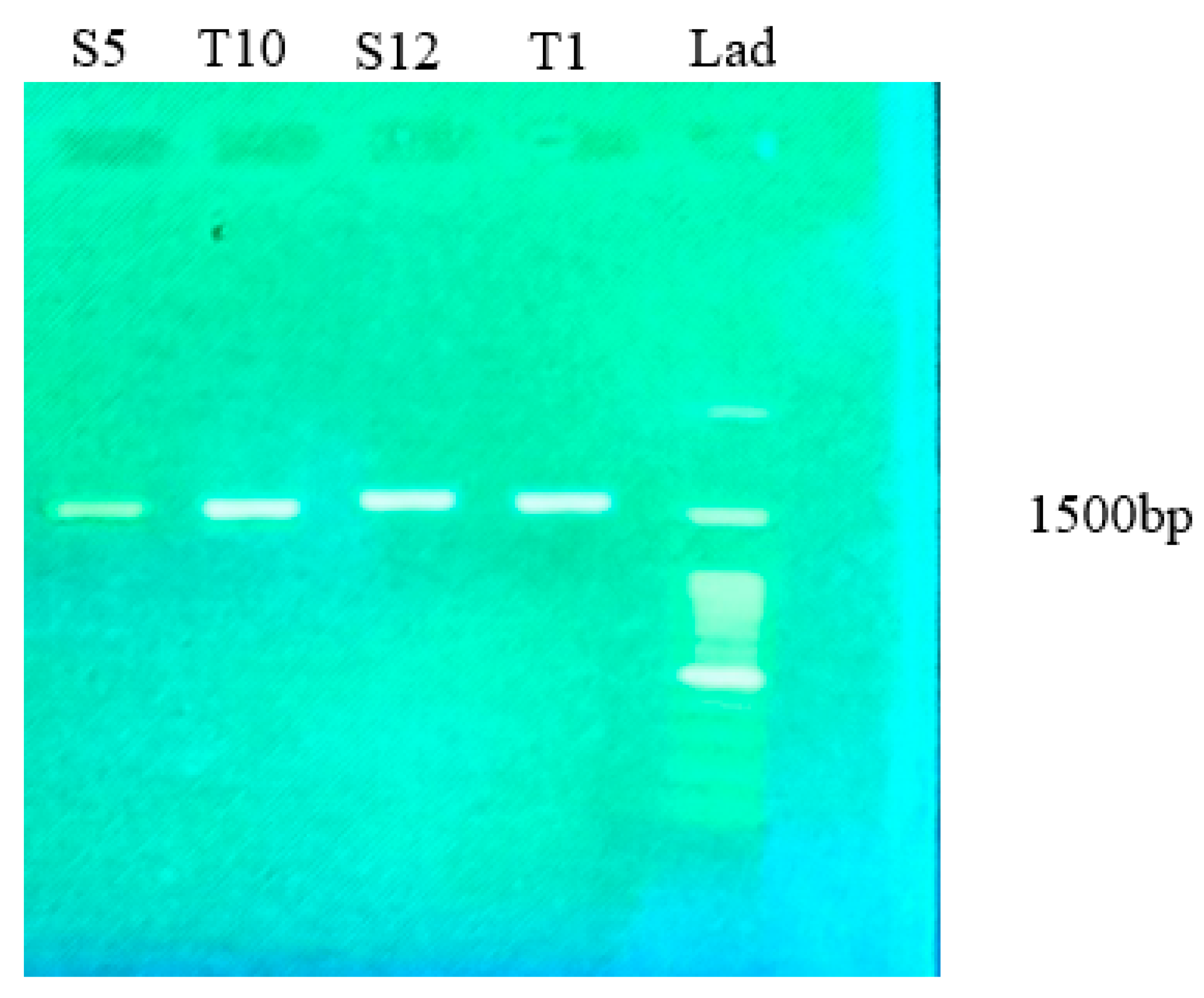
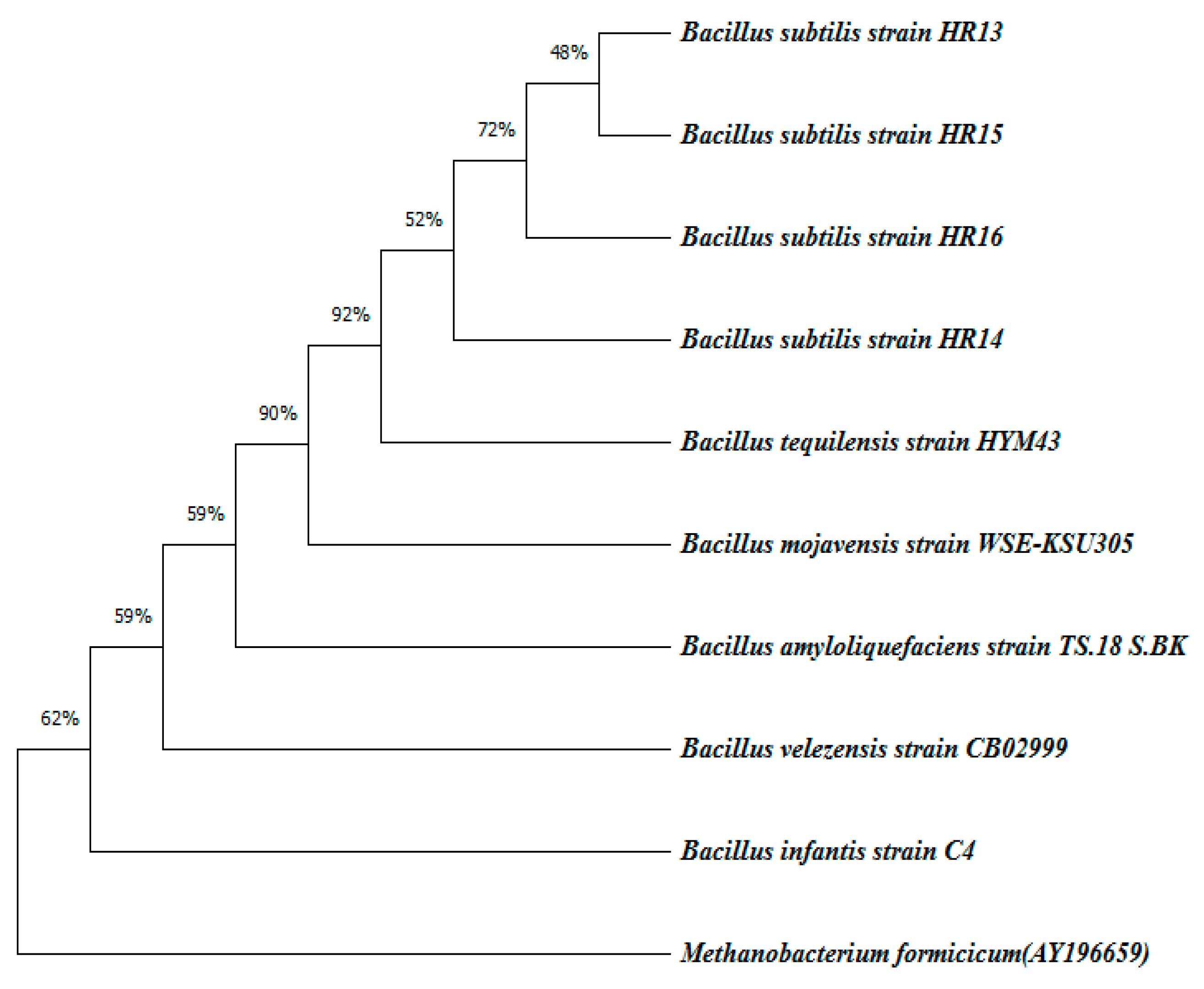
2.2. Molecular Identification of Potential α-Amylase-Producing Bacterial Strains
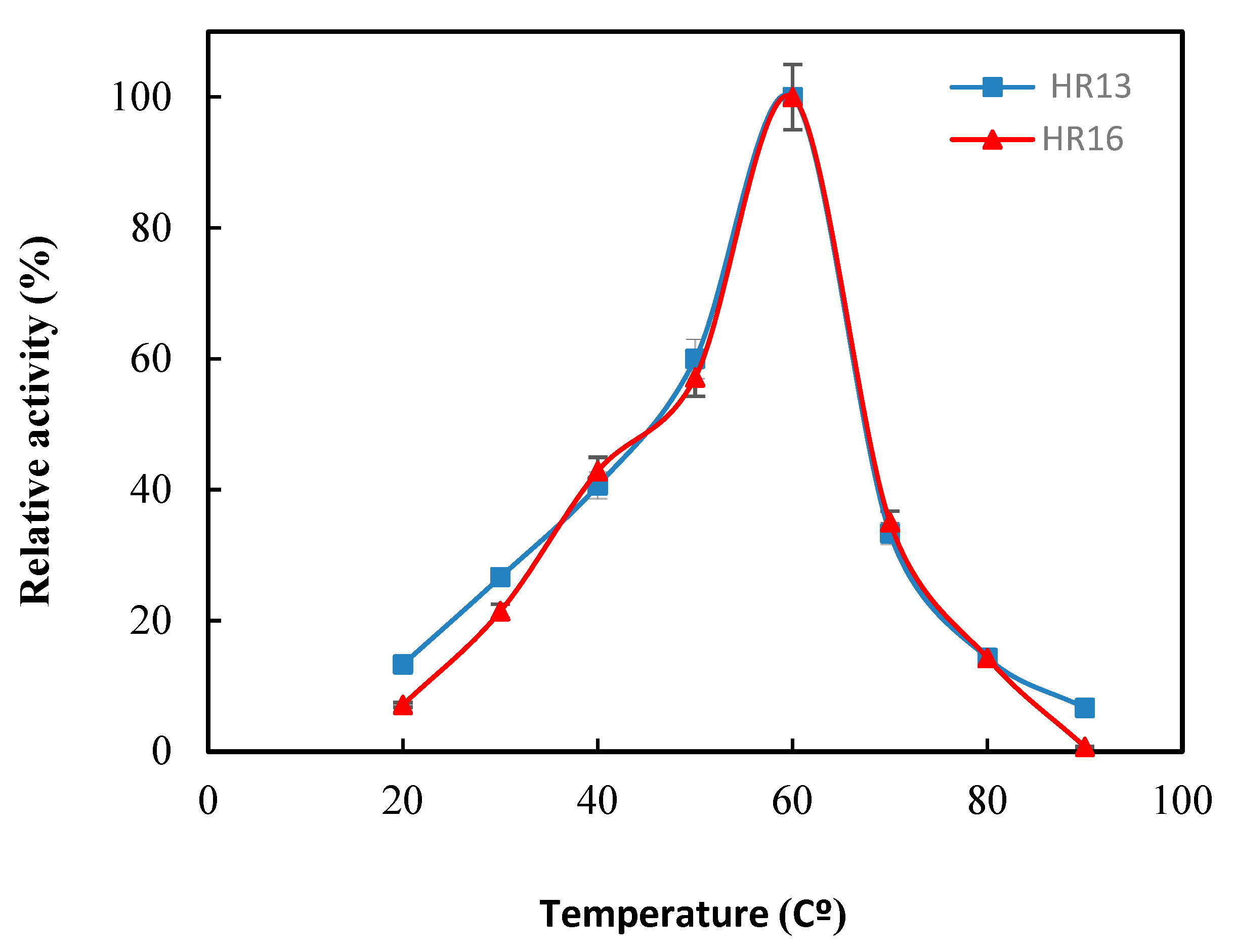
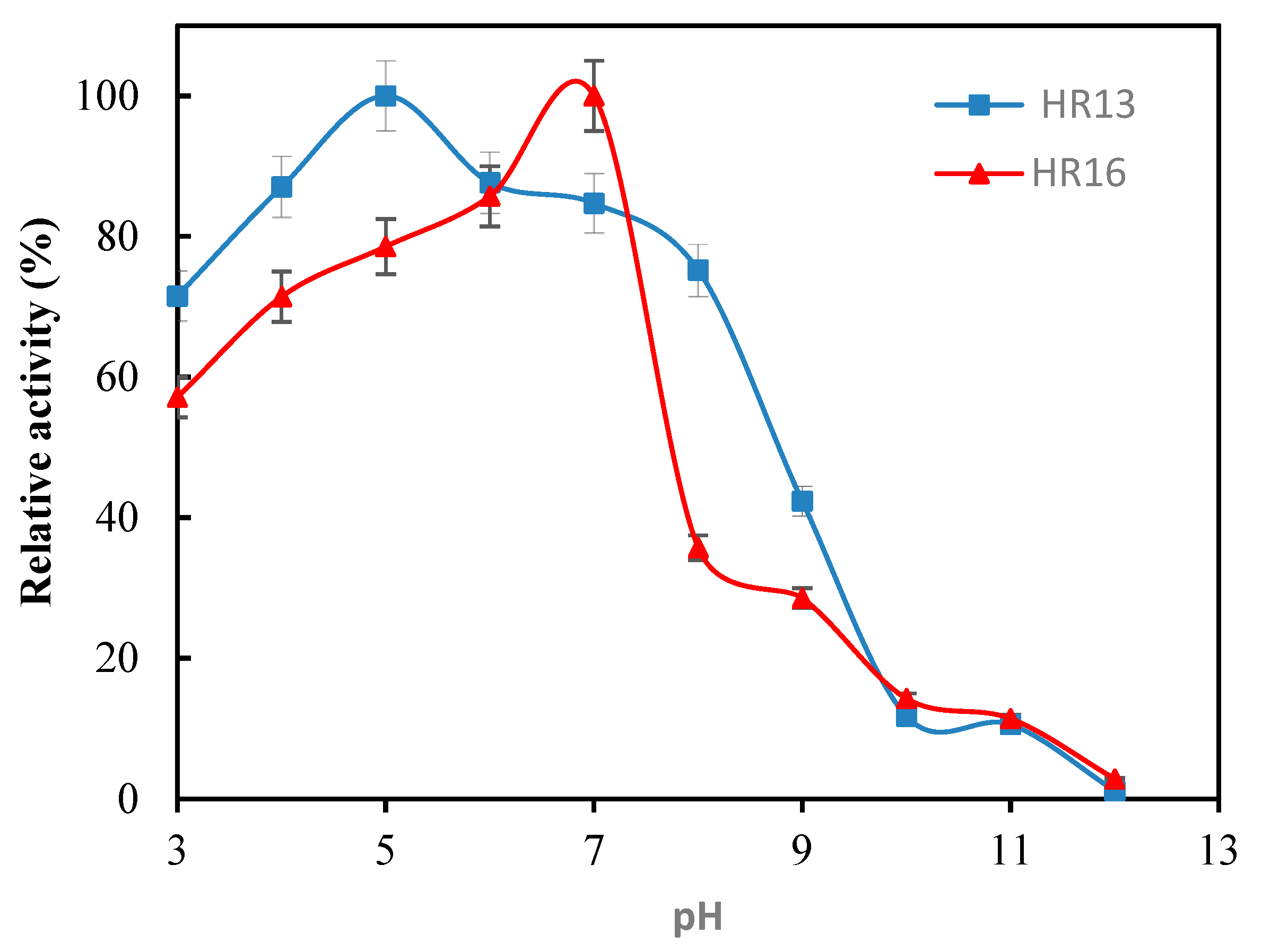
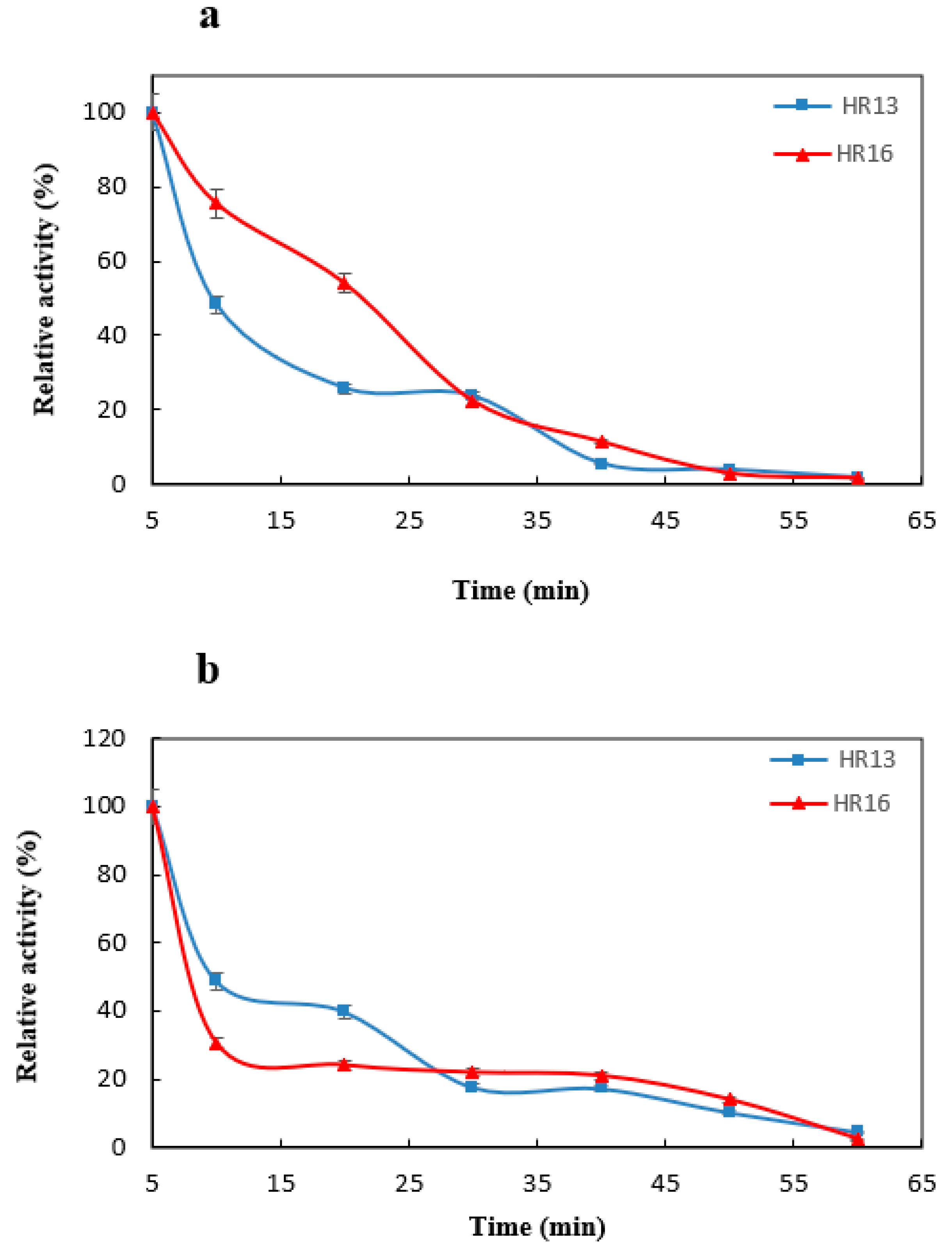
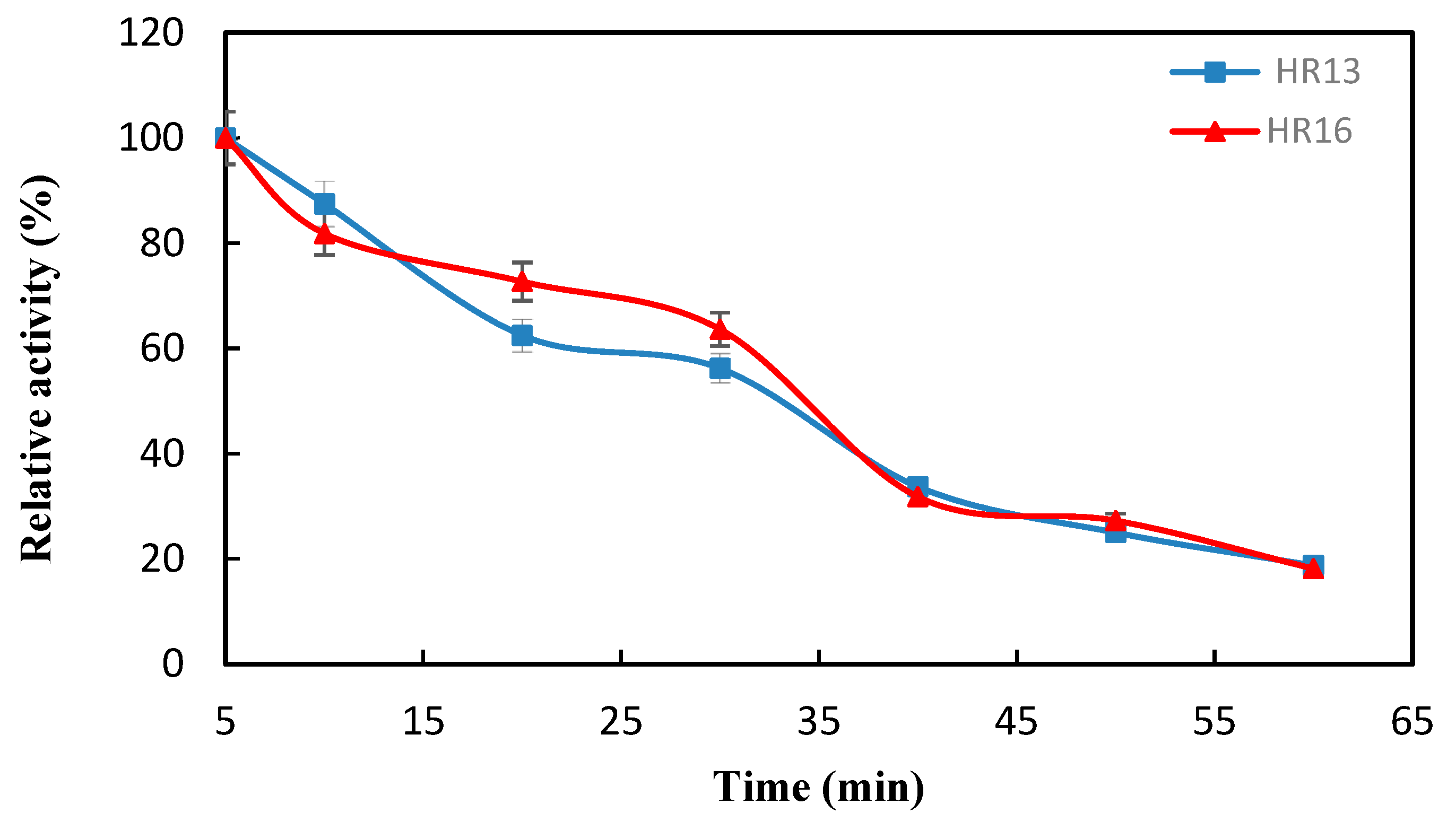
2.3. The Effect of Temperature and pH on the Activity and Stability of the α-Amylase Enzyme
3. Materials and Methods
3.1. Materials Used for the Collection of Short Fish Samples
3.2. Collection of Sillago sihama and Rastrelliger kanagurta Fish
3.3. Isolation of Intestinal Bacteria from Sillago sihama and Rastrelliger kanagurta Fish
3.4. Primary Screening of α-Amylase-Producing Bacteria
3.5. Secondary Screening of α-Amylase-Producing Bacteria and Measurement of Enzyme Activity
3.6. Identification of Potential α-Amylase Enzyme-Producing Bacteria
3.7. Effect of pH and Temperature on α-Amylase Activity
3.8. Effects of pH and Temperature on Enzyme Stability
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qeshmi, F.I.; Homaei, A.; Fernandes, P.; Hemmati, R.; Dijkstra, B.W.; Khajeh, K. Xylanases from marine microorganisms: A brief overview on scope, sources, features and potential applications. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2020, 1868, 140312. [Google Scholar] [CrossRef] [PubMed]
- Homaei, A. Immobilization of Penaeus merguiensis alkaline phosphatase on gold nanorods for heavy metal detection. Ecotoxicol. Environ. Saf. 2017, 136, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Homaei, A. Purification and biochemical properties of highly efficient alkaline phosphatase from Fenneropenaeus merguiensis brain. J. Mol. Catal. B Enzym. 2015, 118, 16–22. [Google Scholar] [CrossRef]
- Homaei, A.; Qeshmi, F.I. Purification and characterization of a robust thermostable protease isolated from Bacillus subtilis strain HR02 as an extremozyme. J. Appl. Microbiol. 2022, 133, 2779–2789. [Google Scholar] [CrossRef]
- Izadpanah Qeshmi, F.; Homaei, A.; Khajeh, K.; Kamrani, E.; Fernandes, P. Production of a Novel Marine Pseudomonas aeruginosa Recombinant L-Asparaginase: Insight on the Structure and Biochemical Characterization. Mar. Biotechnol. 2022, 24, 559–613. [Google Scholar] [CrossRef]
- Pasalari, A.; Homaei, A. Isolation and Molecular Identification of Xylanase-Producing Bacteria from Ulva flexuosa of the Persian Gulf. Processes 2022, 10, 1834. [Google Scholar] [CrossRef]
- Cahill, M.M. Bacterial flora of fishes: A review. Microb. Ecol. 1990, 19, 21–41. [Google Scholar] [CrossRef]
- Balaji, N.; Rajasekaran, K.M.; Kanipandian, N.; Vignesh, V.; Thirumurugan, R. Isolation and screening of proteolytic bacteria from freshwater fish Cyprinus carpio. Int. Multidiscip. Res. J. 2012, 2, 56–59. [Google Scholar]
- Beygmoradi, A.; Homaei, A.; Hemmati, R.; Santos-Moriano, P.; Hormigo, D.; Fernández-Lucas, J. Marine chitinolytic enzymes, a biotechnological treasure hidden in the ocean? Appl. Microbiol. Biotechnol. 2018, 102, 9937–9948. [Google Scholar] [CrossRef]
- Homaei, A.; Ghanbarzadeh, M.; Monsef, F. Biochemical features and kinetic properties of α-amylases from marine organisms. Int. J. Biol. Macromol. 2016, 83, 306–314. [Google Scholar] [CrossRef]
- Reddy, N.; Nimmagadda, A.; Rao, K.S. An overview of the microbial α-amylase family. Afr. J. Biotechnol. 2003, 2, 645–648. [Google Scholar]
- Sivaramakrishnan, S.; Gangadharan, D.; Nampoothiri, K.M.; Soccol, C.R.; Pandey, A. α-Amylases from microbial sources—An overview on recent developments. Food Technol. Biotechnol. 2006, 44, 173–184. [Google Scholar]
- Makhdoumi, A. Bacterial diversity in south coast of the Caspian Sea: Culture-dependent and culture-independent survey. Casp. J. Environ. Sci. 2018, 16, 259–269. [Google Scholar]
- Jabir, T.; Jesmi, Y.; Vipindas, P.V.; Hatha, A.M. Diversity of nitrogen fixing bacterial communities in the coastal sediments of southeastern Arabian Sea (SEAS). Deep. Sea Res. Part II Top. Stud. Oceanogr. 2018, 156, 51–59. [Google Scholar] [CrossRef]
- Jackson, C.R.; Churchill, P.F.; Roden, E.E. Successional changes in bacterial assemblage structure during epilithic biofilm development. Ecology 2001, 82, 555–566. [Google Scholar] [CrossRef]
- Castro, R.A.; Quecine, M.C.; Lacava, P.T.; Batista, B.D.; Luvizotto, D.M.; Marcon, J.; Ferreira, A.; Melo, I.S.; Azevedo, J.L. Isolation and enzyme bioprospection of endophytic bacteria associated with plants of Brazilian mangrove ecosystem. SpringerPlus 2014, 3, 382. [Google Scholar] [CrossRef] [Green Version]
- Ktari, N.; Ben Khaled, H.; Nasri, R.; Jellouli, K.; Ghorbel, S.; Nasri, M. Trypsin from zebra blenny (Salaria basilisca) viscera: Purification, characterisation and potential application as a detergent additive. Food Chem. 2012, 130, 467–474. [Google Scholar] [CrossRef]
- Debashish, G.; Malay, S.; Barindra, S.; Joydeep, M. Marine enzymes. Mar. Biotechnol. I 2005, 96, 189–218. [Google Scholar]
- Stoll, B.J.; Hansen, N.I.; Sánchez, P.J.; Faix, R.G.; Poindexter, B.B.; Van Meurs, K.P. Early onset neonatal sepsis: The burden of group B Streptococcal and E. coli disease continues. Pediatrics 2011, 127, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Ye, F.; Yang, R.; Hua, X.; Shen, Q.; Zhao, W.; Zhang, W. Modification of stevioside using transglucosylation activity of Bacillus amyloliquefaciens α-amylase to reduce its bitter aftertaste. LWT—Food Sci. Technol. 2013, 51, 524–530. [Google Scholar] [CrossRef]
- Pérez-Carrillo, E.; Serna-Saldívar, S.O.; Alvarez, M.M.; Cortes-Callejas, M.L. Effect of sorghum decortication and use of protease before liquefaction with thermoresistant α-amylase on efficiency of bioethanol production. Cereal. Chem. 2008, 85, 792–798. [Google Scholar] [CrossRef]
- Demirkan, E.S.; Mikami, B.; Adachi, M.; Higasa, T.; Utsumi, S. α-Amylase from B. amyloliquefaciens: Purification, characterization, raw starch degradation and expression in E. coli. Process Biochem. 2005, 40, 2629–2636. [Google Scholar] [CrossRef]
- Dong, G.; Vieille, C.; Savchenko, A.; Zeikus, J.G. Cloning, sequencing, and expression of the gene encoding extracellular alpha-amylase from Pyrococcus furiosus and biochemical characterization of the recombinant enzyme. Appl. Environ. Microbiol. 1997, 63, 3569–3576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahri, S.; Homaei, A.; Mosaddegh, E. Zinc sulfide-chitosan hybrid nanoparticles as a robust surface for immobilization of Sillago sihama α-amylase. Colloids Surf. B Biointerfaces 2022, 218, 112754. [Google Scholar] [CrossRef] [PubMed]
- Fajingbesi, A.O.; Anzaku, A.A.; Akande, M.; Emmanuel, I.A.; Akwashiki, O. Production of protease enzyme from fish guts using Pseudomonas fluorescens, Enterobacter cloacae and Bacillus megaterium. J. Clin. Path. Lab. Med. 2018, 2, 1–7. [Google Scholar]
- Farha, A.K.; Tr, T.; Purushothaman, A.; Salam, J.A.; Hatha, A.M. Phylogenetic diversity and biotechnological potentials of marine bacteria from continental slope of eastern Arabian Sea. J. Genet. Eng. Biotechnol. 2018, 16, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Ramezani-Pour, N.; Badoei-Dalfard, A.; Namaki-Shoushtari, A.; Karami, Z. Nitrile-metabolizing potential of Bacillus cereus strain FA12; Nitrilase production, purification, and characterization. Biocatal. Biotransformation 2015, 33, 156–166. [Google Scholar] [CrossRef]
- Izadpanah Qeshmi, F.; Javadpour, S.; Malekzadeh, K.; Tamadoni Jahromi, S.; Rahimzadeh, M. Persian gulf is a bioresource of potent L-asparaginase producing bacteria: Isolation & molecular differentiating. Int. J. Environ. Res. 2014, 8, 813–818. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Isolated Bacterial Code | Investigating the Growth of Bacteria in the Culture Medium Containing Starch | The Diameter of the Halo Formed after Lugol’s Addition |
|---|---|---|---|
| 1 | T1 | Positive | Strong |
| 2 | T2 | Negative | No halo |
| 3 | T3 | Negative | No halo |
| 4 | T4 | Negative | No halo |
| 5 | T5 | S5 Positive | Strong |
| 6 | T6 | Negative | Weak |
| 7 | T7 | Positive | Medium |
| 8 | T8 | Negative | No halo |
| Number | Isolated Bacterial Code | Investigating the Growth of Bacteria in the Culture Medium Containing Starch | The Diameter of the Halo Formed after Lugol’s Addition |
|---|---|---|---|
| 1 | S1 | Positive | Weak |
| 2 | S2 | Negative | No halo |
| 3 | S3 | Negative | No halo |
| 4 | S4 | Negative | No halo |
| 5 | S5 | S5 Positive | Strong |
| 6 | S6 | Negative | No halo |
| 7 | S7 | Positive | Medium |
| 8 | S8 | Negative | No halo |
| 9 | S9 | Negative | No halo |
| 10 | S10 | S Negative | No halo |
| 11 | S11 | Positive | Medium |
| 12 | S12 | Positive | Strong |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erfanimoghadam, M.R.; Homaei, A. Identification of New Amylolytic Enzymes from Marine Symbiotic Bacteria of Bacillus Species. Catalysts 2023, 13, 183. https://doi.org/10.3390/catal13010183
Erfanimoghadam MR, Homaei A. Identification of New Amylolytic Enzymes from Marine Symbiotic Bacteria of Bacillus Species. Catalysts. 2023; 13(1):183. https://doi.org/10.3390/catal13010183
Chicago/Turabian StyleErfanimoghadam, Mohammad Reza, and Ahmad Homaei. 2023. "Identification of New Amylolytic Enzymes from Marine Symbiotic Bacteria of Bacillus Species" Catalysts 13, no. 1: 183. https://doi.org/10.3390/catal13010183
APA StyleErfanimoghadam, M. R., & Homaei, A. (2023). Identification of New Amylolytic Enzymes from Marine Symbiotic Bacteria of Bacillus Species. Catalysts, 13(1), 183. https://doi.org/10.3390/catal13010183