


Advances and Prospects of d-Tagatose Production Based on a Biocatalytic Isomerization Pathway


 and
and 
Abstract
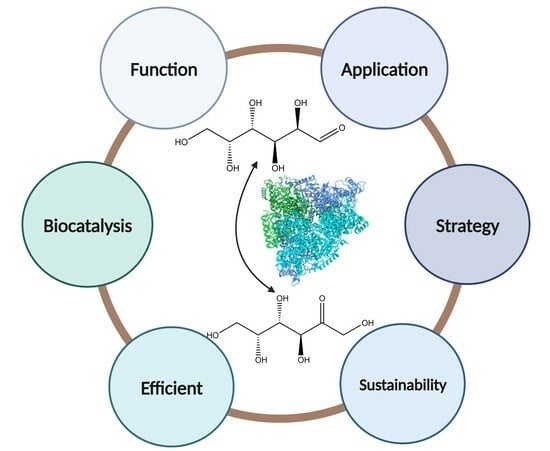
:
1. Introduction
2. The Production of d-Tagatose by L-AIs
2.1. Molecular Structure and Catalytic Mechanism of L-AIs
2.2. Properties of L-AIs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Source | Temparature Optima (°C) | pH Optima | Metal Ion Requirement | d-Tagatose Yield (%) | kcat/KM (mM−1 min−1) (d-galactose) | Reference |
|---|---|---|---|---|---|---|
| Lactobacillus plantarum NC8 | 60 | 7.5 | Mn2+, Co2+ | 30 | 1.6 | [42] |
| Anoxybacillus flavithermus | 95 | 10.5 | none | 60 | 5.16 | [43] |
| Bacillus coagulans NL01 | 60 | 7.5 | Mn2+, Co2+ | 32 | 1.0 | [44] |
| Pediococcus pentosaceus PC-5 | 50 | 6.0 | Mn2+, Co2+ | 50 | 2.9 | [45] |
| Clostridium hylemonae | 50 | 7.5 | Mg2+ | 46 | 3.69 | [46] |
| Lactobacillus sakei 23K | 35 | 5 | Mn2+, Mg2+ | 36 | 10.3 | [47] |
| Lactobacillus fermentum CGMCC2921 | 65 | 6.5 | Mn2+, Co2+ | 55 | 9.02 | [48] |
| Bifidobacterium adolescentis | 55 | 6.5 | Mn2+, Fe2+, Zn2+ Ca2+ | 56.7 | 9.3 | [49] |
| Thermotoga maritima | 90 | 7.5 | Mn2+, Co2+ | 56 | 8.5 | [50] |
| Thermotoga neapolitana | 85 | 7.0 | Mn2+, Co2+ | 68 | 3.24 | [51] |
| Lactococcus lactis | 50 | 8.0 | Mg2+, Mn2+,Co2+ | 42.4 | NA | [52] |
| Bacillus thermoglucosidasius | 40 | 7.0 | Mn2+ | 45.6 | 2.8 | [53] |
| Arthrobacter species 22c | 52 | 8.0 | Mg2+, Mn2+, Ca2+ | 30 | 0.14 | [54] |
| Shewanella species ANA-3 | 15–35 | 5.5–6.5 | Mn2+ | 34 | NA | [55] |
| Bacillus licheniformis | 50 | 7.5 | Mn2+, Co2+ | NA | slight activity | [56] |
| Bacillus subtilis 168 | 32 | 7.5 | Mn2+ | NA | NA | [57] |
2.3. Production of d-Tagatose Using Lactose as Raw Material
3. Strategies for Improving the Production of d-Tagatose by L-AIs
3.1. Protein Engineering
3.2. Immobilization
3.3. Application of Chemicals
3.4. Membrane Selection Difference
3.5. Multiple Strategy Combinations
4. Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Beerens, K.; Desmet, T.; Soetaert, W. Enzymes for the biocatalytic production of rare sugars. J. Ind. Microbiol. Biotechnol. 2012, 39, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Chikkerur, J.; Roy, S.C.; Dhali, A.; Kolte, A.P.; Sridhar, M.; Samanta, A.K. Tagatose as a potential nutraceutical: Production, properties, biological roles, and applications. J. Food Sci. 2018, 83, 2699–2709. [Google Scholar] [CrossRef] [PubMed]
- Donner, T.W.; Magder, L.S.; Zarbalian, K. Dietary supplementation with d-tagatose in subjects with type 2 diabetes leads to weight loss and raises high-density lipoprotein cholesterol. Nutr. Res. 2010, 30, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Buemann, B.; Toubro, S.; Raben, A.; Blundell, J.; Astrup, A. The acute effect of d-tagatose on food intake in human subjects. Br. J. Nutr. 2007, 84, 227–231. [Google Scholar] [CrossRef]
- Ensor, M.; Banfield, A.B.; Smith, R.R.; Williams, J.; Lodder, R.A. Safety and efficacy of d-tagatose in glycemic control in subjects with type 2 diabetes. J. Endocrinol. Diabetes Obes. 2015, 3, 1065. [Google Scholar]
- Seoane, J.; Gomez-Foix, A.M.; O’Doherty, R.M.; Gomez-Ara, C.; Newgard, C.B.; Guinovart, J.J. Glucose 6-phosphate produced by glucokinase, but not hexokinase I, promotes the activation of hepatic glycogen synthase. J. Biol. Chem. 1996, 271, 23756–23760. [Google Scholar] [CrossRef]
- Donner, T.; Wilber, J.; Ostrowski, D. d-tagatose, a novel hexose: Acute effects on carbohydrate tolerance in subjects with and without type 2 diabetes. Diabetes Obes. Metab. 1999, 1, 285–291. [Google Scholar] [CrossRef]
- Espinosa, I.; Fogelfeld, L. Tagatose: From a sweetener to a new diabetic medication? Expert Opin. Investig. Drugs 2010, 19, 285–294. [Google Scholar] [CrossRef]
- Levin, G.V. Tagatose, the new GRAS sweetener and health product. J. Med. Food 2002, 5, 23–36. [Google Scholar] [CrossRef]
- Lu, Y.; Levin, G.V. Removal and prevention of dental plaque with d-tagatose. Int. J. Cosmet. Sci. 2002, 24, 225–234. [Google Scholar] [CrossRef]
- Mayumi, S.; Kuboniwa, M.; Sakanaka, A.; Hashino, E.; Ishikawa, A.; Ijima, Y.; Amano, A. Potential of prebiotic d-tagatose for prevention of oral disease. Front. Cell Infect. Microbiol. 2021, 11, 767944. [Google Scholar] [CrossRef] [PubMed]
- Hasibul, K.; Nakayama-Imaohji, H.; Hashimoto, M.; Yamasaki, H.; Ogawa, T.; Waki, J.; Tada, A.; Yoneda, S.; Tokuda, M.; Miyake, M.; et al. d-Tagatose inhibits the growth and biofilm formation of Streptococcus mutans. Mol. Med. Rep. 2018, 17, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Ensor, M.; Williams, J.; Smith, R.; Banfield, A.; Lodder, R.A. Effects of three low-doses of d-tagatose on glycemic control over six months in subjects with mild type 2 diabetes mellitus under control with diet and exercise. J. Endocrinol. Diabetes Obes. 2014, 2, 1057. [Google Scholar] [PubMed]
- Levin, G.V.; Zehner, L.R.; Saunders, J.P.; Beadle, J.R. Sugar substitutes: Their energy values, bulk characteristics, and potential health benefits. Am. J. Clin. Nutr. 1995, 62, 1161S–1168S. [Google Scholar] [CrossRef]
- Normen, L.; Laerke, H.N.; Jensen, B.B.; Langkilde, A.M.; Andersson, H. Small-bowel absorption of d-tagatose and related effects on carbohydrate digestibility: An ileostomy study. Am. J. Clin. Nutr. 2001, 73, 105–110. [Google Scholar] [CrossRef]
- Laerke, H.N.; Jensen, B.B.; Hojsgaard, S. In vitro fermentation pattern of d-tagatose is affected by adaptation of the microbiota from the gastrointestinal tract of pigs. J. Nutr. 2000, 130, 1772–1779. [Google Scholar] [CrossRef]
- Fujimaru, T.; Park, J.-H.; Lim, J. Sensory characteristics and relative sweetness of tagatose and other sweeteners. J. Food Sci. 2012, 77, S323–S328. [Google Scholar] [CrossRef]
- Armstrong, L.M.; Luecke, K.J.; Bell, L.N. Consumer evaluation of bakery product flavour as affected by incorporating the prebiotic tagatose. Int. J. Food Sci. Technol. 2009, 44, 815–819. [Google Scholar] [CrossRef]
- Torrico, D.D.; Tam, J.; Fuentes, S.; Gonzalez Viejo, C.; Dunshea, F.R. d-tagatose as a sucrose substitute and its effect on the physico-chemical properties and acceptability of strawberry-flavored yogurt. Foods 2019, 8, 256. [Google Scholar] [CrossRef]
- Hirst, E.; Hough, L.; Jones, J. Composition of the gum of Sterculia setigera: Occurrence of d-tagatose in nature. Nature 1949, 163, 177. [Google Scholar] [CrossRef]
- Drabo, P.; Delidovich, I. Catalytic isomerization of galactose into tagatose in the presence of bases and Lewis acids. Catal. Commun. 2018, 107, 24–28. [Google Scholar] [CrossRef]
- Gao, D.-M.; Kobayashi, T.; Adachi, S. Production of rare sugars from common sugars in subcritical aqueous ethanol. Food Chem. 2015, 175, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Milasing, N.; Khuwijitjaru, P.; Adachi, S. Isomerization of galactose to tagatose using arginine as a green catalyst. Food Chem. 2023, 398, 133858. [Google Scholar] [CrossRef] [PubMed]
- Murzin, D.Y.; Murzina, E.V.; Aho, A.; Kazakova, M.A.; Selyutin, A.G.; Kubicka, D.; Kuznetsov, V.L.; Simakova, I.L. Aldose to ketose interconversion: Galactose and arabinose isomerization over heterogeneous catalysts. Catal. Sci. Technol. 2017, 7, 5321–5331. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Z.; Jin, Q.; Feng, D.; Lee, J. Isomerization of galactose to tagatose: Recent advances in non-enzymatic isomerization. J. Agric. Food Chem. 2023, 71, 4228–4234. [Google Scholar] [CrossRef] [PubMed]
- Granstrom, T.B.; Takata, G.; Tokuda, M.; Izumori, K. Izumoring: A novel and complete strategy for bioproduction of rare sugars. J. Biosci. Bioeng. 2004, 97, 89–94. [Google Scholar] [CrossRef]
- Shin, K.-C.; Lee, T.-E.; Seo, M.-J.; Kim, D.W.; Kang, L.-W.; Oh, D.-K. Development of tagaturonate 3-epimerase into tagatose 4-epimerase with a biocatalytic route from fructose to tagatose. ACS Catal. 2020, 10, 12212–12222. [Google Scholar] [CrossRef]
- Jeon, E.J.; Lee, Y.-M.; Choi, E.J.; Kim, S.-B.; Jeong, K.J. Production of tagatose by whole-cell bioconversion from fructose using Corynebacterium glutamicum. Biotechnol. Bioprocess. Eng. 2023, 28, 419–427. [Google Scholar] [CrossRef]
- Lu, F.; Xu, W.; Zhang, W.; Guang, C.; Mu, W. Polyol dehydrogenases: Intermediate role in the bioconversion of rare sugars and alcohols. Appl. Microbiol. Biotechnol. 2019, 103, 6473–6481. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Singh, R.; Kang, Y.C.; Zhao, H.; Lee, J.K. Cloning and characterization of a galactitol 2-dehydrogenase from Rhizobium legumenosarum and its application in d-tagatose production. Enzym. Microb. Technol. 2014, 58–59, 44–51. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, J.; Chen, Q.; Wu, H.; Mu, W. Sugar alcohols derived from lactose: Lactitol, galactitol, and sorbitol. Appl. Microbiol. Biotechnol. 2020, 104, 9487–9495. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Bedekar, A.A.; Liu, J.J.; Jin, Y.S.; Rao, C.V. Production of galactitol from galactose by the oleaginous yeast Rhodosporidium toruloides IFO0880. Biotechnol. Biofuels 2019, 12, 250. [Google Scholar] [CrossRef]
- Zhang, G.; An, Y.; Zabed, H.M.; Yun, J.; Parvez, A.; Zhao, M.; Zhang, C.; Ravikumar, Y.; Li, J.; Qi, X. Rewiring Bacillus subtilis and bioprocess optimization for oxidoreductive reaction-mediated biosynthesis of d-tagatose. Bioresour. Technol. 2023, 389, 129843. [Google Scholar] [CrossRef]
- Liu, J.-J.; Zhang, G.-C.; Kwak, S.; Oh, E.J.; Yun, E.J.; Chomvong, K.; Cate, J.H.D.; Jin, Y.-S. Overcoming the thermodynamic equilibrium of an isomerization reaction through oxidoreductive reactions for biotransformation. Nat. Commun. 2019, 10, 1356. [Google Scholar] [CrossRef]
- Lee, S.H.; Hong, S.H.; Kim, K.R.; Oh, D.K. High-yield production of pure tagatose from fructose by a three-step enzymatic cascade reaction. Biotechnol. Lett. 2017, 39, 1141–1148. [Google Scholar] [CrossRef]
- Han, P.; Wang, X.; Li, Y.; Wu, H.; Shi, T.; Shi, J. Synthesis of a healthy sweetener d-tagatose from starch catalyzed by semiartificial cell factories. J. Agric. Food Chem. 2023, 71, 3813–3820. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Li, S.; Feng, X.; Liang, J.; Xu, H. l-Arabinose isomerase and its use for biotechnological production of rare sugars. Appl. Microbiol. Biotechnol. 2014, 98, 8869–8878. [Google Scholar] [CrossRef] [PubMed]
- Manjasetty, B.A.; Chance, M.R. Crystal structure of Escherichia coli l-arabinose isomerase (ECAI), the putative target of biological tagatose production. J. Mol. Biol. 2006, 360, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Li, S.; Feng, X.; Zhan, Y.; Xu, H. Function of aspartic acid residues in optimum pH control of l-arabinose isomerase from Lactobacillus fermentum. Appl. Microbiol. Biotechnol. 2014, 98, 3987–3996. [Google Scholar] [CrossRef]
- Shin, K.C.; Seo, M.J.; Kim, S.J.; Kim, Y.S.; Park, C.S. Characterization of l-Arabinose Isomerase from Klebsiella pneumoniae and Its Application in the Production of d-Tagatose from d-Galactose. Appl. Sci. 2022, 12, 4696. [Google Scholar] [CrossRef]
- Prabhu, P.; Jeya, M.; Lee, J.-K. Probing the molecular determinant for the catalytic efficiency of l-arabinose isomerase from Bacillus licheniformis. Appl. Environ. Microbiol. 2010, 76, 1653–1660. [Google Scholar] [CrossRef] [PubMed]
- Chouayekh, H.; Bejar, W.; Rhimi, M.; Jelleli, K.; Mseddi, M.; Bejar, S. Characterization of an l-arabinose isomerase from the Lactobacillus plantarum NC8 strain showing pronounced stability at acidic pH. FEMS Microbiol. Lett. 2007, 277, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, Y.; Liu, A.; Sun, Y. Identification and characterization of a novel l-arabinose isomerase from Anoxybacillus flavithermus useful in d-tagatose production. Extremophiles 2011, 15, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Mei, W.; Wang, L.; Zang, Y.; Zheng, Z.; Ouyang, J. Characterization of an l-arabinose isomerase from Bacillus coagulans NL01 and its application for d-tagatose production. BMC Biotechnol. 2016, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Men, Y.; Zhu, Y.; Zhang, L.; Kang, Z.; Izumori, K.; Sun, Y.; Ma, Y. Enzymatic conversion of d-galactose to d-tagatose: Cloning, overexpression and characterization of l-arabinose isomerase from Pediococcus pentosaceus PC-5. Microbiol. Res. 2014, 169, 171–178. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Hong, M.G.; Chang, P.S.; Lee, B.H.; Yoo, S.H. Biochemical properties of l-arabinose isomerase from Clostridium hylemonae to produce d-tagatose as a functional sweetener. PLoS ONE 2018, 13, e0196099. [Google Scholar] [CrossRef]
- Rhimi, M.; Ilhammami, R.; Bajic, G.; Boudebbouze, S.; Maguin, E.; Haser, R.; Aghajari, N. The acid tolerant l-arabinose isomerase from the food grade Lactobacillus sakei 23K is an attractive d-tagatose producer. Bioresour. Technol. 2010, 101, 9171–9177. [Google Scholar] [CrossRef]
- Xu, Z.; Qing, Y.; Li, S.; Feng, X.; Xu, H.; Ouyang, P. A novel l-arabinose isomerase from Lactobacillus fermentum CGMCC2921 for d-tagatose production: Gene cloning, purification and characterization. J. Mol. Catal. B Enzym. 2011, 70, 1–7. [Google Scholar] [CrossRef]
- Zhang, G.; An, Y.; Parvez, A.; Zabed, H.M.; Yun, J.; Qi, X. Exploring a highly d-galactose specific l-arabinose isomerase from Bifidobacterium adolescentis for d-tagatose production. Front. Bioeng. Biotechnol. 2020, 8, 377. [Google Scholar] [CrossRef]
- Lee, D.W.; Jang, H.J.; Choe, E.A.; Kim, B.C.; Lee, S.J.; Kim, S.B.; Hong, Y.H.; Pyun, Y.R. Characterization of a thermostable l-arabinose (d-galactose) isomerase from the hyperthermophilic eubacterium Thermotoga maritima. Appl. Environ. Microbiol. 2004, 70, 1397–1404. [Google Scholar] [CrossRef]
- Kim, B.-C.; Lee, Y.-H.; Lee, H.-S.; Lee, D.-W.; Choe, E.-A.; Pyun, Y.-R. Cloning, expression and characterization of l-arabinose isomerase from Thermotoga neapolitana: Bioconversion of d-galactose to d-tagatose using the enzyme. FEMS Microbiol. Lett. 2002, 212, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, Z.; Ma, M.; Zhao, G.; Chang, R.; Si, H.; Dai, M. A novel Lactococcus lactis l-arabinose isomerase for d-tagatose production from lactose. Food Biosci. 2022, 48, 101765. [Google Scholar] [CrossRef]
- Seo, M.J. Characterization of an l-arabinose isomerase from Bacillus thermoglucosidasius for d-tagatose production. Biosci. Biotechnol. Biochem. 2013, 77, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Wanarska, M.; Kur, J. A method for the production of d-tagatose using a recombinant Pichia pastoris strain secreting β-d-galactosidase from Arthrobacter chlorophenolicus and a recombinant l-arabinose isomerase from Arthrobacter sp. 22c. Microb. Cell Factories 2012, 11, 113. [Google Scholar] [CrossRef]
- Rhimi, M.; Bajic, G.; Ilhammami, R.; Boudebbouze, S.; Maguin, E.; Haser, R.; Aghajari, N. The acid-tolerant l-arabinose isomerase from the mesophilic Shewanella sp. ANA-3 is highly active at low temperatures. Microb. Cell Factories 2011, 10, 96. [Google Scholar] [CrossRef]
- Prabhu, P.; Tiwari, M.K.; Jeya, M.; Gunasekaran, P.; Kim, I.W.; Lee, J.K. Cloning and characterization of a novel l-arabinose isomerase from Bacillus licheniformis. Appl. Microbiol. Biotechnol. 2008, 81, 283–290. [Google Scholar] [CrossRef]
- Kim, J.H.; Prabhu, P.; Jeya, M.; Tiwari, M.K.; Moon, H.J.; Singh, R.K.; Lee, J.K. Characterization of an l-arabinose isomerase from Bacillus subtilis. Appl. Microbiol. Biotechnol. 2010, 85, 1839–1847. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, W.; Zhang, T.; Jiang, B.; Mu, W. l-arabinose isomerases: Characteristics, modification, and application. Trends Food Sci. Technol. 2018, 78, 25–33. [Google Scholar] [CrossRef]
- Jayamuthunagai, J.; Gautam, P.; Srisowmeya, G.; Chakravarthy, M. Biocatalytic production of d-tagatose: A potential rare sugar with versatile applications. Crit. Rev. Food Sci. Nutr. 2017, 57, 3430–3437. [Google Scholar] [CrossRef]
- Fan, C.; Liu, K.; Zhang, T.; Zhou, L.; Xue, D.; Jiang, B.; Mu, W. Biochemical characterization of a thermostable l-arabinose isomerase from a thermoacidophilic bacterium, Alicyclobacillus hesperidum URH17-3-68. J. Mol. Catal. B Enzym. 2014, 102, 120–126. [Google Scholar] [CrossRef]
- Hong, Y.-H.; Lee, D.-W.; Pyun, Y.-R.; Lee, S.H. Creation of metal-independent hyperthermophilic l-arabinose isomerase by homologous recombination. J. Agric. Food Chem. 2011, 59, 12939–12947. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.M.; Lee, Y.-J.; Cao, T.-P.; Shin, S.-M.; Park, M.-K.; Lee, H.-S.; di Luccio, E.; Kim, S.-B.; Lee, S.-J.; Lee, S.J.; et al. Structure of the thermophilic l-Arabinose isomerase from Geobacillus kaustophilus reveals metal-mediated intersubunit interactions for activity and thermostability. Arch. Biochem. Biophys. 2016, 596, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fan, C.; Zhang, T.; Jiang, B.; Mu, W. Cloning, expression, and characterization of a novel l-arabinose isomerase from the psychrotolerant bacterium Pseudoalteromonas haloplanktis. Mol. Biotechnol. 2016, 58, 695–706. [Google Scholar] [CrossRef]
- Patel, M.J.; Patel, A.T.; Akhani, R.; Dedania, S.; Patel, D.H. Bioproduction of d-tagatose from d-galactose using phosphoglucose isomerase from Pseudomonas aeruginosa PAO1. Appl. Biochem. Biotechnol. 2016, 179, 715–727. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lee, D.W.; Lee, S.J.; Choe, E.A.; Kim, S.B.; Lee, Y.H.; Cheigh, C.I.; Pyun, Y.R. Production of d-tagatose at high temperatures using immobilized Escherichia coli cells expressing l-arabinose isomerase from Thermotoga neapolitana. Biotechnol. Lett. 2007, 29, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Kim, P. Current studies on biological tagatose production using l-arabinose isomerase: A review and future perspective. Appl. Microbiol. Biotechnol. 2004, 65, 243–249. [Google Scholar] [CrossRef]
- Dobbs, C.M.; Bell, L.N. Storage stability of tagatose in buffer solutions of various compositions. Food Res. Int. 2010, 43, 382–386. [Google Scholar] [CrossRef]
- Luecke, K.J.; Bell, L.N. Thermal stability of tagatose in solution. J. Food Sci. 2010, 75, C346–C351. [Google Scholar] [CrossRef]
- Lim, B.-C.; Kim, H.-J.; Oh, D.-K. Tagatose production with pH control in a stirred tank reactor containing immobilized l-arabinose isomerase from Thermotoga neapolitana. Appl. Biochem. Biotechnol. 2007, 149, 245–253. [Google Scholar] [CrossRef]
- Rhimi, M.; Chouayekh, H.; Gouillouard, I.; Maguin, E.; Bejar, S. Production of d-tagatose, a low caloric sweetener during milk fermentation using l-arabinose isomerase. Bioresour. Technol. 2011, 102, 3309–3315. [Google Scholar] [CrossRef]
- Zhang, G.; Zabed, H.M.; Yun, J.; Yuan, J.; Zhang, Y.; Wang, Y.; Qi, X. Two-stage biosynthesis of d-tagatose from milk whey powder by an engineered Escherichia coli strain expressing l-arabinose isomerase from Lactobacillus plantarum. Bioresour. Technol. 2020, 305, 123010. [Google Scholar] [CrossRef] [PubMed]
- Rhimi, M.; Messaoud, E.B.; Borgi, M.A.; Khadra, K.B.; Bejar, S. Co-expression of l-arabinose isomerase and d-glucose isomerase in E. coli and development of an efficient process producing simultaneously d-tagatose and d-fructose. Enzym. Microb. Technol. 2007, 40, 1531–1537. [Google Scholar] [CrossRef]
- Zheng, Z.; Xie, J.; Liu, P.; Li, X.; Ouyang, J. Elegant and efficient biotransformation for dual production of d-tagatose and bioethanol from cheese whey powder. J. Agric. Food Chem. 2019, 67, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zabed, H.M.; An, Y.; Yun, J.; Huang, J.; Zhang, Y.; Li, X.; Wang, J.; Ravikumar, Y.; Qi, X. Biocatalytic conversion of a lactose-rich dairy waste into d-tagatose, d-arabitol and galactitol using sequential whole cell and fermentation technologies. Bioresour. Technol. 2022, 358, 127422. [Google Scholar] [CrossRef]
- Ravikumar, Y.; Ponpandian, L.N.; Zhang, G.; Yun, J.; Qi, X. Harnessing l-arabinose isomerase for biological production of d-tagatose: Recent advances and its applications. Trends Food Sci. Technol. 2021, 107, 16–30. [Google Scholar] [CrossRef]
- Jayaraman, A.B.; Kandasamy, T.; Venkataraman, D.; Meenakshisundaram, S. Rational design of Shewanella sp. l-arabinose isomerase for d-galactose isomerase activity under mesophilic conditions. Enzym. Microb. Technol. 2021, 147, 109796. [Google Scholar] [CrossRef]
- Laksmi, F.A.; Arai, S.; Tsurumaru, H.; Nakamura, Y.; Saksono, B.; Tokunaga, M.; Ishibashi, M. Improved substrate specificity for d-galactose of l-arabinose isomerase for industrial application. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 1084–1091. [Google Scholar] [CrossRef]
- Kim, B.-J.; Hong, S.-H.; Shin, K.-C.; Jo, Y.-S.; Oh, D.-K. Characterization of a F280N variant of l-arabinose isomerase from Geobacillus thermodenitrificans identified as ad-galactose isomerase. Appl. Microbiol. Biotechnol. 2014, 98, 9271–9281. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, J.H.; Oh, H.J.; Oh, D.K. Characterization of a mutated Geobacillus stearothermophilus l-arabinose isomerase that increases the production rate of d-tagatose. J. Appl. Microbiol. 2006, 101, 213–221. [Google Scholar] [CrossRef]
- Zhao, L.; Ma, Z.; Wang, Q.; Hu, M.; Zhang, J.; Chen, L.; Shi, G.; Ding, Z. Engineering the thermostability of sucrose synthase by reshaping the subunit interaction contributes to efficient UDP-glucose production. J. Agric. Food Chem. 2023, 71, 3832–3841. [Google Scholar] [CrossRef]
- Han, Z.; Li, N.; Xu, H.; Xu, Z. Improved thermostability and robustness of l-arabinose isomerase by C-terminal elongation and its application in rare sugar production. Biochem. Biophys. Res. Commun. 2022, 637, 224–231. [Google Scholar] [CrossRef]
- Staudigl, P.; Haltrich, D.; Peterbauer, C.K. l-Arabinose Isomerase and d-Xylose Isomerase from Lactobacillus reuteri: Characterization, Coexpression in the Food Grade Host Lactobacillus plantarum, and Application in the Conversion of d-Galactose and d-Glucose. J. Agric. Food Chem. 2014, 62, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Rhimi, M.; Aghajari, N.; Juy, M.; Chouayekh, H.; Maguin, E.; Haser, R.; Bejar, S. Rational design of Bacillus stearothermophilus US100 l-arabinose isomerase: Potential applications for d-tagatose production. Biochimie 2009, 91, 650–653. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Chen, M.; Liu, X.; Zhai, Y.; Liu, X.W.; Zhang, H.; Xiao, M.; Wang, P. Bioconversion of d-galactose to d-tagatose: Continuous packed bed reaction with an immobilized thermostable l-arabinose isomerase and efficient purification by selective microbial degradation. Appl. Microbiol. Biotechnol. 2012, 93, 1469–1474. [Google Scholar] [CrossRef]
- Bortone, N.; Fidaleo, M. Stabilization of immobilized l-arabinose isomerase for the production of d-tagatose from d-galactose. Biotechnol. Prog. 2020, 36, e3033. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, S.; Xu, H.; Wu, L.; Xu, Z.; Liu, J.; Feng, X. Efficient production of d-tagatose using a food-grade surface display system. J. Agric. Food Chem. 2014, 62, 6756–6762. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; An, Y.; Yun, J.; Yang, M.; Magocha, T.A.; Zhu, J.; Xue, Y.; Qi, Y.; Hossain, Z.; Sun, W.; et al. Enhanced d-tagatose production by spore surface-displayed l-arabinose isomerase from isolated Lactobacillus brevis PC16 and biotransformation. Bioresour. Technol. 2018, 247, 940–946. [Google Scholar] [CrossRef]
- Jebors, S.; Tauran, Y.; Aghajari, N.; Boudebbouze, S.; Maguin, E.; Haser, R.; Coleman, A.W.; Rhimi, M. Supramolecular stabilization of acid tolerant l-arabinose isomerase from Lactobacillus sakei. Chem. Commun. 2011, 47, 12307–12309. [Google Scholar] [CrossRef]
- Oh, D.-K. Tagatose: Properties, applications, and biotechnological processes. Appl. Microbiol. Biotechnol. 2007, 76, 1–8. [Google Scholar] [CrossRef]
- Lim, B.C.; Kim, H.J.; Oh, D.K. High production of d-tagatose by the addition of boric acid. Biotechnol. Prog. 2007, 23, 824–828. [Google Scholar] [CrossRef]
- Kim, J.H.; Lim, B.C.; Yeom, S.J.; Kim, Y.S.; Kim, H.J.; Lee, J.K.; Lee, S.H.; Kim, S.W.; Oh, D.K. Differential selectivity of the Escherichia coli cell membrane shifts the equilibrium for the enzyme-catalyzed isomerization of galactose to tagatose. Appl. Environ. Microbiol. 2008, 74, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.C.; Sim, D.H.; Seo, M.J.; Oh, D.K. Increased Production of Food-Grade d-Tagatose from d-Galactose by Permeabilized and Immobilized Cells of Corynebacterium glutamicum, a GRAS Host, Expressing d-Galactose Isomerase from Geobacillus thermodenitrificans. J. Agric. Food Chem. 2016, 64, 8146–8153. [Google Scholar] [CrossRef] [PubMed]
- Bober, J.R.; Nair, N.U. Galactose to tagatose isomerization at moderate temperatures with high conversion and productivity. Nat. Commun. 2019, 10, 4548. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xu, W.; Wu, H.; Guang, C.; Zhang, W.; Mu, W. An overview of d-galactose utilization through microbial fermentation and enzyme-catalyzed conversion. Appl. Microbiol. Biotechnol. 2021, 105, 7161–7170. [Google Scholar] [CrossRef]
- Tsermoula, P.; Khakimov, B.; Nielsen, J.H.; Engelsen, S.B. WHEY—The waste-stream that became more valuable than the food product. Trends Food Sci. Technol. 2021, 118, 230–241. [Google Scholar] [CrossRef]
- Kim, H.M.; Song, Y.; Wi, S.G.; Bae, H.J. Production of d-tagatose and bioethanol from onion waste by an intergrating bioprocess. J. Biotechnol. 2017, 260, 84–90. [Google Scholar] [CrossRef]
- Sha, F.; Zheng, Y.; Chen, J.; Chen, K.; Cao, F.; Yan, M.; Ouyang, P. d-Tagatose manufacture through bio-oxidation of galactitol derived from waste xylose mother liquor. Green. Chem. 2018, 20, 2382–2391. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, T.; Xin, Y.; Qin, L.; Kong, J. Biotechnological production of d-tagatose from lactose using metabolically engineering Lactiplantibacillus plantarum. LWT 2021, 142, 110995. [Google Scholar] [CrossRef]
- Cheon, J.; Kim, S.B.; Park, S.W.; Han, J.K.; Kim, P. Characterization of l-Arabinose Isomerase in Bacillus subtilis, a GRAS Host, for the Production of Edible Tagatose. Food Biotechnol. 2009, 23, 8–16. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, R.; Wang, Q.; Hu, M.; Li, Z.; Xu, M.; Yang, T.; Zhang, R.; Rao, Z. Production of d-tagatose by whole-cell conversion of recombinant Bacillus subtilis in the absence of antibiotics. Biology 2021, 10, 1343. [Google Scholar] [CrossRef]






| Methods | Advantages | Disadvantages | Significance | |
|---|---|---|---|---|
| Chemical methods | Low cost | High temperature and pressure; environmental pollution; Low yield | d-galactose to d-tagatose | |
| Biological methods | Tagatose 4-epimerase | Low cost; Mild reaction conditions | Low yield; Rare sources | d-fructose to d-tagatose |
| Galactitol dehydrogenase | High yield; Mild reaction conditions | High cost; Need for cofactor Rare sources | d-galactitol to d-tagatose | |
| l-arabinose isomerase | Low cost; Mild reaction conditions; High yield; Many sources | Need for metal ions | d-galactose to d-tagatose | |
| phosphorylation-dephosphorylation cascade | High yield; Mild reaction conditions | Need for multiple enzymes and steps; Need for ATP | d-fructose or starch to d-tagatose | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, P.; Wang, Q.; Ren, K.; Zhang, Z.; Xu, T.; Xu, M.; Zhang, X.; Rao, Z. Advances and Prospects of d-Tagatose Production Based on a Biocatalytic Isomerization Pathway. Catalysts 2023, 13, 1437. https://doi.org/10.3390/catal13111437
Miao P, Wang Q, Ren K, Zhang Z, Xu T, Xu M, Zhang X, Rao Z. Advances and Prospects of d-Tagatose Production Based on a Biocatalytic Isomerization Pathway. Catalysts. 2023; 13(11):1437. https://doi.org/10.3390/catal13111437
Chicago/Turabian StyleMiao, Peiyu, Qiang Wang, Kexin Ren, Zigang Zhang, Tongtong Xu, Meijuan Xu, Xian Zhang, and Zhiming Rao. 2023. "Advances and Prospects of d-Tagatose Production Based on a Biocatalytic Isomerization Pathway" Catalysts 13, no. 11: 1437. https://doi.org/10.3390/catal13111437
APA StyleMiao, P., Wang, Q., Ren, K., Zhang, Z., Xu, T., Xu, M., Zhang, X., & Rao, Z. (2023). Advances and Prospects of d-Tagatose Production Based on a Biocatalytic Isomerization Pathway. Catalysts, 13(11), 1437. https://doi.org/10.3390/catal13111437