1. Introduction
Beyond serving as the fundamental components of life, proteins and DNA are also key building blocks for nanoscale self-assemblies. Biomolecular assemblies, ranging from 2D arrays to 3D crystals, are useful tools for structural biology, bio-catalysis, and biomedical applications [
1,
2,
3]. Porous biomolecular crystals can even act as macromolecular scaffolds [
4], providing structural details to guest macromolecules [
5]. However, downstream applications of interest, including X-ray diffraction, are hindered by crystal fragility and intolerance to solvent conditions other than the crystal growth solution. In this study, we establish a protocol for the chemical ligation of DNA inside of crystals and we demonstrate structural resilience of crosslinked co-crystals which may further their application utility.
DNA assembly stability is a limiting factor for DNA nanotechnology and DNA crystals. While coding DNA sticky base overhangs can drive self-assembly, the non-covalent DNA base stacking interactions and Watson–Crick hydrogen bonds that stabilize the junctions are only stable under specific conditions. For example, crystallization conditions for DNA crystals typically feature high concentrations of divalent cations such as Mg(II) to balance the negative phosphate backbone of DNA [
6]. DNA–protein co-crystals may be similarly reliant on counterions, particularly if counterions stabilize the DNA–protein binding event [
7]. Crystal forms that bring DNA building blocks into close proximity are very sensitive to the counterion environment, and often dissolve or convert into a disordered aggregate when placed in water. To maximize application versatility, DNA structures should ideally be robust to solution variations, not just ionic strength but also temperature and pH [
1]. Introducing covalent bonds across DNA–DNA interfaces has the potential to dramatically improve crystal macro-structure stability and could also improve X-ray diffraction.
Bioconjugation, or crosslinking, is a well-established strategy to improve the structural integrity of protein and DNA crystals [
8]. The protein–protein interfaces found within protein crystals tend to be rich in primary amines and carboxylic acids. If all neighboring building blocks can be covalently linked, the resulting covalent organic framework can be a robust material. In traditional protein X-ray crystallography, glutaraldehyde, a highly reactive crosslinker, can increase crystal stability in varying solution conditions, and can even improve diffraction resolution [
9,
10]. In our previous work on protein crystals, we have found that glyoxal offers an effective alternative to glutaraldehyde [
11,
12]. Chemical crosslinking and photo-crosslinking methods for DNA crystals are also established in the literature [
13,
14,
15]; however, we wanted to focus on a protocol in which the crosslinking does not require a specific sequence of DNA and does not add atoms to the structure (a zero-length crosslink).
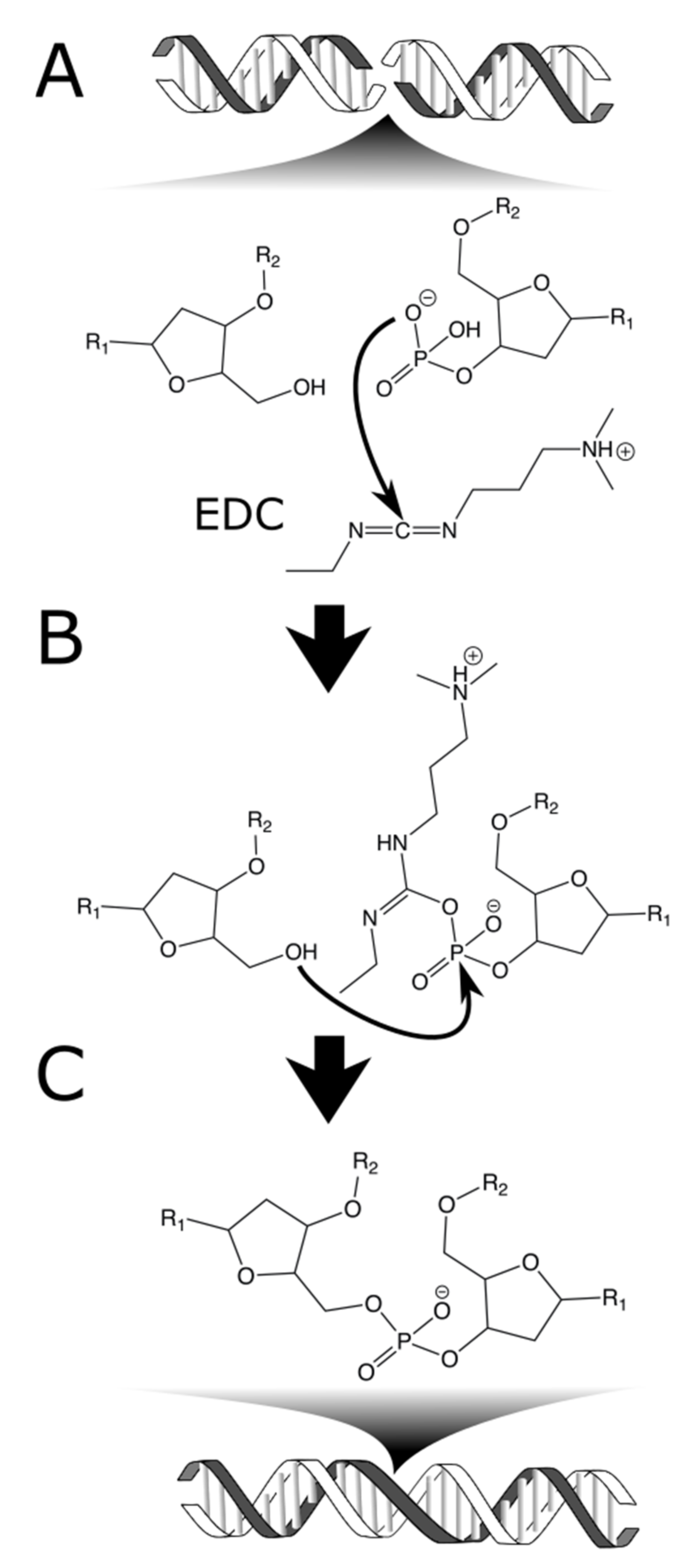
Arguably the most natural form of sequence-independent DNA crosslinking is
ligation, where the nicks dividing stacked dsDNA blocks are removed to generate longer contiguous DNA strands. For example, Li et al. used T4 DNA Ligase to ligate the DNA junctions within highly porous DNA crystals [
16]. This elegant approach is limited to crystals that have large enough solvent channels for enzyme ingress. Here, we sought to optimize a chemical ligation alternative to the use of ligase that would be applicable to crystals with both large and small pores.
Our chemical ligation chemistry relies on 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide (EDC), a water-soluble carbodiimide [
8]. EDC is widely used, especially in protein conjugation, to crosslink primary amines to carboxylic acids. A less common chemistry for EDC is the activation of a terminal phosphate such that a suitably placed nucleophile can displace the leaving group [
17]. When that nucleophile is the hydroxyl of a neighboring DNA strand, this chemistry results in a zero-length crosslink: a scar-less chemical ligation of DNA (
Figure 1). EDC has been used to ligate dsDNA hairpins in solution [
17], to link the phosphate backbone of stacked DNA in liquid crystals [
18] and to stabilize a 600 nucleotide DNA origami structure [
19]. Our work represents the first ligation via EDC of co-crystals containing protein and DNA. We show that EDC crosslinking dramatically increases crystal stability at the macroscale and does not prevent destroy the crystal nanostructure (i.e., treated crystals are still suitable for study via X-ray diffraction).
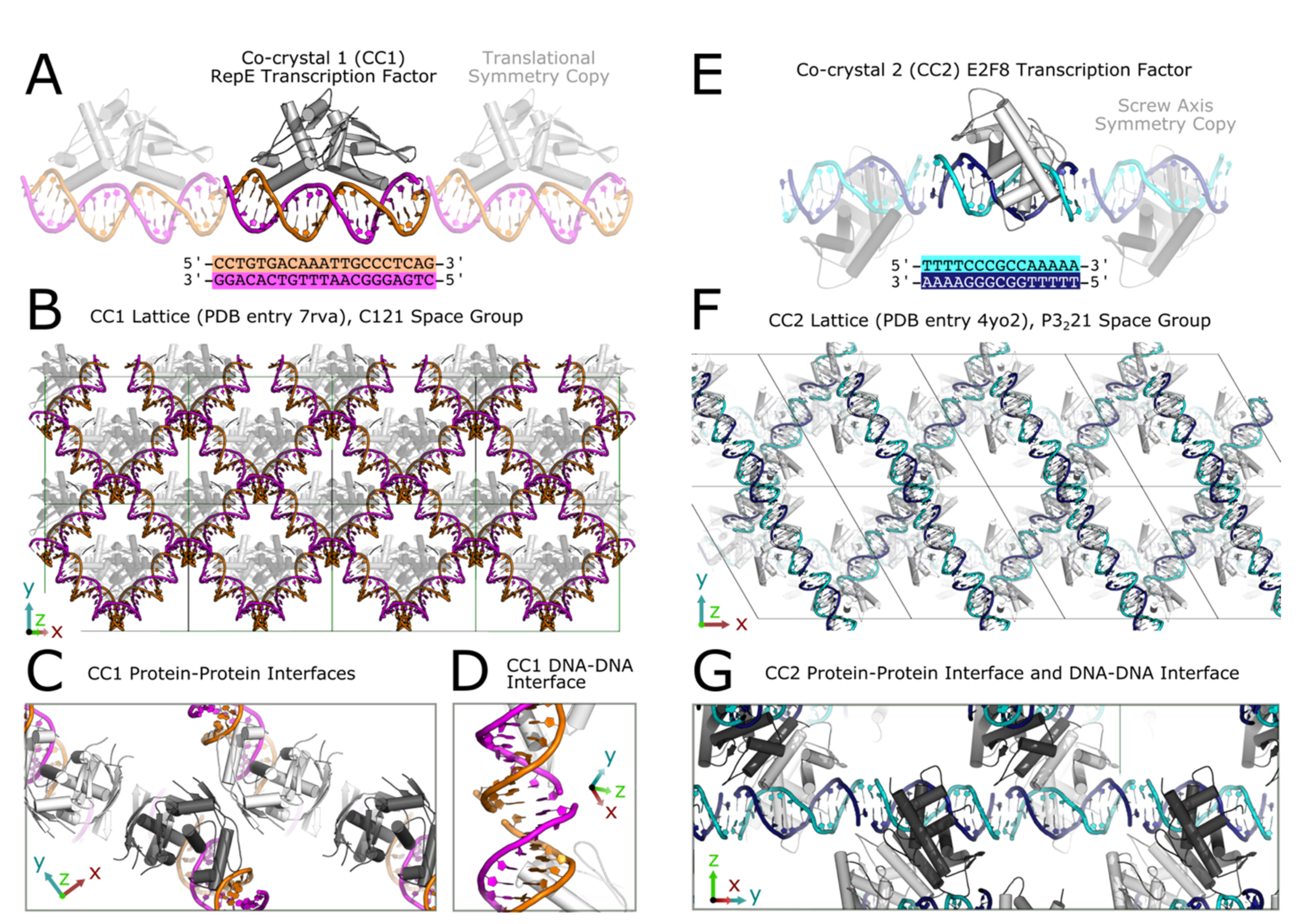
To demonstrate generality, we chemically ligate two different co-crystals of DNA-binding proteins containing stacked DNA–DNA interfaces (
Figure 2). For convenience, we will refer to crystals of the RepE54 transcription factor bound to cognate 21-mer dsDNA as Co-Crystal One (CC1) (
Figure 2A–D) and we will refer to crystals of the E2F8 transcription factor bound to cognate 15-mer dsDNA as Co-Crystal Two (CC2) (
Figure 2E–G). The asymmetric unit for each co-crystal consists of a DNA-binding protein and short, cognate DNA duplex. Both co-crystals have existing models in the Protein Data Bank (PDB). CC1 is closely related to existing PDB entry 1rep, though the 1rep model corresponds to a crystal with differing DNA at the junction (
Table S1). CC2 is identical to existing PDB entry 4yo2. The CC1 and CC2 crystals used in this study consist of dsDNA that is either blunt-ended or carries terminal 5′ or 3′ phosphates (
Figure 3). In each co-crystal system, the crosslinking variables tested were terminal 5′ vs. 3′ phosphates, crosslinking time, EDC concentration, and repeated EDC dose. After EDC crosslinking, co-crystals had dramatically increased structural integrity with respect to changes in the solution condition.
To show foundational feasibility for biomedical applications, we demonstrated that crosslinked co-crystals remain robust in aqueous environments, blood serum, and at pH values found in the stomach (pH 2.0) or lysosomes (pH 4.5). Therefore, the EDC crosslinking results provided here may justify further investigation of chemically ligated co-crystals or pure DNA crystals as biomaterials. For scaffold-assisted crystallography [
3] it is also important to note that the crosslinked co-crystals still diffracted X-rays. The crosslinked crystals tested here diffracted nearly as well as non-crosslinked crystals (anecdotally, a typical ~0.3 Å resolution difference). Additionally, we showed that this chemical ligation method is independent of the DNA sequence at the DNA–DNA junction. For example, despite differing DNA sequences at the junctions of CC1 and CC2, chemical ligation was effective in both cases. In summary, EDC ligation is a practical approach for crosslinking DNA inside of crystals and the optimized chemical crosslinking shown can provide the stability needed for diverse downstream applications.
3. Results
3.1. Chemical Ligation in Co-Crystals
Within our two co-crystal families (CC1 and CC2), we observed clear evidence of chemical ligation of stacked DNA duplexes. Both co-crystals demonstrated broadly similar ligation results, emphasizing the generality of this ligation method to co-crystals in which blunt-ended DNA blocks are suitably positioned to resemble contiguous DNA. As shown in
Table 3, the PDB entries for the parent structures of both CC1 (7rva) and CC2 (4yo2) have junction step geometry that is reasonably comparable to contiguous B-DNA as calculated using x3DNA [
29]. Except for the twist and roll across the CC2 junction (as seen in PDB entry 4yo2), all step geometry parameters are within 2 standard deviations of the B-DNA mean. It is possible that other co-crystals in which the DNA–DNA junctions have a geometry less like contiguous DNA would resist ligation. Additionally, in the preliminary crosslinking tests shown here, the crosslinking was successfully independent of the sequence at the DNA ends. CC1 has GC/CG flanking ends while CC2 has AT/TA flanking ends. The sequence independence of this ligation strategy is advantageous for DNA structure design projects where the junction sequence may be constrained for functional reasons.
Table 3 also reports an interesting asymmetry between the two nick sites at the DNA–DNA junctions within the CC1 family of structures. We report the distance between C5′ and O3′ to avoid relying on the less certain O5′ position. For calibration, an idealized B-DNA model from x3dna had C5′ to O3′ distances of 2.73 Å for contiguous bases, but this span is variable (2.99 ± 0.17 Å) elsewhere within the dsDNA of PDB entry 7rva. One of the two CC1 nick sites, chain B, was invariably closer than chain A (e.g., 3.75 Å rather than 4.22 Å in CC1-3′P), and electron density suggested that this shorter gap (chain B) was more readily ligated.
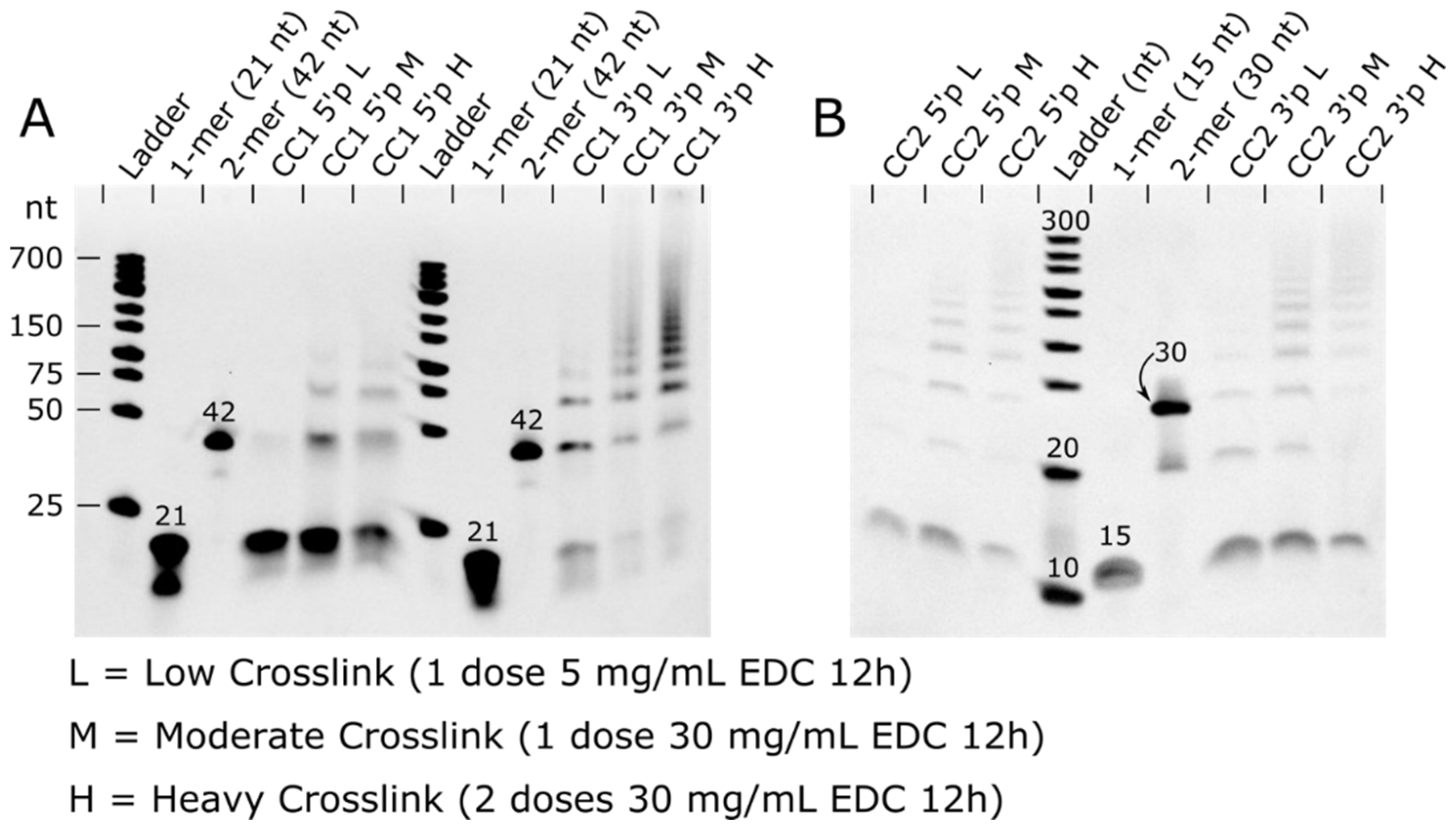
EDC crosslinking was tested for both 5′ and 3′ phosphate laden crystals. For CC1, the 3′ phosphate resulted in superior ligation yield than the 5′ phosphate in each trial (
Figure 4 and
Figures S7–S11). On the other hand, CC2 ligation yields had a modest difference in the ligation yield for 3′ and 5′ phosphates. Given the limited dataset, it is premature to conclude that 3′ phosphates will typically give a higher ligation yield within co-crystals.
In both systems, DNA ligation was dependent on the presence of the terminal phosphates as well as on the crystal template; control crystals lacking terminal phosphates yielded no observable ligation products (
Figure S10). Additionally, freely diffusing DNA blocks carrying terminal phosphates (but lacking the co-crystal scaffold) also yielded no observable ligation products when exposed to EDC (
Figure S10). This second control demonstrated that the scaffold was necessary for ensuring efficient ligation of blunt-ended DNA blocks. The absence of observable ligation for building blocks in the absence of the crystal “scaffold”, precludes a systematic study of the effects of precursor ligation on crystal growth. Future work will determine, as a function of sticky overhang length, the extent to which blocks with sticky overhangs can be ligated within crystals and in solution.
Crosslinking reaction time was clearly and directly related to ligation reaction yield during the first 12 h (
Figure S7). It was less clear if reaction yield was further improved by incubation beyond 12 h. Therefore, 12 h crosslinking incubations were used for the subsequent ligation optimization trials.
In the next series of experiments, we optimized EDC concentration for maximum ligation yield. We assayed the ligation product distribution as a function of concentration from 5 mg/mL EDC to 80 mg/mL EDC. As hypothesized, increasing the concentration of EDC increases the ligation of DNA duplexes in the co-crystals (
Figure 4 and
Figure S8). In CC1 trials, we did not see a noticeable increase in ligation beyond 30 mg/mL. However, in CC2 trials, there was improved ligation at 60 mg/mL. We also subjected the co-crystals to multiple fresh doses of EDC (30 mg/mL) to determine if we could achieve near 100% ligation. For both co-crystal systems, multiple doses of EDC did increase ligation yields (
Figure S9) but did not approach 100% ligation yields.
Reaction buffer components were critical for successful ligation. We observed, at the outset of this project, that the presence of magnesium chloride in the crosslinking buffer appeared to interfere with the crosslinking reaction. This was problematic because the CC1 crystal growth conditions contain a significant amount of magnesium chloride. In our crystallization trials, 30–120 mM magnesium chloride was required for growth [
21]. Additionally, there is a structural Mg(II) at the DNA–protein interface coordinated by Glu77 and Asp81. To circumvent the apparent deleterious role of Mg(II) on CC1 crosslinking, we replaced magnesium chloride with sodium chloride in the wash solution for all CC1 crosslinking trials. At the conclusion of the project, we again confirmed that Mg(II) was deleterious to ligation by adding Mg(II) to the optimized ligation protocol. Specifically, we verified that supplementing the crosslinking incubation buffer with 90 mM or 110 mM MgCl
2 noticeably reduced the ligation yield (
Figure S11). The exact role of Mg(II) in inhibiting the ligation reaction is not clear, but might involve reduced availability of the nucleophilic phosphate groups.
3.2. Ligation Model Compared to Experimental Co-Crystal Ligation
The ligation product distributions we experimentally obtained should shed light on the stochastic process of ligation. Using a destructive assay, densitometric analysis of electrophoresis results on ssDNA recovered from dissolved crystals, we quantified the population ratio of bands assigned to non-modified DNA blocks as well as fused 2-mer, 3-mer, etc. For selected gels, we also obtained TapeStation results (
Figure S1). The relative population of the end-product distribution was fairly consistent for gel band populations measured with TBE-urea gels in ImageJ compared to the automated TapeStation analysis (
Figure S1).
Next, we sought to calculate a global performance metric for the ligation yield,
PLIG, as the fraction of all possible DNA–DNA nick sites throughout a crystal that were ligated. To quantify the ligation yield throughout an entire crystal, we analyzed the implications of the final DNA product distribution recovered after the crystal is dissolved and the protein components are removed. If we count the number of DNA oligos of each length (
) that were present in the crystal, and we ignore edge effects, we can estimate the total number of single-stranded breaks (SSB) as
. For the same crystal, the estimated total number of original single-strand breaks (regardless of final ligation status) would be
. For example, adding a single fused 3-mer to the crystal increases the SSB tally by one, but increases the tally of all possible junctions by three. Then, to compute the total probability of encountering SSB, we calculate:
In the final equation,
is the mole fraction for
-mer oligos. Therefore, to estimate the
, we can use estimated mole fractions from electrophoresis and densitometry (
Figure S2 and
Figure 4 and
Table 4). Accurately calculating
does require including the small mole fractions for higher-order products (
Table S3) since longer products contribute proportionally more to
. To estimate the uncertainty in each
, we used 500 numerical trials in which random noise was added to
to mimic the densitometry measurement error. We used noise comparable to
for the highest-order ligation products (normal variate with standard deviation 0.03), such that the smallest
values would regularly fall to 0 after the addition of random noise.
Given the probability of encountering a single-stranded break in the crystal (PSSB), it is trivial to calculate the probability of each terminal phosphate having undergone ligation (PLIG), since PLIG = 1 − PSSB. In the context of the random ligation model (RLM), ligation events throughout the crystal are independent and occur with equal probability at all nick sites. This is a physically plausible model if the intra-crystal transport rate for EDC exceeds the rate of reaction. Therefore, the incidence of double-stranded breaks within the crystal should occur with the joint probability of independent events, PDSB = (PSSB)2.
This analysis of the electrophoresis experiments suggests that ~75% of the terminal phosphates within the most thoroughly crosslinked CC1-3′P crystal have undergone ligation. Furthermore, ~94% of the DNA–DNA junctions in this crystal had at least one ligated chain. The similarity in ligation yield for the medium- and high-dose cases leads to an important question. What factors are limiting the yield? Incomplete ligation could result if a random population of terminal phosphates are missing, or otherwise incapable of on-target ligation. We used simulations to verify that the predicted RLM product ratio did not change when we postulated that a random subset of nick sites is incapable of ligation. This makes sense because junctions that are randomly selected to be incapable of ligation are functionally equivalent to sites that are randomly selected to be ligated last.
It may also be possible that ligating one phosphate at a DNA–DNA junction would negatively affect neighboring ligation probabilities. However, evidence for such allostery is lacking. Instead, the observed product distributions for CC1 ligation outcomes (
Table 4), were close to the distributions predicted by the RLM (
Figure S12). One small but consistent deviation from the RLM was a lower 2-mer, and higher 3-mer population than predicted. This observation seems to preclude the simplest negative allostery scenario (where one ligation event would reduce the probability at flanking sites). We cannot rule out the possibility that this discrepancy is an artifact associated with the gel electrophoresis densitometry.
The CC2 ligation outcomes (
Table 4) were significantly less consistent with distributions predicted by the RLM. Once more, the 3-mer population was often higher than expected, frequently exceeding the 2-mer population (which never happens in the RLM). This effect also seemed to extend to anomalously common 4-mers. A more striking divergence from the RLM prediction was the high population of non-ligated 1-mer blocks. Regardless of the RLM fit, the significant difference between the 1-mer mole fractions and the
PSSB values obtained from all the mole fractions strongly implicates that the RLM is lacking.
To investigate, we tested biased ligation model simulations. One possible explanation is that the ligation outcomes were driven partially by kinetics and molecular transport phenomena. Hypothetically, ligation sites near the crystal exterior might be more likely to be ligated than possible sites near the crystal center since reactive molecules must traverse the outer layers to react the interior. To determine the likely implications of this scenario, we conducted biased random ligation simulations (Protocol S3) that increased the probability of ligation events near the surface, decreased the probability at the center, and terminated the random ligation process at a set PSSB threshold. Perhaps counterintuitively, this spatial bias increased the predicted 1-mer mole fraction. A high 1-mer fraction is partially consistent with the observed product distribution for CC2. The overall lower ligation yield achieved for CC2 crystals compared to CC1 is also consistent with the hypothesis that the CC2 crystal interior is systematically under-ligated. Alternately, it could be the case that one of the two symmetry-distinct nick sites in the CC2 lattice has a significantly lower ligation yield, and therefore one of the two DNA oligos will be over-represented in the 1-mer population.
3.3. Ligation Structural Details
Co-crystal structural details were revealed with X-ray diffraction at the Advanced Light Source beamline 4.2.2. Electrophoresis data (
Figure 4) suggest that the CC2 DNA is stacked as intended. However, while high-resolution diffraction for CC2 crystals should be possible (3.07 Å reported by Morgunova et al. (22)), our CC2 crystals have, to date, yielded poor diffraction (>10 Å). Therefore, we chose to focus on the CC1 crystals as the model crystals to observe ligation via X-ray diffraction”.
Here, we report five new crystal structures for CC1. We obtained a 1.89 Å dataset for the original co-crystal, which revealed additional details beyond the original model (PDB code: 1rep, 2.60 Å). Komori et al. varied the DNA building block to optimize resolution (20), finding that dangling Ts resulted in the best data. Our updated structure provides a rationale for this empirical observation. Specifically, one of the dangling T bases is resolved, and participates in a crystallographic contact. Removing the dangling Ts decreased the resolution of our native structures from 1.9 Å to 2.7 Å (CC1-5′p) or 3.01 Å (CC1-3′p). Once crystals were crosslinked with low (15 mg/mL 12 h) and heavy (2 doses 30 mg/mL 12 h) EDC, the crystals maintained diffraction, albeit with a moderate loss in diffraction (3.14 Å and 3.28 Å, respectively).
Models were refined with PHENIX [
26] and COOT [
27]. The electron density for the heavily ligated DNA junction was consistent with contiguous DNA, despite omitting the terminal phosphate throughout prior refinement calculations.
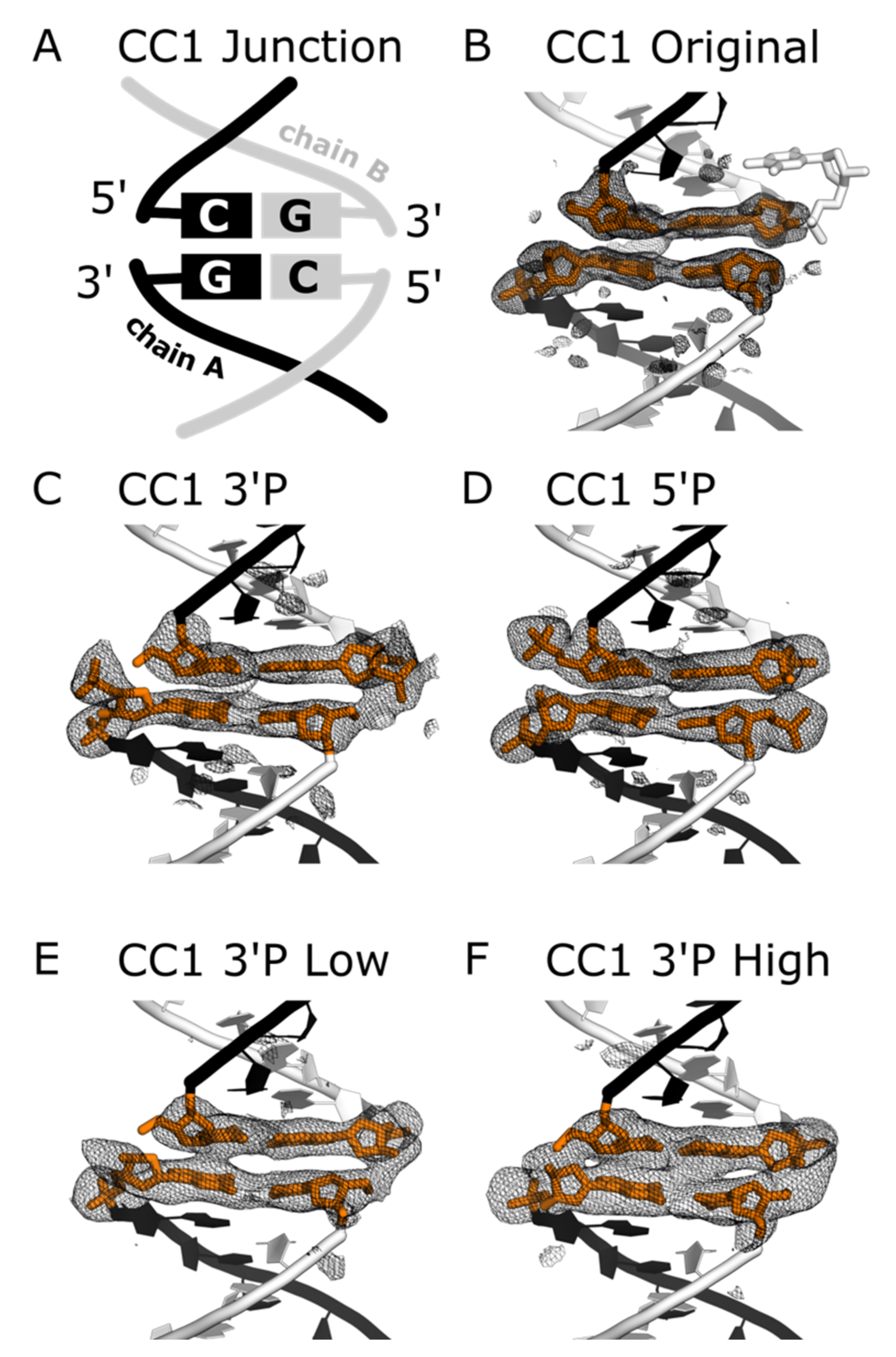
Figure 5 shows omit maps where any terminal phosphates are omitted, along with the bases flanking the junctions. The potential for overlapping electron density contributions from non-ligated and ligated phosphates makes it difficult to quantify occupancy. Nonetheless, we observed clear trends. Prior to ligation, the positions of 3′ phosphates (
Figure 5C) or 5′ phosphates (
Figure 5D) were reasonably clear.
Consistent with the lower distance between C5′ and O3′ for chain B (
Table 3), the electron density was invariably higher for the right hand nick (chain B:chain B). When contoured at 3.0 rmsd, the omit map electron density was even contiguous for the non-ligated CC1 3′P case (
Figure 5C). Notably, the maps for crystals subjected to EDC (
Figure 5E,F) are discovery maps in the sense that the models were refined in the absence of terminal 3′ phosphates. After light ligation (
Figure 5E), the omit map was not clearly changed. However, after heavy ligation (
Figure 5F), there was very strong electron density on the right and solid electron density in the left. Phosphates were added prior to submission to the PDB (entry 7spm) and our final refinement calculation for CC1 3′P High included bond length restraints between the model and its symmetry neighbor to ensure a reasonable phosphate geometry.
It is somewhat remarkable that ligation was visible in the electron density trend (
Figure 5C–F), despite the incomplete ligation yield suggested by the electrophoresis data (
Table 4). In principle, the clarity of the ligation sites in the electron density maps may vary depending on whether the X-ray beam is diffracting from a highly ligated region of the crystal.
3.4. Co-Crystal Stabilization Effects from Ligation and Crosslinking
To determine if crosslinked co-crystals may be suitable for various applications, including biomedical applications at physiologically relevant conditions, the co-crystals were crosslinked (20 h, 15 mg/mL EDC) and subjected to a panel of harsh conditions: a stomach acid mimic, a lysosomal fluid mimic, blood serum (bovine calf), and deionized water (
Figure 6). The conditions chosen, especially the stomach acid mimic and deionized water, were challenging for native crystals (no crosslink) since DNA-containing crystals typically require stabilizing counterions.
In the stomach acid mimic (0.01 M hydrochloric acid pH 2), the non-crosslinked co-crystals were observed to convert to an aggregate (
Figure S4). Remarkably, in the stomach acid solution, the entire set of crosslinked crystals demonstrated enhanced stability, not dissolving even after 7 days. The 3′ phosphate crosslinked crystals did not change macro-structure for at least 5 days in the harshly acidic environment (
Figure S4). Co-crystals without phosphates were also crosslinked and these crystals expanded dramatically in the acidic environment after 24 h (~430 ± 70% volume change), demonstrating the importance of the DNA ligation for crystal stability. Crosslinked co-crystals also maintained integrity in a lysosomal mimic buffer (pH 4.5) and blood serum with no measurable changes to the crystal dimensions after 24 and 72 h, respectively (
Figures S5 and S6).
In deionized water, the co-crystal stability resulting from crosslinking was exceptional (
Figure 7 and
Figure S3). Within one minute of transferring co-crystals to deionized water, non-crosslinked crystals (except for interesting exception CC2-3′P) completely dissolved or were converted to an aggregate. When the co-crystals were crosslinked (20 h, 15 mg/mL EDC), the crystals remained intact and lacked observable changes to their surface quality or dimensions for at least 7 days (
Figure 7 and
Figure S3). Interestingly, crosslinked co-crystals without terminal phosphates remained unperturbed, just like the 3′ and 5′ phosphorylated crystals. These results indicate that the protein–protein crosslinks created within the co-crystals were sufficient to maintain macroscopic crystal integrity in water. The distinct stability of crosslinked crystals in water confirmed our hypothesis that crystals can be stabilized with new covalent crosslinks. Specifically, the non-covalent interactions that make up crystals can be stabilized with chemical crosslinking and prevent crystals from degrading rapidly in an ion-environment (deionized water).
4. Discussion
Our strategy in this work was to identify an EDC ligation protocol (EDC concentration, incubation time, and repeated dosage regimen) that optimized reaction yield, without chasing diminishing returns. Accordingly, our final protocol uses 30 mg/mL EDC, an incubation time of 12 h, and two repeated doses within reaction sizes of approximately 200 microliters to ligate the DNA present within approximately 500 ng of co-crystals. Under these conditions, stacked DNA within co-crystals was reliably ligated to a significant extent. We used gel densitometry and detailed Gaussian peak fitting to estimate the fraction of the population for each ligated species (
Figure S2).
Global analysis of the ligation product distribution suggested that the most thoroughly crosslinked CC1 crystals feature ligation of approximately 75% of all possible ligation sites, covalently linking about 94% of the DNA–DNA junctions through one or more covalent bond. Ligation was corroborated by single-crystal XRD where we could directly observe ligation in electron density omit maps (
Figure 5).
Apart from small systematic deviations, the random ligation model (RLM,
Protocol S2) was able to fit the ligation product distribution for CC1 (
Figure S12). In contrast, the CC2 ligation results could not be fit to the RLM as accurately (
Figure S12). In particular, the CC2 crystals appeared to have a 1-mer mole fraction that was significantly larger than the total PDSB, which is inconsistent with the RLM. This could be explained by invoking transport limitations. Specifically, one way to boost the 1-mer mole fraction is if the exterior of the crystal has a higher ligation probability than the interior (
Protocol S3).
Previously mentioned in the introduction, EDC ligation of DNA has been reported in the literature in the context of DNA hairpins in solution, liquid DNA crystals, and DNA origami. Notably, there has not been a consensus for whether 5′ or 3′ phosphate placement results in a superior yield. Fraccia et al. used 3′ phosphates for the EDC ligation of liquid DNA crystals (15), whereas Kramer and Richert used 5′ phosphates for the EDC ligation of a DNA origami structure (14). Giving a comparison of 5′ versus 3′ phosphates, Obianyor et al. showed EDC ligation of a hairpin DNA structure and reported 95% ligation yield for DNA with 3′ phosphates whereas the 5′ phosphates yielded 40% ligation [
17]. They hypothesized the 3′ phosphate ligation reaction could benefit from a primary alcohol nucleophile (
Figure 1B) and the geometry difference of the two phosphate positions could contribute to reaction yields. Our data suggest that 3′ phosphates may be superior in the context of a crystal, though comparison between CC1 and CC2 suggests that the results may be system dependent. Our XRD data (
Figure 5) furthermore suggest that the results may vary for different nick sites within the same crystal.
It is not clear why 3′ phosphates were more readily ligated than 5′ phosphates in CC1. Conceivably, the rate limiting step for the ligation reaction may be the attack of the hydroxyl on the activated EDC intermediate. Perhaps the short-arm 3′-EDC intermediate is more accessible to the long-arm 5′ hydroxyl than a long-arm 5′-EDC intermediate is to a short-arm 3′ hydroxyl. Notably, one of the 3′ phosphates in the CC1 lattice (chain B) is close (5.65 Å) to a symmetry copy of itself (
Figure S13), whereas the 5′ phosphate is farther (9.29 Å). Therefore, 3′ phosphate ligation might be favored due to the greater reduction in electrostatic repulsion upon ligation. However, the CC2 ligation results were more balanced (albeit still favoring 3′ phosphates), suggesting that the relative efficacy of 3′ or 5′ phosphates will be system dependent.
Analyses via gel electrophoresis showed that the ligation yield increased concomitant with the EDC incubation time, but also that the reaction yield appeared to plateau short of full ligation. The cause is unclear. Transport considerations and EDC conjugation to protein sites complicate reaction modeling. One consideration is that the predicted active half-life for EDC in water at 298 K is sixteen hours [
17]. However, the ligation yields also appeared to plateau for repeated EDC dosing. Perhaps incomplete ligation is due to a small fraction of DNA strands lacking the necessary terminal phosphate. Alternately, perhaps some EDC-activated phosphates have been ligated to third-party molecules. Perhaps a small DNA strand population is missing a base. Further investigation may be worthwhile prior to future work that depends on near 100% ligation.
In addition to optimizing conditions for our two co-crystals, we have established a set of generalizable guidelines for DNA ligation within co-crystals regarding optimal reaction conditions, phosphate composition, and concentration of EDC. First, it is imperative to optimize the wash solution for each respective system, eliminating components that could interfere with crosslinking. Reactive amines and carboxylic acids are obvious components to eliminate, to avoid forming off-target species. Additionally, we empirically found that it was important to minimize the concentration of the standard divalent cation Mg(II). Re-introducing 90–110 mM Mg(II) into our optimized protocol, we observed a dramatic reduction in the yield (
Figure S11). Second, since we found that the best phosphate for ligation may depend on subtle geometry differences, we recommend testing both 5′ and 3′ phosphates for new co-crystal systems. Finally, the EDC concentration used for ligation of a new co-crystal may need to be optimized. Our co-crystals did not dissolve when introduced to crosslinking agents, with the highest concentration at 80 mg/mL. However, in past experiments, we found that the concentration of EDC in the crosslinking reaction drop can affect the integrity of co-crystals. Biomolecular crystals are typically fragile, and a drastic change in solution conditions can cause crystals to fall apart. Therefore, when working with a new system, we recommend testing a range of EDC concentrations. Dosing experiments may be necessary for systems that need a “gentle”, multistep transition to harsher conditions. These guidelines may apply to crystals composed of only DNA, as well.
With data for two example co-crystals, generalization is difficult. CC1 and CC2 differ in numerous ways (e.g., DNA length of 21 bp vs. 15 bp, crystal space group, different base pairs spanning the DNA–DNA junction, different DNA sequences in general including flanking base pairs) which makes it difficult to determine which variables may be predictive of ligation yield. Given our observation that ligation may be very sensitive to the nick geometry (
Table 3 and
Figure 5), we hypothesize that several factors will be particularly important due to their influence on the nick geometry. The DNA sequence at the junction, and to a lesser extent the flanking bases, will affect the base pair stacking energy, which would be expected to change the nick geometry probability distribution. Other nick-site ligation yield differences may be driven by the crystallographic symmetry, particularly the presence or absence of neighboring groups in addition to intrinsic geometry differences between the nick site (e.g., a slightly higher nick distance for chain A nick sites in CC1 crystals).
The crystal stability produced after the chemical ligation of stacked DNA within crystals opens the door for downstream applications, especially for DNA nanotechnology efforts. As shown here, even incomplete ligation can result in dramatic stabilization effects with tangible benefits to suitable application targets. No obvious EDC-induced crosslinks were visible at the two distinct protein–protein interfaces in the CC1 system. Further experiments will be needed to specifically seek and identify any EDC-induced protein–protein or DNA–protein conjugation.
It is possible that DNA ligation provided strong stabilizing effects because both CC1 and CC2 are held together by DNA–DNA junctions in two dimensions (
Figure 6 and
Figure 7). Essentially, by ligating the stacked DNA in these cases we are forming longer “threads” that are woven together. Stabilization of devices or materials is intriguing if this stabilization allows them to provide or preserve functionality in various biomedical contexts (e.g., in the digestive system, the blood stream, or within lysosomes). It may also be useful if crosslinking allows crystals to remain stable and diffract to high resolution under buffer conditions that mimic physiological conditions (e.g., inside the nucleus), thereby allowing XRD structure determination under conditions besides the idiosyncratic conditions that allow for co-crystal growth.
Along the same lines, one traditional concern crystallographers have regarding crosslinking chemistry is that subjecting a crystal to handling, buffer changes, and reactive chemicals, can degrade the diffraction resolution. For example, subjecting crystals to the common crosslinking agent, glutaraldehyde, can rapidly degrade diffraction resolution. However, supplying aldehydes via gentle vapor diffusion [
9] can improve outcomes. We have observed that using glyoxal and EDC can likewise result in negligible diffraction loss, particularly if the reactive chemistry is quenched [
11,
12]. In the case of CC1, we have once again found that carefully optimized crosslinking protocols can maintain diffraction. Another notable benefit of the EDC crosslinking method is that crystals were not “damaged” during the reaction chemistry. For comparison, when crosslinking HEWL crystals with glutaraldehyde, careful optimization was required to avoid forming cracks in the crystals [
31].
Future work may determine if the ligation yield differs for sticky overhang junctions compared to the blunt end junctions used in this work. Similarly, yield may also depend on the DNA bases that span the junction. That said, the current work suggests that the method may be sequence independent because the CC1 junction has a GC/CG and the CC2 junction has an AT/TA. In summary, the reported protocol is a reliable crosslinking strategy using the zero-length crosslinking agent EDC to affect DNA ligation at blunt-end DNA–DNA junctions held together by the co-crystal lattice. Post-ligation stability paves the way for biomedical applications.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}