
Mitochondrial Dysfunction Induced by Zinc Oxide Nanoparticles



Abstract
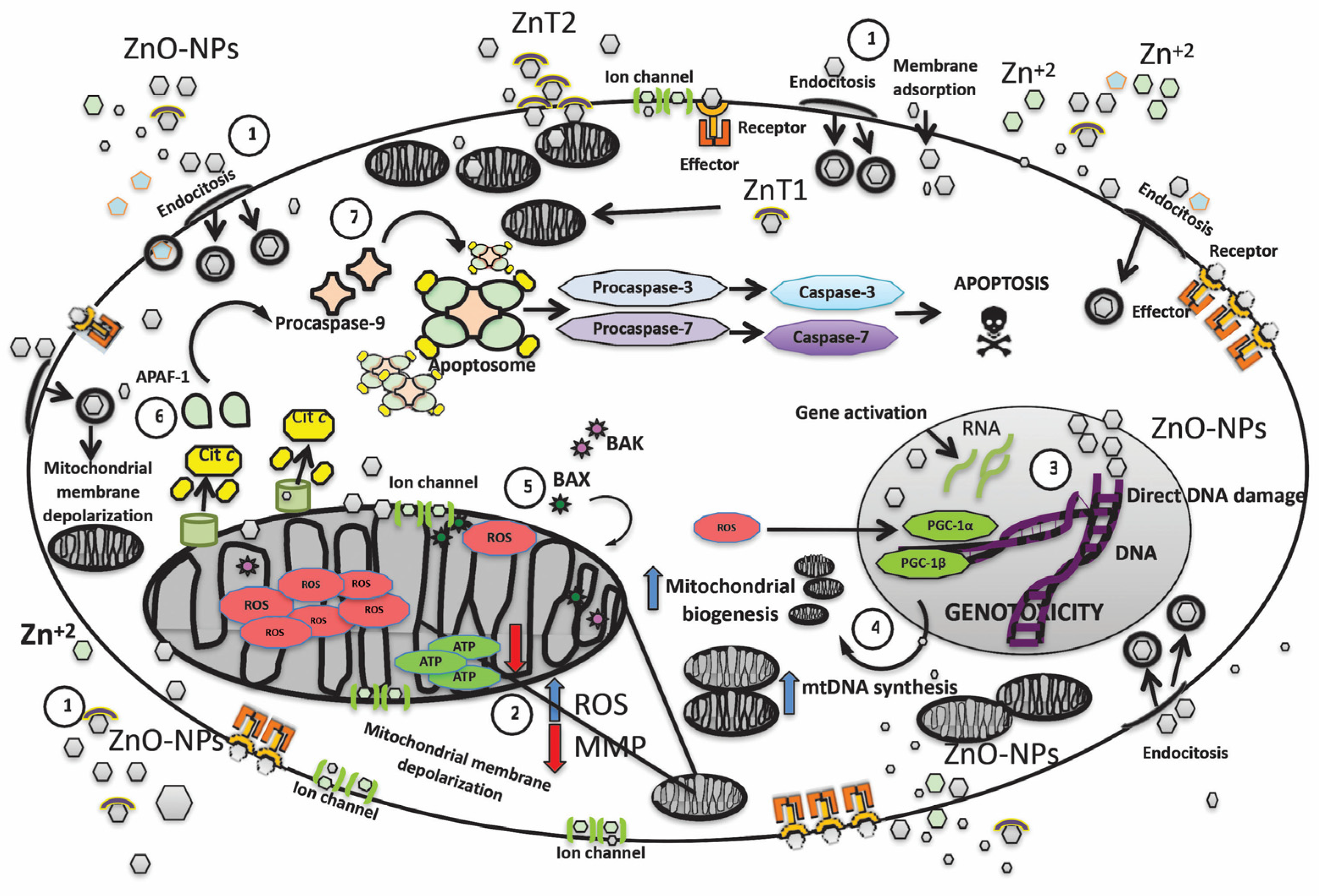
:1. Introduction
2. Impairment of Mitochondrial Biogenesis Induced by ZnO-NPs
3. Apoptosis Induction ZnO-NPs
4. Caspases Expression and ZnO-NPs
5. Effect of ZnO-NPs in Mitochondrial Membrane Potential
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mathew, E.N.; Hurst, M.N.; Wang, B.; Murthy, V.; Zhang, Y.; De Long, R.K. Interaction of Ras Binding Domain (RBD) by chemotherapeutic zinc oxide nanoparticles: Progress towards RAS pathway protein interference. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Bharathi, D.; Bhuvaneshwari, V. Synthesis of zinc oxide nanoparticles (ZnO NPs) using pure bioflavonoid rutin and their biomedical applications: Antibacterial, antioxidant and cytotoxic activities. Res. Chem. Intermed. 2019, 45, 2065–2078. [Google Scholar] [CrossRef]
- Danielsen, P.H.; Cao, Y.; Roursgaard, M.; Møller, P.; Loft, S. Endothelial cell activation, oxidative stress and inflammation induced by a panel of metal-based nanomaterials. Nanotoxicology 2015, 9, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Tang, M. Toxic effects and involved molecular pathways of nanoparticles on cells and subcellular organelles. J. Appl. Toxicol. 2020, 40, 16–36. [Google Scholar] [CrossRef]
- Hu, X.; Cook, S.; Wang, P.; Hwang, H.-M. In vitro evaluation of cytotoxicity of engineered metal oxide nanoparticles. Sci. Total Environ. 2009, 407, 3070–3072. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Shukla, R.K.; Saxena, N.; Parmar, D.; Das, M.; Dhawan, A. DNA damaging potential of zinc oxide nanoparticles in human epidermal cells. Toxicol. Lett. 2009, 185, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, K.; Stone, V.; Clouter, A.; Renwick, L.; Macnee, W. Ultrafine particles. Occup. Environ. Med. 2001, 58, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Wang, G.; Huang, J.; Zhang, Y.; Cheng, X.; Chuai, M.; Brand-Saberi, B.; Chen, G.; Jiang, X.; Yang, X. Zinc oxide nanoparticles exposure-induced oxidative stress restricts cranial neural crest development during chicken embryogenesis. Ecotoxicol. Environ. Saf. 2020, 194, 110415. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Liu, Y.; Wang, J.; Nie, Y.; Si, B.; Liu, Y.; Wang, X.; Chen, S.; Hei, T.K.; et al. Subcellular Targets of Zinc Oxide Nanoparticles during the Aging Process: Role of Cross-Talk between Mitochondrial Dysfunction and Endoplasmic Reticulum Stress in the Genotoxic Response. Toxicol. Sci. 2019, 171, 159–171. [Google Scholar] [CrossRef]
- Limo, M.J.; Sola-Rabada, A.; Boix, E.; Thota, V.; Westcott, Z.C.; Puddu, V.; Perry, C.C. Interactions between Metal Oxides and Biomolecules: From Fundamental Understanding to Applications. Chem. Rev. 2018, 118, 11118–11193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popov, L.D. Mitochondrial biogenesis: An update. J. Cell. Mol. Med. 2020, 24, 4892–4899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Sumberaz, P. Mitochondrial DNA Damage: Prevalence, Biological Consequence, and Emerging Pathways. Chem. Res. Toxicol. 2020, 33, 2491–2502. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.B.; Pekkurnaz, G. Mechanisms Orchestrating Mitochondrial Dynamics for Energy Homeostasis. J. Mol. Biol. 2018, 430, 3922–3941. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, D.; Zhang, D.; Li, P.; Gao, Y. Mitochondrial Protein Translation: Emerging Roles and Clinical Significance in Disease. Front. Cell Dev. Biol. 2021, 9, 675465. [Google Scholar] [CrossRef] [PubMed]
- Yokokawa, T.; Kido, K.; Suga, T.; Isaka, T.; Hayashi, T.; Fujita, S. Exercise-induced mitochondrial biogenesis coincides with the expression of mitochondrial translation factors in murine skeletal muscle. Physiol. Rep. 2018, 6, e13893. [Google Scholar] [CrossRef]
- Holt, A.G.; Davies, A.M. The significance of mitochondrial DNA half-life to the lifespan of post-mitotic cells. bioRxiv 2020. [Google Scholar] [CrossRef]
- Byrne, J.J.; Soh, M.S.; Chandhok, G.; Vijayaraghavan, T.; Teoh, J.S.; Crawford, S.; Cobham, A.E.; Yapa, N.M.B.; Mirth, C.K.; Neumann, B. Disruption of mitochondrial dynamics affects behaviour and lifespan in Caenorhabditis elegans. Cell. Mol. Life Sci. 2019, 76, 1967–1985. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Lendahl, U.; Nistér, M.; Zhao, J. Regulation of Mammalian Mitochondrial Dynamics: Opportunities and Challenges. Front. Endocrinol. 2020, 11, 374. [Google Scholar] [CrossRef]
- Zhang, C.S.; Lin, S.C. AMPK promotes autophagy by facilitating mitochondrial fission. Cell Metab. 2016, 23, 399–401. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Gureev, A.P.; Shaforostova, E.A.; Popov, V.N. Regulation of mitochondrial biogenesis as a way for active longevity: Interaction between the Nrf2 and PGC-1α signaling pathways. Front. Genet. 2019, 10, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, F.; Zhang, L.; Zhang, C.; Peng, H.; Lan, F.; Peng, S.; Liu, C.; Guo, J. Zinc oxide nanoparticles induce mitochondrial biogenesis impairment and cardiac dysfunction in human ipsc-derived cardiomyocytes. Int. J. Nanomed. 2020, 15, 2669–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousef, M.I.; Mutar, T.F.; Kamel, M.A.E.N. Hepato-renal toxicity of oral sub-chronic exposure to aluminum oxide and/or zinc oxide nanoparticles in rats. Toxicol. Rep. 2019, 6, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; McIntyre, R.L.; Janssens, G.E.; Houtkooper, R.H. Mitochondrial fission and fusion: A dynamic role in aging and potential target for age-related disease. Mech. Ageing Dev. 2020, 186, 111212. [Google Scholar] [CrossRef]
- Li, J.; Zhang, B.; Chang, X.; Gan, J.; Li, W.; Niu, S.; Kong, L.; Wu, T.; Zhang, T.; Tang, M.; et al. Silver nanoparticles modulate mitochondrial dynamics and biogenesis in HepG2 cells. Environ. Pollut. 2020, 256, 113430. [Google Scholar] [CrossRef] [PubMed]
- Aung, L.H.H.; Jumbo, J.C.C.; Wang, Y.; Li, P. Therapeutic potential and recent advances on targeting mitochondrial dynamics in cardiac hypertrophy: A concise review. Mol. Ther. Nucleic Acids 2021, 25, 416–443. [Google Scholar] [CrossRef]
- Babele, P.K.; Thakre, P.K.; Kumawat, R.; Tomar, R.S. Zinc oxide nanoparticles induce toxicity by affecting cell wall integrity pathway, mitochondrial function and lipid homeostasis in Saccharomyces cerevisiae. Chemosphere 2018, 213, 65–75. [Google Scholar] [CrossRef]
- Kumar Babele, P. Zinc oxide nanoparticles impose metabolic toxicity by de-regulating proteome and metabolome in Saccharomyces cerevisiae. Toxicol. Rep. 2019, 6, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.J.; Wilson, S. Zinc homeostasis in the secretory pathway in yeast. Curr. Opin. Chem. Biol. 2020, 55, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Chevallet, M.; Gallet, B.; Fuchs, A.; Jouneau, P.H.; Um, K.; Mintz, E.; Michaud-Soret, I. Metal homeostasis disruption and mitochondrial dysfunction in hepatocytes exposed to sub-toxic doses of zinc oxide nanoparticles. Nanoscale 2016, 8, 18495–18506. [Google Scholar] [CrossRef]
- Yu, K.N.; Yoon, T.J.; Minai-Tehrani, A.; Kim, J.E.; Park, S.J.; Jeong, M.S.; Ha, S.W.; Lee, J.K.; Kim, J.S.; Cho, M.H. Zinc oxide nanoparticle induced autophagic cell death and mitochondrial damage via reactive oxygen species generation. Toxicol. Vitr. 2013, 27, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.; Siddiqui, S.; Zia, Q.; Ahmad, E.; Jafri, A.; Arshad, M.; Jamal, A.; Alam, M.M.; Banawas, S.; Alshehri, B.A.; et al. Characterization and in vitro cytotoxic assessment of zinc oxide nano-particles in human epidermoid carcinoma cells. J. Environ. Chem. Eng. 2021, 9, 105636. [Google Scholar] [CrossRef]
- Liang, S.; Sun, K.; Wang, Y.; Dong, S.; Wang, C.; Liu, L.X.; Wu, Y.H. Role of Cyt-C/caspases-9,3, Bax/Bcl-2 and the FAS death receptor pathway in apoptosis induced by zinc oxide nanoparticles in human aortic endothelial cells and the protective effect by alpha-lipoic acid. Chem. Biol. Interact. 2016, 258, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, D.; Wang, Z.; Yin, X.; Wei, H.; Hu, W.; Chen, R.; Chen, C. Zinc oxide nanoparticles induce human tenon fibroblast apoptosis through reactive oxygen species and caspase signaling pathway. Arch. Biochem. Biophys. 2020, 683, 108324. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Naqvi, A.H.; Ahmad, M. Comparative study of the cytotoxic and genotoxic potentials of zinc oxide and titanium dioxide nanoparticles. Toxicol. Rep. 2015, 2, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Czyżowska, A.; Dyba, B.; Rudolphi-Szydło, E.; Barbasz, A. Structural and biochemical modifications of model and native membranes of human immune cells in response to the action of zinc oxide nanoparticles. J. Appl. Toxicol. 2021, 41, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Czyżowska, A.; Barbasz, A. Cytotoxicity of zinc oxide nanoparticles to innate and adaptive human immune cells. J. Appl. Toxicol. 2021, 41, 1425–1437. [Google Scholar] [CrossRef] [PubMed]
- Ahlam, A.A.; Shaniba, V.S.; Jayasree, P.R.; Manish Kumar, P.R. Spondias pinnata (L.f.) Kurz Leaf Extract Derived Zinc Oxide Nanoparticles Induce Dual Modes of Apoptotic-Necrotic Death in HCT 116 and K562 Cells. Biol. Trace Elem. Res. 2021, 199, 1778–1801. [Google Scholar] [CrossRef]
- Wang, S.W.; Lee, C.H.; Lin, M.S.; Chi, C.W.; Chen, Y.J.; Wang, G.S.; Liao, K.W.; Chiu, L.P.; Wu, S.H.; Huang, D.M.; et al. ZnO nanoparticles induced caspase-dependent apoptosis in gingival squamous cell carcinoma through mitochondrial dysfunction and p70s6K signaling pathway. Int. J. Mol. Sci. 2020, 21, 1612. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Gao, S.; Wang, S.; Xu, Z.; Wei, L. Zinc oxide nanoparticles induce toxicity in CAL 27 oral cancer cell lines by activating PINK1/Parkin-mediated mitophagy. Int. J. Nanomed. 2018, 13, 3441–3450. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Song, Y.; Vogt, R.D.; Liu, Y.; Luo, J.; Li, T. Bioavailability and cytotoxicity of Cerium- (IV), Copper- (II), and Zinc oxide nanoparticles to human intestinal and liver cells through food. Sci. Total Environ. 2020, 702, 134700. [Google Scholar] [CrossRef] [PubMed]
- Ananthalakshmi, R.; Rathinam, S.R.X.R.; Sadiq, A.M. Apoptotic Signalling of Huh7 Cancer Cells by Biofabricated Zinc Oxide Nanoparticles. J. Inorg. Organomet. Polym. Mater. 2021, 31, 1764–1773. [Google Scholar] [CrossRef]
- Wahab, R.; Siddiqui, M.A.; Saquib, Q.; Dwivedi, S.; Ahmad, J.; Musarrat, J.; Al-Khedhairy, A.A.; Shin, H.S. ZnO nanoparticles induced oxidative stress and apoptosis in HepG2 and MCF-7 cancer cells and their antibacterial activity. Colloids Surfaces B Biointerfaces 2014, 117, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Arasu, M.V.; Madankumar, A.; Theerthagiri, J.; Salla, S.; Prabu, S.; Kim, H.S.; Al-Dhabi, N.A.; Arokiyaraj, S.; Duraipandiyan, V. Synthesis and characterization of ZnO nanoflakes anchored carbon nanoplates for antioxidant and anticancer activity in MCF7 cell lines. Mater. Sci. Eng. C 2019, 102, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.J.; Alhadlaq, H.A.; Alshamsan, A.; Majeed Khan, M.A.; Ahamed, M. Aluminum doping tunes band gap energy level as well as oxidative stress-mediated cytotoxicity of ZnO nanoparticles in MCF-7 cells. Sci. Rep. 2015, 5, 13876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farasat, M.; Niazvand, F.; Khorsandi, L. Zinc oxide nanoparticles induce necroptosis and inhibit autophagy in MCF-7 human breast cancer cells. Biologia 2020, 75, 161–174. [Google Scholar] [CrossRef]
- Li, Z.; Guo, D.; Yin, X.; Ding, S.; Shen, M.; Zhang, R.; Wang, Y.; Xu, R. Zinc oxide nanoparticles induce human multiple myeloma cell death via reactive oxygen species and Cyt-C/Apaf-1/Caspase-9/Caspase-3 signaling pathway in vitro. Biomed. Pharmacother. 2020, 122, 109712. [Google Scholar] [CrossRef] [PubMed]
- Gowdhami, B.; Jaabir, M.; Archunan, G.; Suganthy, N. Anticancer potential of zinc oxide nanoparticles against cervical carcinoma cells synthesized via biogenic route using aqueous extract of Gracilaria edulis. Mater. Sci. Eng. C 2019, 103, 109840. [Google Scholar] [CrossRef]
- Chen, H.; Luo, L.; Fan, S.; Xiong, Y.; Ling, Y.; Peng, S. Zinc oxide nanoparticles synthesized from Aspergillus terreus induces oxidative stress-mediated apoptosis through modulating apoptotic proteins in human cervical cancer HeLa cells. J. Pharm. Pharmacol. 2021, 73, 221–232. [Google Scholar] [CrossRef]
- Dulta, K.; Koşarsoy Ağçeli, G.; Chauhan, P.; Jasrotia, R.; Chauhan, P.K. A Novel Approach of Synthesis Zinc Oxide Nanoparticles by Bergenia ciliata Rhizome Extract: Antibacterial and Anticancer Potential. J. Inorg. Organomet. Polym. Mater. 2021, 31, 180–190. [Google Scholar] [CrossRef]
- Bai, D.P.; Zhang, X.F.; Zhang, G.L.; Huang, Y.F.; Gurunathan, S. Zinc oxide nanoparticles induce apoptosis and autophagy in human ovarian cancer cells. Int. J. Nanomed. 2017, 12, 6521–6535. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Xia, H.; Liang, W.; Huo, X.; Wei, X. Synthesis and characterization of zinc oxide nanoparticles from Morus nigra and its anticancer activity of AGS gastric cancer cells. J. Photochem. Photobiol. B Biol. 2020, 202, 111698. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, X.; Qiu, L.; Li, Y.; Marraiki, N.; Elgorban, A.M.; Xue, L. Green synthesized zinc oxide nanoparticles regulates the apoptotic expression in bone cancer cells MG-63 cells. J. Photochem. Photobiol. B Biol. 2020, 202, 111644. [Google Scholar] [CrossRef] [PubMed]
- Patrón-Romero, L.; Luque, P.A.; Soto-Robles, C.A.; Nava, O.; Vilchis-Nestor, A.R.; Barajas-Carrillo, V.W.; Martínez-Ramírez, C.E.; Chávez Méndez, J.R.; Alvelais Palacios, J.A.; Leal Ávila, M.; et al. Synthesis, characterization and cytotoxicity of zinc oxide nanoparticles by green synthesis method. J. Drug Deliv. Sci. Technol. 2020, 60, 101925. [Google Scholar] [CrossRef]
- Aude-Garcia, C.; Dalzon, B.; Ravanat, J.L.; Collin-Faure, V.; Diemer, H.; Strub, J.M.; Cianferani, S.; Van Dorsselaer, A.; Carrière, M.; Rabilloud, T. A combined proteomic and targeted analysis unravels new toxic mechanisms for zinc oxide nanoparticles in macrophages. J. Proteomics 2016, 134, 174–185. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, G.; Luo, X.; Zhang, P.; Peng, S.; Cheng, X.; Wang, M.; Yang, X. Endoplasmic reticulum stress-related calcium imbalance plays an important role on Zinc oxide nanoparticles-induced failure of neural tube closure during embryogenesis. Environ. Int. 2021, 152, 106495. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Song, L.; Yang, X.; Huang, Z.; Mou, X.; Syed, A.; Bahkali, A.H.; Zheng, L. Anticancer and genotoxicity effect of (Clausena lansium (Lour.) Skeels) Peel ZnONPs on neuroblastoma (SH-SY5Y) cells through the modulation of autophagy mechanism. J. Photochem. Photobiol. B Biol. 2020, 203, 111748. [Google Scholar] [CrossRef]
- Han, Z.; Yan, Q.; Ge, W.; Liu, Z.G.; Gurunathan, S.; De Felici, M.; Shen, W.; Zhang, X.F. Cytotoxic effects of ZnO nanoparticles on mouse testicular cells. Int. J. Nanomed. 2016, 11, 5187–5203. [Google Scholar] [CrossRef] [Green Version]
- Saber, M.; Hayaei-Tehrani, R.S.; Mokhtari, S.; Hoorzad, P.; Esfandiari, F. In vitro cytotoxicity of zinc oxide nanoparticles in mouse ovarian germ cells. Toxicol. Vitr. 2021, 70, 105032. [Google Scholar] [CrossRef]
- Sruthi, S.; Nury, T.; Millot, N.; Lizard, G. Evidence of a non-apoptotic mode of cell death in microglial BV-2 cells exposed to different concentrations of zinc oxide nanoparticles. Environ. Sci. Pollut. Res. 2021, 28, 12500–12520. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Ladokhin, A.S. Regulation of Apoptosis by the Bcl-2 Family of Proteins: Field on a Brink. Cells 2020, 9, 2121. [Google Scholar] [CrossRef] [PubMed]
- Morrish, E.; Brumatti, G.; Silke, J. Future Therapeutic Directions for Smac-Mimetics. Cells 2020, 9, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef]
- Wanner, E.; Thoppil, H.; Riabowol, K. Senescence and Apoptosis: Architects of Mammalian Development. Front. Cell Dev. Biol. 2021, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Rajashekara, S.; Shrivastava, A.; Sumhitha, S.; Kumari, S. Biomedical Applications of Biogenic Zinc Oxide Nanoparticles Manufactured from Leaf Extracts of Calotropis gigantea (L.) Dryand. Bionanoscience 2020, 10, 654–671. [Google Scholar] [CrossRef]
- Shobha, N.; Nanda, N.; Giresha, A.S.; Manjappa, P.; Sophiya, P.; Dharmappa, K.K.; Nagabhushana, B.M. Synthesis and characterization of Zinc oxide nanoparticles utilizing seed source of Ricinus communis and study of its antioxidant, antifungal and anticancer activity. Mater. Sci. Eng. C 2019, 97, 842–850. [Google Scholar] [CrossRef]
- Moratin, H.; Scherzad, A.; Gehrke, T.; Ickrath, P.; Radeloff, K.; Kleinsasser, N.; Hackenberg, S. Toxicological characterization of ZnO nanoparticles in malignant and non-malignant cells. Environ. Mol. Mutagen. 2018, 59, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Julien, O.; Wells, J.A. Caspases and their substrates. Cell Death Differ. 2017, 24, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Boice, A.; Bouchier-Hayes, L. Targeting apoptotic caspases in cancer. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118688. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.I.; Kuranaga, E. Caspase-dependent non-apoptotic processes in development. Cell Death Differ. 2017, 24, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- McComb, S.; Chan, P.K.; Guinot, A.; Hartmannsdottir, H.; Jenni, S.; Dobay, M.P.; Bourquin, J.P.; Bornhauser, B.C. Efficient apoptosis requires feedback amplification of upstream apoptotic signals by effector caspase-3 or -7. Sci. Adv. 2019, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kopeina, G.S.; Prokhorova, E.A.; Lavrik, I.N.; Zhivotovsky, B. Alterations in the nucleocytoplasmic transport in apoptosis: Caspases lead the way. Cell Prolif. 2018, 51, e12467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordo, R.; Nasrallah, G.K.; Al-Jamal, O.; Paliogiannis, P.; Pintus, G. Resveratrol inhibits oxidative stress and prevents mitochondrial damage induced by zinc oxide nanoparticles in zebrafish (Danio rerio). Int. J. Mol. Sci. 2020, 21, 3838. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, L. Aged Zinc Oxide Nanoparticles Did Not Induce Cytotoxicity Through Apoptosis Signaling Pathway as Fresh NPs. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in Cell Death, Inflammation, and Disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef]
- Suski, J.; Lebiedzinska, M.; Bonora, M.; Pinton, P.; Duszynski, J.; Wieckowski, M.R. Relation between mitochondrial membrane potential and ROS formation. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2018; Volume 1782, pp. 357–381. [Google Scholar]


{kind=link}
| Cell Type | Size and Shape of ZnO-NP | Synthesis and Characterization of ZnO-NPs | Time of Exposition (h) | Doses mg/L | IC50 | Biological Characterization | Mitochondrial Pathway | Reference |
|---|---|---|---|---|---|---|---|---|
| Human embryonic kidney cells (HEK293T) Chicken embryo (cranial neural crest cells HH10) | <50 nm Spherical shape | Synthesis method ND/OC. TEM, SEM-EDS, DLS | 12 h | 12.5, 25, 50, 100, 200 μg/mL | 50 μg/mL ND * | Cell culture, CCK-8, CNCCs, ARS, IHC, RNA-seq, q-PCR, DAPI, FM/IFA | Ion release triggered ROS production, which further induced cell toxicity, inflammation and apoptosis, which are mediated by NF-κB signaling cascades and mitochondria dysfunction. Increase in the expression of Nrf2, HO-1, NQO1, Cat, GLXR and NOS. Increase in Pax7 and Casp-3 expression. | [8] |
| Human cardiomyocytes (hiPSC-CMs) | 40 nm–60 nm Spheroid and rod shaped | Synthesis method ND/OC. TEM, DLS | 0, 2, 6 h | 0–200 μg/mL | 62.5 μg/mL | Cell culture, LDH, HCA, CCK-8, MMP, ROS, q-PCR, WB, MEA | ROS generation and induced mitochondrial dysfunction. Impair mitochondrial biogenesis and inhibit the PGC-1α pathway. Cardiac electrophysiological alterations. | [22] |
| Human epidermoid carcinoma cell A431 Normal kidney epithelial NRK-52E cells | <40 nm Hexagonal structure | Chemical synthesis by reduction Zinc acetate dyhidrate XRD, TEM, SEM, UV–Vis | 24 h 24 h | 0–25 μg/mL | 24 μg/mL 15 μg/mL NS * | Cell culture, MTT, FM, ROS, DAPI, Casp-3 and cell morphology | The anti-proliferative activity, morphological changes, ROS generation, nuclear apoptosis and caspase-3 in a dose-dependent manner. | [32] |
| Human aortic endothelial cells (HAECs) | 70 nm Rod shaped | Synthesis method ND/OC. TEM, XRD | 12 or 24 h | 8–50 μg/mL | 50 μg/mL | Cell culture, MTT, LDH, ROS, FM, MMP, LA, CACAk, Cas-3, Cas-9, Cyt-c, ICC, IFA | Apoptosis was confirmed using reactive oxygen species (ROS). Decrease in MMP. Increased release of Cyt-c. Caspase-3, caspase-9, BAX, BCL-2 and FAS receptor expression. The antioxidant LA was able to protect HAECs from apoptosis induced by ZnO-NPs. | [33] |
| Human tenon fibroblast (HTF) | 56 nm ND | Synthesis method ND/OC. TEM | 24 h 48 h 72 h | 0–16.0 μg/mL | 1.51 μg/mL 1.03 μg/mL 0.57 μg/mL | Cell culture, CCK-8, RT-CES, ROS, FM, MMP, qPCR, FC, Apaf-1, Cas-3, Cas-9, FSP-1 | Inhibit the viability of HTFs and decrease MMP. Elevated ROS, caspase-3, caspase-9, and apoptotic Apaf-1 expression. Decrease the levels of FSP-1, collagen III, and E-cadherin expression, leading to HTF apoptosis. | [34] |
| Human erythrocytes | 47.8–52.5 nm Rod shaped | Chemical synthesis by Pechini method XRD, TEM, FTIR | 1 h | 50–500 μg/mL | 200 μg/mL | Cell culture, H, SOD, CAT, LPO, GST, GSH, ROS, OH, O2, CA, IFA/FM | Concentration-dependent hemolytic activity to human erythrocytes. ROS generation. Depletion of glutathione and GST levels. Increased SOD, CAT and lipid peroxidation in dose-dependent manner. | [35] |
| Human monocytes U-937 Human promyelocytes HL-60 Human B lymphocytes COLO-720L Human T lymphocytes HUT-78 | 15 nm ND | Synthesis method ND/OC. TEM, DLS | 24 h | 1.6–25 μg/mL | 1.5625 μg/mL 3.125 μg/mL 6.25 μg/mL 12.5 μg/mL 25 μg/mL | Cell culture, MTT, LDH, MDA | ZnO-NPs caused lipid peroxidation of all cells and correlated with apoptosis. The level of cholesterol in membranes strongly modifies the effect exerted by ZnO-NPs. | [36] |
| Human monocytes U-937 Human promyelocytes HL-60 Human B lymphocytes COLO-720L Human T lymphocytes HUT-78 | 100–130 nm ND | Synthesis method ND/OC. TEM | 24 h | 0–25 μg/mL | 1.56 μg/mL 3.12 μg/mL 6.25 μg/mL 12.5 μg/mL 25 μg/mL | Cell culture, MTT, LDH, NO, MDA, Casp-9, IL-6, TNF-α | Activation of the mitochondrial apoptosis pathway. TNF-α concentration increased for both cell lines. IL-6 concentration rose on average 5-fold in HL-60 cells in all experimental variants, the opposite trend was observed for COLO-720L. ZnO-NPs are cytotoxic to immune system and cause peroxidation od membrane lipids. | [37] |
| Human colon carcinoma (HCT116) Human myelogenous leukemic (K562) | 30–48.5 nm Hexagonal structure | Green Synthesis method (Spondias pinnata) UV–Vis, FTIR, XRD, FESEM, HRTEM, EDX | 24 and 48 h | 0.25–200 μg/mL | 82 and 60μg/mL 55 and 35μg/mL | Cell culture, MTT, IFA/FM, VWH, MMP, FC, CA, DNAf, RT-qPCR, WB, H | Upregulation of pro-apoptotic (PUMA, Bax, Cyt-c, cas-9, cas-3, and PARP) and downregulation of anti-apoptotic (Bcl-2 and survivin) genes. Elevation in the expression of Cyt-c, cas-9, and cas-3 genes involved in mitochondrial (intrinsic) apoptotic pathway. | [38] |
| Human keratinocytes (HaCaT) Human gingival fibroblast (HGF-1) Human gingival carcinomas (Ca9-22) Human gingival carcinomas (OECM-1) | 100 nm ND | Synthesis method ND/OC. SEM | 24 h | 0–100 μg/mL | 100 μg/mL 100 μg/mL 17.4 ± 0.6 μg/mL 51.0 ± 0.6 μg/mL | Cell culture, MTT, ROS, MMP, WB, ZXA, FC | Mitochondrial oxidative damage and p70S6K signaling pathway inhibition, caspase-3, caspase-8, caspase-9 and PARP activation. Induce sub-G1 arrest of the cell cycle followed by apoptosis in human GSCC. ROS is essential for the anti-cancer activity. Did not affect the expression of pro-survival Bcl-2 members (Bcl-2, Bcl-xl, and Mcl-1) as well as pro-apoptosis Bcl-2 members (Bax, Bad, and Bid). | [39] |
| Human oral caner (CAL 27) | 50 nm Hexagonal prism | Synthesis method ND/OC. TEM, XRD and DLS | 24 h | 0–100 μg/mL | 25 μg/mL | Cell culture, CCK-8, LC3, P62, GAPDH, PINK1, Parkin, IHC, ROS, MDC, MMP, WB, JC-I, ROS | Increased ROS levels, decreased MMP and mitochondrial dysfunction time-dependent manner. Increased levels of LC3-II, PINK1, mito-Parkin and decreased P62 and Parkin. | [40] |
| Human hepatocytes (HepG2) | 237 nm 79 nm Rod shaped and spherical | Synthesis method ND/OC. TEM, ICP-AES, EDX and DLS | 24 h | 0–300 μg/mL | >150 μg/mL | Cell culture, qRT-PCR, AVO, MMP, TBEA, SOD, CAT | Increased mitochondrial zinc transporter 1 and 2 expression levels. Response and the upregulation of MET, ZnT1, ZnT2, no change in CAT, SOD1 and SOD2 expression, nor in the enzymatic activities of catalase and SOD, implying a minimal activation of oxidative stress. Abnormal mitochondria morphologies and autophagy vesicles in response to ZnO-NPs. | [30] |
| Human hepatocytes (HL-7702) Human colorectal (Caco-2) | <100 nm ND | Synthesis method ND/OC. ICP-MS, TEM, SEM | 3 h | 0.665 μg/mL | NS * NS * | Cell culture, MTT, LDH, MMP, ROS, qRT-PCR | Increased ROS levels due to NP exposure, overexpression of HMOX1 in response to the increased oxidative stress. The cytotoxicity induced by high ROS levels, oxidative stress and depolarization of mitochondrial membrane. | [41] |
| Huh7 hepatocytes cancer cell line | 18 nm Polycrystalline nature | Green synthesis (Luff acutangula) UV–Vis, HRTEM, SAED | 8 h | 0–60 μg/mL | 40 µg/mL | Cell culture, MTT, ROS, MMP, DNAf. FM | Huh 7 liver cancer cells undergo apoptosis as a result of generation of ROS molecules. Stimulation of apoptotic signaling pathway in a dose-dependent manner. Condensed chromatin and fragmented DNA. | [42] |
| Human hepatocytes (HepG2) Human breast cancer (MCF-7) | ∼13 ± 2 nm ND | Chemical synthesis by reduction Zinc acetate dihydrate FESEM, TEM, XRD, FTIR, UV–Vis | 24 and 48 h | 0–100 μg/mL | 25 and 10 µg/mL 25 µg/mL | FC, MTT, FACS, qRT-PCR, cell morphology, Casp-3, p53,Bax,Bcl-2 | Significant upregulation of mRNA expression levels of Bax, p53, and caspase-3 and the downregulation of the anti-apoptotic gene Bcl-2. | [43] |
| Human breast cancer (MCF-7) | 41 nm flakes Hexagonal | Chemical synthesis Triethanolamine XRD, FESEM, EDS | 24 h | 1–5 μg/mL | 3 μg/mL | Cell culture, MTT, DPPH, AO/EB, PI | Dose-dependent loss of cell viability. ZnO-NP exposure increases necrosis and apoptosis of MCF cells. | [44] |
| Human breast cancer (MCF-7) | 43 nm Spherical | Synthesis using sol-gel method. XRD, HR-TEM, EDS, TEM, XRD | 24 h | 0–100 μg/mL | 44 μg/mL | Cell culture, MTT, LDH, ROS, GSH, TSH, MMP, IFA/FM, q-PCR | Upregulation of apoptotic genes (p53, Bax/Bcl2 ratio, caspase-3 and caspase-9). Loss of MMP and apoptosis in MCF-7 cells through the mitochondrial pathway. | [45] |
| Human breast cancer (MCF-7) Human Non-tumorigenic human mammary epithelial cell line (MCF-10A) | 63.7 ±6.5 nm Spherical | Synthesis method ND/OC. DLS, AFM | 24 and 48 h | 0.5–20 μg/mL | 20 and 10 µg/mL >90% IC50 with max doses | MTT, DAPI, qRT-PCR, IFA/FM and cell morphology | The increased expression of Bax/Bcl-2 ratio confirmed the induction of apoptosis in MCF-7 cells. ZnO-NPs induced casp3- and casp-8 upregulation. | [46] |
| Human multiple myeloma cell (RPMI8226) Peripheral blood mononuclear cell | 30 nm | Synthesis method ND/OC. TEM | 24 and 48 h | 0–60 μg/mL | 34 and 29 µg/mL 120 and79 µg/mL | ROS, FC, q-PCR, ATP assay, cell morphology | Increase expression and proteins levels of caspase-3, caspase-9, Apaf-1, and Cyt-c, decrease ATP levels in time- and dose-dependent manner, ROS generation. | [47] |
| Human cervical cancer cell (SiHa) | 20–50 nm | Green synthesis (Gracilaria edulis) UV–Vis, FTIR, XRD, FESEM, XPS, SEM, EDS, EDX and HRTEM | 24 h | 0–200 μg/mL | 35 µg/mL | MTT, FC, ROS, MMP AO/EB, IFA/FM, CA, JC-I | Apoptotic and necrotic effect. ROS elevation, DNA damage, activation of mitochondrial intrinsic pathway. | [48] |
| Human cervical cancer cell line (HeLa) | 10–70 nm | Green synthesis (Aspergillus terreus) UV–Vis, XRD, EDX, FTIR and TEM. | 24 h | 5–80 μg/mL | 20 µg/mL | MTT, WB, SOD, CAT, ROS, MMP, p53, BAX, casp-3, casp-9, Cyt-c. Cell morphology. | Reduced SOD, CAT and GPO, increased ROS, diminished MMP. Upregulation of p53, casp-9, BAX, RAD51 and downregulation of BCL-2. | [49] |
| Human cervical cancer (HeLa) Human colon cancer (HT-29) | 30 nm | Green synthesis (Bergenia ciliate) UV–Vis, FTIR, DLS, EDX, XDR and SEM | 24 h | 25–200 μg/mL | 101.7 μg/mL 124.3 μg/mL | MTT, DPPH and ABTS assay | Increased ROS and cytotoxicity against cancer cells. | [50] |
| Human ovarian cancer (SKOV3) | 20 nm | Synthesis method ND/OC. UV–Vis, TEM, XRD, FTIR and AFM | 24 h | 0–30 μg/mL | 20–30 μg/mL | Cell morphology, MTT, ICC, JC-I assay, TUNEL, WB, ROS levels and IFA/FM | Genotoxicity, double DNA strand breaks and apoptosis. Loss of MMP, ROS increased levels. Upregulation of p53, casp-9, BAX, RAD51 and downregulation of BCL-2. | [51] |
| AGS gastric cancer cells | 100 nm | Green synthesis (Morus nigra) UV–Vis, TEM, SEM, FT-IR, EDX and XDR | 24 h | 0–25 μg/mL | 10 µg/mL | MTT, MMP, AO/EB, ROS, FC, CAT, TBARS, GSH SOD, RT-PCR | MMP decreased. Apoptosis induced by ROS generation and gene expressions of apoptosis markers effects, increased lipid peroxidation, cell cycle arrest. | [52] |
| Human osteosarcoma cell line (MG-63) | 10–12 nm | Green synthesis (Radix Rehmanniae) UV–Vis, TEM, XRD and FTIR | 24 h | 5–80μg/mL | 30 µg/mL | MTT, MMP, WB. ROS, BAX, casp-3 and casp-9 levels. Morphology assay AO/EB | Increased levels of BAX, casp-3, casp-9. MMP decreased and ROS generation. | [53] |
| Human lymphocytes Mouse fibroblast cell (C2C12) Mouse myoblast cell (L929) | 16–24 nm | Green synthesis (Beta vulgaris) HRTEM, FTIR, XRD | 24 and 48 h | 1 × 10−6 100 μg/mL | 37 and 30 μg/mL 32 and 23 μg/mL 28 and 20 μg/mL | MTT, TBEA, cell kinetic and morphology assay | Abnormal cell morphology, adherence and viability manner. Mitochondrial function. Increased doses-dependent mortality rate. | [54] |
| Mouse macrophage line (J774A1) | 140 ± 16 nm | Synthesis method ND/OC. TEM, SEM DLS | 24 h | 200 µg/mL | 8 µg/mL | IFA/FM. MMP, GAPDH assay, mass spectrometry, HPLC, 2D and SDS electrophoresis. | Mitochondrial proteomics abnormalities, MMP proteins loss, nuclear DNA damage, genotoxic effect. GAPDH inhibition, increased levels od methylglyoxal-associated DNA damage. | [55] |
| Chicken embryo HHO Neural stem cells Neuroblastoma SH-SY5Y cell | 50 m | Synthesis method ND/OC. TEM, SEM, EDS and DLS | 24 h 12 h 12 h | 12.5–50 µg/mL 0–50 µg/mL 0–50 µg/mL | ND * 25 µg/mL 25 µg/mL | Cell/embryo IFA, FC, WB, RNA-seq, q-PCR, IFA/FM | Endoplasmic reticulum stress, increased Ca2+ levels, Casp-3, increased levels, abnormal mitochondrial morphology. ZnO-NPs induced failure of neural tube closure. | [56] |
| Neuroblastoma SH-SY5Y cell | 24–30 nm | Green synthesis (Clausena lansium) UV–Vis, XRD, FT-IR and TEM | 24 h | 10–20 µg/mL | 15 µg/mL | Cell viability, MTT, ROS levels, WB, CA (DNA damage) | Increased ROS levels, increased levels of BAX, caspase-3, and BCL-2 proteins. Autophagy (beclin-1, LC3-I, LCEII and ATG4B) increased levels. DNA loss and damage. NAC prevents ROS in SH-SY5Y. | [57] |
| Mouse Leydig cells Mouse Sertoli cells | 70 nm | Synthesis method ND/OC. TEM, AFM, XRD, FTIR, UV–Vis, DLS | 24 h 24 h | 0, 5, 10, 15, and 20 μg/mL | 15 μg/mL 15 μg/mL | MTT, LDH, ROS levels, TUNEL and JC-I assay, WB, sperm morphology | Increased ROS levels, MMP abnormalities, increased apoptotic proteins, loss of MMP, nuclear DNA damage and breakage. | [58] |
| Mouse ovarian germ cells (CHO-K1) | 80 nm | Co-precipitation method SEM, TEM | 24 h (1 day) 168 h (7day) | 10, 20 and 30 μg/mL | 30 μg/mL 10 μg/mL | IFA/FM; FC. Quantification of ROS. qRT-PCR of Prdm1, Dppa3, Ifitm3, Ddx4 and Dazl | ROS level, cell membrane integrity. Significant increase in expression of premeiotic germ cells markers but a decrease in meiotic and post-meiotic markers. | [59] |
| Murine microglial BV-2 cells | 20 nm | Synthesised by using the wet chemical method. TEM, SEM, XRD, XPS | 6 h 24 h | 0–80 μg/mL | 20 μg/mL 10 μg/mL | MTT, TBEA, FC. MMP, ROS levels, PMP levels, cell cycle analysis, WB | Absence of caspase-3 cleavage and PARP fragmentation. Mitochondrial dysfunction and lysosomal alteration increased PMP. Accumulation of ROS. Non-apoptotic hallmarks, necrotic markers in BV-2 cells. | [60] |
| S. Cerevisiae (yeast BY4743) | 20–80 nm | Synthesis method ND/OC. SEM, HRFESEM, UV–Vis and DLS analysis. | 3 h | 5–20 μg/mL | 10 μg/mL | Cell death assay. WB, ROS levels, chitin and lipid droplets measurement. Vacuolar organelle morphology. RT-PCR | Mitochondrial morphology and lipid homeostasis abnormalities, ROS elevation. Perturbations in peripheral endoplasmic reticulum. ZnO-NPs do not affect histone epigenetic marks. Increased levels of Hog1. Inhibition of cell growth due alterations in CWI and HOG signaling pathway. | [27] |
| S. Cerevisiae (yeast BY4741) | 20–80 nm | Synthesis method ND/OC. SEM, HRFESEM, UV–Vis | 3 h | 10 μg/mL | 10 μg/mL | Cell culture, WB, HNMR, RT-PCR. RNA and metabolites identification/quantification | Mitochondrial expression of PSD1, SOD1, SOD2, KGD1 TCA cycle genes (ACO1 and KDG1) Oxidative pathway abnormalities (SOD1, SOD2 and YAP1) Lipid biosynthesis pathway abnormalities (INO2, INO4, CHO1, PSD1 and CRD1) | [28] |
| Mitochondrial Pathway | ZnO-NPs Effect |
|---|---|
| Mitochondrial biogenesis | ↑ mDNA replication and transcription systems. |
| ↑ PSD1 to mitochondrial maintenance. | |
| ↑ Mitochondrial biogenesis by PGC-1. | |
| ↑ Mitophagy. | |
| Apoptosis induction | ↑ p53, BAX, BAK, SMAC and cytochrome-c levels to induce apoptosis. |
| ↑ DNA fragmentation and cytoplasmic reduction. | |
| ↑ Anti-BCL2. | |
| Caspases induction | ↑ Pro-caspases-9 and pro-caspase-3 to convert in activated caspase-9. |
| ↑ Apaf-1. | |
| ↑ Apoptosome. | |
| Mitochondrial membrane potential | ↑ Mitochondrial membrane potential. |
| ↑ ATP synthase. | |
| ↑ Reactive oxygen species. | |
| ↑ Mitochondrial pore abnormalities with delivery of pro-apoptotic proteins. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patrón-Romero, L.; Luque-Morales, P.A.; Loera-Castañeda, V.; Lares-Asseff, I.; Leal-Ávila, M.Á.; Alvelais-Palacios, J.A.; Plasencia-López, I.; Almanza-Reyes, H. Mitochondrial Dysfunction Induced by Zinc Oxide Nanoparticles. Crystals 2022, 12, 1089. https://doi.org/10.3390/cryst12081089
Patrón-Romero L, Luque-Morales PA, Loera-Castañeda V, Lares-Asseff I, Leal-Ávila MÁ, Alvelais-Palacios JA, Plasencia-López I, Almanza-Reyes H. Mitochondrial Dysfunction Induced by Zinc Oxide Nanoparticles. Crystals. 2022; 12(8):1089. https://doi.org/10.3390/cryst12081089
Chicago/Turabian StylePatrón-Romero, Leslie, Priscy Alfredo Luque-Morales, Verónica Loera-Castañeda, Ismael Lares-Asseff, María Ángeles Leal-Ávila, Jorge Arturo Alvelais-Palacios, Ismael Plasencia-López, and Horacio Almanza-Reyes. 2022. "Mitochondrial Dysfunction Induced by Zinc Oxide Nanoparticles" Crystals 12, no. 8: 1089. https://doi.org/10.3390/cryst12081089
APA StylePatrón-Romero, L., Luque-Morales, P. A., Loera-Castañeda, V., Lares-Asseff, I., Leal-Ávila, M. Á., Alvelais-Palacios, J. A., Plasencia-López, I., & Almanza-Reyes, H. (2022). Mitochondrial Dysfunction Induced by Zinc Oxide Nanoparticles. Crystals, 12(8), 1089. https://doi.org/10.3390/cryst12081089