
Soft Polymer-Based Technique for Cellular Force Sensing


Abstract
:1. Introduction
2. Soft Polymer-Based Cellular Force Sensing Technique
2.1. Synthetic Soft Polymer-Based Techniques
2.1.1. Deformable Membrane (DM)
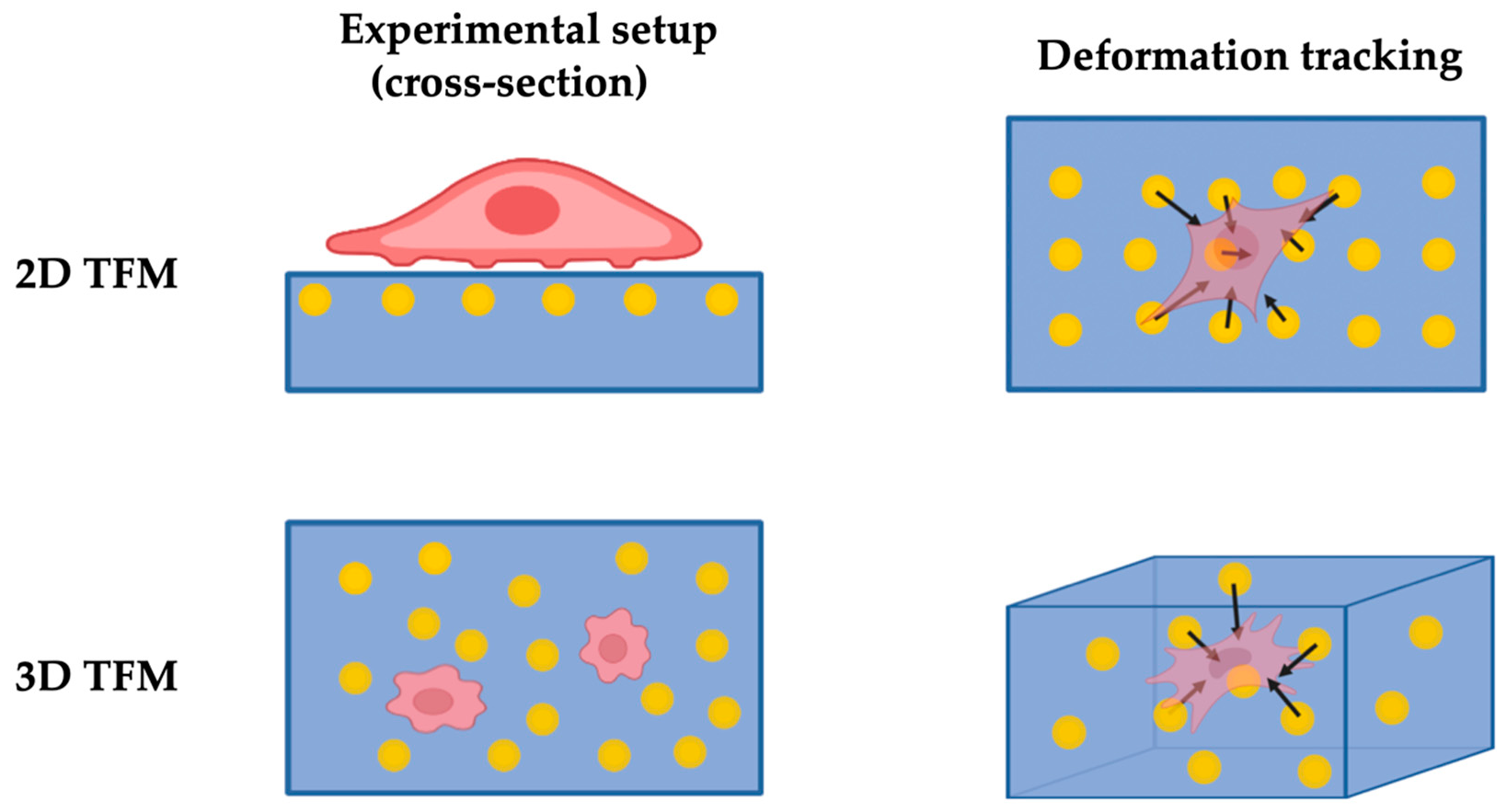
2.1.2. Traction Force Microscopy (TFM)
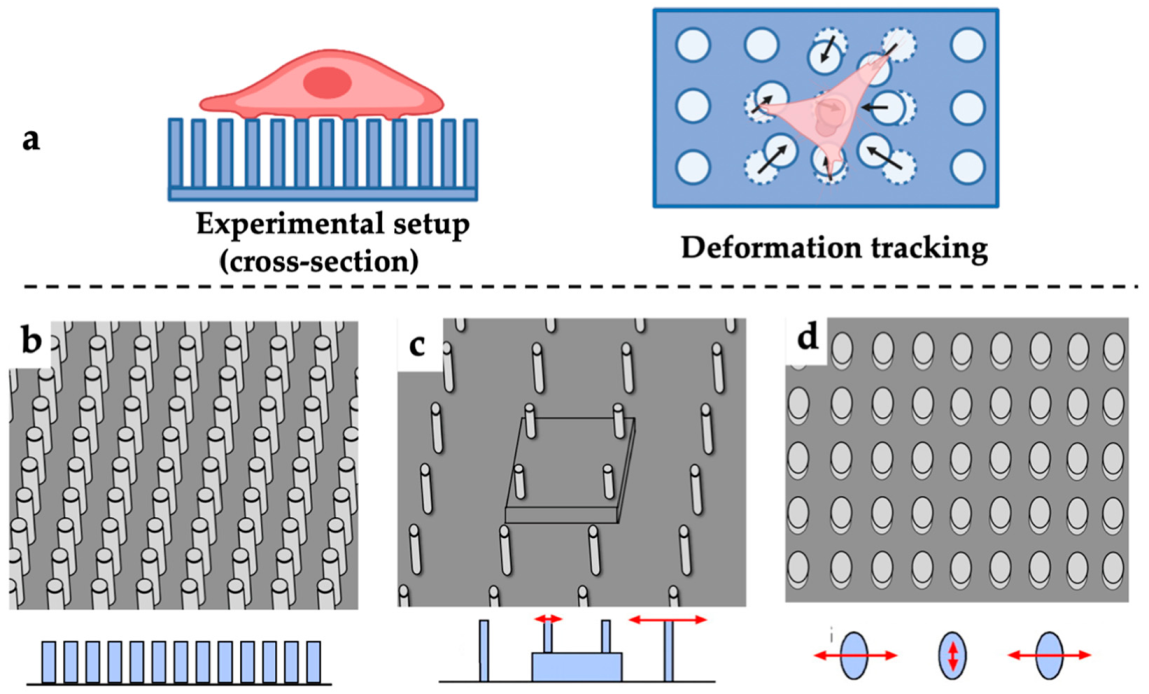
2.1.3. Elastic Micro-Pillar Technique (EMP)
2.2. Natural Soft Polymer-Based Techniques
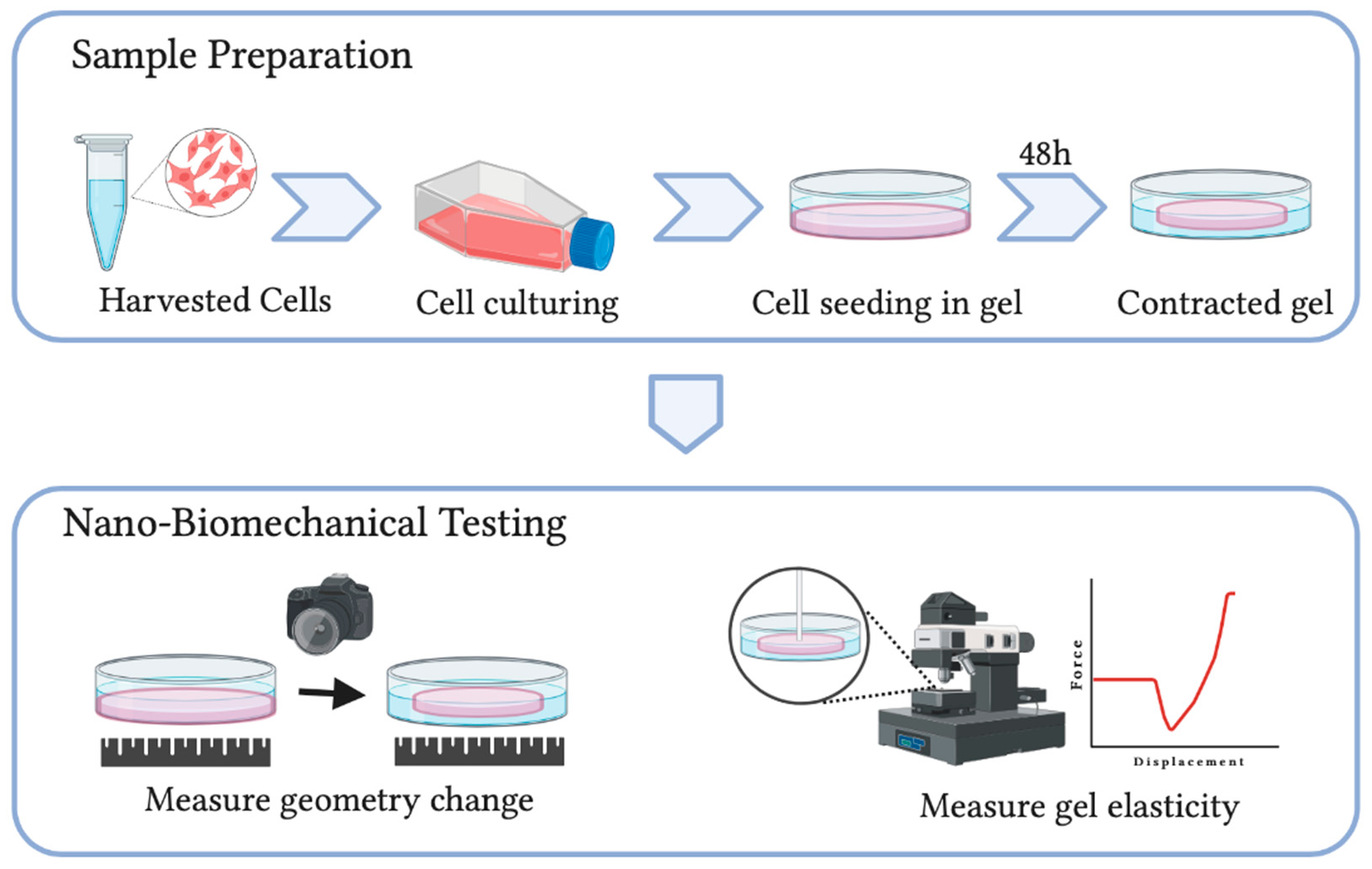
2.2.1. Collagen Gel-Based Contraction Assay (CGCA)
2.2.2. Culture Force Monitor (CFM)
3. Dynamic Interactions between Cells and Soft Polymer-Based Matrix
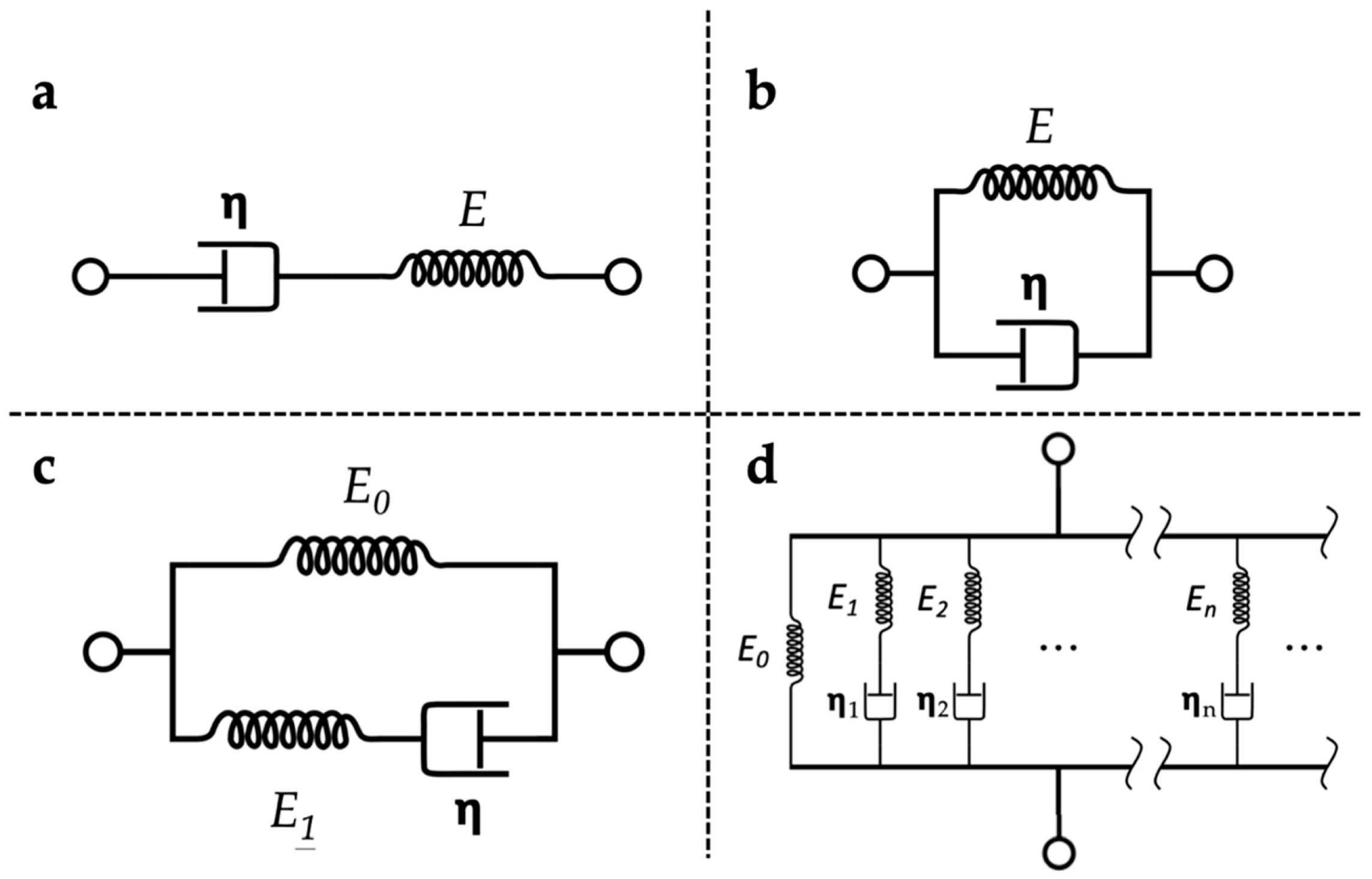
4. Viscoelastic Properties of the Soft Polymer Matrix
5. Intricate Problems in Cellular Force Sensing
5.1. 2D Substrate or 3D Matrix
5.2. Measurement Accuracy
6. Prospects
6.1. Extended Choice of Natural Materials
6.2. Bioactive Modification of Synthetic Materials
6.3. 3D Printing of the Soft Polymer Scaffold
6.4. Improvements of Methodological Design on Soft Polymers
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, C.H.; Singla, A.; Lee, Y. Biomedical applications of collagen. Int. J. Pharm. 2001, 221, 1–22. [Google Scholar] [CrossRef]
- Behring, J.; Junker, R.; Walboomers, X.F.; Chessnut, B.; Jansen, J.A. Toward guided tissue and bone regeneration: Morphology, attachment, proliferation, and migration of cells cultured on collagen barrier membranes. A systematic review. Odontology 2008, 96, 1–11. [Google Scholar] [CrossRef]
- Mondschein, R.J.; Kanitkar, A.; Williams, C.B.; Verbridge, S.S.; Long, T.E. Polymer structure-property requirements for stereolithographic 3D printing of soft tissue engineering scaffolds. Biomaterials 2017, 140, 170–188. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.-N.; Abdollahi, S.; Laurenti, M.; Fang, D.; Tran, S.D.; Cerruti, M.; Tamimi, F. Scaffolds for epithelial tissue engineering customized in elastomeric molds. J. Biomed. Mater. Res. Part B Appl. Biomater. 2018, 106, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, J.M.; Hollister, S.J. Tailoring the mechanical properties of 3D-designed poly(glycerol sebacate) scaffolds for cartilage applications. J. Biomed. Mater. Res. Part A 2010, 94A, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Huang, C.; Tomsia, A.P. Freeze Casting for Assembling Bioinspired Structural Materials. Adv. Mater. 2017, 29, 1703155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Hong, Z.; Yu, T.; Chen, X.; Jing, X. In vivo mineralization and osteogenesis of nanocomposite scaffold of poly(lactide-co-glycolide) and hydroxyapatite surface-grafted with poly(l-lactide). Biomaterials 2009, 30, 58–70. [Google Scholar] [CrossRef]
- Lee, K.Y.; Jeong, L.; Kang, Y.O.; Lee, S.J.; Park, W.H. Electrospinning of polysaccharides for regenerative medicine. Adv. Drug Deliv. Rev. 2009, 61, 1020–1032. [Google Scholar] [CrossRef]
- Visser, C.W.; Amato, D.N.; Mueller, J.; Lewis, J.A. Architected Polymer Foams via Direct Bubble Writing. Adv. Mater. 2019, 31, 1904668. [Google Scholar] [CrossRef]
- Bajaj, P.; Schweller, R.M.; Khademhosseini, A.; West, J.L.; Bashir, R. 3D biofabrication strategies for tissue engineering and regenerative medicine. Annu. Rev. Biomed. Eng. 2014, 16, 247–276. [Google Scholar] [CrossRef] [Green Version]
- Martinez, P.R.; Goyanes, A.; Basit, A.W.; Gaisford, S. Fabrication of drug-loaded hydrogels with stereolithographic 3D printing. Int. J. Pharm. 2017, 532, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.R.; Zhang, X. Microsystems for cellular force measurement: A review. J. Micromech. Microeng. 2011, 21, 54003. [Google Scholar] [CrossRef]
- Polacheck, W.J.; Chen, C.S. Measuring cell-generated forces: A guide to the available tools. Nat. Methods 2016, 13, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siamantouras, E.; Hills, C.E.; Squires, P.E.; Liu, K.K. Quantifying cellular mechanics and adhesion in renal tubular injury using single cell force spectroscopy. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1013–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Liu, Z.; Liu, H.; Nash, M.A. Next Generation Methods for Single-Molecule Force Spectroscopy on Polyproteins and Receptor-Ligand Complexes. Front. Mol. Biosci. 2020, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Harlepp, S.; Thalmann, F.; Follain, G.; Goetz, J.G. Hemodynamic forces can be accurately measured in vivo with optical tweezers. Mol. Biol. Cell 2017, 28, 3252–3260. [Google Scholar] [CrossRef]
- Arbore, C.; Perego, L.; Sergides, M.; Capitanio, M. Probing force in living cells with optical tweezers: From single-molecule mechanics to cell mechanotransduction. Biophys. Rev. 2019, 11, 765–782. [Google Scholar] [CrossRef]
- Smith, S.B.; Finzi, L.; Bustamante, C. Direct mechanical measurements of the elasticity of single DNA molecules by using magnetic beads. Science 1992, 258, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.; Ritchie, K.; Merkel, R. Sensitive force technique to probe molecular adhesion and structural linkages at biological interfaces. Biophys. J. 1995, 68, 2580–2587. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.K.; Wild, P.; Stopak, D. Silicone rubber substrata: A new wrinkle in the study of cell locomotion. Science 1980, 208, 177–179. [Google Scholar] [CrossRef]
- Harris, A.K.; Stopak, D.; Wild, P. Fibroblast traction as a mechanism for collagen morphogenesis. Nature 1981, 290, 249–251. [Google Scholar] [CrossRef]
- Pelham, R.J.; Wang, Y.L. Cell locomotion and focal adhesions are regulated by substrate flexibility. Proc. Natl. Acad. Sci. USA 1997, 94, 13661–13665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munevar, S.; Wang, Y.L.; Dembo, M. Traction force microscopy of migrating normal and H-ras transformed 3T3 fibroblasts. Biophys. J. 2001, 80, 1744–1757. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.L.; Tien, J.; Pirone, D.M.; Gray, D.S.; Bhadriraju, K.; Chen, C.S. Cells lying on a bed of microneedles: An approach to isolate mechanical force. Proc. Natl. Acad. Sci. USA 2003, 100, 1484–1489. [Google Scholar] [CrossRef] [Green Version]
- Vernon, R.B.; Gooden, M.D. An improved method for the collagen gel contraction assay. Vitr. Cell. Dev. Biol. Anim. 2002, 38, 97–101. [Google Scholar] [CrossRef]
- Moon, A.G.; Tranquillo, R.T. Fibroblast-populated collagen microsphere assay of cell traction force: Part 1. Continuum model. AIChE J. 1993, 39, 163–177. [Google Scholar] [CrossRef]
- Jin, T.; Li, L.; Siow, R.C.M.; Liu, K.-K. A novel collagen gel-based measurement technique for quantitation of cell contraction force. J. R. Soc. Interface 2015, 12, 20141365. [Google Scholar] [CrossRef]
- Yu, Z.; Smith, M.J.; Siow, R.C.M.; Liu, K.-K. Ageing modulates human dermal fibroblast contractility: Quantification using nano-biomechanical testing. Biochim. Biophys. Acta-Mol. Cell Res. 2021, 1868, 118972. [Google Scholar] [CrossRef]
- Delvoye, P.; Wiliquet, P.; Levêque, J.L.; Nusgens, B.V.; Lapière, C.M. Measurement of mechanical forces generated by skin fibroblasts embedded in a three-dimensional collagen gel. J. Investig. Dermatol. 1991, 97, 898–902. [Google Scholar] [CrossRef]
- Campbell, B.H.; Clark, W.W.; Wang, J.H.C. A multi-station culture force monitor system to study cellular contractility. J. Biomech. 2003, 36, 137–140. [Google Scholar] [CrossRef]
- Boys, A.J.; Owens, R.M. Measuring cellular contraction: Current progress and a future in bioelectronics. APL Mater. 2021, 9, 040903. [Google Scholar] [CrossRef]
- Rodriguez, M.L.; McGarry, P.J.; Sniadecki, N.J. Review on cell mechanics: Experimental and modeling approaches. Appl. Mech. Rev. 2013, 65, 060801. [Google Scholar] [CrossRef] [Green Version]
- Tibbitt, M.W.; Anseth, K.S. Hydrogels as extracellular matrix mimics for 3D cell culture. Biotechnol. Bioeng. 2009, 103, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, Y. Chapter Two-Single-Cell Imaging of Mechanotransduction in Endothelial Cells. In Mechanotransduction; Engler, A.J., Kumar, S., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 126, pp. 25–51. ISBN 1877-1173. [Google Scholar]
- Curtze, S.; Dembo, M.; Miron, M.; Jones, D.B. Dynamic changes in traction forces with DC electric field in osteoblast-like cells. J. Cell Sci. 2004, 117, 2721–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, A.D.; Lee, J. Cyclic changes in keratocyte speed and traction stress arise from Ca2+-dependent regulation of cell adhesiveness. J. Cell Sci. 2005, 118, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Ng, S.S.; Li, C.; Chan, V. Experimental and numerical determination of cellular traction force on polymeric hydrogels. Interface Focus 2011, 1, 777–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ng, S.S.; Wang, Y.; Feng, H.; Chen, W.N.; Chan-Park, M.B.; Li, C.; Chan, V. Collective cell traction force analysis on aligned smooth muscle cell sheet between three-dimensional microwalls. Interface Focus 2014, 4, 20130056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Botvinick, E.L.; Zhao, Y.; Berns, M.W.; Usami, S.; Tsien, R.Y.; Chien, S. Visualizing the mechanical activation of Src. Nature 2005, 434, 1040–1045. [Google Scholar] [CrossRef]
- Iwadate, Y.; Yumura, S. Actin-based propulsive forces and myosin-II-based contractile forces in migrating Dictyostelium cells. J. Cell Sci. 2008, 121, 1314–1324. [Google Scholar] [CrossRef] [Green Version]
- Beningo, K.A.; Dembo, M.; Kaverina, I.; Small, J.V.; Wang, Y.L. Nascent focal adhesions are responsible for the generation of strong propulsive forces in migrating fibroblasts. J. Cell Biol. 2001, 153, 881–888. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, M.L.; Knecht, D.A.; Dembo, M.; Lee, J. Traction force microscopy in Dictyostelium reveals distinct roles for myosin II motor and actin-crosslinking activity in polarized cell movement. J. Cell Sci. 2007, 120, 1624–1634. [Google Scholar] [CrossRef] [Green Version]
- Rösel, D.; Brábek, J.; Tolde, O.; Mierke, C.T.; Zitterbart, D.P.; Raupach, C.; Bicanová, K.; Kollmannsberger, P.; Paňková, D.; Veselý, P.; et al. Up-regulation of Rho/ROCK signaling in sarcoma cells drives invasion and increased generation of protrusive forces. Mol. Cancer Res. 2008, 6, 1410–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.B.; Dembo, M.; Wang, Y.L. Substrate flexibility regulates growth and apoptosis of normal but not transformed cells. Am. J. Physiol. Cell Physiol. 2000, 279, c1345–c1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakely, B.L.; Dumelin, C.E.; Trappmann, B.; McGregor, L.M.; Choi, C.K.; Anthony, P.C.; Duesterberg, V.K.; Baker, B.M.; Block, S.M.; Liu, D.R.; et al. A DNA-based molecular probe for optically reporting cellular traction forces. Nat. Methods 2014, 11, 1229–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimatsu, M.; Mekhdjian, A.H.; Adhikari, A.S.; Dunn, A.R. Molecular tension sensors report forces generated by single integrin molecules in living cells. Nano Lett. 2013, 13, 3985–3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurchenko, C.; Salaita, K.S. Lighting Up the Force: Investigating Mechanisms of Mechanotransduction Using Fluorescent Tension Probes. Mol. Cell. Biol. 2015, 35, 2570–2582. [Google Scholar] [CrossRef] [Green Version]
- Yasunaga, A.; Murad, Y.; Li, I.T.S. Quantifying molecular tension—Classifications, interpretations and limitations of force sensors. Phys. Biol. 2019, 17, 11001. [Google Scholar] [CrossRef] [PubMed]
- Cukierman, E.; Pankov, R.; Stevens, D.R.; Yamada, K.M. Taking cell-matrix adhesions to the third dimension. Science 2001, 294, 1708–1712. [Google Scholar] [CrossRef]
- Bloom, R.J.; George, J.P.; Celedon, A.; Sun, S.X.; Wirtz, D. Mapping local matrix remodeling induced by a migrating tumor cell using three-dimensional multiple-particle tracking. Biophys. J. 2008, 95, 4077–4088. [Google Scholar] [CrossRef] [Green Version]
- Legant, W.R.; Miller, J.S.; Blakely, B.L.; Cohen, D.M.; Genin, G.M.; Chen, C.S. Measurement of mechanical tractions exerted by cells in three-dimensional matrices. Nat. Methods 2010, 7, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Wang, X. Recent advances in cell adhesive force microscopy. Sensors 2020, 20, 7128. [Google Scholar] [CrossRef]
- Maskarinec, S.A.; Franck, C.; Tirrell, D.A.; Ravichandran, G. Quantifying cellular traction forces in three dimensions. Proc. Natl. Acad. Sci. USA 2009, 106, 22108–22113. [Google Scholar] [CrossRef] [Green Version]
- Mulligan, J.A.; Feng, X.; Adie, S.G. Quantitative reconstruction of time-varying 3D cell forces with traction force optical coherence microscopy. Sci. Rep. 2019, 9, 4086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, T.M.; Münster, S.; Bonakdar, N.; Butler, J.P.; Fabry, B. 3D traction forces in cancer cell invasion. PLoS ONE 2012, 7, e33476. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Naruse, K.; Stamenović, D.; Fredberg, J.J.; Mijailovich, S.M.; Tolić-Nørrelykke, I.M.; Polte, T.; Mannix, R.; Ingber, D.E. Mechanical behavior in living cells consistent with the tensegrity model. Proc. Natl. Acad. Sci. USA 2001, 98, 7765–7770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, U.S.; Balaban, N.Q.; Riveline, D.; Bershadsky, A.; Geiger, B.; Safran, S.A. Calculation of forces at focal adhesions from elastic substrate data: The effect of localized force and the need for regularization. Biophys. J. 2002, 83, 1380–1394. [Google Scholar] [CrossRef] [Green Version]
- Colin-York, H.; Shrestha, D.; Felce, J.H.; Waithe, D.; Moeendarbary, E.; Davis, S.J.; Eggeling, C.; Fritzsche, M. Super-Resolved Traction Force Microscopy (STFM). Nano Lett. 2016, 16, 2633–2638. [Google Scholar] [CrossRef] [PubMed]
- Stubb, A.; Laine, R.F.; Miihkinen, M.; Hamidi, H.; Guzmán, C.; Henriques, R.; Jacquemet, G.; Ivaska, J. Fluctuation-Based Super-Resolution Traction Force Microscopy. Nano Lett. 2020, 20, 2230–2245. [Google Scholar] [CrossRef]
- Morimatsu, M.; Mekhdjian, A.H.; Chang, A.C.; Tan, S.J.; Dunn, A.R. Visualizing the Interior Architecture of Focal Adhesions with High-Resolution Traction Maps. Nano Lett. 2015, 15, 2220–2228. [Google Scholar] [CrossRef] [Green Version]
- Barnhart, E.; Lee, K.-C.; Allen, G.M.; Theriot, J.A.; Mogilner, A. Balance between cell−substrate adhesion and myosin contraction determines the frequency of motility initiation in fish keratocytes. Proc. Natl. Acad. Sci. USA 2015, 112, 5045–5050. [Google Scholar] [CrossRef] [Green Version]
- Legant, W.R.; Choi, C.K.; Miller, J.S.; Shao, L.; Gao, L.; Betzig, E.; Chen, C.S. Multidimensional traction force microscopy reveals out-of-plane rotational moments about focal adhesions. Proc. Natl. Acad. Sci. USA 2013, 110, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Mpiv-MatLab PIV Toolbox. Available online: http://www.oceanwave.jp/softwares/mpiv/ (accessed on 1 August 2021).
- The MatLab Particle Tracking Code Repository. Available online: http://site.physics.georgetown.edu/matlab/ (accessed on 1 August 2021).
- Barrasa-Fano, J.; Shapeti, A.; Jorge-Peñas, Á.; Barzegari, M.; Sanz-Herrera, J.A.; Van Oosterwyck, H. TFMLAB: A MATLAB toolbox for 4D traction force microscopy. SoftwareX 2021, 15, 100723. [Google Scholar] [CrossRef]
- Du Roure, O.; Saez, A.; Buguin, A.; Austin, R.H.; Chavrier, P.; Siberzan, P.; Ladoux, B. Force mapping in epithelial cell migration. Proc. Natl. Acad. Sci. USA 2005, 102, 2390–2395. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Sniadecki, N.J.; Chen, C.S. Mechanical forces in endothelial cells during firm adhesion and early transmigration of human monocytes. Cell. Mol. Bioeng. 2010, 3, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legant, W.R.; Chen, C.S.; Vogel, V. Force-induced fibronectin assembly and matrix remodeling in a 3D microtissue model of tissue morphogenesis. Integr. Biol. 2012, 4, 1164–1174. [Google Scholar] [CrossRef]
- West, A.R.; Zaman, N.; Cole, D.J.; Walker, M.J.; Legant, W.R.; Boudou, T.; Chen, C.S.; Favreau, J.T.; Gaudette, G.R.; Cowley, E.A.; et al. Development and characterization of a 3D multicell microtissue culture model of airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 304, L4–L16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.G.; Han, S.J.; Regnier, M.; Sniadecki, N.J. Substrate stiffness increases twitch power of neonatal cardiomyocytes in correlation with changes in myofibril structure and intracellular calcium. Biophys. J. 2011, 101, 2455–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakar, M.S.; Neal, D.; Boudou, T.; Borochin, M.A.; Li, Y.; Weiss, R.; Kamm, R.D.; Chen, C.S.; Asada, H.H. Formation and optogenetic control of engineered 3D skeletal muscle bioactuators. Lab Chip 2012, 12, 4976–7985. [Google Scholar] [CrossRef] [Green Version]
- Boudou, T.; Legant, W.R.; Mu, A.; Borochin, M.A.; Thavandiran, N.; Radisic, M.; Zandstra, P.W.; Epstein, J.A.; Margulies, K.B.; Chen, C.S. A microfabricated platform to measure and manipulate the mechanics of engineered cardiac microtissues. Tissue Eng. Part A 2012, 18, 910–919. [Google Scholar] [CrossRef]
- Kajzar, A.; Cesa, C.M.; Kirchgeßner, N.; Hoffmann, B.; Merkel, R. Toward Physiological Conditions for Cell Analyses: Forces of Heart Muscle Cells Suspended Between Elastic Micropillars. Biophys. J. 2008, 94, 1854–1866. [Google Scholar] [CrossRef] [Green Version]
- Ricart, B.G.; Yang, M.T.; Hunter, C.A.; Chen, C.S.; Hammer, D.A. Measuring traction forces of motile dendritic cells on micropost arrays. Biophys. J. 2011, 101, 2620–2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.T.; Sniadecki, N.J.; Chen, C.S. Geometric considerations of micro- To nanoscale elastomeric post arrays to study cellular traction forces. Adv. Mater. 2007, 19, 3119–3123. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Di Lullo, G.A.; Sweeney, S.M.; Körkkö, J.; Ala-Kokko, L.; San Antonio, J.D. Mapping the ligand-binding sites and disease-associated mutations on the most abundant protein in the human, type I collagen. J. Biol. Chem. 2002, 277, 4223–4231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, E.; Ivarsson, B.; Merrill, C. Production of a tissue-like structure by contraction of collagen lattices by human fibroblasts of different proliferative potential in vitro. Proc. Natl. Acad. Sci. USA 1979, 76, 1274–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo, P.; Ramalingam, P.; Phillips, J.A.; Furuta, G.T. Collagen Gel Contraction Assay. In Cell-Cell Interactions: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2006; pp. 103–109. ISBN 978-1-59745-113-0. [Google Scholar]
- Tomasek, J.J.; Haaksma, C.J.; Eddy, R.J.; Vaughan, M.B. Fibroblast contraction occurs on release of tension in attached collagen lattices: Dependency on an organized actin cytoskeleton and serum. Anat. Rec. 1992, 232, 359–368. [Google Scholar] [CrossRef]
- Ehrlich, H.P.; Rajaratnam, J.B.M. Cell locomotion forces versus cell contraction forces for collagen lattice contraction: An in vitro model of wound contraction. Tissue Cell 1990, 22, 407–417. [Google Scholar] [CrossRef]
- Módis, L. Organization of the Extracellular Matrix: A Polarization Microscopic Approach, 1st ed.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9781351075282. [Google Scholar]
- Travis, J.A.; Hughes, M.G.; Wong, J.M.; Wagner, W.D.; Geary, R.L. Hyaluronan enhances contraction of collagen by smooth muscle cells and adventitial fibroblasts role of CD44 and implications for constrictive remodeling. Circ. Res. 2001, 88, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Fitts, R.H. The cross-bridge cycle and skeletal muscle fatigue. J. Appl. Physiol. 2008, 104, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Horie, M.; Saito, A.; Yamauchi, Y.; Mikami, Y.; Sakamoto, M.; Jo, T.; Nakajima, J.; Takizawa, H.; Nagase, T.; Kohyama, T. Histamine induces human lung fibroblast-mediated collagen gel contraction via histamine H1 receptor. Exp. Lung Res. 2014, 40, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Morales, S.A.; Mareninov, S.; Prasad, P.; Wadehra, M.; Braun, J.; Gordon, L.K. Collagen gel contraction by ARPE-19 cells is mediated by a FAK-Src dependent pathway. Exp. Eye Res. 2007, 85, 790–798. [Google Scholar] [CrossRef]
- Yamato, M.; Yamamoto, K.; Hayashi, T. Age-related changes in collagen gel contraction by cultured human lung fibroblasts resulting in cross-over of contraction curves between young and aged cells. Mech. Ageing Dev. 1993, 67, 149–158. [Google Scholar] [CrossRef]
- Hu, K.; Shi, H.; Zhu, J.; Deng, D.; Zhou, G.; Zhang, W.; Cao, Y.; Liu, W. Compressed collagen gel as the scaffold for skin engineering. Biomed. Microdevices 2010, 12, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Tingstrom, A.; Heldin, C.H.; Rubin, K. Regulation of fibroblast-mediated collagen gel contraction by platelet-derived growth factor, interleukin-1 α and transforming growth factor-β1. J. Cell Sci. 1992, 102, 315–322. [Google Scholar] [CrossRef]
- Ehrlich, H.P.; Hunt, T.K. Collagen Organization Critical Role in Wound Contraction. Adv. Wound Care 2012, 1, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, T.; Li, L.; Siow, R.C.M.; Liu, K. Collagen matrix stiffness influences fibroblast contraction force. Biomed. Phys. Eng. Express 2016, 2, 047002. [Google Scholar] [CrossRef] [Green Version]
- Wilks, B.T.; Evans, E.B.; Nakhla, M.N.; Morgan, J.R. Directing fibroblast self-assembly to fabricate highly-aligned, collagen-rich matrices. Acta Biomater. 2018, 81, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Duan, L.; Zhu, W.; Xiong, J.; Wang, D. Extracellular matrix production in vitro in cartilage tissue engineering. J. Transl. Med. 2014, 12, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastwood, M.; McGrouther, D.A.; Brown, R.A. A culture force monitor for measurement of contraction forces generated in human dermal fibroblast cultures: Evidence for cell-matrix mechanical signalling. BBA Gen. Subj. 1994, 1201, 186–192. [Google Scholar] [CrossRef]
- Peperzak, K.A.; Gilbert, T.W.; Wang, J.H.C. A multi-station dynamic-culture force monitor system to study cell mechanobiology. Med. Eng. Phys. 2004, 26, 355–358. [Google Scholar] [CrossRef]
- Freyman, T.M.; Yannas, I.V.; Yokoo, R.; Gibson, L.J. Fibroblast contraction of a collagen-GAG matrix. Biomaterials 2001, 22, 2883–2891. [Google Scholar] [CrossRef]
- Kolodney, M.S.; Wysolmerski, R.B. Isometric contraction by fibroblasts and endothelial cells in tissue culture: A quantitative study. J. Cell Biol. 1992, 117, 73–82. [Google Scholar] [CrossRef]
- Zimmermann, W.H.; Fink, C.; Kralisch, D.; Remmers, U.; Weil, J.; Eschenhagen, T. Three-dimensional engineered heart tissue from neonatal rat cardiac myocytes. Biotechnol. Bioeng. 2000, 68, 106–114. [Google Scholar] [CrossRef]
- Eichinger, J.F.; Haeusel, L.J.; Paukner, D.; Aydin, R.C.; Humphrey, J.D.; Cyron, C.J. Mechanical homeostasis in tissue equivalents: A review. Biomech. Model. Mechanobiol. 2021, 20, 833–850. [Google Scholar] [CrossRef]
- Petersen, O.W.; Ronnov-Jessen, L.; Howlett, A.R.; Bissell, M.J. Interaction with basement membrane serves to rapidly distinguish growth and differentiation pattern of normal and malignant human breast epithelial cells. Proc. Natl. Acad. Sci. USA 1992, 89, 9064–9068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulrich, T.A.; De Juan Pardo, E.M.; Kumar, S. The mechanical rigidity of the extracellular matrix regulates the structure, motility, and proliferation of glioma cells. Cancer Res. 2009, 69, 4167–4174. [Google Scholar] [CrossRef] [Green Version]
- Jaalouk, D.E.; Lammerding, J. Mechanotransduction gone awry. Nat. Rev. Mol. Cell Biol. 2009, 10, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Ingber, D.E. Cell tension, matrix mechanics, and cancer development. Cancer Cell 2005, 8, 175–176. [Google Scholar] [CrossRef] [Green Version]
- Discher, D.E.; Janmey, P.; Wang, Y.L. Tissue cells feel and respond to the stiffness of their substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Marklein, R.A.; Burdick, J.A. Spatially controlled hydrogel mechanics to modulate stem cell interactions. Soft Matter 2010, 6, 136–143. [Google Scholar] [CrossRef]
- Banerjee, A.; Arha, M.; Choudhary, S.; Ashton, R.S.; Bhatia, S.R.; Schaffer, D.V.; Kane, R.S. The influence of hydrogel modulus on the proliferation and differentiation of encapsulated neural stem cells. Biomaterials 2009, 30, 4695–4699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.K.; Umino, T.; Liu, X.D.; Wang, H.J.; Romberger, D.J.; Spurzem, J.R.; Rennard, I.; In, S.; Cellular, V.; Animal, D.B.; et al. Contraction of fibroblast-containing collagen cels: Initial collagen concentration regulates the degree of contraction and cell survival. In Vitro Cell. Dev. Biol. Anima. 2001, 37, 10–16. [Google Scholar] [CrossRef]
- Ahearne, M.; Wilson, S.L.; Liu, K.-K.; Rauz, S.; El Haj, A.J.; Yang, Y. Influence of cell and collagen concentration on the cell-matrix mechanical relationship in a corneal stroma wound healing model. Exp. Eye Res. 2010, 91, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewoldt, R.H.; Hosoi, A.E.; McKinley, G.H. Nonlinear viscoelastic biomaterials: Meaningful characterization and engineering inspiration. Integr. Comp. Biol. 2009, 49, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Fratzl, P. Collagen: Structure and Mechanics; Springer: Boston, MA, USA, 2008; ISBN 9780387739052. [Google Scholar]
- Ahearne, M. Introduction to cell-hydrogel mechanosensing. Interface Focus 2014, 4, 20130038. [Google Scholar] [CrossRef]
- Chaudhuri, O.; Cooper-White, J.; Janmey, P.A.; Mooney, D.J.; Shenoy, V.B. Effects of extracellular matrix viscoelasticity on cellular behaviour. Nature 2020, 584, 535–546. [Google Scholar] [CrossRef]
- Xu, Q.; Engquist, B. A mathematical model for fitting and predicting relaxation modulus and simulating viscoelastic responses. Proc. R. Soc. Math. Phys. Eng. Sci. 2018, 474, 20170540. [Google Scholar] [CrossRef] [Green Version]
- Tirella, A.; Mattei, G.; Ahluwalia, A. Strain rate viscoelastic analysis of soft and highly hydrated biomaterials. J. Biomed. Mater. Res. Part A 2014, 102, 3352–3360. [Google Scholar] [CrossRef] [Green Version]
- Kalyanam, S.; Yapp, R.D.; Insana, M.F. Poro-viscoelastic behavior of gelatin hydrogels under compression-implications for bioelasticity imaging. J. Biomech. Eng. 2009, 131, 081005. [Google Scholar] [CrossRef] [Green Version]
- Ahearne, M.; Siamantouras, E.; Yang, Y.; Liu, K.-K. Mechanical characterization of biomimetic membranes by micro-shaft poking. J. R. Soc. Interface 2009, 6, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahearne, M.; Yang, Y.; El Haj, A.J.; Then, K.Y.; Liu, K.-K. Characterizing the viscoelastic properties of thin hydrogel-based constructs for tissue engineering applications. J. R. Soc. Interface 2005, 2, 455–463. [Google Scholar] [CrossRef]
- Cheng, L.; Xia, X.; Scriven, L.E.; Gerberich, W.W. Spherical-tip indentation of viscoelastic material. Mech. Mater. 2005, 37, 213–226. [Google Scholar] [CrossRef]
- Mattice, J.M.; Lau, A.G.; Oyen, M.L.; Kent, R.W. Spherical indentation load-relaxation of soft biological tissues. J. Mater. Res. 2006, 21, 2003–2010. [Google Scholar] [CrossRef]
- Friedrich, C. Relaxation and retardation functions of the Maxwell model with fractional derivatives. Rheol. Acta 1991, 30, 151–158. [Google Scholar] [CrossRef]
- Eldred, L.B.; Baker, W.P.; Palazotto, A.N. Kelvin-Voigt versus fractional derivative model as constitutive relations for viscoelastic materials. AIAA J. 1995, 33, 547–550. [Google Scholar] [CrossRef]
- Tanner, R.I. Engineering Rheology, 2nd ed.; Oxford University Press: Oxford, UK, 1982. [Google Scholar]
- Li, S.; Patwardhan, A.G.; Amirouche, F.M.L.; Havey, R.; Meade, K.P. Limitations of the standard linear solid model of intervertebral discs subject to prolonged loading and low-frequency vibration in axial compression. J. Biomech. 1995, 28, 779–790. [Google Scholar] [CrossRef]
- Abuzeid, O.M.; Eberhard, P. Linear Viscoelastic Creep Model for the Contact of Nominal Flat Surfaces Based on Fractal Geometry: Standard Linear Solid (SLS) Material. J. Tribol. 2007, 129, 461–466. [Google Scholar] [CrossRef]
- Wiechert, E. Gesetze der elastischen Nachwirkung für constante Temperatur. Ann. Phys. 1893, 286, 546–570. [Google Scholar] [CrossRef]
- Gieni, R.S.; Hendzel, M.J. Mechanotransduction from the ECM to the genome: Are the pieces now in place? J. Cell. Biochem. 2008, 104, 1964–1987. [Google Scholar] [CrossRef] [PubMed]
- Tigges, J.; Krutmann, J.; Fritsche, E.; Haendeler, J.; Schaal, H.; Fischer, J.W.; Kalfalah, F.; Reinke, H.; Reifenberger, G.; Stühler, K.; et al. The hallmarks of fibroblast ageing. Mech. Ageing Dev. 2014, 138, 26–44. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zünd, G.; Benedikt, P.; Jockenhoevel, S.; Hoerstrup, S.P.; Sakyama, S.; Hubbell, J.A.; Turina, M. Fibrin gel as a three dimensional matrix in cardiovascular tissue engineering. Eur. J. Cardio-Thorac. Surg. 2000, 17, 587–591. [Google Scholar] [CrossRef]
- Shaikh, F.M.; Callanan, A.; Kavanagh, E.G.; Burke, P.E.; Grace, P.A.; McGloughlin, T.M. Fibrin: A natural biodegradable scaffold in vascular tissue engineering. Cells. Tissues. Organs 2008, 188, 333–346. [Google Scholar] [CrossRef] [PubMed]
- de la Puente, P.; Ludeña, D. Cell culture in autologous fibrin scaffolds for applications in tissue engineering. Exp. Cell Res. 2014, 322, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.N.; Birkinshaw, C. Hyaluronic acid based scaffolds for tissue engineering—A review. Carbohydr. Polym. 2013, 92, 1262–1279. [Google Scholar] [CrossRef]
- Seidlits, S.K.; Drinnan, C.T.; Petersen, R.R.; Shear, J.B.; Suggs, L.J.; Schmidt, C.E. Fibronectin–hyaluronic acid composite hydrogels for three-dimensional endothelial cell culture. Acta Biomater. 2011, 7, 2401–2409. [Google Scholar] [CrossRef]
- Perng, C.-K.; Wang, Y.-J.; Tsi, C.-H.; Ma, H. In Vivo Angiogenesis Effect of Porous Collagen Scaffold with Hyaluronic Acid Oligosaccharides. J. Surg. Res. 2011, 168, 9–15. [Google Scholar] [CrossRef]
- Trujillo, S.; Gonzalez-Garcia, C.; Rico, P.; Reid, A.; Windmill, J.; Dalby, M.J.; Salmeron-Sanchez, M. Engineered 3D hydrogels with full-length fibronectin that sequester and present growth factors. Biomaterials 2020, 252, 120104. [Google Scholar] [CrossRef]
- Zhu, J. Bioactive modification of poly(ethylene glycol) hydrogels for tissue engineering. Biomaterials 2010, 31, 4639–4656. [Google Scholar] [CrossRef] [Green Version]
- Petrini, P.; Farè, S.; Piva, A.; Tanzi, M.C. Design, synthesis and properties of polyurethane hydrogels for tissue engineering. J. Mater. Sci. Mater. Med. 2003, 14, 683–686. [Google Scholar] [CrossRef]
- Hong, S.; Sycks, D.; Chan, H.F.; Lin, S.; Lopez, G.P.; Guilak, F.; Leong, K.W.; Zhao, X. 3D Printing of Highly Stretchable and Tough Hydrogels into Complex, Cellularized Structures. Adv. Mater. 2015, 27, 4035–4040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wu, C.; Chu, P.K.; Gelinsky, M. 3D printing of hydrogels: Rational design strategies and emerging biomedical applications. Mater. Sci. Eng. R Rep. 2020, 140, 100543. [Google Scholar] [CrossRef]
- Parada, G.A.; Zhao, X. Ideal reversible polymer networks. Soft Matter 2018, 14, 5186–5196. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.R.; Frith, J.E.; Cooper-White, J.J. The influence of substrate creep on mesenchymal stem cell behaviour and phenotype. Biomaterials 2011, 32, 5979–5993. [Google Scholar] [CrossRef]
- Charrier, E.E.; Pogoda, K.; Wells, R.G.; Janmey, P.A. Control of cell morphology and differentiation by substrates with independently tunable elasticity and viscous dissipation. Nat. Commun. 2018, 9, 449. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Ma, H.; Tu, H.-C.; Wang, H.-R.; Lin, P.-C.; Anseth, K.S. Adaptable Fast Relaxing Boronate-Based Hydrogels for Probing Cell–Matrix Interactions. Adv. Sci. 2018, 5, 1800638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marozas, I.A.; Anseth, K.S.; Cooper-White, J.J. Adaptable boronate ester hydrogels with tunable viscoelastic spectra to probe timescale dependent mechanotransduction. Biomaterials 2019, 223, 119430. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Huebsch, N.; Mooney, D.J.; Suo, Z. Stress-relaxation behavior in gels with ionic and covalent crosslinks. J. Appl. Phys. 2010, 107, 63509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dooling, L.J.; Buck, M.E.; Zhang, W.-B.; Tirrell, D.A. Programming Molecular Association and Viscoelastic Behavior in Protein Networks. Adv. Mater. 2016, 28, 4651–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sniadecki, N.J.; Anguelouch, A.; Yang, M.T.; Lamb, C.M.; Liu, Z.; Kirschner, S.B.; Liu, Y.; Reich, D.H.; Chen, C.S. Magnetic microposts as an approach to apply forces to living cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14553–14558. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-C.; Kramer, C.M.; Chen, C.S.; Reich, D.H. Probing cellular traction forces with magnetic nanowires and microfabricated force sensor arrays. Nanotechnology 2012, 23, 75101. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polymer Origin | Polymer Type | Principles | Advantages/ Disadvantages | |
|---|---|---|---|---|
| Synthetic | DM | Silicon rubber | Length and patterns of wrinkles on film shows force generation | Simple and cheap/ Not cell-friendly; not quantitative |
| 2D TFM | Poly- acrylamide; PEG | Use fluorescent microbeads to track substrate deformation due to cells seeded on the surface | Highly quantitative/ Not 3D; not cell-friendly | |
| 3D TFM | Tracking of matrix deformation due to embedded cells in 3D | Highly quantitative, 3D/ Not mimicking in vivo environment; computationally extensive | ||
| EMP | PDMS; PMMA | Optically measure the deflection of micro-pillar array | Highly quantitative/ Not 3D; not cell-friendly | |
| Natural | CGCA | Collagen gel | Measure the geometry of cell-embedded collagen hydrogel | Highly cell friendly; 3D/ Qualitative |
| CFM | Collagen gel | Continuously measure the force generated through attached strain gauges | 3D; qualitative/ Complex; disturbance to cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Liu, K.-K. Soft Polymer-Based Technique for Cellular Force Sensing. Polymers 2021, 13, 2672. https://doi.org/10.3390/polym13162672
Yu Z, Liu K-K. Soft Polymer-Based Technique for Cellular Force Sensing. Polymers. 2021; 13(16):2672. https://doi.org/10.3390/polym13162672
Chicago/Turabian StyleYu, Zhuonan, and Kuo-Kang Liu. 2021. "Soft Polymer-Based Technique for Cellular Force Sensing" Polymers 13, no. 16: 2672. https://doi.org/10.3390/polym13162672
APA StyleYu, Z., & Liu, K. -K. (2021). Soft Polymer-Based Technique for Cellular Force Sensing. Polymers, 13(16), 2672. https://doi.org/10.3390/polym13162672