
Surface and Thermal Characterization of Cotton Fibers of Phenotypes Differing in Fiber Length





Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Sample Preparation
2.2. Microscopically Morphologic Measurements
2.3. Water Contact Angle Measurement
2.4. Attenuated Total Reflection Reflectance Fourier Transform Infrared (ATR FT-IR) Spectroscopy
2.5. Thermogravimetric (TG) and Differential Thermogravimetric (DTG) Measurements
2.6. Data Treatments and Statistical Analysis
3. Results and Discussion
3.1. Optical Microscopic Images
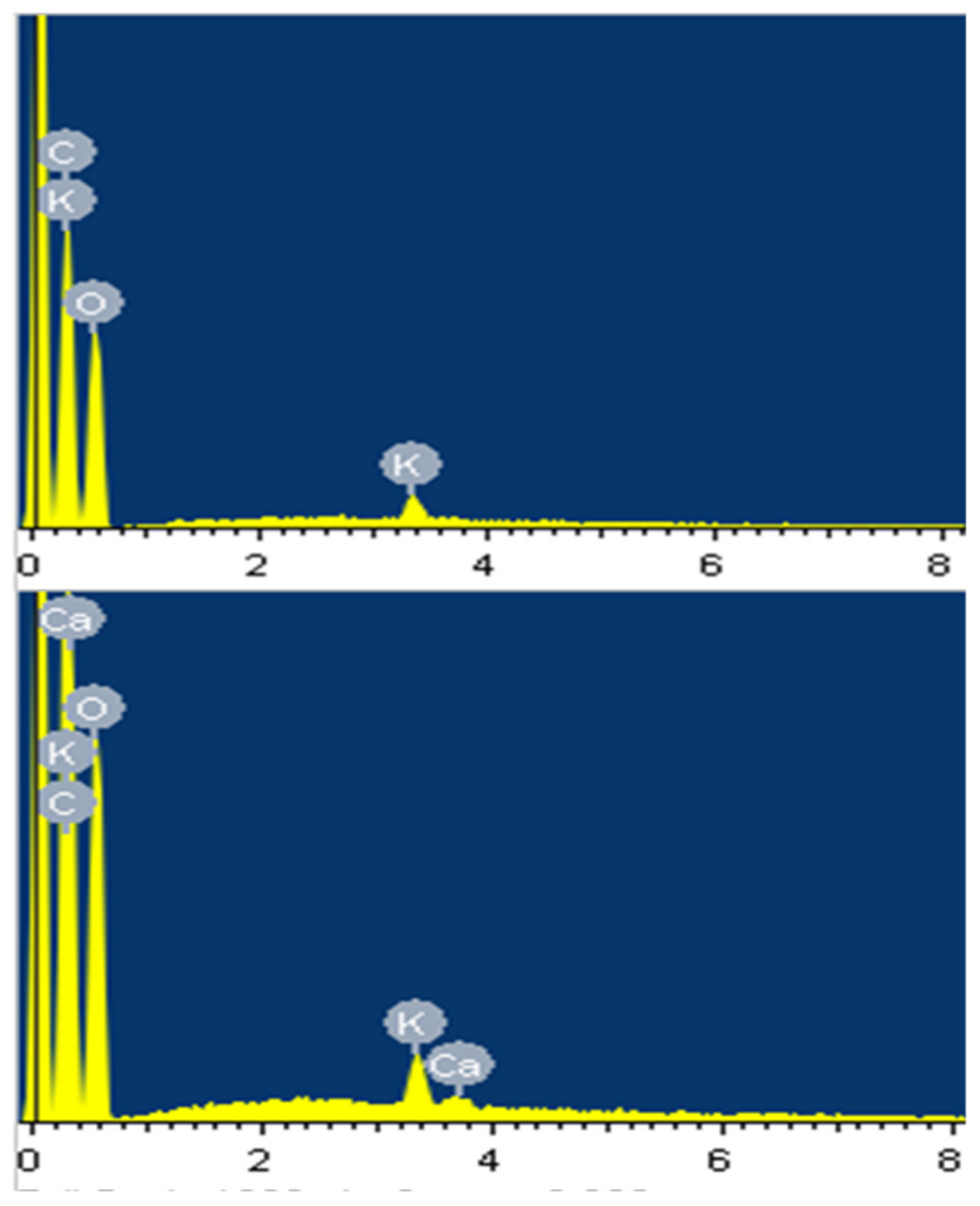
3.2. SEM and SEM–EDS Morphological Characteristics
3.3. Wetting Behavior
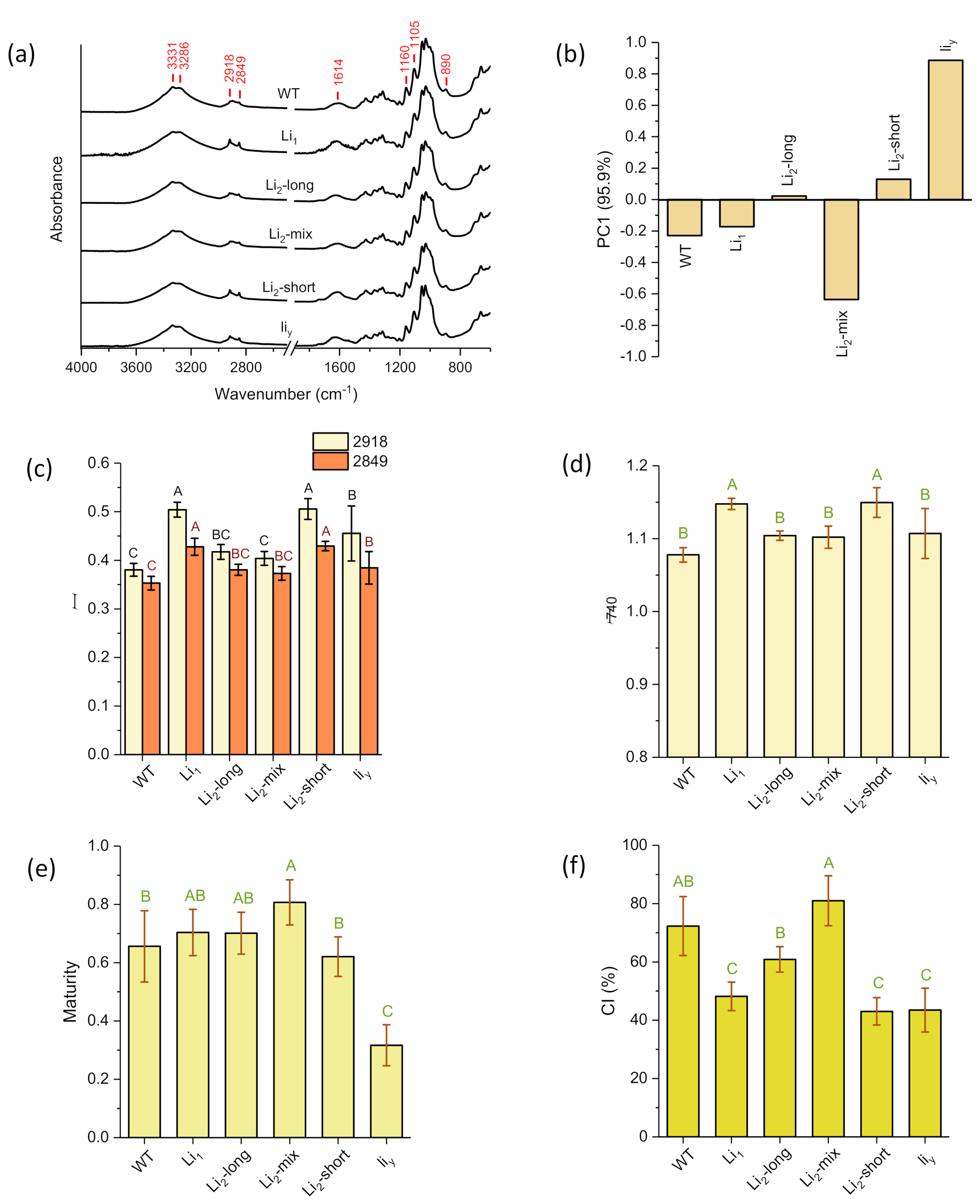
3.4. FT-IR Analysis
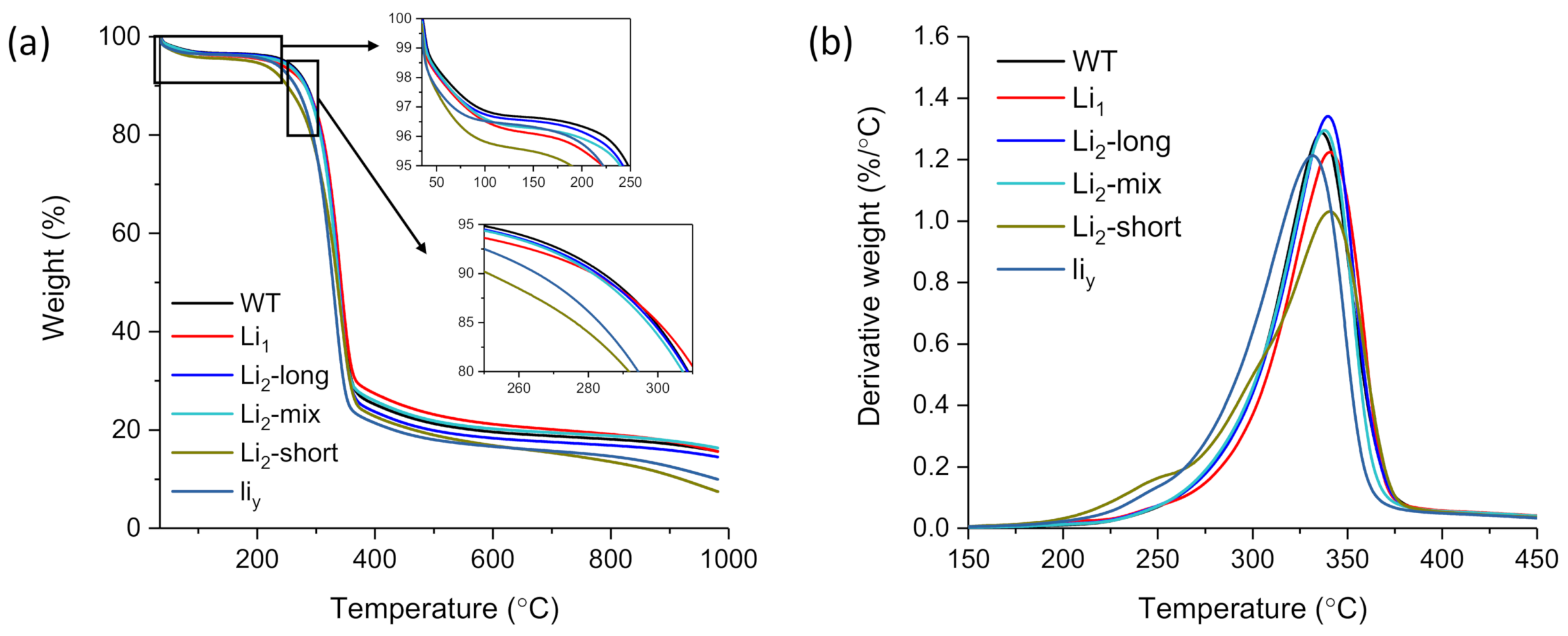
3.5. Thermogravimetric Analysis
3.6. Correlation Analysis of the Two Sets of ATR FT-IR and Thermogravimetric Measurements
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, D.D.; Percy, R.G. Cotton, 2nd ed.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2015. [Google Scholar]
- Liu, Y.; Kim, H.-J. Fourier Transform Infrared Spectroscopy (FT-IR) and Simple Algorithm Analysis for Rapid and Non-Destructive Assessment of Developmental Cotton Fibers. Sensors 2017, 17, 1469. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.K.; Turley, R.B.; Kim, H.J.; Li, P.; Thyssen, G.; Tang, Y.; Delhom, C.D.; Naoumkina, M.; Fang, D.D. Transcript profiling by microarray and marker analysis of the short cotton (Gossypium hirsutum L.) fiber mutant Ligon lintless-1 (Li 1). BMC Genom. 2013, 14, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, D.D.; Naoumkina, M.; Thyssen, G.N.; Bechere, E.; Li, P.; Florane, C.B. An EMS-induced mutation in a tetratricopeptide repeat-like superfamily protein gene (Ghir_A12G008870) on chromosome A12 is responsible for the liy short fiber phenotype in cotton. Theor. Appl. Genet. 2020, 133, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Jiang, Y.; Cao, Y.; Lin, L.; He, S.; Zhou, W.; Rong, J. Gene expression profile analysis of Ligon lintless-1 (Li1) mutant reveals important genes and pathways in cotton leaf and fiber development. Gene 2014, 535, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Bechere, E.; Fang, D.D.; Naoumkina, M. A new Ligon-lintless mutant (liy) in upland cotton. J. Cotton Sci. 2020, 24, 168–174. [Google Scholar]
- Fang, D.D.; Naoumkina, M.; Kim, H.J. Unraveling Cotton Fiber Development Using Fiber Mutants in the Post-Genomic Era. Crop. Sci. 2018, 58, 2214–2228. [Google Scholar] [CrossRef]
- Turley, R.B.; Kloth, R.H. The inheritance model for the fiberless trait in upland cotton (Goss-ypium hirsutum L.) line SL1-7-1: Variation on a theme. Euphytica 2008, 164, 123–132. [Google Scholar] [CrossRef]
- Zhang, T.Z.; Pan, J.J. Genetic analysis of a fuzzless-lintless mutant in Gossypium hirsutum L. Jiangsu J. Agr. Sci. 1991, 7, 13–16. [Google Scholar]
- Kearney, T.H.; Harrison, G.J. Inheritance of smooth seeds in cotton. J. Agric. Res. 1927, 3, 193–217. [Google Scholar]
- Ware, J.O.; Benedict, L.I.; Rolfe, W.H. A Recessive Naked-Seed Character in Upland Cotton. J. Hered. 1947, 38, 313–320. [Google Scholar] [CrossRef]
- Kohel, R.J. Linkage Tests in Upland Cotton, Gossypium hirsutum L. II. 1. Crop. Sci. 1972, 12, 66–69. [Google Scholar] [CrossRef]
- Narbuth, E.V.; Kohel, R.J. Inheritance and Linkage Analysis of a New Fiber Mutant in Cotton. J. Hered. 1990, 81, 131–133. [Google Scholar] [CrossRef]
- Cai, C.; Tong, X.; Liu, F.; Lv, F.; Wang, H.; Zhang, T.; Guo, W. Discovery and identification of a novel Ligon lintless-like mutant (Lix) similar to the Ligon lintless (Li1) in allotetraploid cotton. Theor. Appl. Genet. 2013, 126, 963–970. [Google Scholar] [CrossRef]
- Thyssen, G.N.; Fang, D.D.; Turley, R.B.; Florane, C.B.; Li, P.; Mattison, C.P.; Naoumkina, M. A Gly65Val substitution in an actin, Ghact_Gi1, disrupts cell polarity and f-actin organization resulting in dwarf, lintless cotton plants. Plant J. 2017, 90, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.K.; Kim, H.J.; Tang, Y.; Naoumkina, M.; Fang, D.D. Comparative transcriptome analysis of short fiber mutants Ligon-lintless 1 and 2 reveals common mechanisms pertinent to fiber elongation in cotton (Gossypium hirsutum L.). PLoS ONE 2014, 9, e95554. [Google Scholar] [CrossRef]
- An, C.; Jenkins, J.N.; McCarty, J.C., Jr.; Saha, S. Atypical Ligon lintless-2 phenotype in cotton. J. Cotton Sci. 2010, 14, 13–16. [Google Scholar]
- Patel, J.D.; Huang, X.; Lin, L.; Das, S.; Chandnani, R.; Khanal, S.; Adhikari, J.; Shehzad, T.; Guo, H.; Roy-Zokan, E.M.; et al. The Ligon lintless-2 Short Fiber Mutation Is Located within a Terminal Deletion of Chromosome 18 in Cotton. Plant Physiol. 2020, 183, 277–288. [Google Scholar] [CrossRef]
- Naoumkina, M.; Bechere, E.; Fang, D.D.; Thyssen, G.N.; Florane, C.B. Genome-wide analysis of gene expression of EMS-induced short fiber mutant Ligon lintless-y (liy) in cotton (Gossypium hirsutum L.). Genomics 2017, 109, 320–329. [Google Scholar] [CrossRef]
- Hinchliffe, D.J.; Turley, R.B.; Naoumkina, M.; Kim, H.J.; Tang, Y.; Yeater, K.M.; Li, P.; Fang, D.D. A combined functional and structural genomics approach identified an EST-SSR marker with complete linkage to the Ligon lintless-2 genetic locus in cotton (Gossypium hirsutum L.). BMC Genom. 2011, 12, 445. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Thibodeaux, D.; Gamble, G. Development of Fourier transform infrared spectroscopy in direct, non-destructive, and rapid determination of cotton fiber maturity. Text. Res. J. 2011, 81, 1559–1567. [Google Scholar] [CrossRef]
- Liu, Y.; Thibodeaux, D.; Gamble, G.; Bauer, P.; Vanderveer, D. Comparative Investigation of Fourier Transform Infrared (FT-IR) Spectroscopy and X-ray Diffraction (XRD) in the Determination of Cotton Fiber Crystallinity. Appl. Spectrosc. 2012, 66, 983–986. [Google Scholar] [CrossRef]
- Seagull, R.W.; Oliveri, V.; Murphy, K.; Binder, A.; Kothari, S. Cotton fiber growth and development 2. Changes in cell diameter and wall birefringence. J. Cotton Sci. 2000, 4, 97–104. [Google Scholar]
- Liyanage, S.; Abidi, N. Molecular weight and organization of cellulose at different stages of cotton fiber development. Text. Res. J. 2018, 89, 726–738. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, C.M.; Dazen, K.; Delhom, C.D.; Liu, Y.; Rodgers, J.E.; French, A.D.; Kim, S.H. Comparative physical and chemical analyses of cotton fibers from two near isogenic upland lines differing in fiber wall thickness. Cellulose 2017, 24, 2385–2401. [Google Scholar] [CrossRef]
- Seagull, R.W. Cytoskeletal involvement in cotton fiber growth and development. Micron 1993, 24, 643–660. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, Z.; Zhou, J.; Zhang, Y. Reuse of waste cotton cloth for the extraction of cellulose nanocrystals. Carbohydr. Polym. 2017, 157, 945–952. [Google Scholar] [CrossRef]
- Zhao, H.; Kwak, J.H.; Zhang, Z.C.; Brown, H.M.; Arey, B.W.; Holladay, J.E. Studying cellulose fiber structure by SEM, XRD, NMR and acid hydrolysis. Carbohydr. Polym. 2007, 68, 235–241. [Google Scholar] [CrossRef]
- He, Z.; Cheng, H.N.; Olanya, O.M.; Uknalis, J.; Zhang, X.; Koplitz, B.D.; He, J. Surface Characterization of Cottonseed Meal Products by SEM, SEM-EDS, XRD and XPS Analysis. J. Mater. Sci. Res. 2017, 7, 28–40. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Zhang, H.; Fang, D.D.; Zeng, L.; Jenkins, J.N.; McCarty, J.C. Effects of inter-species chromosome substitution on cottonseed mineral and protein nutrition profiles. Agron. J. 2020, 112, 3963–3974. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Tewolde, H.; Shankle, M. Chemical Characterization of Cotton Plant Parts for Multiple Uses. Agric. Environ. Lett. 2017, 2, 110044. [Google Scholar] [CrossRef] [Green Version]
- Gamble, G.R. Regional, varietal, and crop year variations of metal contents associated with the separate structural components of upland cotton (Gossypium hirsutum) fiber. J. Cotton Sci. 2009, 13, 221–226. [Google Scholar]
- Guo, K.; Tu, L.; He, Y.; Deng, J.; Wang, M.; Huang, H.; Li, Z.; Zhang, X. Interaction between calcium and potassium modulates elongation rate in cotton fiber cells. J. Exp. Bot. 2017, 68, 5161–5175. [Google Scholar] [CrossRef]
- Nam, S.; Park, Y.; Hillyer, M.B.; Hron, R.J.; Ernst, N.; Chang, S.; Condon, B.D.; Hinchliffe, D.J.; Ford, E.; Gibb, B.C. Thermal properties and surface chemistry of cotton varieties mineralized with calcium carbonate polymorphs by cyclic dipping. RSC Adv. 2020, 10, 35214–35225. [Google Scholar] [CrossRef]
- Wang, J.; Liu, S. Remodeling of raw cotton fiber into flexible, squeezing-resistant macroporous cellulose aerogel with high oil retention capability for oil/water separation. Sep. Purif. Technol. 2019, 221, 303–310. [Google Scholar] [CrossRef]
- Abidi, N.; Cabrales, L.; Haigler, C.H. Changes in the cell wall and cellulose content of developing cotton fibers investigated by FTIR spectroscopy. Carbohydr. Polym. 2014, 100, 9–16. [Google Scholar] [CrossRef]
- Liu, Y.; He, Z.; Shankle, M.; Tewolde, H. Compositional features of cotton plant biomass fractions characterized by attenuated total reflection Fourier transform infrared spectroscopy. Ind. Crop. Prod. 2016, 79, 283–286. [Google Scholar] [CrossRef]
- Ling, Z.; Wang, T.; Makarem, M.; Cintrón, M.S.; Cheng, H.N.; Kang, X.; Bacher, M.; Potthast, A.; Rosenau, T.; King, H.; et al. Effects of ball milling on the structure of cotton cellulose. Cellulose 2019, 26, 305–328. [Google Scholar] [CrossRef]
- Nam, S.; Condon, B.D.; Liu, Y.; He, Q. Natural resistance of raw cotton fiber to heat evidenced by the suppressed depolymerization of cellulose. Polym. Degrad. Stab. 2017, 138, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Liu, Y.; Fang, D.D.; Delhom, C.D. Feasibility assessment of phenotyping cotton fiber maturity using infrared spectroscopy and algorithms for genotyping analyses. J. Cotton Res. 2019, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Campbell, B.T.; Delhom, C.; Martin, V. Comparative Relationship of Fiber Strength and Yarn Tenacity in Four Cotton Cultivars. J. Mater. Sci. Res. 2015, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Du, C.; Zhou, J. Structural and bonding environments derived from infrared spectroscopic studies. In Environmental Chemistry of Animal Manure; He, Z., Ed.; Nova Science Publishers: New York, NY, USA, 2011; pp. 23–42. [Google Scholar]
- Smidt, E.; Lechner, P.; Schwanninger, M.; Haberhauer, G.; Gerzabek, M.H. Characterization of Waste Organic Matter by FT-IR Spectroscopy: Application in Waste Science. Appl. Spectrosc. 2002, 56, 1170–1175. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Cheng, H.N.; Nam, S. Comparison of the wood bonding performance of water- and alkali-soluble cottonseed protein fractions. J. Adhes. Sci. Technol. 2020, 1–18. [Google Scholar] [CrossRef]
- Arslan, F.N.; Akin, G.; Elmas, Ş.N.K.; Yilmaz, I.; Janssen, H.-G.; Kenar, A. Rapid detection of authenticity and adulteration of cold pressed black cumin seed oil: A comparative study of ATR–FTIR spectroscopy and synchronous fluorescence with multivariate data analysis. Food Control. 2019, 98, 323–332. [Google Scholar] [CrossRef]
- Abidi, N.; Cabrales, L.; Hequet, E. Fourier transform infrared spectroscopic approach to the study of the secondary cell wall development in cotton fiber. Cellulose 2010, 17, 309–320. [Google Scholar] [CrossRef]
- Nelson, M.L.; O’Connor, R.T. Relation of certain infrared bands to cellulose crystallinity and crystal lattice type. Part II. A new infrared ratio for estimation of crystallinity in celluloses I and II. J. Appl. Polym. Sci. 1964, 8, 1325–1341. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Zhang, S.; Gao, Q.; Lu, Q.; Peng, R.; Xu, P.; Shang, H.; Yuan, Y.; Zou, H. Micro-FTIR combined with curve fitting method to study cellulose crystallinity of developing cotton fibers. Anal. Bioanal. Chem. 2021, 413, 1313–1320. [Google Scholar] [CrossRef]
- Abidi, N.; Hequet, E.; Cabrales, L.; Gannaway, J.; Wilkins, T.; Wells, L.W. Evaluating cell wall structure and composition of developing cotton fibers using Fourier transform infrared spectroscopy and thermogravimetric analysis. J. Appl. Polym. Sci. 2007, 107, 476–486. [Google Scholar] [CrossRef]
- Chen, L.; Carpita, N.C.; Reiter, W.-D.; Wilson, R.H.; Jeffries, C.; McCann, M.C. A rapid method to screen for cell-wall mutants using discriminant analysis of Fourier transform infrared spectra. Plant J. 1998, 16, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.M.; Kafle, K.; Belias, D.W.; Park, Y.B.; Glick, R.E.; Haigler, C.H.; Kim, S.H. Comprehensive analysis of cellulose content, crystallinity, and lateral packing in Gossypium hirsutum and Gossypium barbadense cotton fibers using sum frequency generation, infrared and Raman spectroscopy, and X-ray diffraction. Cellulose 2015, 22, 971–989. [Google Scholar] [CrossRef]
- Tian, C.M.; Shi, Z.H.; Zhang, H.Y.; Xu, J.Z.; Shi, J.R.; Guo, H.Z. Thermal Degradation of Cotton Cellulose. J. Therm. Anal. Calorim. 1999, 55, 93–98. [Google Scholar] [CrossRef]
- Carrillo, I.; Mendonça, R.T.; Ago, M.; Rojas, O.J. Comparative study of cellulosic components isolated from different Eucalyptus species. Cellulose 2018, 25, 1011–1029. [Google Scholar] [CrossRef]
- Mironova, M.; Makarov, I.; Golova, L.; Vinogradov, M.; Shandryuk, G.; Levin, I. Improvement in Carbonization Efficiency of Cellulosic Fibres Using Silylated Acetylene and Alkoxysilanes. Fibers 2019, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- He, Z.-Q.; Cao, X.-Y.; Mao, J.-D.; Ohno, T.; Waldrip, H. Analysis of Carbon Functional Groups in Mobile Humic Acid and Recalcitrant Calcium Humate Extracted from Eight US Soils. Pedosphere 2013, 23, 705–716. [Google Scholar] [CrossRef]
- Waldrip, H.M.; He, Z.; Todd, R.W.; Hunt, J.F.; Rhoades, M.B.; Cole, N.A. Characterization of Organic Matter in Beef Feedyard Manure by Ultraviolet-Visible and Fourier Transform Infrared Spectroscopies. J. Environ. Qual. 2014, 43, 690–700. [Google Scholar] [CrossRef]
- He, Z.; Guo, M.; Fortier, C.; Cao, X.; Schmidt-Rohr, K. Fourier Transform Infrared and Solid State 13C Nuclear Magnetic Resonance Spectroscopic Characterization of Defatted Cottonseed Meal-Based Biochars. Mod. Appl. Sci. 2021, 15, 108–121. [Google Scholar] [CrossRef]
- Liu, Y.; He, Z.; Uchimiya, M. Comparison of Biochar Formation from Various Agricultural By-Products Using FTIR Spectroscopy. Mod. Appl. Sci. 2014, 9, p246. [Google Scholar] [CrossRef]
- Abidi, N.; Manike, M. X-ray diffraction and FTIR investigations of cellulose deposition during cotton fiber development. Text. Res. J. 2018, 88, 719–730. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | WT | Li1 | liy | Li2-long | Li2-medium | Li2-short | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W | A | W | A | W | A | W | A | W | A | W | A | |
| C | 51.45 | 58.67 | 52.49 | 59.63 | 53.34 | 60.54 | 49.94 | 57.20 | 51.34 | 58.53 | 52.39 | 60.15 |
| O | 48.08 | 41.16 | 47.20 | 40.26 | 46.09 | 39.27 | 49.57 | 42.62 | 48.30 | 41.34 | 46.47 | 39.65 |
| K | 0.47 | 0.16 | 0.31 | 0.11 | 0.47 | 0.17 | 0.50 | 0.18 | 0.36 | 0.12 | 0.44 | 0.15 |
| Ca | ND 1 | ND | ND | ND | 0.10 | 0.03 | ND | ND | ND | ND | 0.16 | 0.05 |
| To (°C) | WLo (%) | Tp (°C) | WLp (%) | Dp(%/°C) | Te (°C) | WLe (%) | Char (%) C | |
|---|---|---|---|---|---|---|---|---|
| WT | 251.4 A (0.1) B | 5.8 A (0.5) | 335.9 D (0.4) | 47.5 C (1.0) | 1.256 BC (0.031) | 360.4 C (0.1) | 69.4 C (0.5) | 19.6 C (0.5) |
| Li1 | 248.5 A (3.8) | 6.1 A (0.4) | 341.2 A (0.4) | 47.5 C (0.8) | 1.217 C (0.007) | 363.1 B (0.785) | 68.1 D (0.6) | 21.1 A (0.1) |
| Li2-long | 249.1 A (0.6) | 5.6 A (0.1) | 339.6 B (0.1) | 51.3 B (0.5) | 1.328 A (0.014) | 360.3 C (0.2) | 71.4 B (0.1) | 18.3 D (0.3) |
| Li2-mix | 252.3 A (1.3) | 5.8 A (0.1) | 338.2 C (0.2) | 49.8 B (0.4) | 1.288 AB (0.010) | 358.6 D (0.4) | 69.3 C (0.3) | 20.4 B (0.1) |
| Li2-short | 209.7 B (0.2) | 5.4 A (0.3) | 340.9 A (0.7) | 54.7 A (0.6) | 1.066 D (0.035) | 366.6 A (0.9) | 74.2 A (0.4) | 16.8 e (0.1) |
| liy | 210.5 B (0.5) | 4.5 B (0.1) | 331.4 e (0.4) | 54.3 A (0.1) | 1.206 C (0.008) | 353.8 e (0.1) | 74.2 A (0.2) | 16.7 e (0.1) |
| PC1 | I2918 | I2849 | I740 | Maturity | CI | |
|---|---|---|---|---|---|---|
| T0 | −0.790 | −0.571 | 0.664 | −0.415 | −0.942 ** | 0.767 |
| WL0 | −0.889 * | −0.087 | 0.761 | 0.104 | −0.875 * | 0.485 |
| Tp | −0.596 | 0.386 | 0.631 | 0.601 | −0.478 | 0.001 |
| WLp | 0.690 | 0.354 | −0.689 | 0.286 | 0.886 * | −0.575 |
| Dp | −0.352 | −0.766 | 0.354 | −0.680 | −0.620 | 0.680 |
| Te | 0.178 | 0.586 | 0.236 | 0.697 | 0.410 | −0.452 |
| WLe | 0.780 | 0.303 | −0.782 | 0.182 | 0.866 * | −0.606 |
| Char | −0.791 | −0.219 | 0.804 | −0.084 | −0.793 | 0.573 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Nam, S.; Fang, D.D.; Cheng, H.N.; He, J. Surface and Thermal Characterization of Cotton Fibers of Phenotypes Differing in Fiber Length. Polymers 2021, 13, 994. https://doi.org/10.3390/polym13070994
He Z, Nam S, Fang DD, Cheng HN, He J. Surface and Thermal Characterization of Cotton Fibers of Phenotypes Differing in Fiber Length. Polymers. 2021; 13(7):994. https://doi.org/10.3390/polym13070994
Chicago/Turabian StyleHe, Zhongqi, Sunghyun Nam, David D. Fang, Huai N. Cheng, and Jibao He. 2021. "Surface and Thermal Characterization of Cotton Fibers of Phenotypes Differing in Fiber Length" Polymers 13, no. 7: 994. https://doi.org/10.3390/polym13070994
APA StyleHe, Z., Nam, S., Fang, D. D., Cheng, H. N., & He, J. (2021). Surface and Thermal Characterization of Cotton Fibers of Phenotypes Differing in Fiber Length. Polymers, 13(7), 994. https://doi.org/10.3390/polym13070994