
Refining the Design of Diblock Elastin-Like Polypeptides for Self-Assembly into Nanoparticles

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Production of ELP-[M1V3-i] (i = 20, 40, 60, 80, 100) Monoblocks
2.3. Production and Post-Modification of ELP-[I-j] (j = 20, 40, 60) Monoblocks
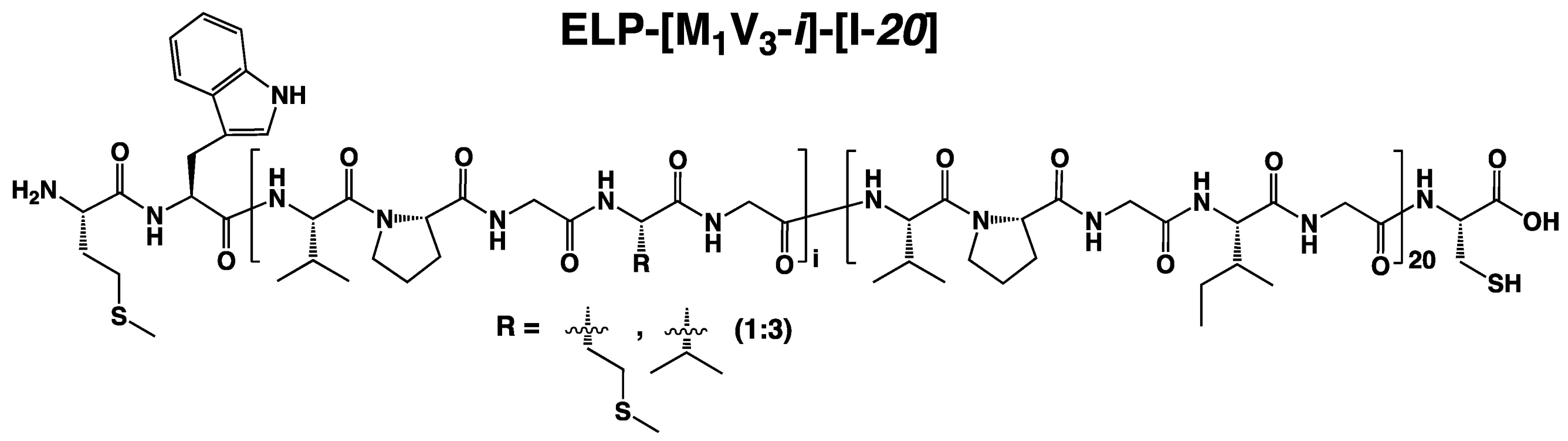
2.4. Production and Post-Modification of ELP-[M1V3-i]-[I-20] (i = 20, 40, 60)
2.4.1. Construction of the Expression Vector ELP-[M1V3-i]-[I-20] (i = 20, 40, 60)
2.4.2. Production
2.4.3. Purification
2.4.4. NEM Capping of Cysteine C-Terminal
2.5. Production and Post-Modification of ELP-[M1V3-i]-[I-90], (i = 40, 60)
2.5.1. Construction of the Expression Vector ELP-[M1V3-i]-[I-90] (i = 40, 60)
2.5.2. Production of ELP-[M1V3-i]-[I-90], (i = 40, 60)
2.5.3. Purification
2.6. Chemical Oxidation of Diblock ELPs
2.7. Characterization Techniques
2.7.1. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.7.2. Mass Spectrometry Analysis
2.7.3. Nuclear Magnetic Resonance (NMR)
2.7.4. Size-Exclusion Chromatography (SEC)
2.7.5. Turbidity Analysis
2.7.6. Dynamic Light Scattering
3. Results and Discussion
3.1. Development and Thermal Study of the First Series of Diblock ELPs: ELP-[M1V3-i]-[I-20] (i = 20, 40, 60)
3.1.1. Production of Diblock ELPs and of a Library of Related Monoblock ELPs
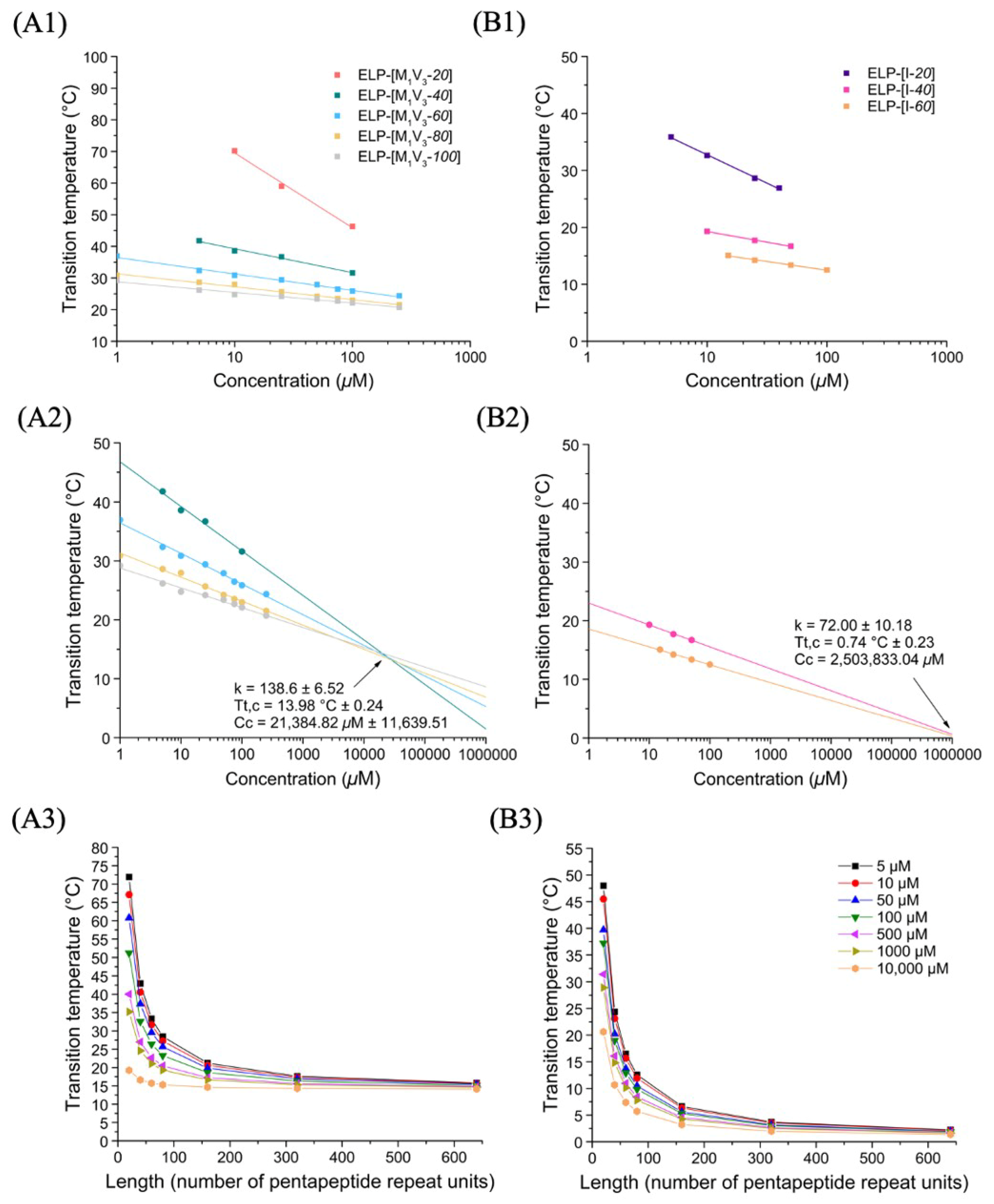
3.1.2. Thermal Properties of ELP-[M1V3-i]-[I-20] Diblocks and of the Monoblock Library
Thermal Behavior of Monoblock ELPs
Thermal Behavior of ELP-[M1V3-i]-[I-20] Diblocks
3.2. Development and Thermal Study of the Second Series of Diblock ELPs: ELP-[*M1V3-i]-[I-20] (i = 20, 40, 60)
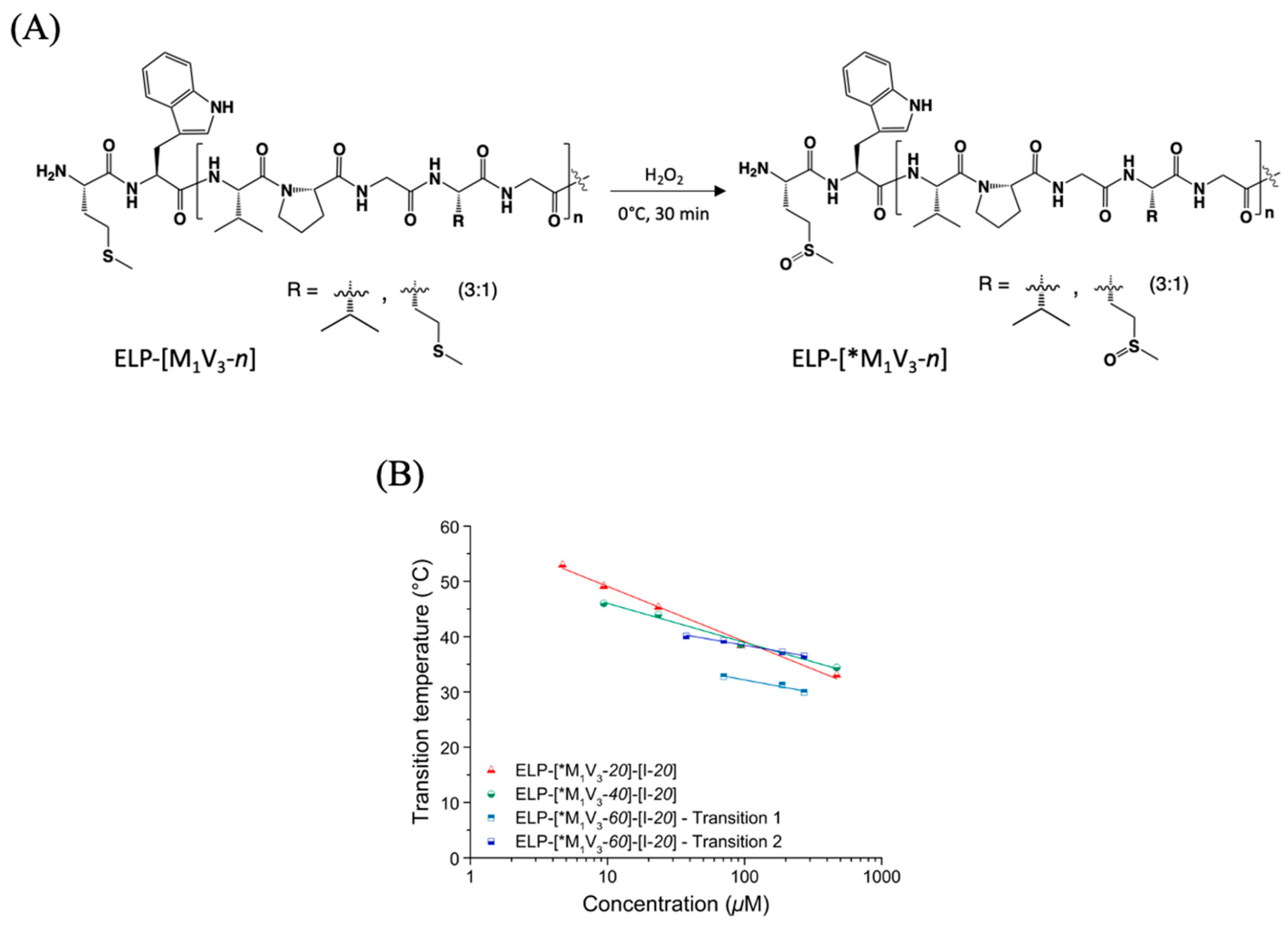
3.2.1. Chemical Oxidation at Methionine Residues of Diblock ELPs
3.2.2. Thermal Properties of Oxidized Diblock ELPs
3.3. Development and Thermal Study of the Third Series of Diblock ELPs: ELP-[*M1V3-i]-[I-90] (i = 40, 60)
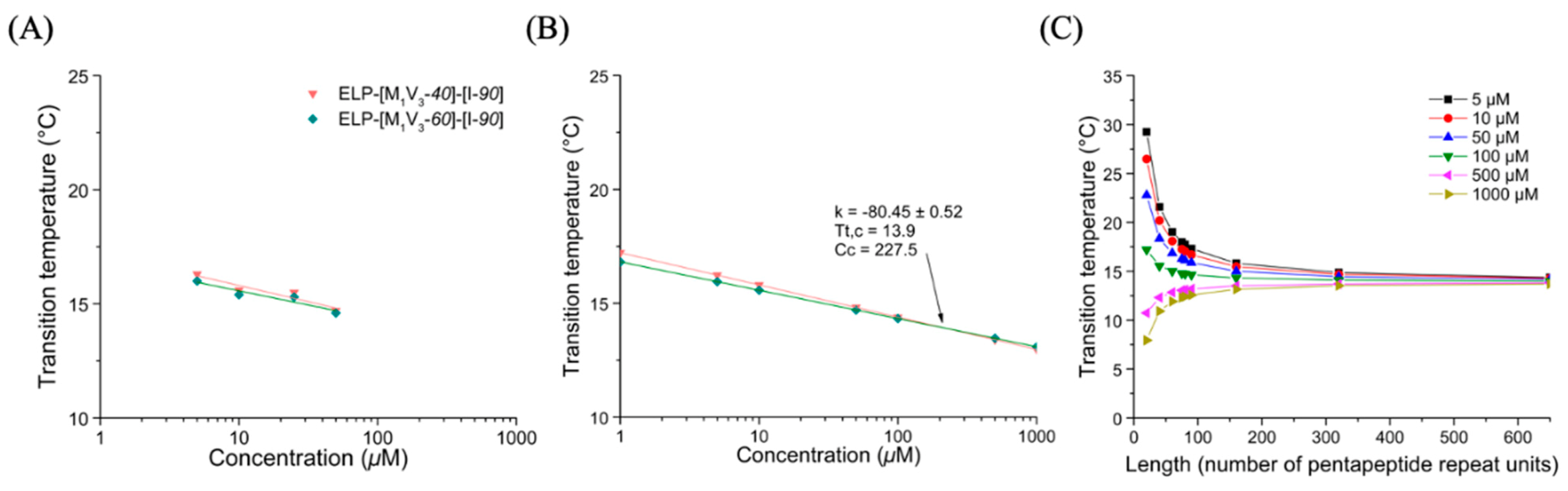
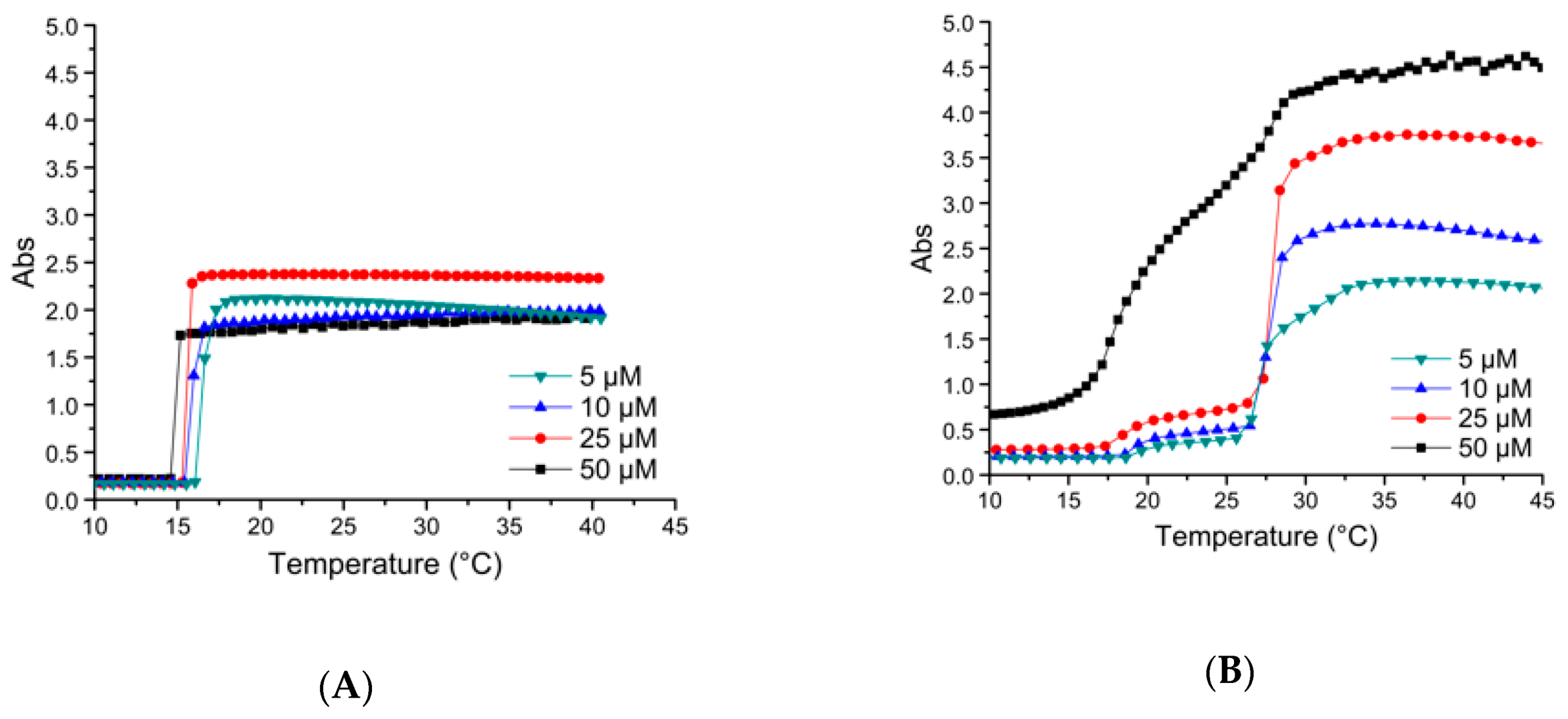
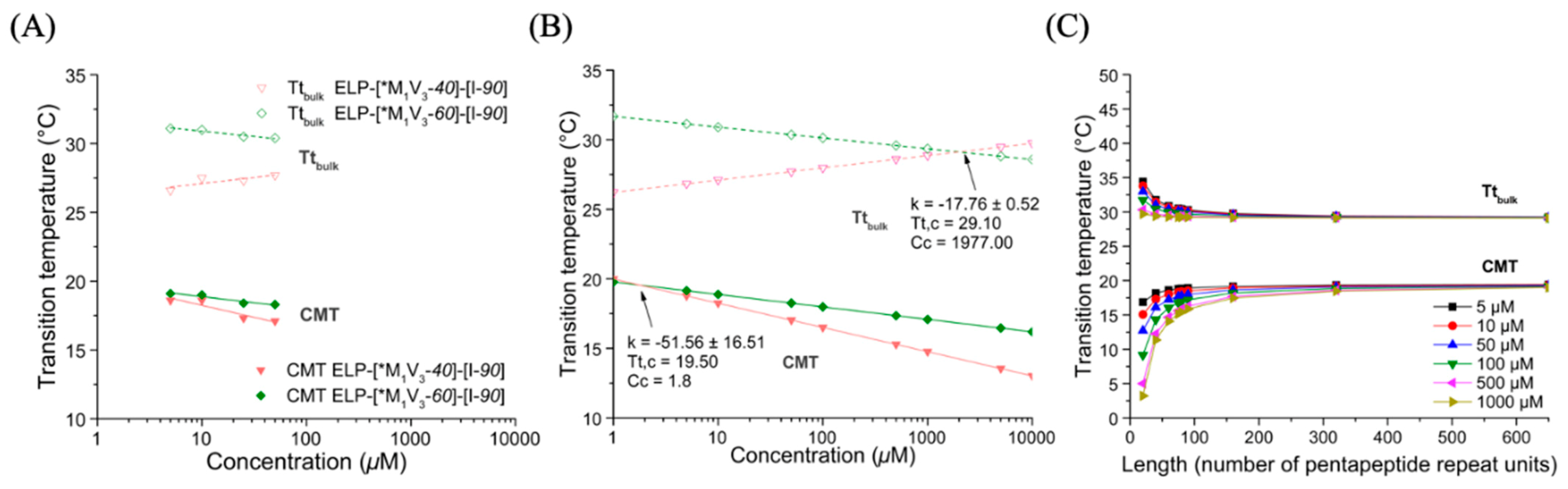
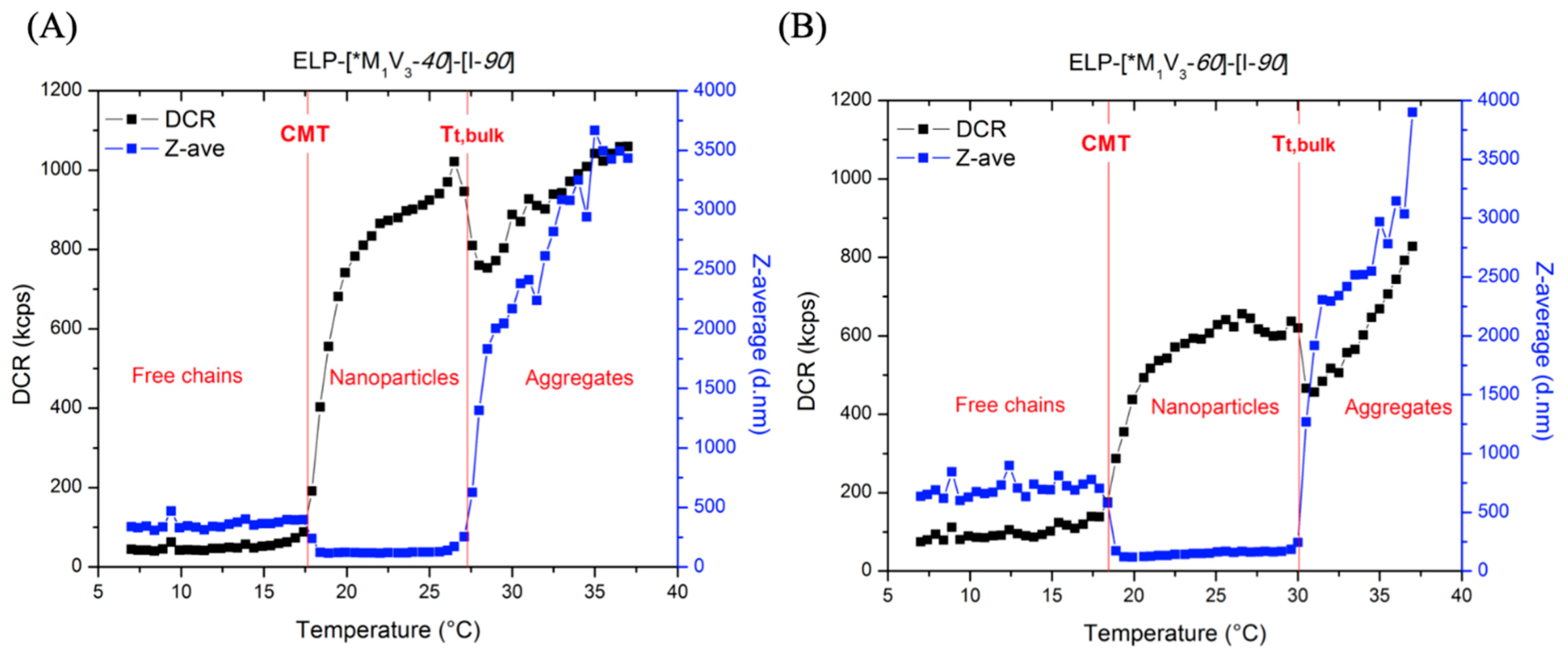
3.3.1. Thermal Properties of ELP-[M1V3-i]-[I-90] Diblock ELPs (I = 40, 60)
3.3.2. Chemoselective Modification of ELP-[*M1V3-i]-[I-90] Methionine Residues
3.3.3. Thermal Properties of Oxidized Diblocks ELP-[*M1V3-i]-[I-90]
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ward, M.A.; Georgiou, T.K. Thermoresponsive polymers for biomedical applications. Polymers 2011, 3, 1215–1242. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Matsunaga, Y.T. Thermo-responsive polymers and their application as smart biomaterials. J. Mater. Chem. B 2017, 5, 4307–4321. [Google Scholar] [CrossRef]
- Bordat, A.; Boissenot, T.; Nicolas, J.; Tsapis, N. Thermoresponsive polymer nanocarriers for biomedical applications. Adv. Drug Deliv. Rev. 2019, 138, 167–192. [Google Scholar] [CrossRef] [PubMed]
- Matanović, M.R.; Kristl, J.; Grabnar, P.A. Thermoresponsive polymers: Insights into decisive hydrogel characteristics, mechanisms of gelation, and promising biomedical applications. Int. J. Pharm. 2014, 472, 262–275. [Google Scholar] [CrossRef]
- Sponchioni, M.; Capasso Palmiero, U.; Moscatelli, D. Thermo-responsive polymers: Applications of smart materials in drug delivery and tissue engineering. Mater. Sci. Eng. C 2019, 102, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Sarwan, T.; Kumar, P.; Choonara, Y.E.; Pillay, V. Hybrid Thermo-Responsive Polymer Systems and Their Biomedical Applications. Front. Mater. 2020, 7. [Google Scholar] [CrossRef]
- Karimi, M.; Sahandi Zangabad, P.; Ghasemi, A.; Amiri, M.; Bahrami, M.; Malekzad, H.; Ghahramanzadeh Asl, H.; Mahdieh, Z.; Bozorgomid, M.; Ghasemi, A.; et al. Temperature-Responsive Smart Nanocarriers for Delivery of Therapeutic Agents: Applications and Recent Advances. ACS Appl. Mater. Interfaces 2016, 8, 21107–21133. [Google Scholar] [CrossRef] [Green Version]
- Seuring, J.; Agarwal, S. Polymers with upper critical solution temperature in aqueous solution. Macromol. Rapid Commun. 2012, 33, 1898–1920. [Google Scholar] [CrossRef]
- Qi, J.; Li, W.; Lu, K.; Jin, F.; Liu, D.; Xu, X.; Wang, X.; Kang, X.; Wang, W.; Shu, G.; et al. PH and Thermal Dual-Sensitive Nanoparticle-Mediated Synergistic Antitumor Effect of Immunotherapy and Microwave Thermotherapy. Nano Lett. 2019, 19, 4949–4959. [Google Scholar] [CrossRef]
- Halperin, A.; Kröger, M.; Winnik, F.M. Poly(N-isopropylacrylamide) Phase Diagrams: Fifty Years of Research. Angew. Chemie Int. Ed. 2015, 54, 15342–15367. [Google Scholar] [CrossRef]
- Vancoillie, G.; Frank, D.; Hoogenboom, R. Thermoresponsive poly(oligo ethylene glycol acrylates). Prog. Polym. Sci. 2014, 39, 1074–1095. [Google Scholar] [CrossRef]
- Hoogenboom, R.; Schlaad, H. Thermoresponsive poly(2-oxazoline)s, polypeptoids, and polypeptides. Polym. Chem. 2017, 8, 24–40. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Li, Z. Peptoid Applications in Biomedicine and Nanotechnology; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; ISBN 9780081007426. [Google Scholar]
- Winninger, J.; Iurea, D.M.; Atanase, L.I.; Salhi, S.; Delaite, C.; Riess, G. Micellization of novel biocompatible thermo-sensitive graft copolymers based on poly(ε-caprolactone), poly(N-vinylcaprolactam) and poly(N-vinylpyrrolidone). Eur. Polym. J. 2019, 119, 74–82. [Google Scholar] [CrossRef]
- Bonacucina, G.; Cespi, M.; Mencarelli, G.; Giorgioni, G.; Palmieri, G.F. Thermosensitive self-assembling block copolymers as drug delivery systems. Polymers 2011, 3, 779–811. [Google Scholar] [CrossRef] [Green Version]
- Atanase, L.I. Micellar Drug Delivery Systems Based on Natural Biopolymers. Polymers 2021, 13, 477. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cabello, J.C.; Martín, L.; Alonso, M.; Arias, F.J.; Testera, A.M. “Recombinamers” as advanced materials for the post-oil age. Polymer (Guildf) 2009, 50, 5159–5169. [Google Scholar] [CrossRef] [Green Version]
- Urry, D.W. Physical chemistry of biological free energy transduction as demonstrated by elastic protein-based polymers. J. Phys. Chem. B 1997, 101, 11007–11028. [Google Scholar] [CrossRef]
- Dzuricky, M.; Roberts, S.; Chilkoti, A. Convergence of Artificial Protein Polymers and Intrinsically Disordered Proteins. Biochemistry 2018, 57, 2405–2414. [Google Scholar] [CrossRef]
- Urry, D.W.; Long, M.M.; Cox, B.A.; Ohnishi, T.; Mitchell, L.W.; Jacobs, M. The synthetic polypentapeptide of elastin coacervates and forms filamentous aggregates. Biochim. Biophys. Acta Protein Struct. 1974, 371, 597–602. [Google Scholar] [CrossRef]
- Urry, D.W.; Ohnishi, T. Studies on the conformations and interactions of elastin. Proton magnetic resonance of the repeating tetramer. Biopolymers 1974, 13, 1223–1242. [Google Scholar] [CrossRef]
- Urry, D.W.; Gowda, D.C.; Parker, T.M.; Luan, C.-H.; Reid, M.C.; Harris, C.M.; Pattanaik, A.; Harris, R.D. Hydrophobicity scale for proteins based on inverse temperature transitions. Biopolymers 1992, 32, 1243–1250. [Google Scholar] [CrossRef]
- Urry, D.W. Free energy transduction in polypeptides and proteins based on inverse temperature transitions. Prog. Biophys. Mol. Biol. 1992, 57, 23–57. [Google Scholar] [CrossRef]
- Mcpherson, D.T.; Morrow, C.; Minehan, D.S.; Wu, J.; Hunter, E.; Urry, D.W. Production and Purification of a Recombinant Elastomeric Polypeptide, G-(VPGVG)19-VPGV, from Escherichia coli. Biotechnol. Prog. 1992, 8, 347–352. [Google Scholar] [CrossRef]
- Meyer, D.E.; Chilkoti, A. Genetically Encoded Synthesis of Protein-Based Polymers with Precisely Specified Molecular Weight and Sequence by Recursive Directional Ligation: Examples from the Elastin-like Polypeptide System. Biomacromolecules 2002, 3, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.; Fernández-Colino, A.; López, I.M.; Rodríguez-Cabello, J.C.; Arias, F.J. Elastin-like recombinamers: Biosynthetic strategies and biotechnological applications. Biotechnol. J. 2011, 6, 1174–1186. [Google Scholar] [CrossRef]
- Meyer, D.E.; Chilkoti, A. Quantification of the effects of chain length and concentration on the thermal behavior of elastin-like polypeptides. Biomacromolecules 2004, 5, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, J.; Bellucci, J.J.; Weitzhandler, I.; McDaniel, J.R.; Spasojevic, I.; Li, X.; Lin, C.-C.; Chi, J.-T.A.; Chilkoti, A. A paclitaxel-loaded recombinant polypeptide nanoparticle outperforms Abraxane in multiple murine cancer models. Nat. Commun. 2015, 6, 7939. [Google Scholar] [CrossRef] [PubMed]
- Luginbuhl, K.M.; Schaal, J.L.; Umstead, B.; Mastria, E.M.; Li, X.; Banskota, S.; Arnold, S.; Feinglos, M.; D’Alessio, D.; Chilkoti, A. One-week glucose control via zero-order release kinetics from an injectable depot of glucagon-like peptide-1 fused to a thermosensitive biopolymer. Nat. Biomed. Eng. 2017, 1. [Google Scholar] [CrossRef]
- Schaal, J.L.; Li, X.; Mastria, E.; Bhattacharyya, J.; Zalutsky, M.R.; Chilkoti, A.; Liu, W. Injectable polypeptide micelles that form radiation crosslinked hydrogels in situ for intratumoral radiotherapy. J. Control. Release 2016, 228, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pille, J.; van Lith, S.A.M.; van Hest, J.C.M.; Leenders, W.P.J. Self-Assembling VHH-Elastin-Like Peptides for Photodynamic Nanomedicine. Biomacromolecules 2017, 18, 1302–1310. [Google Scholar] [CrossRef]
- MacEwan, S.R.; Weitzhandler, I.; Hoffmann, I.; Genzer, J.; Gradzielski, M.; Chilkoti, A. Phase Behavior and Self-Assembly of Perfectly Sequence-Defined and Monodisperse Multiblock Copolypeptides. Biomacromolecules 2017, 18, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Georgilis, E.; Abdelghani, M.; Pille, J.; Aydinlioglu, E.; van Hest, J.C.M.; Lecommandoux, S.; Garanger, E. Nanoparticles based on natural, engineered or synthetic proteins and polypeptides for drug delivery applications. Int. J. Pharm. 2020, 586. [Google Scholar] [CrossRef]
- Deng, C.; Wu, J.; Cheng, R.; Meng, F.; Klok, H.A.; Zhong, Z. Functional polypeptide and hybrid materials: Precision synthesis via α-amino acid N-carboxyanhydride polymerization and emerging biomedical applications. Prog. Polym. Sci. 2014, 39, 330–364. [Google Scholar] [CrossRef]
- Kim, W.; Chaikof, E.L. Recombinant elastin-mimetic biomaterials: Emerging applications in medicine. Adv. Drug Deliv. Rev. 2010, 62, 1468–1478. [Google Scholar] [CrossRef] [Green Version]
- MacEwan, S.R.; Chilkoti, A. Elastin-like polypeptides: Biomedical applications of tunable biopolymers. Biopolymers 2010, 94, 60–77. [Google Scholar] [CrossRef]
- Nettles, D.L.; Chilkoti, A.; Setton, L.A. Applications of elastin-like polypeptides in tissue engineering. Adv. Drug Deliv. Rev. 2010, 62, 1479–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janib, S.M.; Pastuszka, M.F.; Aluri, S.; Folchman-Wagner, Z.; Hsueh, P.Y.; Shi, P.; Lin, Y.A.; Cui, H.; MacKay, J.A. A quantitative recipe for engineering protein polymer nanoparticles. Polym. Chem. 2014, 5, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Garanger, E.; MacEwan, S.R.; Sandre, O.; Bataille, L.; Chilkoti, A. Structural Evolution of a Stimulus-Responsive Diblock Polypeptide Micelle by Temperature Tunable Compaction of its Core. Macromolecules 2015. [Google Scholar] [CrossRef] [Green Version]
- Hassouneh, W.; Zhulina, E.B.; Chilkoti, A.; Rubinstein, M. Elastin-like Polypeptide Diblock Copolymers Self-Assemble into Weak Micelles. Macromolecules 2015, 48, 4183–4195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petitdemange, R.; Garanger, E.; Bataille, L.; Bathany, K.; Garbay, B.; Deming, T.J.; Lecommandoux, S. Tuning Thermoresponsive Properties of Cationic Elastin-like Polypeptides by Varying Counterions and Side-Chains. Bioconjug. Chem. 2017, 28, 1403–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, J.R.; Petitdemange, R.; Bataille, L.; Bathany, K.; Wirotius, A.-L.L.; Garbay, B.; Deming, T.J.; Garanger, E.; Lecommandoux, S. Quantitative Side-Chain Modifications of Methionine-Containing Elastin-Like Polypeptides as a Versatile Tool to Tune Their Properties. ACS Macro Lett. 2015, 4, 1283–1286. [Google Scholar] [CrossRef] [Green Version]
- Bataille, L.; Dieryck, W.; Hocquellet, A.; Cabanne, C.; Bathany, K.; Lecommandoux, S.; Garbay, B.; Garanger, E. Recombinant production and purification of short hydrophobic Elastin-like polypeptides with low transition temperatures. Protein Expr. Purif. 2016, 121, 81–87. [Google Scholar] [CrossRef]
- Dai, M.; Goudounet, G.; Zhao, H.; Garbay, B.; Garanger, E.; Pecastaings, G.; Schultze, X.; Lecommandoux, S. Thermosensitive Hybrid Elastin-like Polypeptide-Based ABC Triblock Hydrogel. Macromolecules 2020. [Google Scholar] [CrossRef]
- Meyer, D.E.; Chilkoti, A. Protein Purification by Inverse Transition Cycling. In Protein–Protein Interactions: AMolecular Cloning Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2002; Chapter 18; pp. 329–344. [Google Scholar]
- Meyer, D.E.; Chilkoti, A. Purification of recombinant proteins by fusion with thermally- responsive polypeptides. Nat. Biotechnol. 1999, 17, 1112–1115. [Google Scholar] [CrossRef]
- Rosselin, M.; Xiao, Y.; Belhomme, L.; Lecommandoux, S.; Garanger, E. Expanding the Toolbox of Chemoselective Modifications of Protein-Like Polymers at Methionine Residues. ACS Macro Lett. 2019, 8, 1648–1653. [Google Scholar] [CrossRef]
- Rosselin, M.; Chinoy, Z.S.; Bravo-Anaya, L.M.; Lecommandoux, S.; Garanger, E. Multivalent Elastin-Like Glycopolypeptides: Subtle Chemical Structure Modifications with High Impact on Lectin Binding Affinity. ACS Macro Lett. 2021, 65–70. [Google Scholar] [CrossRef]
- Petitdemange, R.; Garanger, E.; Bataille, L.; Dieryck, W.; Bathany, K.; Garbay, B.; Deming, T.J.; Lecommandoux, S. Selective Tuning of Elastin-like Polypeptide Properties via Methionine Oxidation. Biomacromolecules 2017, 18, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Pinedo-Martín, G.; Castro, E.; Martín, L.; Alonso, M.; Rodríguez-Cabello, J.C. Effect of surfactants on the self-Assembly of a model elastin-like block corecombinamer: From micelles to an aqueous two-phase system. Langmuir 2014, 30, 3432–3440. [Google Scholar] [CrossRef]
- Martín, L.; Castro, E.; Ribeiro, A.; Alonso, M.; Rodríguez-Cabello, J.C. Temperature-triggered self-assembly of elastin-like block co-recombinamers: The controlled formation of micelles and vesicles in an aqueous medium. Biomacromolecules 2012, 13, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Dreher, M.R.; Simnick, A.J.; Fischer, K.; Smith, R.J.; Patel, A.; Schmidt, M.; Chilkoti, A. Temperature triggered self-assembly of polypeptides into multivalent spherical micelles. J. Am. Chem. Soc. 2008, 130, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Dreher, M.R.; Liu, W.; Michelich, C.R.; Dewhirst, M.W.; Chilkoti, A. Thermal cycling enhances the accumulation of a temperature-sensitive biopolymer in solid tumors. Cancer Res. 2007, 67, 4418–4424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukerji, R.; Schaal, J.; Li, X.; Bhattacharyya, J.; Asai, D.; Zalutsky, M.R.; Chilkoti, A.; Liu, W. Spatiotemporally photoradiation-controlled intratumoral depot for combination of brachytherapy and photodynamic therapy for solid tumor. Biomaterials 2016, 79, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ELP Notation | Amino Acid Sequence | Theoretical MW (Da) | Experimental a MW (Da) |
|---|---|---|---|
| Monoblock library | |||
| ELP-[M1V3-20] | MW[VPGVGVPGMG(VPGVG)2]5 | 8685 | 8686 |
| ELP-[M1V3-40] | MW[VPGVGVPGMG(VPGVG)2]10 | 17,035 | 17,035 |
| ELP-[M1V3-60] | MW[VPGVGVPGMG(VPGVG)2]15 | 25,385 | 25,387 |
| ELP-[M1V3-80] | MW[VPGVGVPGMG(VPGVG)2]20 | 33,735 | 33,737 |
| ELP-[M1V3-100] | MW[VPGVGVPGMG(VPGVG)2]25 | 42,085 | 42,088 |
| ELP-[I-20] | MW[VPGIG]20C(N-EtSucc) b | 9034 | 9034 |
| ELP-[I-40] | MW[VPGIG]40C(N-EtSucc) b | 17,504 | 17,504 |
| ELP-[I-60] | MW[VPGIG]60C(N-EtSucc) b | 25,974 | 25,974 |
| Diblock library | |||
| ELP-[M1V3-20]-[I-20] | MW[VPGVGVPGMG(VPGVG)2]5[VPGIG]20C(N-EtSucc) b | 17,384 | n.d. c |
| ELP-[M1V3-40]-[I-20] | MW[VPGVGVPGMG(VPGVG)2]10[VPGIG]20C(N-EtSucc) b | 25,734 | n.d. c |
| ELP-[M1V3-60]-[I-20] | MW[VPGVGVPGMG(VPGVG)2]15[VPGIG]20C(N-EtSucc) b | 34,084 | n.d. c |
| ELP-[*M1V3-20]-[I-20] d | M(O)W[VPGVGVPGM(O)G(VPGVG)2]5[IGVPG]20C(N-EtSucc) b,d | 17,480 | n.d. e |
| ELP-[*M1V3-40]-[I-20] d | M(O)W[VPGVGVPGM(O)G(VPGVG)2]10[VPGIG]20C(N-EtSucc) b,d | 25,910 | n.d. e |
| ELP-[*M1V3-60]-[I-20] d | M(O)W[VPGVGVPGM(O)G(VPGVG)2]15[VPGIG]20C(N-EtSucc) b,d | 34,340 | n.d. e |
| ELP-[M1V3-40]-[I-90] | MW[VPGVGVPGMG(VPGVG)2]10[IGVPG]90Y | 55,315 | 55,314 |
| ELP-[M1V3-60]-[I-90] | MW[VPGVGVPGMG(VPGVG)2]15[IGVPG]90Y | 63,665 | 63,664 |
| ELP-[*M1V3-40]-[I-90] d | M(O)W[VPGVGVPGM(O)G(VPGVG)2]10[IGVPG]90Y | 55,491 | 55,523 f |
| ELP-[*M1V3-60]-[I-90] d | M(O)W[VPGVGVPGM(O)G(VPGVG)2]15[IGVPG]90Y | 63,921 | 63,949 f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, M.; Georgilis, E.; Goudounet, G.; Garbay, B.; Pille, J.; van Hest, J.C.M.; Schultze, X.; Garanger, E.; Lecommandoux, S. Refining the Design of Diblock Elastin-Like Polypeptides for Self-Assembly into Nanoparticles. Polymers 2021, 13, 1470. https://doi.org/10.3390/polym13091470
Dai M, Georgilis E, Goudounet G, Garbay B, Pille J, van Hest JCM, Schultze X, Garanger E, Lecommandoux S. Refining the Design of Diblock Elastin-Like Polypeptides for Self-Assembly into Nanoparticles. Polymers. 2021; 13(9):1470. https://doi.org/10.3390/polym13091470
Chicago/Turabian StyleDai, Michèle, Evangelos Georgilis, Guillaume Goudounet, Bertrand Garbay, Jan Pille, Jan C. M. van Hest, Xavier Schultze, Elisabeth Garanger, and Sébastien Lecommandoux. 2021. "Refining the Design of Diblock Elastin-Like Polypeptides for Self-Assembly into Nanoparticles" Polymers 13, no. 9: 1470. https://doi.org/10.3390/polym13091470
APA StyleDai, M., Georgilis, E., Goudounet, G., Garbay, B., Pille, J., van Hest, J. C. M., Schultze, X., Garanger, E., & Lecommandoux, S. (2021). Refining the Design of Diblock Elastin-Like Polypeptides for Self-Assembly into Nanoparticles. Polymers, 13(9), 1470. https://doi.org/10.3390/polym13091470