Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean

Abstract
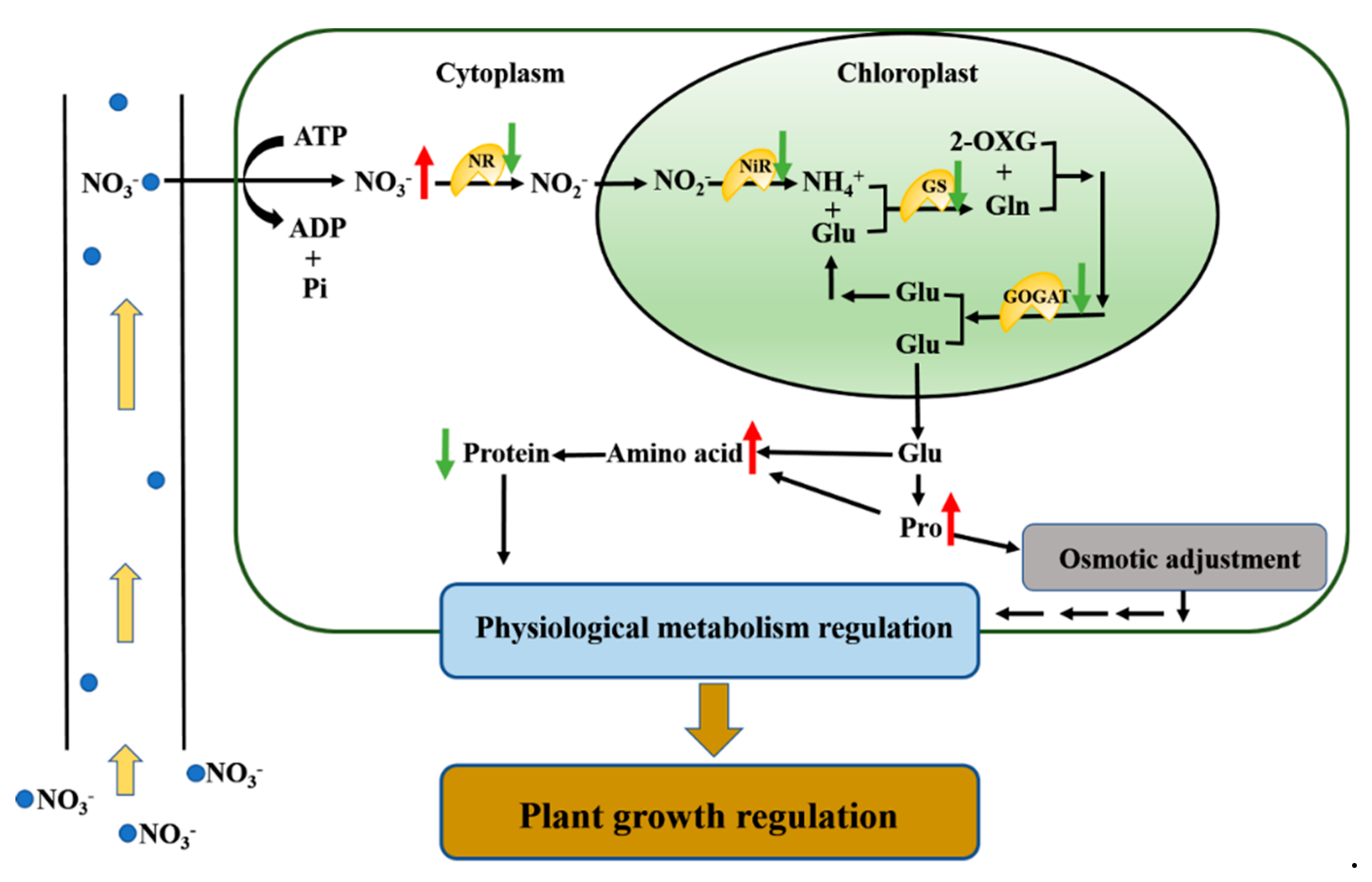
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatment
2.2. Sampling
2.3. Agronomic Traits Measurement
2.4. Measurement of Nitrogen and Nitrate Concentrations
2.5. Measurement of Soluble Protein, Free Amino Acid, Proline, and Soluble Sugar Concentrations
2.6. N-Metabolizing Enzymes Extraction and Analysis
2.7. Antioxidant Enzymes Activities and MDA Concentration
2.8. RNA Extraction and Complementary DNA Synthesis
2.9. Gene Expression by qRT-PCR
2.10. Statistical Analysis
3. Results
3.1. Effect of Drought Stress on Dry Matter Accumulation and Seed Yield
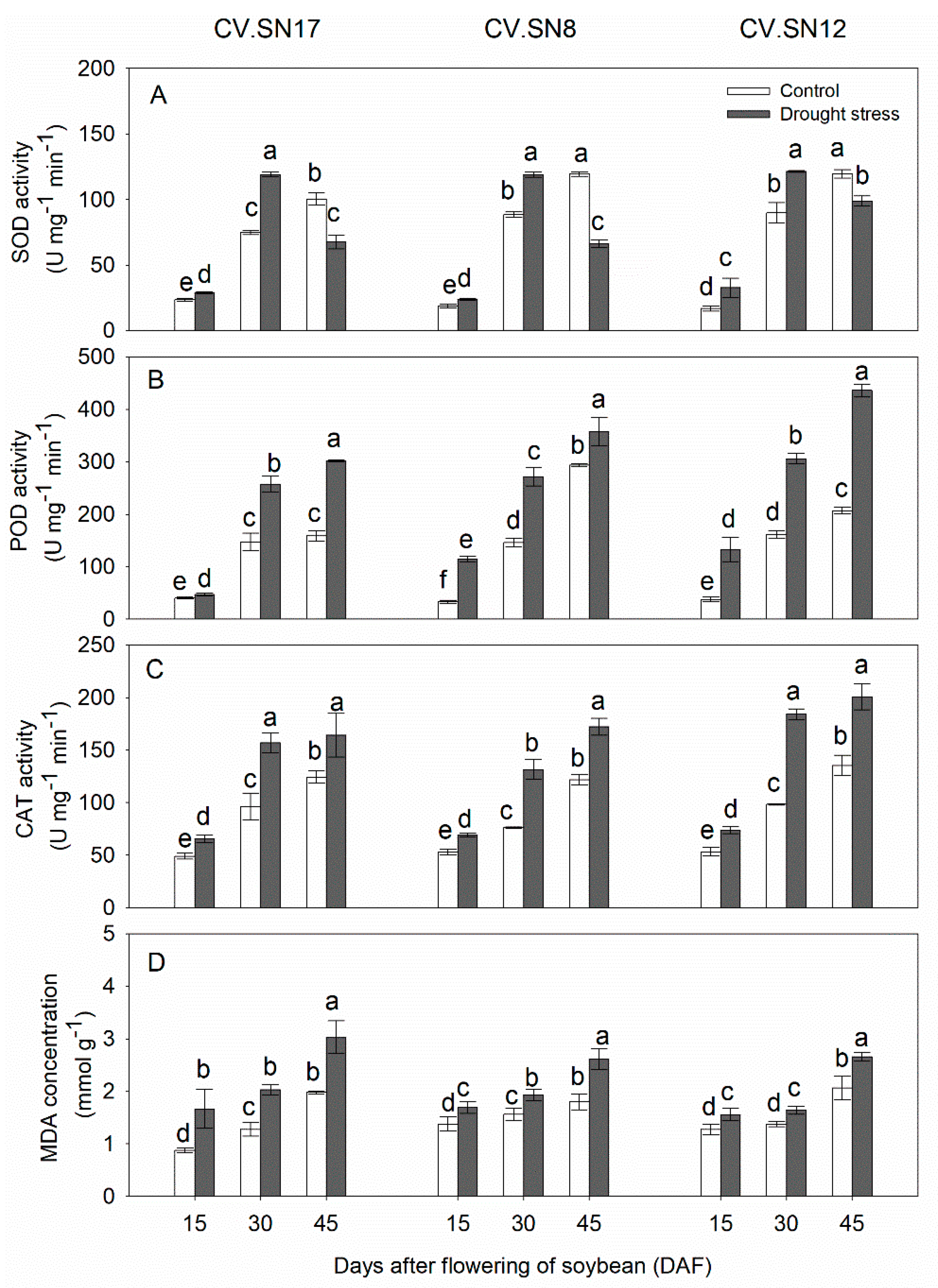
3.2. Effect of Drought Stress on Antioxidant Enzymes Activities and MDA Contents in Leaves
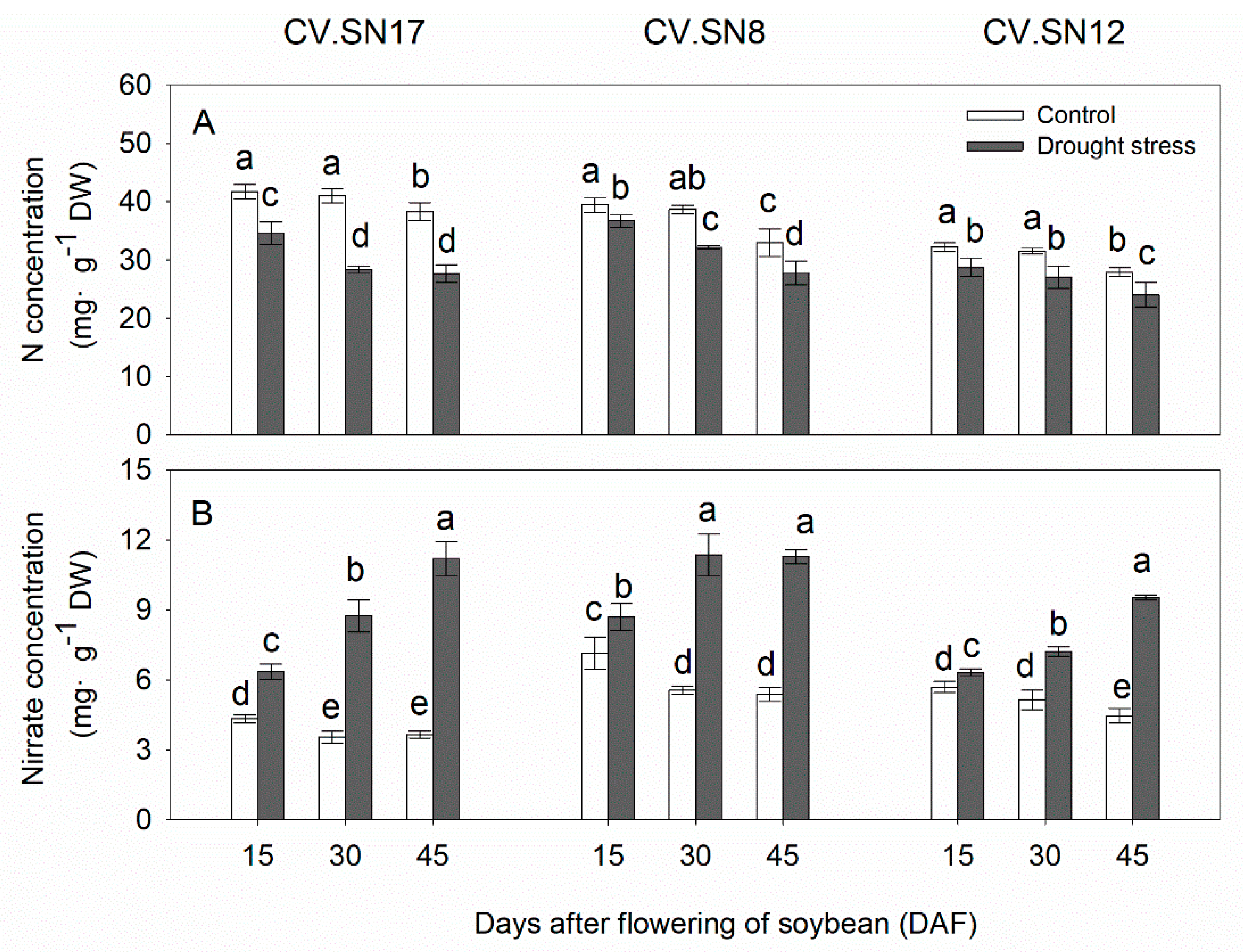
3.3. Effect of Drought Stress on Nitrogen and Nitrate Contents in Leaves
3.4. Effect of Drought Stress on the Activities of Enzymes Related to Nitrogen Metabolism in Leaves
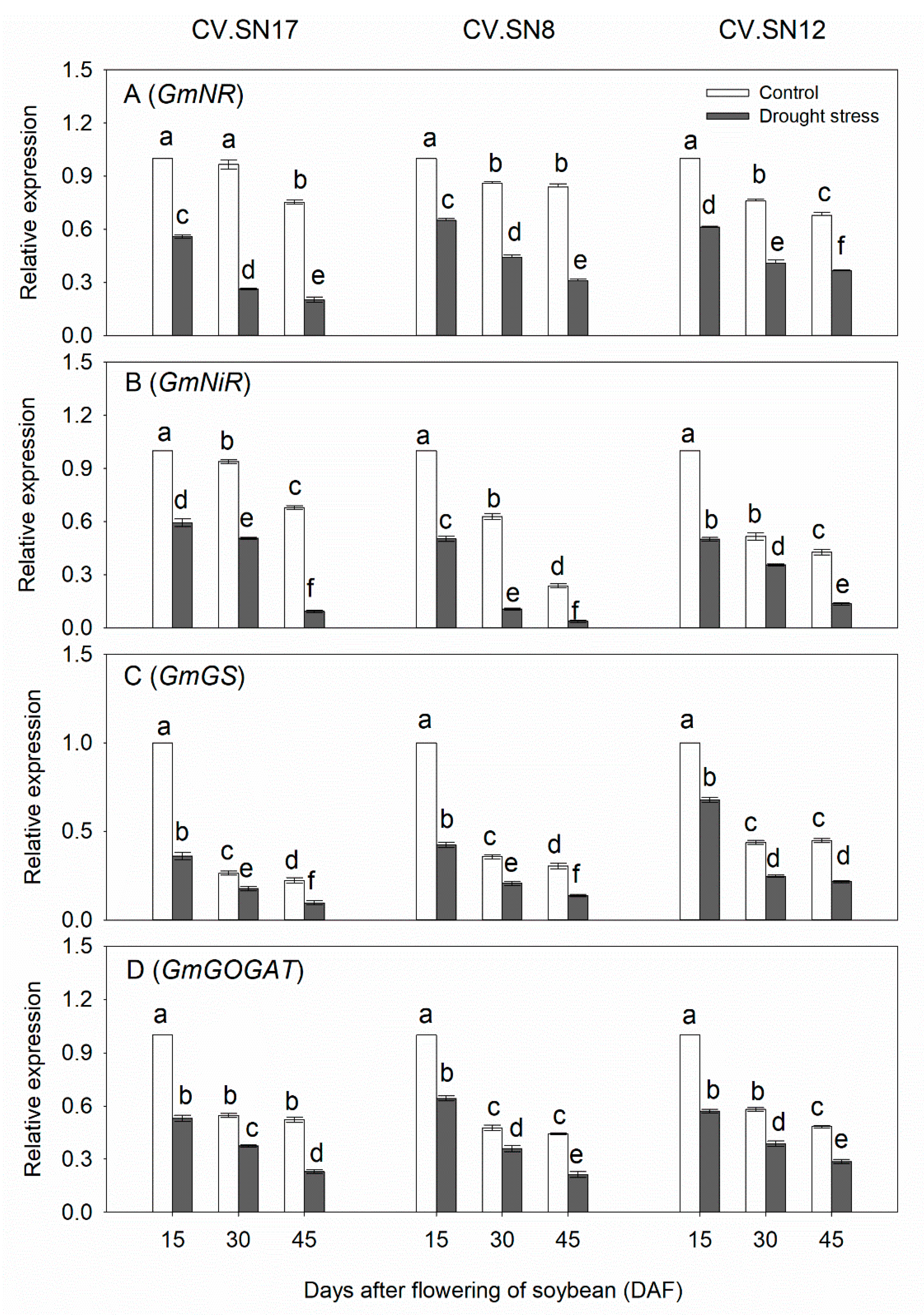
3.5. Effect of Drought Stress on Expression Levels of Genes Involved in Nitrogen Metabolism in Leaves
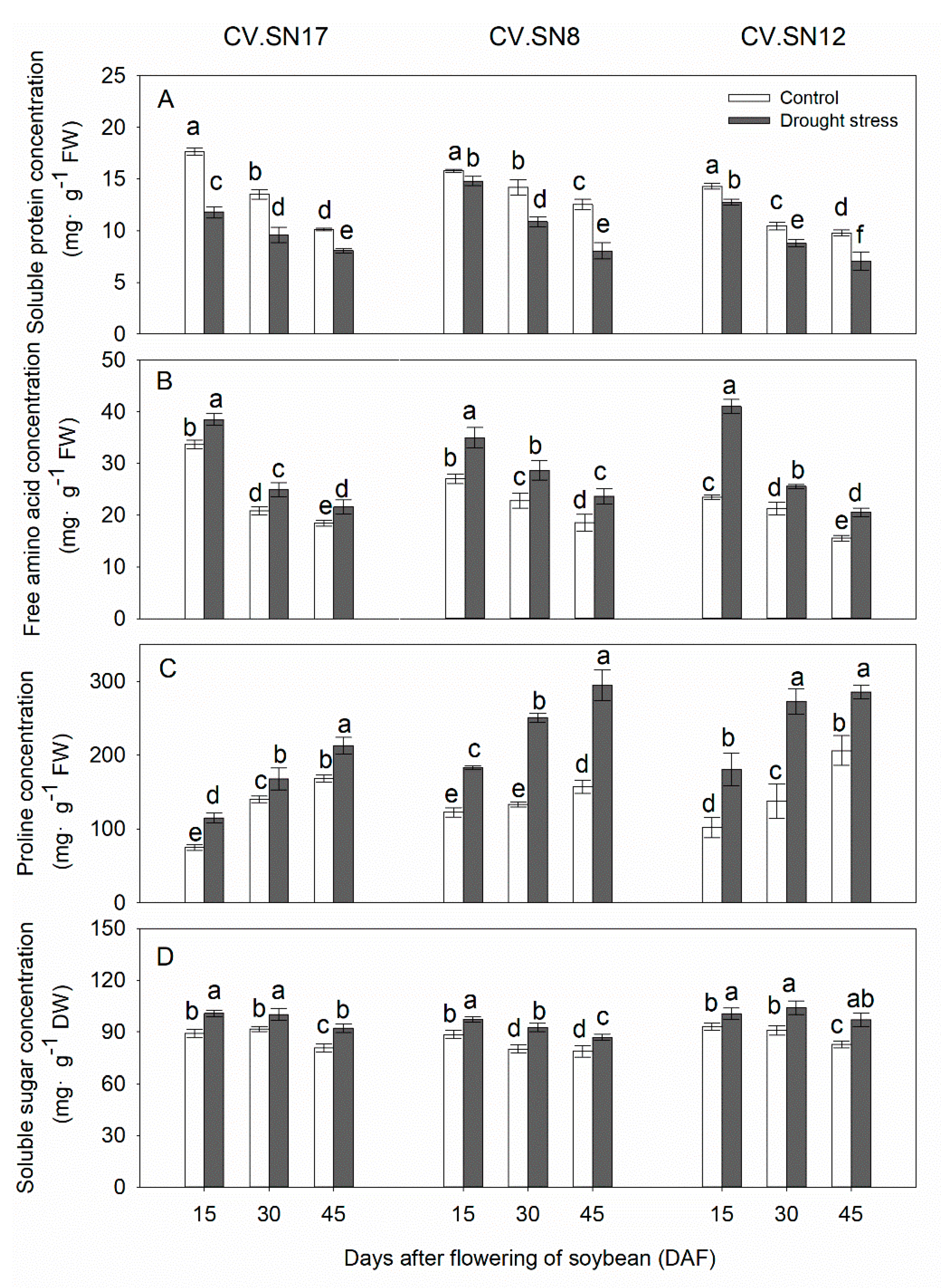
3.6. Effect of Drought Stress on Soluble Protein, Free Amino Acid, Proline, and Soluble Sugar Contents in Leaves
3.7. Correlations of Seed Weight with Physiological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chakhchar, A.; Lamaoui, M.; Aissam, S.; Ferradous, A.; Wahbi, S.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; El Modafar, C. Differential physiological and antioxidative responses to drought stress and recovery among four contrasting Argania spinosa ecotypes. J. Plant Interact. 2016, 11, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.R.; Chaitanya, K.; Jutur, P.; Sumithra, K. Differential antioxidative responses to water stress among five mulberry (Morus alba L.) cultivars. Environ. Exp. Bot. 2004, 52, 33–42. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, S.; Liu, Z.; Yang, F.; Yin, G. Drought tolerance in alfalfa (Medicago sativa L.) varieties is associated with enhanced antioxidative protection and declined lipid peroxidation. J. Plant Physiol. 2019, 232, 226–240. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant 2015, 37, 9. [Google Scholar] [CrossRef]
- Fougereux, J.-A.; Doré, T.; Ladonne, F.; Fleury, A. Water stress during reproductive stages affects seed quality and yield of pea (Pisum sativum L.). Crop Sci. 1997, 37, 1247–1252. [Google Scholar] [CrossRef]
- Begcy, K.; Walia, H. Drought stress delays endosperm development and misregulates genes associated with cytoskeleton organization and grain quality proteins in developing wheat seeds. Plant Sci. 2015, 240, 109–119. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Muller, B.; Pantin, F.; Génard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Blumwald, E. The roles of ROS and ABA in systemic acquired acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Cicco, N.; Paraggio, M.; Scopa, A. Regulation of the ascorbate—Glutathione cycle in plants under drought stress. In Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Anjum, N.A., Chan, M.T., Umar, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 137–189. [Google Scholar]
- Xu, L.; Han, L.; Huang, B. Antioxidant enzyme activities and gene expression patterns in leaves of Kentucky bluegrass in response to drought and post-drought recovery. J. Am. Soc. Hortic. Sci. 2011, 136, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Kumar, S. Stipules are the principal photosynthetic organs in the papilionoid species Lathyrus aphaca. Natl. Acad. Sci. Lett. 2012, 35, 75–78. [Google Scholar] [CrossRef]
- Ge, T.D.; Sui, F.G.; Bai, L.P.; Lu, Y.Y.; Zhou, G.S. Effects of water stress on the protective enzyme activities and lipid peroxidation in roots and leaves of summer maize. Agric. Sci. China 2006, 5, 291–298. [Google Scholar] [CrossRef]
- Wang, R.; Gao, M.; Ji, S.; Wang, S.; Meng, Y.; Zhou, Z. Carbon allocation, osmotic adjustment, antioxidant capacity and growth in cotton under long-term soil drought during flowering and boll-forming period. Plant Physiol. Biochem. 2016, 107, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Zhong, C.; Zhu, C.; Zhu, L.; Zhang, J.; Wu, L.; Jin, Q. Ammonium uptake and metabolism alleviate PEG-induced water stress in rice seedlings. Plant Physiol. Biochem. 2018, 132, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Cao, X.; Bai, Z.; Zhang, J.; Zhu, L.; Huang, J.; Jin, Q. Nitrogen metabolism correlates with the acclimation of photosynthesis to short-term water stress in rice (Oryza sativa L.). Plant Physiol. Biochem. 2018, 125, 52–62. [Google Scholar] [CrossRef]
- Zhong, C.; Bai, Z.G.; Zhu, L.F.; Zhang, J.H.; Zhu, C.Q.; Huang, J.L.; Jin, Q.Y.; Cao, X.C. Nitrogen-mediated alleviation of photosynthetic inhibition under moderate water deficit stress in rice (Oryza sativa L.). Environ. Exp. Bot. 2019, 157, 269–282. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res. 2008, 108, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Yao, X.; Liu, W.; Wu, Z.; Zhao, Q.; Zhang, H.; Wang, H.; Ao, X.; Zhao, M.; Xie, F. Precise nitrogen topdressing upregulates nitrogen metabolism and improves soybean (Glycine max) grain yield. Crop Pasture Sci. 2019, 70, 334–343. [Google Scholar] [CrossRef]
- Xu, Z.Z.; Zhou, G.S. Combined effects of water stress and high temperature on photosynthesis, nitrogen metabolism and lipid peroxidation of a perennial grass Leymus chinensis. Planta 2006, 224, 1080–1090. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Shim, I.S.; Kobayashi, K.; Usui, K. Regulation of ammonium accumulation during salt stress in rice (Oryza sativa L.) seedlings. Plant Prod. Sci. 2005, 8, 397–404. [Google Scholar] [CrossRef]
- Foyer, C.H.; Valadier, M.H.; Migge, A.; Becker, T.W. Drought-induced effects on nitrate reductase activity and mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Physiol. 1998, 117, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rodríguez, E.; del Mar Rubio-Wilhelmi, M.; Ríos, J.J.; Blasco, B.; Rosales, M.Á.; Melgarejo, R.; Romero, L.; Ruiz, J.M. Ammonia production and assimilation: Its importance as a tolerance mechanism during moderate water deficit in tomato plants. J. Plant Physiol. 2011, 168, 816–823. [Google Scholar] [CrossRef] [PubMed]
- del Mar Rubio-Wilhelmi, M.; Sanchez-Rodriguez, E.; Leyva, R.; Blasco, B.; Romero, L.; Blumwald, E.; Ruiz, J.M. Response of carbon and nitrogen-rich metabolites to nitrogen deficiency in PSARK∷IPT tobacco plants. Plant Physiol. Biochem. 2012, 57, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.; Németh, E.; Guóth, A.; Bona, L.; Wodala, B.; Pécsváradi, A. Metabolic indicators of drought stress tolerance in wheat: Glutamine synthetase isoenzymes and Rubisco. Plant Physiol. Biochem. 2013, 67, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Ghosh, S. Regulation of glutamine synthetase isoforms in two differentially drought-tolerant rice (Oryza sativa L.) cultivars under water deficit conditions. Plant Cell Rep. 2013, 32, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Coruzzi, G.M.; Zhou, L. Carbon and nitrogen sensing and signaling in plants: Emerging ‘matrix effects’. Curr. Opin. Plant Biol. 2001, 4, 247–253. [Google Scholar] [CrossRef]
- Ashraf, M.F.M.R.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B.J.B.P. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef]
- Zahoor, R.; Zhao, W.; Abid, M.; Dong, H.; Zhou, Z. Potassium application regulates nitrogen metabolism and osmotic adjustment in cotton (Gossypium hirsutum L.) functional leaf under drought stress. J. Plant Physiol. 2017, 215, 30–38. [Google Scholar] [CrossRef] [PubMed]
- de Souza Miranda, R.; Gomes-Filho, E.; Prisco, J.T.; Alvarez-Pizarro, J.C. Ammonium improves tolerance to salinity stress in Sorghum bicolor plants. Plant Growth Regul. 2016, 78, 121–131. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef]
- Sperandio, M.V.L.; Santos, L.A.; Bucher, C.A.; Fernandes, M.S.; de Souza, S.R. Isoforms of plasma membrane H+-ATPase in rice root and shoot are differentially induced by starvation and resupply of NO3− or NH4+. Plant Sci. 2011, 180, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.L.; Berhow, M.A.; Kim, J.T.; Chi, H.Y.; Lee, S.J.; Chung, I.M. Evaluation of soyasaponin, isoflavone, protein, lipid, and free sugar accumulation in developing soybean seeds. J. Agric. Food Chem. 2006, 54, 10003–10010. [Google Scholar] [CrossRef]
- Sakthivelu, G.; Akitha Devi, M.; Giridhar, P.; Rajasekaran, T.; Ravishankar, G.; Nikolova, M.; Angelov, G.; Todorova, R.; Kosturkova, G. Isoflavone composition, phenol content, and antioxidant activity of soybean seeds from India and Bulgaria. J. Agric. Food Chem. 2008, 56, 2090–2095. [Google Scholar] [CrossRef]
- Liu, F.; Jensen, C.R.; Andersen, M.N. Drought stress effect on carbohydrate concentration in soybean leaves and pods during early reproductive development: Its implication in altering pod set. Field Crops Res. 2004, 86, 1–13. [Google Scholar] [CrossRef]
- Lobato, A.; Neto, M.; Meirelles, A.; Silva, L.; Marochio, C.; Monteiro, E.; Zeni Neto, H.; Maleia, M.; Moiana, L.; Bronzato, A. Relationship between leaf relative water content and total soluble proteins in soybean exposed to short water deficit. Res. J. Biol. Sci. 2009, 4, 1061–1067. [Google Scholar]
- Masoumi, H.; Masoumi, M.; Darvish, F.; Daneshian, J.; Nourmohammadi, G.; Habibi, D. Change in several antioxidant enzymes activity and seed yield by water deficit stress in soybean (Glycine max L.) cultivars. Not. Bot. Horti Agrobot. Cluj Napoca 2010, 38, 86–94. [Google Scholar]
- Fehr, W.; Caviness, C.; Burmood, D.; Pennington, J. Stage of development descriptions for soybeans, Glycine Max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct signalling pathways and transcriptome response signatures differentiate ammonium-and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hendrix, D.L. Rapid extraction and analysis of nonstructural carbohydrates in plant tissues. Crop Sci. 1993, 33, 1306–1311. [Google Scholar] [CrossRef]
- Ding, Y.; Luo, W.; Xu, G. Characterisation of magnesium nutrition and interaction of magnesium and potassium in rice. Ann. Appl. Biol. 2006, 149, 111–123. [Google Scholar] [CrossRef]
- Hageman, R.; Reed, A. Nitrate reductase from higher plants. Methods Enzymol. 1980, 69, 270–280. [Google Scholar]
- Singh, R.P.; Srivastava, H. Increase in glutamate synthase (NADH) activity in maize seedlings in response to nitrate and ammonium nitrogen. Physiol. Plant 1986, 66, 413–416. [Google Scholar] [CrossRef]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; Medeiros, J.V.R.; Gomes-Filho, E. Hydrogen peroxide pre-treatment induces salt-stress acclimation in maize plants. J. Plant Physiol. 2005, 162, 1114–1122. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Havir, E.A.; McHale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jesus Miranda, V.; Coelho, R.R.; Viana, A.A.B.; de Oliveira Neto, O.B.; Carneiro, R.M.D.G.; Rocha, T.L.; de Sa, M.F.G.; Fragoso, R.R. Validation of reference genes aiming accurate normalization of qPCR data in soybean upon nematode parasitism and insect attack. BMC Res. Notes 2013, 6, 196. [Google Scholar]
- Du, Y.; Zhao, Q.; Li, S.; Yao, X.; Xie, F.; Zhao, M. Shoot/root interactions affect soybean photosynthetic traits and yield formation: A case study of grafting with record-yield cultivars. Front. Plant Sci. 2019, 10, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarah, N.; Alqudah, A.; Amayreh, J.; McAndrews, G. The effect of late-terminal drought stress on yield components of four barley cultivars. J. Agron. Crop Sci. 2009, 195, 427–441. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K. Drought stress in grain legumes during reproduction and grain filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Devi, M.A.; Giridhar, P. Variations in physiological response, lipid peroxidation, antioxidant enzyme activities, proline and isoflavones content in soybean varieties subjected to drought stress. Proc. Natl. Acad. Sci. India Sec. B Biol. Sci. 2015, 85, 35–44. [Google Scholar] [CrossRef]
- Zhang, C.; Zeng, G.; Chen, J. Effects of drought stress on the protective enzymes activities and membrane lipid peroxidation in leaves of Brassica parachinensis LH Bailey. J. Plant Resour. Environ. 2000, 9, 23–26, (In Chinese, with English abstract). [Google Scholar]
- Fu, J.; Huang, B. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Saito, A.; Tanabata, S.; Tanabata, T.; Tajima, S.; Ueno, M.; Ishikawa, S.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Effect of nitrate on nodule and root growth of soybean (Glycine max (L.) Merr.). Int. J. Mol. Sci. 2014, 15, 4464–4480. [Google Scholar] [CrossRef]
- La Menza, N.C.; Monzon, J.P.; Specht, J.E.; Grassini, P. Is soybean yield limited by nitrogen supply? Field Crops Res. 2017, 213, 204–212. [Google Scholar] [CrossRef]
- Zapata, F.; Danso, S.; Hardarson, G.; Fried, M. Time course of nitrogen fixation in field-grown soybean using nitrogen-15 methodology. Agron. J. 1987, 79, 172–176. [Google Scholar] [CrossRef]
- Barker, D.W.; Sawyer, J.E. Nitrogen application to soybean at early reproductive development. Agron. J. 2005, 97, 615–619. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Boem, F.H.; Scheiner, J.D.; Rimski-Korsakov, H.; Lavado, R.S. Late season nitrogen fertilization of soybeans: Effects on leaf senescence, yield and environment. Nutr. Cycl. Agroecosyst. 2004, 68, 109–115. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.; Nagarajan, L.; Roy, A.; Rabbinge, R. Revisiting fertilisers and fertilisation strategies for improved nutrient uptake by plants. Biol. Fertil. Soils 2015, 51, 897–911. [Google Scholar] [CrossRef] [Green Version]
- Meng, S.; Zhang, C.; Su, L.; Li, Y.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Armijo, G.; Gutiérrez, R.A. Emerging players in the nitrate signaling pathway. Mol. Plant 2017, 10, 1019–1022. [Google Scholar] [CrossRef]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef]
- Fresneau, C.; Ghashghaie, J.; Cornic, G. Drought effect on nitrate reductase and sucrose-phosphate synthase activities in wheat (Triticum durum L.): Role of leaf internal CO2. J. Exp. Bot. 2007, 58, 2983–2992. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, S.; Bacon, M.A.; Davies, W.J. Nitrate signalling to stomata and growing leaves: Interactions with soil drying, ABA, and xylem sap pH in maize. J. Exp. Bot. 2007, 58, 1705–1716. [Google Scholar] [CrossRef]
- Krouk, G.; Crawford, N.M.; Coruzzi, G.M.; Tsay, Y.F. Nitrate signaling: Adaptation to fluctuating environments. Curr. Opin. Plant Biol. 2010, 13, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.M.; Hilker, C. Nitrate reduction in leaves and roots of young pedunculate oaks (Quercus robur) growing on different nitrate concentrations. Environ. Exp. Bot. 2000, 43, 19–32. [Google Scholar] [CrossRef]
- Hummel, I.; Pantin, F.; Sulpice, R.; Piques, M.; Rolland, G.; Dauzat, M.; Christophe, A.; Pervent, M.; Bouteillé, M.; Stitt, M. Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: An integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol. 2010, 154, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Ben, K.R.; Abdelly, C.; Savouré, A. Proline, a multifunctional amino-acid involved in plant adaptation to environmental constraints. Biol. Aujourd’hui 2012, 206, 291–299. [Google Scholar]
- Matysik, J.; Alia; Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Wingler, A.; Quick, W.; Bungard, R.; Bailey, K.; Lea, P.; Leegood, R. The role of photorespiration during drought stress: An analysis utilizing barley mutants with reduced activities of photorespiratory enzymes. Plant Cell Environ. 1999, 22, 361–373. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2010, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer ID | Primer Sequence |
|---|---|---|
| GmNR | Glyma.06G109200_F | GATCCTCGCCCGTATGAAGG |
| Glyma.06G109200_R | CATACTTGGACCCACCACCC | |
| GmNiR | Glyma.02G132100_F | AACCCCGCCATGTCAAACTT |
| Glyma.02G132100_R | TTGCAGGCATGTAAGCCAGA | |
| GmGS | Glyma.11G215500_F | CTATTCCCACTAACAAG AGGCACGC |
| Glyma.11G215500_R | GCCAACACCACAGTAGTATGGACCG | |
| GmGOGAT | Glyma.03G128300_F | ACACTCTCATCTTCTCTCTCTCTCTCTCTCG |
| Glyma.03G128300_R | CGTTGTGGAGGGAAGGGAATG | |
| GmEF1a | Glyma.05G114900_F | GACCTTCTTCGTTTCTCGCA |
| Glyma.05G114900_R | CGAACCTCTCAATCACACGC | |
| GmEF1b | Glyma.14G039100_F | GTTGAAAAGCCAGGGGACA |
| Glyma.14G039100_R | TCTTACCCCTTGAGCGTGG |
| Genotype | Treatment | Root Biomass (g plant−1) | Stem Biomass (g plant−1) | Pod Wall Weight (g plant−1) | Pod Number (plant−1) | Seed Number (plant−1) | Seed Weight (g plant−1) | 100-Seed Weight (g plant−1) |
|---|---|---|---|---|---|---|---|---|
| CV.SN17 | CK | 8.04 ± 1.41a | 21.32 ± 2.5a | 16.9 ± 0.51a | 75 ± 3a | 147 ± 2b | 38.31 ± 0.25a | 25.03 ± 0.54a |
| DS | 2.40 ± 0.5d | 10.21 ± 1.14d | 5.33 ± 0.22d | 27 ± 3e | 53 ± 1f | 12.03 ± 1.16e | 21.05 ± 0.85c | |
| CV.SN8 | CK | 3.31 ± 0.5bc | 14.07 ± 1.92c | 12.7 ± 1.69c | 70 ± 3b | 126 ± 5c | 33.24 ± 0.87b | 25.54 ± 0.95a |
| DS | 2.40 ± 0.27bc | 6.88 ± 1.82e | 4.06 ± 0.63e | 39 ± 3d | 61 ± 5e | 11.16 ± 1.00e | 22.16 ± 0.60b | |
| CV.SN12 | CK | 3.92 ± 0.42b | 18.6 ± 0.98b | 13.83 ± 0.39b | 80 ± 2a | 227 ± 9a | 38.98 ± 0.67a | 16.82 ± 0.11d |
| DS | 2.96 ± 0.23cd | 11.99 ± 0.48d | 5.05 ± 0.53d | 51 ± 2c | 94 ± 7d | 16.75 ± 0.31d | 15.95 ± 0.11d | |
| Source of variations | ||||||||
| Treatment (T) (df = 1) | 121.85 ** | 235.87 ** | 1262.44 ** | 913.92 ** | 1625.56 ** | 10500.15 ** | 88.06 ** | |
| Genotype (G) (df = 2) | 39.35 ** | 39.05 ** | 34.43 ** | 52.47 ** | 287.27 ** | 203.14 ** | 261.91 ** | |
| T × G (df = 2) | 47.90 ** | 6.84 ** | 12.32 ** | 26.64 ** | 73.59 ** | 35.86 ** | 10.63 ** |
| Cultivars | Treatment | Root/Plant | Stem/Plant | Pod Wall/Plant | Seed/Plant |
|---|---|---|---|---|---|
| CV.SN17 | CK | 9.51 | 25.21 | 19.99 | 45.30 |
| DS | 8.00 ** | 34.06 ** | 17.79 ** | 40.15 ** | |
| CV.SN8 | CK | 5.23 | 22.22 | 20.05 | 52.50 |
| DS | 9.79 ** | 28.10 ** | 16.56 ** | 45.55 ** | |
| CV.SN12 | CK | 5.20 | 24.69 | 18.36 | 51.75 |
| DS | 8.06 ** | 32.62 ** | 13.73 ** | 45.58 ** |
| Traits (/plant) | Source of Variations | ||||||
|---|---|---|---|---|---|---|---|
| Growth Stages (S) (df = 2) | Treatment (T) (df = 1) | Genotype (G) (df = 2) | S × T (df = 2) | S × G (df = 4) | T × G (df = 2) | S × T × G (df = 4) | |
| Nitrogen concentration (mg g−1) | 73.78 ** | 258.81 ** | 119.48 ** | 6.44 ** | 3.38 * | 23.87 ** | 0.97 NS |
| Nitrate concentration (mg g−1) | 32.05 ** | 1104.61 ** | 110.56 ** | 135.7 ** | 6.97 ** | 35.01 ** | 6.09 ** |
| Soluble protein concentration (mg g−1) | 511.26 ** | 474.76 ** | 86.52 ** | 0.39 NS | 3.57 * | 17.8 ** | 21.48 ** |
| Free amino acid concentration (mg g−1) | 572.79 ** | 381.29 ** | 10.40 ** | 31.58 ** | 18.71 ** | 18.37 ** | 14.15 ** |
| Proline concentration (mg g−1) | 536.11 ** | 1021.81 ** | 271.68 ** | 115.43 ** | 105.69 ** | 171.75 ** | 62.22 ** |
| Soluble sugar concentration (mg g−1) | 108.4 ** | 137.38 ** | 36.11 ** | 8.65 ** | 5.00 ** | 0.21 NS | 2.04 NS |
| NR activity (μg−1 FW h−1) | 703.77 ** | 284.74 ** | 62.42 ** | 5.89 ** | 44.00 ** | 2.40 NS | 1.04 NS |
| NiR activity (μmol g−1 FW h−1) | 134.59 ** | 866.12 ** | 1.37 NS | 73.66 ** | 2.25 NS | 8.41 ** | 0.96 NS |
| GS activity (μmol g−1 FW h−1) | 1351.24 ** | 1082.03 ** | 306.95 ** | 22.38 ** | 36.10 ** | 31.98 ** | 34.78 ** |
| GOGAT activity (μmol g−1 FW h−1) | 548 ** | 227.48 ** | 56.22 ** | 2.11 NS | 29.15 ** | 1.43 NS | 0.29 NS |
| GmNR | 2475.46 ** | 18594.23 ** | 118.98 ** | 79.1 ** | 21.85 ** | 360.04 ** | 99.48 ** |
| GmNiR | 8857.47 ** | 17,121.19 ** | 1745.57 ** | 126.06 ** | 364.56 ** | 226.87 ** | 360.32 ** |
| GmGS | 10,763.28 ** | 7859.28 ** | 802.44 ** | 1446.63 ** | 16.56 ** | 22.94 ** | 179.66 ** |
| GmGOGAT | 6747.71 ** | 7460.88 ** | 27.14 ** | 580.73 ** | 49.6 ** | 50.34 ** | 15.31 ** |
| SOD activity (U mg−1 min−1) | 5132.44 ** | 643.77 ** | 34.80 ** | 84.27 ** | 21.18 ** | 6.94 ** | 5.38 ** |
| POD activity (U mg−1 min−1) | 1614.38 ** | 1147.66 ** | 103.75 ** | 61.04 ** | 23.00 ** | 46.86 ** | 24.72 ** |
| CAT activity (U mg−1 min−1) | 1510.18 ** | 1487.9 ** | 30.51 ** | 614.69 ** | 9.69 ** | 12.38 ** | 4.06 ** |
| MDA concentration (mmol g−1) | 156.35 ** | 137.39 ** | 3.00 NS | 12.18 ** | 7.47 ** | 4.10 * | 1.78 NS |
| Traits (/plant) | 15DAF | 30DAF | 45DAF |
|---|---|---|---|
| Nitrogen concentration (mg g−1) | 0.375 | 0.677 ** | 0.609 ** |
| Nitrate concentration (mg g−1) | −0.621 ** | −0.897 ** | −0.992 ** |
| Soluble protein concentration (mg g−1) | 0.674 ** | 0.621 ** | 0.725 ** |
| Free amino acid concentration (mg g−1) | −0.755 ** | −0.858 ** | −0.870 ** |
| Proline concentration (mg g−1) | −0.723 ** | −0.740 ** | −0.506 * |
| Soluble sugar concentration (mg g−1) | −0.803 ** | −0.587* | −0.721 ** |
| NR activity (μg g−1 FW h−1) | 0.712 ** | 0.886 ** | 0.948 ** |
| NiR activity (μmol g−1 FW h−1) | 0.884 ** | 0.956 ** | 0.978 ** |
| GS activity (μmol g−1 FW h−1) | 0.711 ** | 0.639 ** | 0.759 ** |
| GOGAT activity (μmol g−1 FW h−1) | 0.401 | 0.748 ** | 0.891 ** |
| GmNR | 0.968 ** | 0.929 ** | 0.930 ** |
| GmNiR | 0.960 ** | 0.746 ** | 0.871 ** |
| GmGS | 0.961 ** | 0.806 ** | 0.805 ** |
| GmGOGAT | 0.956 ** | 0.967 ** | 0.988 ** |
| SOD activity (U mg−1 min−1) | −0.651 ** | −0.937 ** | 0.844 ** |
| POD activity (U mg−1 min−1) | −0.668 ** | −0.903 ** | −0.779 ** |
| CAT activity (U mg−1 min−1) | −0.874 ** | −0.776 ** | −0.785 ** |
| MDA concentration (mmol g−1) | −0.778 ** | −0.659 ** | −0.859 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Xie, F. Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean. Agronomy 2020, 10, 302. https://doi.org/10.3390/agronomy10020302
Du Y, Zhao Q, Chen L, Yao X, Xie F. Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean. Agronomy. 2020; 10(2):302. https://doi.org/10.3390/agronomy10020302
Chicago/Turabian StyleDu, Yanli, Qiang Zhao, Liru Chen, Xingdong Yao, and Futi Xie. 2020. "Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean" Agronomy 10, no. 2: 302. https://doi.org/10.3390/agronomy10020302
APA StyleDu, Y., Zhao, Q., Chen, L., Yao, X., & Xie, F. (2020). Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean. Agronomy, 10(2), 302. https://doi.org/10.3390/agronomy10020302