The Use of a Tomato Landrace as Rootstock Improves the Response of Commercial Tomato under Water Deficit Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Growth Conditions
2.3. Leaf Carbon Isotope Composition
2.4. Pre-Dawn Leaf Water Potential and Scion Fresh Weight
2.5. Fruit Production, Fruit Quality and Shelf-Life
2.6. Statistical Analysis
3. Results
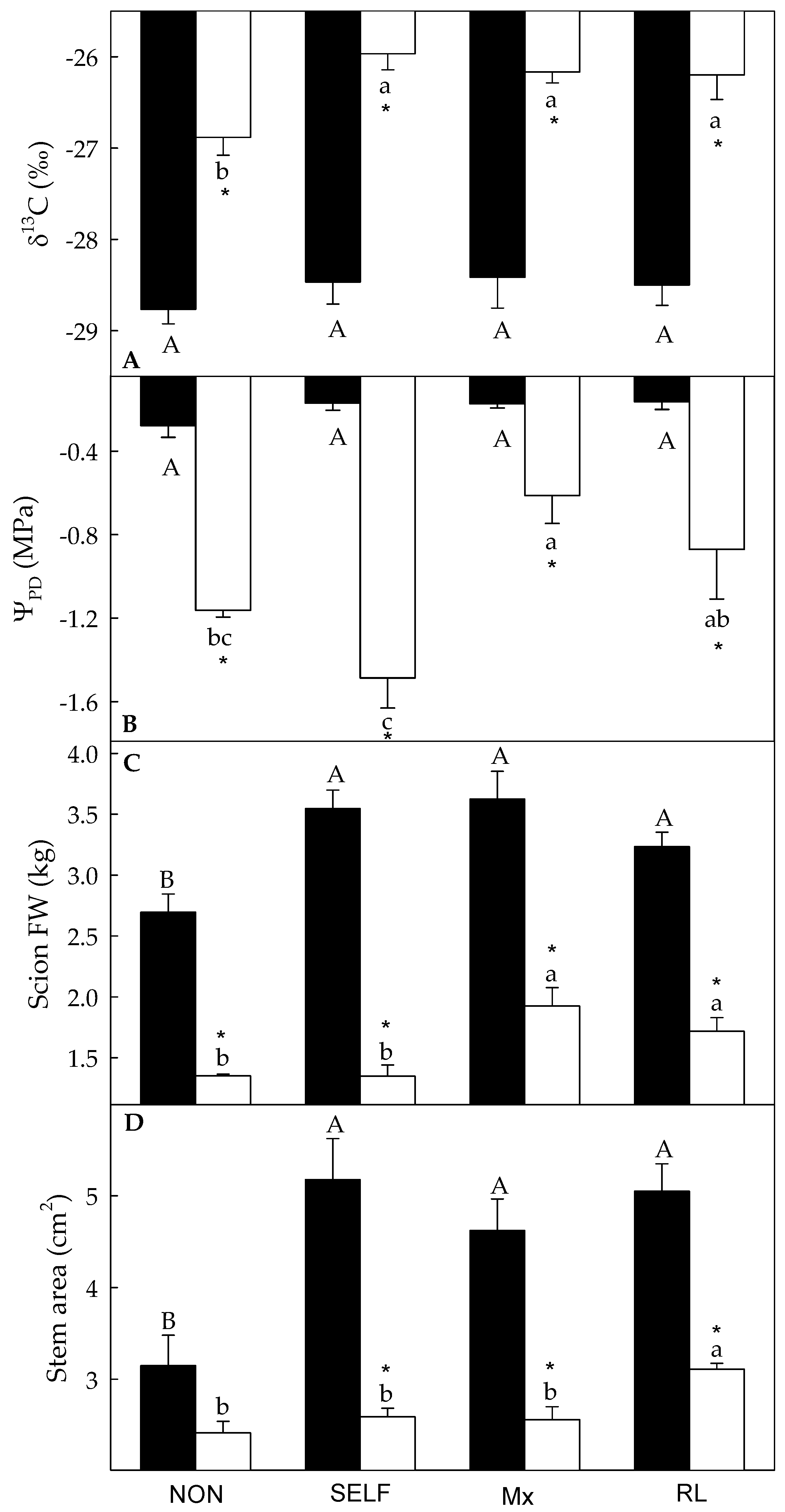
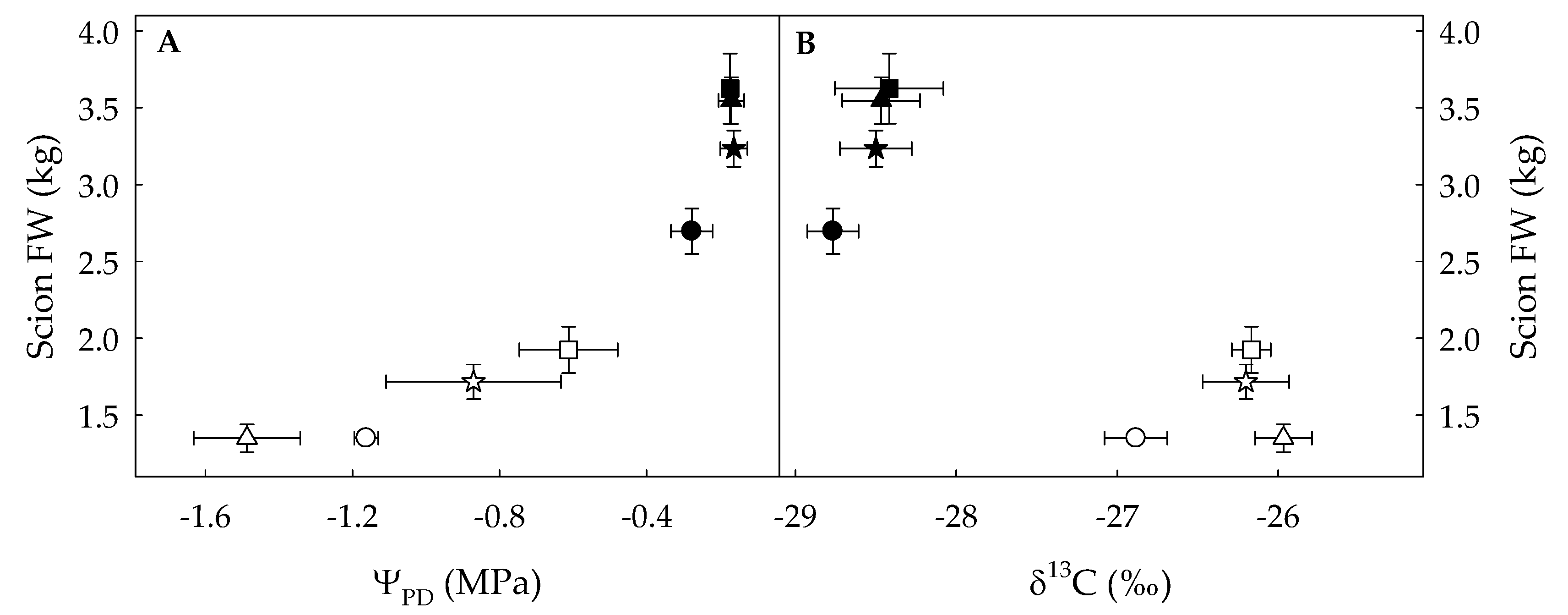
3.1. Analysis of the Variability of δ13C and Growth Parameters under WW and WD Conditions
3.2. Variation in Fruit Production and Its Correlation with δ13C as a Result of Water Deficit, Grafting and the Rootstock Genotype
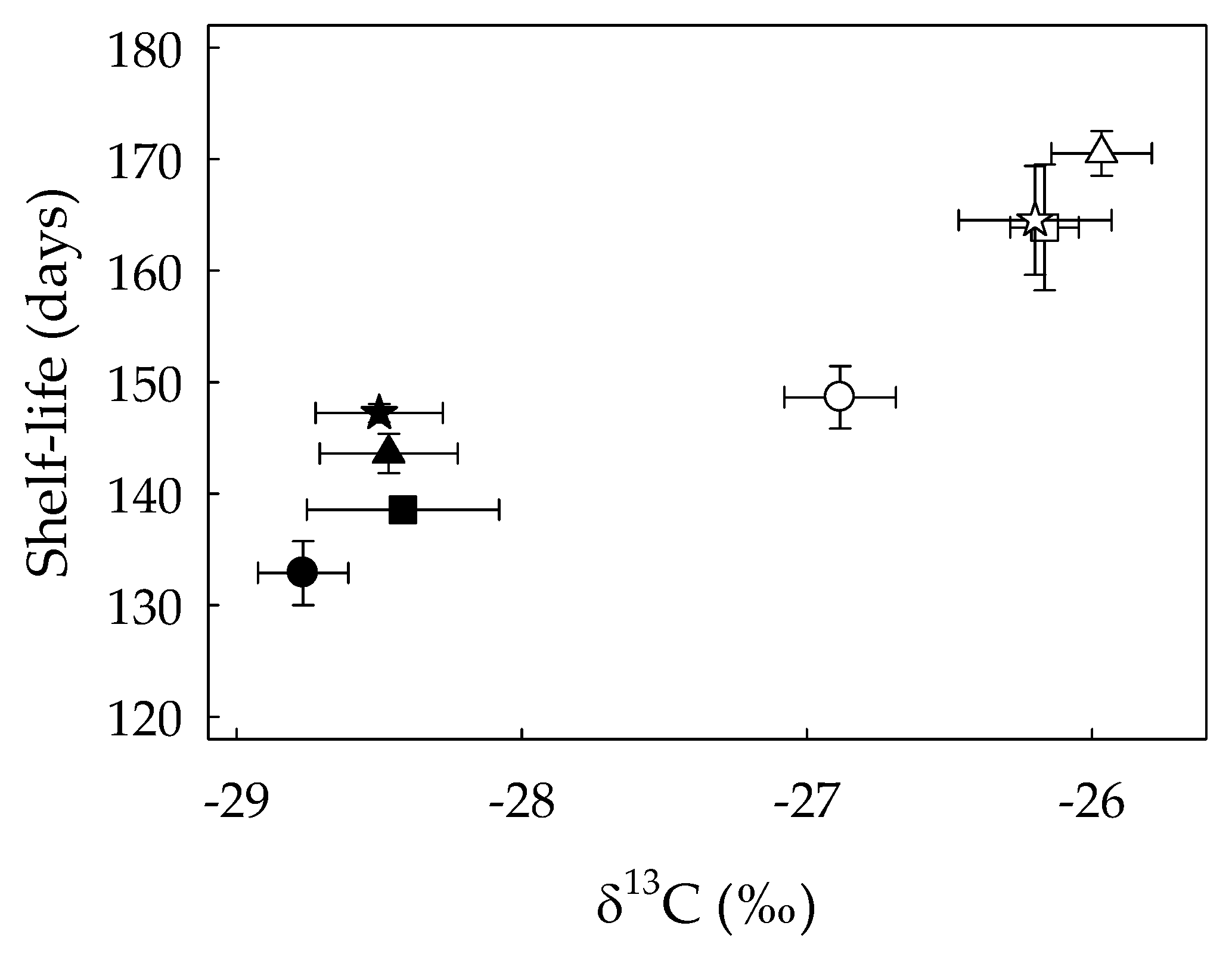
3.3. Modification of the Scion Fruit Quality and Shelf-Life Related to Water Deficit, Grafting and the Rootstock Genotype
4. Discussion
4.1. Grafting Maximized Plant Growth
4.2. Grafting onto RL Increased Fruit Production in Both Treatments
4.3. Grafting Had an Effect on Fruit Quality and Shelf-Life
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baron, D.; Esteves Amaro, A.C.; Pina, A.; Ferreira, G. An overview of grafting re-establishment in woody fruit species. Sci. Hortic. (Amst.) 2019, 243, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Rivard, C.L.; Louws, F.J. Grafting to manage soilborne diseases in heirloom tomato production. Hortic. Rev. (Am. Soc. Hortic. Sci.) 2008, 43, 2104–2111. [Google Scholar] [CrossRef] [Green Version]
- Žanić, K.; Dumičić, G.; Mandušić, M.; Vuletin Selak, G.; Bočina, I.; Urlić, B.; Ljubenkov, I.; Bučević Popović, V.; Goreta Ban, S. Bemisia tabaci MED Population Density as Affected by Rootstock-Modified Leaf Anatomy and Amino Acid Profiles in Hydroponically Grown Tomato. Front. Plant Sci. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, C.; Giuffrida, F. Variation of plant growth and macronutrient uptake in grafted tomatoes and eggplants on three different rootstocks. Eur. J. Hortic. Sci. 2006, 71, 97–101. [Google Scholar]
- Estañ, M.T.; Martinez-Rodriguez, M.M.; Perez-Alfocea, F.; Flowers, T.J.; Bolarin, M.C. Grafting raises the salt tolerance of tomato through limiting the transport of sodium and chloride to the shoot. J. Exp. Bot. 2005, 56, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penella, C.; Nebauer, S.G.; Quiñones, A.; San Bautista, A.; López-Galarza, S.; Calatayud, A. Some rootstocks improve pepper tolerance to mild salinity through ionic regulation. Plant Sci. 2015, 230, 12–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Yu, L.; Wang, L.; Guo, S. Bottle gourd rootstock-grafting promotes photosynthesis by regulating the stomata and non-stomata performances in leaves of watermelon seedlings under NaCl stress. J. Plant Physiol. 2015, 186–187, 50–58. [Google Scholar] [CrossRef]
- Rouphael, Y.; Schwarz, D.; Krumbein, A.; Colla, G. Impact of grafting on product quality of fruit vegetables. Sci. Hortic. (Amst.) 2010, 127, 172–179. [Google Scholar] [CrossRef]
- Fullana-Pericàs, M.; Conesa, M.À.; Pérez-Alfocea, F.; Galmés, J. The influence of grafting on crops’ photosynthetic performance. Plant Sci. 2019, 110250. [Google Scholar] [CrossRef]
- Penella, C.; Nebauer, S.G.; López-Galarza, S.; Quiñones, A.; San Bautista, A.; Calatayud, Á. Grafting pepper onto tolerant rootstocks: An environmental-friendly technique overcome water and salt stress. Sci. Hortic. (Amst.) 2017, 226, 33–41. [Google Scholar] [CrossRef]
- López-Serrano, L.; Penella, C.; San-Bautista, A.; López-Galarza, S.; Calatayud, A. Physiological changes of pepper accessions in response to salinity and water stress. Span. J. Agric. Res. 2017, 15, e0804. [Google Scholar] [CrossRef] [Green Version]
- Aloni, B.; Cohen, R.; Karni, L.; Aktas, H.; Edelstein, M. Hormonal signaling in rootstock-scion interactions. Sci. Hortic. (Amst) 2010, 127, 119–126. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Zhang, X.; Crosby, K. Genetics, breeding and selection of rootstocks for solanaceae and cucurbitaceae. Sci. Hortic. (Amst.) 2010, 127, 106–111. [Google Scholar] [CrossRef]
- FAO. Available online: http://www.fao.org/faostat/en/#data (accessed on 10 April 2020).
- Causse, M.; Friguet, C.; Coiret, C.; Lépicier, M.; Navez, B.; Lee, M.; Holthuysen, N.; Sinesio, F.; Moneta, E.; Grandillo, S. Consumer Preferences for Fresh Tomato at the European Scale: A Common Segmentation on Taste and Firmness. J. Food Sci. 2010, 75, S531–S541. [Google Scholar] [CrossRef] [PubMed]
- Tieman, D.; Zhu, G.; Resende, M.F.R.; Lin, T.; Nguyen, C.; Bies, D.; Rambla, J.L.; Beltran, K.S.O.; Taylor, M.; Zhang, B.; et al. A chemical genetic roadmap to improved tomato flavor. Science (80–) 2017, 355, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Buret, M.; Robini, K.; Verschave, P. Inheritance of Nutritional and Sensory Quality Traits in Fresh Market Tomato and Relation to Consumer Preferences. J. Food Sci. 2003, 68, 2342–2350. [Google Scholar] [CrossRef]
- Galmés, J.; Conesa, M.A.; Ochogavía, J.M.; Perdomo, J.A.; Francis, D.M.; Ribas-Carbó, M.; Savé, R.; Flexas, J.; Medrano, H.; Cifre, J. Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum. Plant Cell Environ. 2011, 34, 245–260. [Google Scholar] [CrossRef]
- Fullana-Pericàs, M.; Conesa, M.À.; Douthe, C.; El Aou-ouad, H.; Ribas-Carbó, M.; Galmés, J. Tomato landraces as a source to minimize yield losses and improve fruit quality under water deficit conditions. Agric. Water Manag. 2019, 223, 105722. [Google Scholar] [CrossRef]
- Ercolano, M.R.; Sacco, A.; Ferriello, F.; D’Alessandro, R.; Tononi, P.; Traini, A.; Barone, A.; Zago, E.; Chiusano, M.L.; Buson, G.; et al. Patchwork sequencing of tomato San Marzano and Vesuviano varieties highlights genome-wide variations. BMC Genomics 2014, 15. [Google Scholar] [CrossRef] [Green Version]
- Suchoff, D.H.; Perkins-Veazie, P.; Sederoff, H.W.; Schultheis, J.R.; Kleinhenz, M.D.; Louws, F.J.; Gunter, C.C. Grafting the indeterminate tomato cultivar moneymaker onto multifort rootstock improves cold tolerance. HortScience 2018, 53, 1610–1617. [Google Scholar] [CrossRef] [Green Version]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an Evolved Concept of Landrace. Front. Plant Sci. 2017, 08, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeven, A.C. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Villa, T.C.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Conesa, M.À.; Fullana-Pericàs, M.; Granell, A.; Galmés, J. Mediterranean Long Shelf-Life Landraces: An Untapped Genetic Resource for Tomato Improvement. Front. Plant Sci. 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Bota, J.; Conesa, M.À.; Ochogavia, J.M.; Medrano, H.; Francis, D.M.; Cifre, J. Characterization of a landrace collection for Tomàtiga de Ramellet (Solanum lycopersicum L.) from the Balearic Islands. Genet. Resour. Crop Evol. 2014, 61, 1131–1146. [Google Scholar] [CrossRef]
- Ochogavía, J.M.; López, M.; Rigo, M.; Garau, M.; March, J.; Moscardó, J.; Jaume, J.; Conesa, M.À.; Galmés, J.; Bota, J.; et al. Caracterització de les Poblacions de Tomàtiga de Ramellet de les Illes Balears; Govern de les Illes Balears: Illes Balears, Spain, 2011; ISBN 9788461472840. [Google Scholar]
- Conesa, M.A.; Galmés, J.; Ochogavía, J.M.; March, J.; Jaume, J.; Martorell, A.; Francis, D.M.; Medrano, H.; Rose, J.K.C.; Cifre, J. The postharvest tomato fruit quality of long shelf-life Mediterranean landraces is substantially influenced by irrigation regimes. Postharvest Biol. Technol. 2014, 93, 11–121. [Google Scholar] [CrossRef]
- Galmés, J.; Ochogavía, J.M.; Gago, J.; Roldán, E.J.; Cifre, J.; Conesa, M.À. Leaf responses to drought stress in Mediterranean accessions of Solanum lycopersicum: Anatomical adaptations in relation to gas exchange parameters. Plant Cell Environ. 2013, 36, 920–935. [Google Scholar] [CrossRef]
- Fullana-Pericàs, M.; Conesa, M.À.; Soler, S.; Ribas-Carbó, M.; Granell, A.; Galmés, J. Variations of leaf morphology, photosynthetic traits and water-use efficiency in Western-Mediterranean tomato landraces. Photosynthetica 2017, 55, 121–133. [Google Scholar] [CrossRef]
- Fullana-Pericàs, M.; Ponce, J.; Conesa, M.À.; Juan, A.; Ribas-Carbó, M.; Galmés, J. Changes in yield, growth and photosynthesis in a drought-adapted Mediterranean tomato landrace (Solanum lycopersicum ‘Ramellet’) when grafted onto commercial rootstocks and Solanum pimpinellifolium. Sci. Hortic. (Amst.) 2018, 233, 70–77. [Google Scholar] [CrossRef]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. (Amst.) 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Alcaraz-López, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological aspects of rootstock-scion interactions. Sci. Hortic. (Amst.) 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Oda, M.; Maruyama, M.; Mori, G. Water Transfer at Graft Union of Tomato Plants Grafted onto Solanum Rootstocks. J. Jpn. Soc. Hortic. Sci. 2005, 74, 458–463. [Google Scholar] [CrossRef] [Green Version]
- Agele, S.; Cohen, S. Effect of genotype and graft type on the hydraulic characteristics and water relations of grafted melon. J. Plant Interact. 2009, 4, 59–66. [Google Scholar] [CrossRef]
- Santa-Cruz, A.; Martinez-Rodriguez, M.M.; Perez-Alfocea, F.; Romero-Aranda, R.; Bolarin, M.C. The rootstock effect on the tomato salinity response depends on the shoot genotype. Plant Sci. 2002, 162, 825–831. [Google Scholar] [CrossRef]
- Farquhar, G.; Richards, R. Isotopic Composition of Plant Carbon Correlates With Water-Use Efficiency of Wheat Genotypes. Funct. Plant Biol. 1984, 11, 539. [Google Scholar] [CrossRef]
- Condon, A.; Farquhar, G.; Richards, R. Genotypic Variation in Carbon Isotope Discrimination and Transpiration Efficiency in Wheat. Leaf Gas Exchange and Whole Plant Studies. Funct. Plant Biol. 1990, 17, 9. [Google Scholar] [CrossRef]
- De Souza, C.R.; Maroco, J.P.; Dos Santos, T.P.; Rodrigues, M.L.; Lopes, C.M.; Pereira, J.S.; Chaves, M.M. Impact of deficit irrigation on water use efficiency and carbon isotope composition (δ13C) of field-grown grapevines under Mediterranean climate. J. Exp. Bot. 2005, 56, 2163–2172. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Vegetable Grafting as a Tool to Improve Drought Resistance and Water Use Efficiency. Front. Plant Sci. 2017, 8, 1130. [Google Scholar] [CrossRef] [Green Version]
- Lykas, C.; Kittas, C.; Zambeka, A. Water and fertilizers use efficiency in grafted and non grafted tomato plants on soilless culture. Acta Hortic. 2008, 801, 1551–1555. [Google Scholar] [CrossRef]
- Cantero-Navarro, E.; Romero-Aranda, R.; Fernández-Muñoz, R.; Martínez-Andújar, C.; Pérez-Alfocea, F.; Albacete, A. Improving agronomic water use efficiency in tomato by rootstock-mediated hormonal regulation of leaf biomass. Plant Sci. 2016, 251, 90–100. [Google Scholar] [CrossRef]
- Borgognone, D.; Colla, G.; Rouphael, Y.; Cardarelli, M.; Rea, E.; Schwarz, D. Effect of nitrogen form and nutrient solution pH on growth and mineral composition of self-grafted and grafted tomatoes. Sci. Hortic. (Amst.) 2013, 149, 61–69. [Google Scholar] [CrossRef]
- Kumar, P.; Edelstein, M.; Cardarelli, M.; Ferri, E.; Colla, G. Grafting affects growth, yield, nutrient uptake, and partitioning under cadmium stress in tomato. HortScience 2015, 50, 1654–1661. [Google Scholar] [CrossRef]
- Blum, A. Drought resistance, water-use efficiency, and yield potential—Are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159–1168. [Google Scholar] [CrossRef]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2011, 63, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Ripoll, J.; Urban, L.; Staudt, M.; Lopez-Lauri, F.; Bidel, L.P.R.; Bertin, N. Water shortage and quality of fleshy fruits-making the most of the unavoidable. J. Exp. Bot. 2014, 65, 4097–4117. [Google Scholar] [CrossRef] [Green Version]
- Bertin, N.; Causse, M.; Brunel, B.; Tricon, D.; Génard, M. Identification of growth processes involved in QTLs for tomato fruit size and composition. J. Exp. Bot. 2009, 60, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Gautier, H.; Lopez-Lauri, F.; Massot, C.; Murshed, R.; Marty, I.; Grasselly, D.; Keller, C.; Sallanon, H.; Génard, M. Impact of ripening and salinity on tomato fruit ascorbate content and enzymatic activities related to ascorbate recycling. Funct. Plant Sci. Biotechnol. 2010, 4, 66–75. [Google Scholar]
- Davis, A.R.; Perkins-Veazie, P.; Hassell, R.; Levi, A.; King, S.R.; Zhang, X. Grafting effects on vegetable quality. HortScience 2008, 43, 1670–1672. [Google Scholar] [CrossRef] [Green Version]
- Fallik, E.; Ilic, Z. Grafted vegetables—the influence of rootstock and scion on postharvest quality. Folia Hortic. 2015, 26, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Matsuzoe, N.; Aida, H.; Hanada, K.; Ali, M.; Okubo, H.; Fujieda, K. Fruit Quality of Tomato Plants Grafted on Solanum Rootstocks. Engei Gakkai Zasshi 1996, 65, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Khah, E.M.; Kakava, E.; Mavromatis, A.; Chachalis, D.; Goulas, C. Effect of grafting on growth and yield of tomato (Lycopersicon esculentum Mill.) in greenhouse and open-field. J. Appl. Hortic. 2006, 8, 3–7. [Google Scholar] [CrossRef]
- Di Gioia, F.; Serio, F.; Buttaro, D.; Ayala, O.; Santamaria, P. Influence of rootstock on vegetative growth, fruit yield and quality in “Cuore di Bue”, an heirloom tomato. J. Hortic. Sci. Biotechnol. 2010, 85, 477–482. [Google Scholar] [CrossRef]
- Malundo, T.M.M.; Shewfelt, R.L.; Scott, J.W. Flavor quality of fresh tomato (Lycopersicon esculentum Mill.) as affected by sugar and acid levels. Postharvest Biol. Technol. 1995, 6, 103–110. [Google Scholar] [CrossRef]
- Causse, M.; Stevens, R.; Amor, B.B.; Faurobert, M.; Muños, S. Breeding for Fruit Quality in Tomato. In Breeding for Fruit Quality; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 279–305. ISBN 9780813810720. [Google Scholar]
- Shepherd, T.; Griffiths, D.W. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.B.B.; Rose, J.K.C. There’s more than one way to skin a fruit: Formation and functions of fruit cuticles. J. Exp. Bot. 2014, 65, 4639–4651. [Google Scholar] [CrossRef] [Green Version]
- Romero, P.; Rose, J.K.C. A relationship between tomato fruit softening, cuticle properties and water availability. Food Chem. 2019, 295, 300–310. [Google Scholar] [CrossRef]
- Sonneveld, C.; Straver, N. Nutrient solutions for vegetables and flowers grown in water or substrates. Voedingspolossingen Glas. 1994, 8, 1–33. [Google Scholar]





{kind=link}
{kind=link}
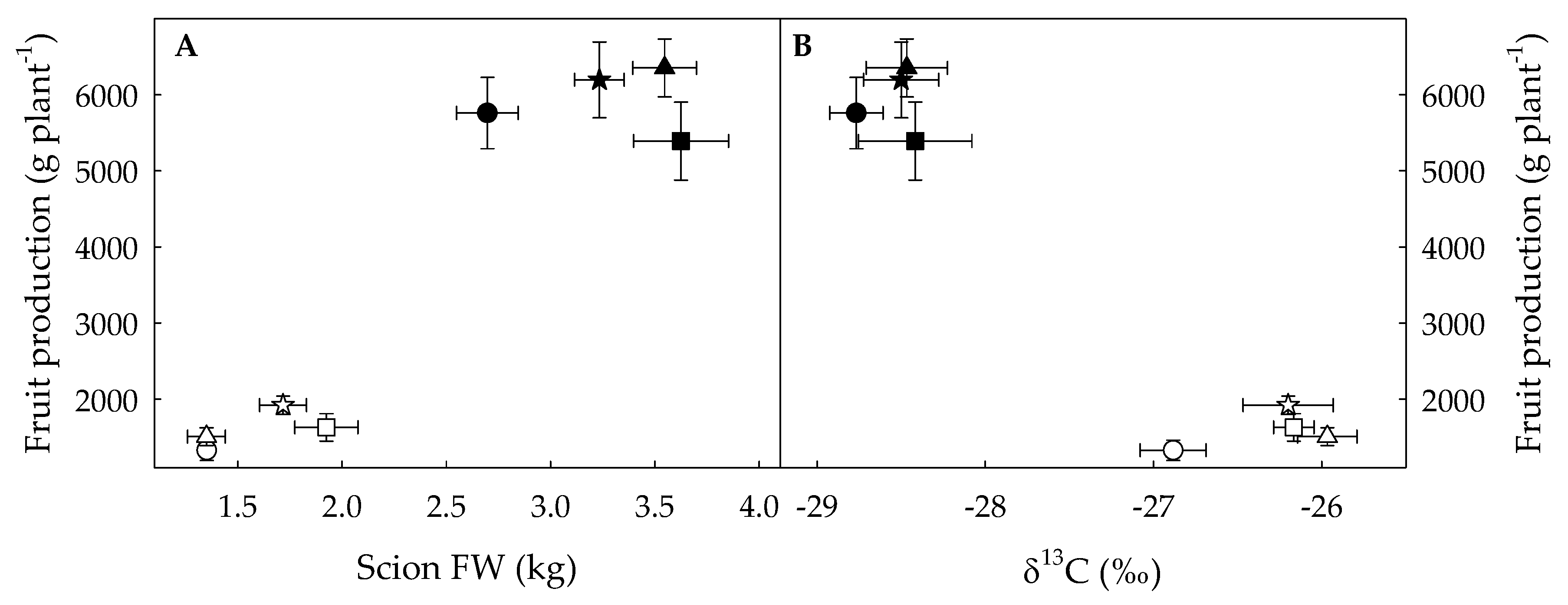{kind=link}
{kind=link}
| Treatment | Graft Combination | Fruit Production | Fruit Number | Fruit Size |
|---|---|---|---|---|
| g plant−1 | fruit plant−1 | g fruit−1 | ||
| WW | ||||
| NON | 5758.8 ± 468.8 a | 88.8 ± 6.5 a | 94.31 ± 11.02 a | |
| SELF | 6352.5 ± 380.8 a | 91.7 ± 6.5 a | 89.31 ± 1.92 a | |
| Mx | 5391.0 ± 512.4 a | 81.7 ± 8.2 a | 77.59 ± 9.30 a | |
| RL | 6195.0 ± 498.5 a | 93.7 ± 4.3 a | 87.04 ± 5.31 a | |
| WD | ||||
| NON | 1327.0 ± 133.9 b* | 20.2 ± 3.1 b* | 67.69 ± 2.08 a* | |
| SELF | 1507.0 ± 117.4 ab* | 32.8 ± 4.3 a* | 56.80 ± 1.34 b* | |
| Mx | 1628.3 ± 180.5 ab* | 38.3 ± 4.0 a* | 58.14 ± 1.29 b* | |
| TR | 1920.0 ± 120.7 a* | 41.5 ± 5.2 a* | 58.39 ± 2.28 b* |
| Treatment | Graft Combination | TSS | Acidity | TSS/Acidity | Hardness | Shelf-Life |
|---|---|---|---|---|---|---|
| °Brix | % Citric Acid | °Shore | Days | |||
| WW | ||||||
| NON | 4.13 ± 0.31 b | 1.14 ± 0.07 a | 3.87 ±0.16 b | 57.97 ± 1.68 a | 132.9 ± 2.9 c | |
| SELF | 5.15 ± 0.10 a | 1.10 ± 0.05 a | 4.81 ± 0.24 a | 50.33 ± 1.33 b | 143.6 ± 1.8 ab | |
| Mx | 4.97 ± 0.22 a | 1.12 ± 0.05 a | 4.44 ± 0.09 ab | 51.45 ± 0.60 b | 138.6 ± 1.1 b | |
| RL | 5.23 ± 0.26 a | 1.22 ± 0.10 a | 4.45 ± 0.31 ab | 51.20 ± 2.73 b | 147.2 ± 0.8 a | |
| WD | ||||||
| NON | 8.43 ± 0.28 a* | 1.52 ± 0.10 b* | 5.39 ± 0.44 a* | 57.47 ± 3.14 a | 148.6 ± 2.8 b* | |
| SELF | 8.35 ± 0.25 a* | 1.60 ± 0.15 b* | 5.33 ± 0.42 ab | 58.73 ± 2.10 a* | 170.5 ± 2.0 a* | |
| Mx | 8.54 ± 0.35 a* | 1.95 ± 0.09 a* | 4.41 ± 0.17 bc | 51.74 ± 2.52 a | 163.9 ± 5.6 a* | |
| RL | 8.10 ± 0.35 a* | 1.92 ± 0.13 a* | 4.27 ± 0.19 c | 54.92 ± 2.98 a | 164.5 ± 4.9 a* |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fullana-Pericàs, M.; Conesa, M.À.; Ribas-Carbó, M.; Galmés, J. The Use of a Tomato Landrace as Rootstock Improves the Response of Commercial Tomato under Water Deficit Conditions. Agronomy 2020, 10, 748. https://doi.org/10.3390/agronomy10050748
Fullana-Pericàs M, Conesa MÀ, Ribas-Carbó M, Galmés J. The Use of a Tomato Landrace as Rootstock Improves the Response of Commercial Tomato under Water Deficit Conditions. Agronomy. 2020; 10(5):748. https://doi.org/10.3390/agronomy10050748
Chicago/Turabian StyleFullana-Pericàs, Mateu, Miquel À. Conesa, Miquel Ribas-Carbó, and Jeroni Galmés. 2020. "The Use of a Tomato Landrace as Rootstock Improves the Response of Commercial Tomato under Water Deficit Conditions" Agronomy 10, no. 5: 748. https://doi.org/10.3390/agronomy10050748
APA StyleFullana-Pericàs, M., Conesa, M. À., Ribas-Carbó, M., & Galmés, J. (2020). The Use of a Tomato Landrace as Rootstock Improves the Response of Commercial Tomato under Water Deficit Conditions. Agronomy, 10(5), 748. https://doi.org/10.3390/agronomy10050748