Differences in Metabolic and Physiological Responses between Local and Widespread Grapevine Cultivars under Water Deficit Stress

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Soil Water Content and Climatic Conditions
2.3. Plant Water Relations and Leaf Mass Area
2.4. Gas Exchange Measurements
2.5. Leaf Carbon Isotope Composition (δ13C)
2.6. Metabolite Profiling
2.7. Data and Statistical Analysis
3. Results
3.1. Physiological Traits and Responses under Well-Watered and Water Deficit Stress Treatments
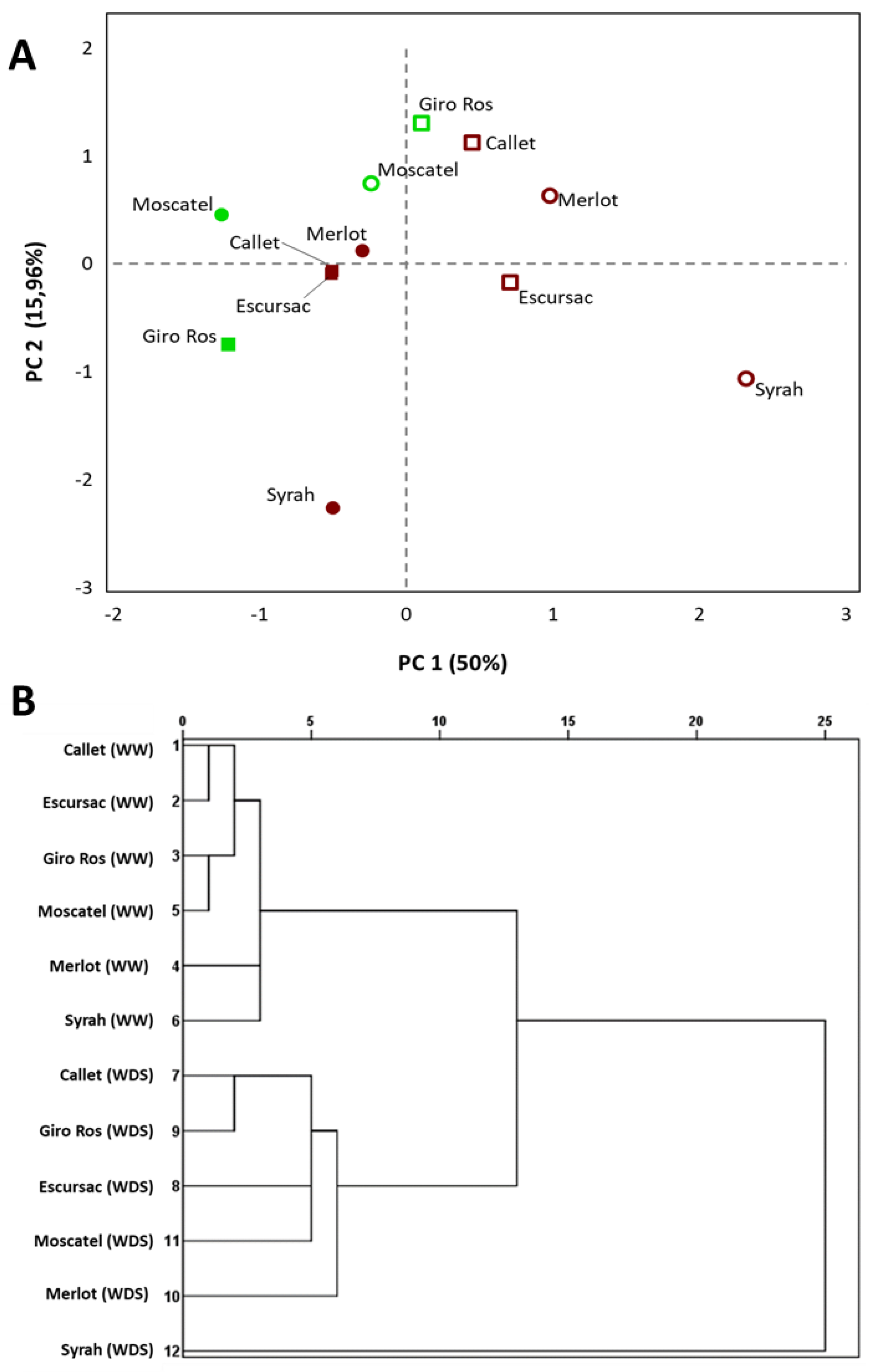
3.2. Multivariate Analysis of the Metabolite Levels under Well-Watered and Water Deficit Stress Treatments
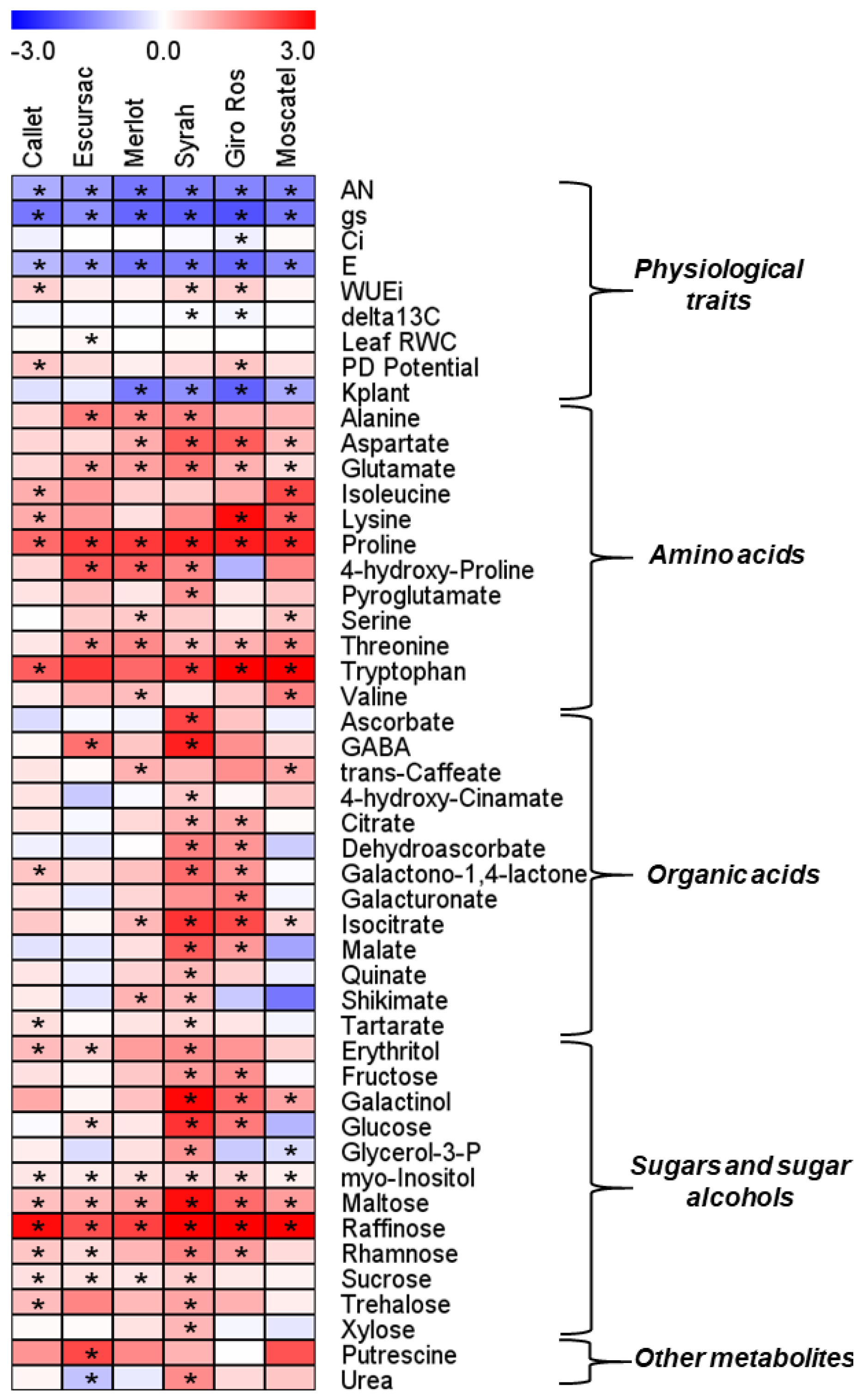
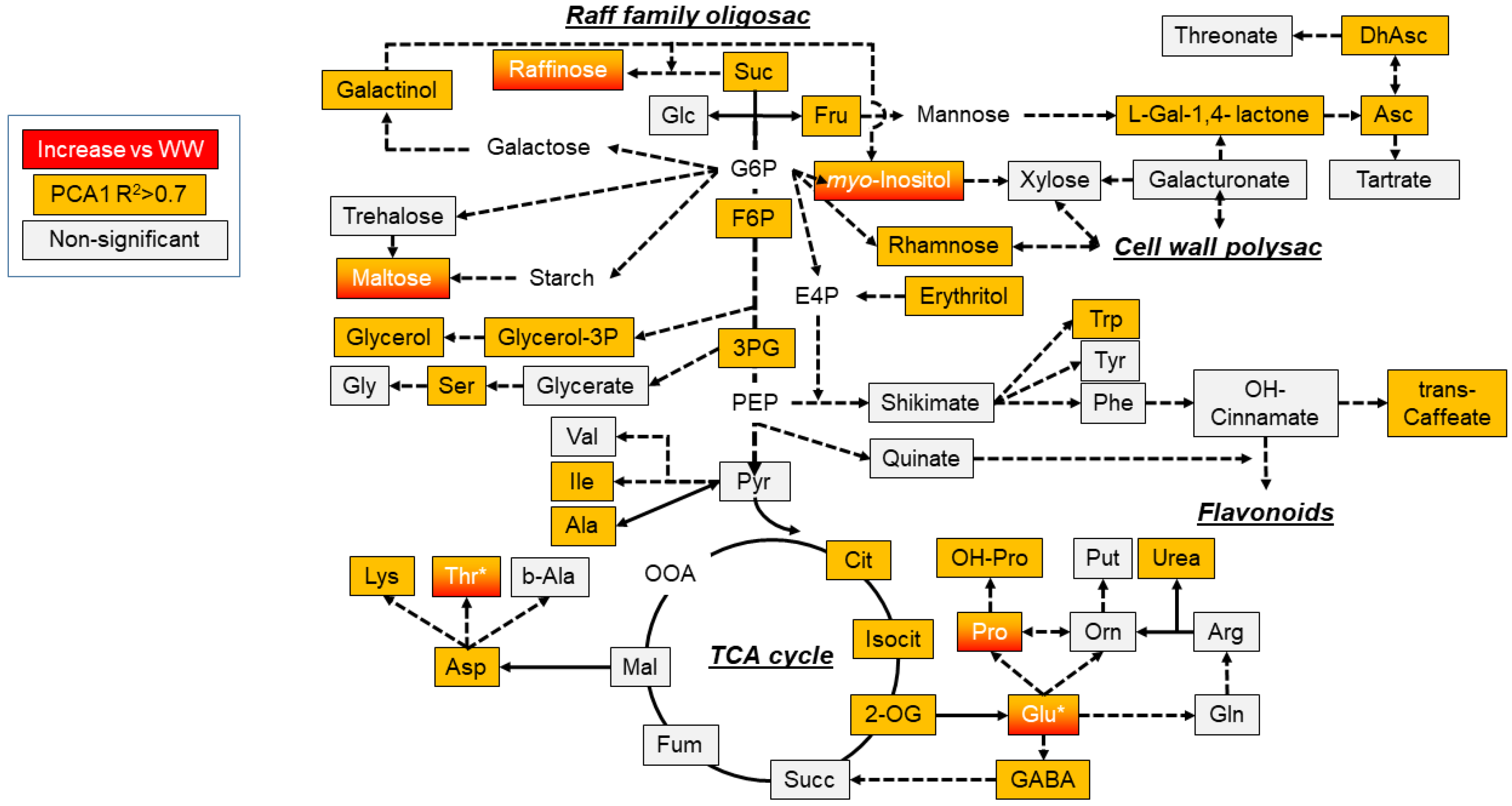
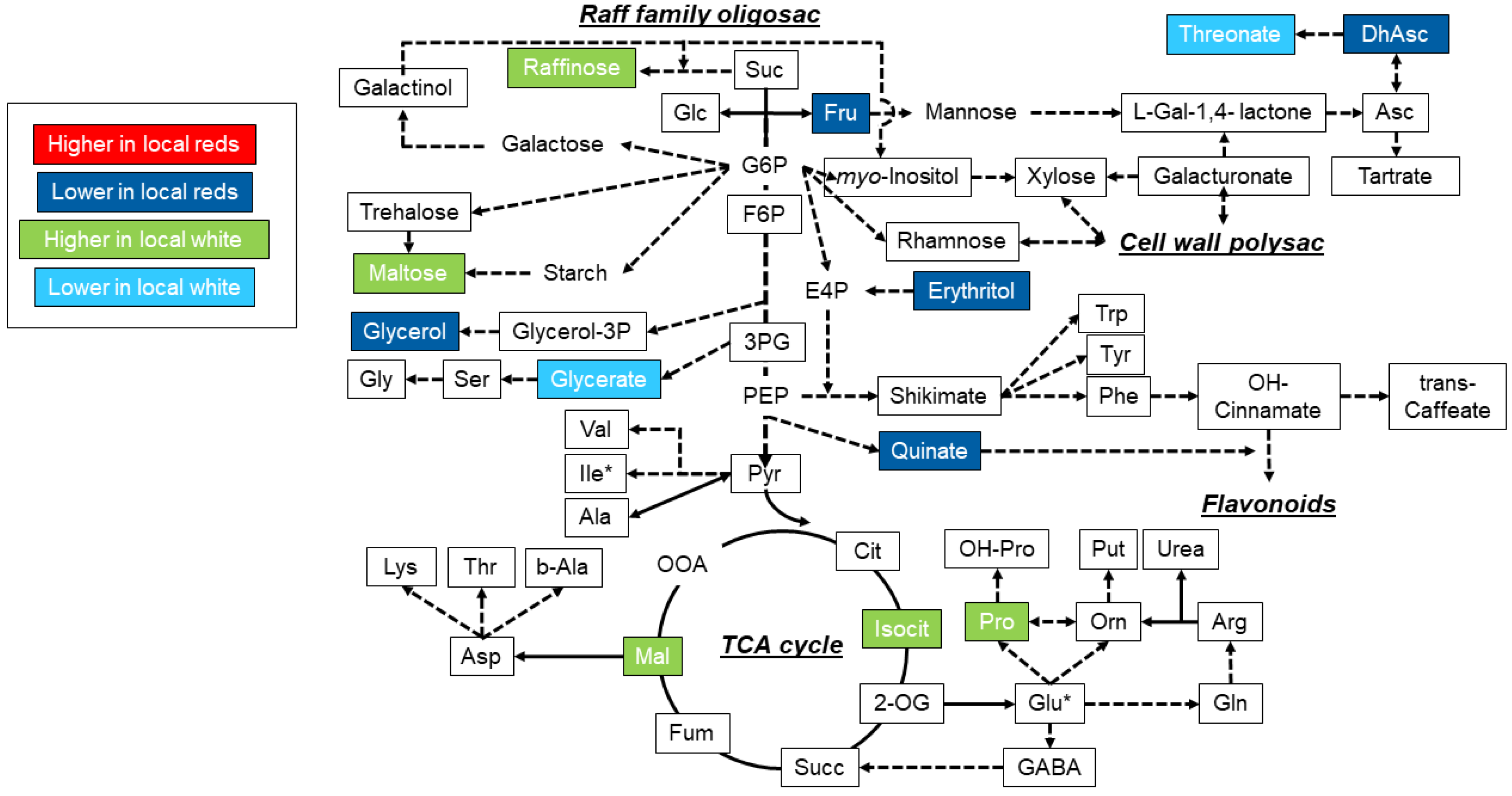
3.3. Metabolite Profiling and Responses under Well-Watered and Water Deficit Stress Treatments
3.4. Correlations between Physiological Parameters and Metabolite Levels under Water Deficit Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The Future of Food and Agriculture—Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Medrano, H.; Tomas, M.; Martorell, S.; Escalona, J.M.; Pou, A.; Fuentes, S.; Flexas, J.; Bota, J. Improving water use efficiency of vineyards in semi-arid regions. A review. Agron. Sustain. Dev. 2015, 35, 499–517. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, C.; Lavoie-Lamoureux, A.; Tramontini, S.; Ferrandino, A. Grapevine adaptations to water stress: New perspectives about soil/plant interactions. Theor. Exp. Plant Phys. 2016, 28, 53–66. [Google Scholar] [CrossRef]
- Costa, J.M.; Vaz, M.; Escalona, J.; Egipto, R.; Lopes, C.; Medrano, H.; Chaves, M.M. Modern viticulture in southern Europe: Vulnerabilities and strategies for adaptation to water scarcity. Agric. Water Manag. 2016, 164, 5–18. [Google Scholar] [CrossRef]
- Costa, J.M.; Egipto, R.; Silvestre, J.; Lopes, C.; Chaves, M.M. Water and Heat Fluxes in Mediterranean Vineyards: Indicators and Relevance for Management. In Water Scarcity and Sustainable Agriculture in Semiarid Enviroment Tools, Strategies, and Challenges for Woody Crops; Academic Press: Cambridge, MA, USA, 2018; pp. 219–245. [Google Scholar]
- Van Leeuwen, C.; Darriet, P. The Impact of Climate Change on Viticulture and Wine Quality. J. Wine Econ. 2016, 11, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Vaz, M.; Coelho, R.; Rato, A.; Samara-Lima, R.; Silva, L.L.; Campostrini, E.; Mota, J.B. Adaptive strategies of two Mediterranean grapevines varieties (Aragonez syn. Tempranillo and Trincadeira) face drought: Physiological and structural responses. Theor. Exp. Plant Phys. 2016, 28, 205–220. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; de Cortazar-Atauri, I.G.; Morales-Castilla, I.; Nicholas, K.A.; Lacombe, T. From Pinot to Xinomavro in the world’s future wine-growing regions. Nat. Clim. Chang. 2018, 8, 29–37. [Google Scholar] [CrossRef]
- Morales-Castilla, I.; Garcia de Cortazar-Atauri, I.; Cook, B.I.; Lacombe, T.; Parker, A.; van Leeuwen, C.; Nicholas, K.A.; Wolkovich, E.M. Diversity buffers winegrowing regions from climate change losses. Proc. Natl. Acad. Sci. USA 2020, 117, 2864–2869. [Google Scholar] [CrossRef]
- Bota, J.; Tomas, M.; Flexas, J.; Medrano, H.; Escalona, J.M. Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress. Agric. Water Manag. 2016, 164, 91–99. [Google Scholar] [CrossRef]
- Schultz, H.R. Water relations and photosynthetic responses of two grapevine cultivars of different geographical origin during water stress. Acta Hortic. 1996, 427, 251–266. [Google Scholar] [CrossRef]
- Schultz, H.R. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 2003, 26, 1393–1405. [Google Scholar] [CrossRef]
- Bota, J.; Flexas, J.; Medrano, H. Genetic variability of photosynthesis and water use in Balearic grapevine cultivars. Ann. Appl. Biol. 2001, 138, 353–361. [Google Scholar] [CrossRef]
- Soar, C.J.; Dry, P.R.; Loveys, B.R. Scion photosynthesis and leaf gas exchange in Vitis vinifera L. cv. Shiraz: Mediation of rootstock effects via xylem sap ABA. Aust. J. Grape Wine Res. 2006, 12, 82–96. [Google Scholar] [CrossRef]
- Tomas, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbo, M.; Flexas, J. Water-use efficiency in grapevine cultivars grown under controlled conditions: Effects of water stress at the leaf and whole-plant level. Aust. J. Grape Wine Res. 2012, 18, 164–172. [Google Scholar] [CrossRef]
- Tomas, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbo, M.; Flexas, J. Variability of water use efficiency in grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Costa, J.M.; Ortuno, M.F.; Lopes, C.M.; Chaves, M.M. Grapevine varieties exhibiting differences in stomatal response to water deficit. Funct. Plant Biol. 2012, 39, 179–189. [Google Scholar] [CrossRef]
- Pou, A.; Flexas, J.; Alsina, M.D.; Bota, J.; Carambula, C.; de Herralde, F.; Galmes, J.; Lovisolo, C.; Jimenez, M.; Ribas-Carbo, M.; et al. Adjustments of water use efficiency by stomatal regulation during drought and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri x V. rupestris). Physiol. Plant. 2008, 134, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.A.; Lebon, E.; Ojeda, H. Stomatal Behavior of Different Grapevine Cultivars in Response to Soil Water Status and Air Water Vapor Pressure Deficit. J. Int. Sci. Vigne Vin 2010, 44, 9–20. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medrano, H.; Tortosa, I.; Montes, E.; Pou, A.; Balda, P.; Bota, J.; Escalona, J.M. Genetic improvement of grapevine (Vitis vinifera L.) water use efficiency. In Water Scarcity and Sustainable Agriculture in Semiarid Environment. Tools, Strategies and Challenges for Woody Crops; Academic Press: Cambridge, MA, USA, 2018; pp. 377–401. [Google Scholar]
- Pou, A.; Medrano, H.; Tomas, M.; Martorell, S.; Ribas-Carbo, M.; Flexas, J. Anisohydric behaviour in grapevines results in better performance under moderate water stress and recovery than isohydric behaviour. Plant Soil 2012, 359, 335–349. [Google Scholar] [CrossRef]
- Klein, T. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Funct. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Garcia-Forner, N. Water potential regulation, stomatal behaviour and hydraulic transport under drought: Deconstructing the iso/anisohydric concept. Plant Cell Environ. 2017, 40, 962–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/Anisohydry: A Plant-Environment Interaction Rather Than a Simple Hydraulic Trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef]
- Habsburg-Lorena, L.S. Die Eigentlichen Balearen [1871] Die Balearen in Wort und Bild geschildert. In Brockhaus. Traducción castellana (1982–1993): Las Baleares por la palabra y el grabado; Caja de Ahorros de Baleares “Sa Nostra”: Palma de Mallorca, Spain; Leipzig, Germany, 1989. [Google Scholar]
- Garcia-Munoz, S.; Lacombe, T.; de Andres, M.T.; Gaforio, L.; Munoz-Organero, G.; Laucou, V.; This, P.; Cabello, F. Grape varieties (Vitis vinifera L.) from the Balearic Islands: Genetic characterization and relationship with Iberian Peninsula and Mediterranean Basin. Genet. Resour. Crop Evol. 2012, 59, 589–605. [Google Scholar] [CrossRef]
- Marsal, G.; Bota, J.; Martorell, A.; Canals, J.M.; Zamora, F.; Fort, F. Local cultivars of Vitis vinifera L. in Spanish islands: Balearic Archipelago. Sci. Hortic. 2017, 226, 122–132. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Vidigal, P.; Amancio, S. Oxidative stress homeostasis in grapevine (Vitis vinifera L.). Front. Environ. Sci. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Obata, T.; Fernie, A.R. The use of metabolomics to dissect plant responses to abiotic stresses. Cell. Mol. Life Sci. 2012, 69, 3225–3243. [Google Scholar] [CrossRef] [Green Version]
- Peters, K.; Worrich, A.; Weinhold, A.; Alka, O.; Balcke, G.; Birkemeyer, C.; Bruelheide, H.; Calf, O.W.; Dietz, S.; Duhrkop, K.; et al. Current Challenges in Plant Eco-Metabolomics. Int. J. Mol. Sci. 2018, 19, 1385. [Google Scholar] [CrossRef] [Green Version]
- Alseekh, S.; Fernie, A.R. Metabolomics 20years on: What have we learned and what hurdles remain? Plant J. 2018, 94, 933–942. [Google Scholar] [CrossRef]
- Flexas, J.; Gago, J. A role for ecophysiology in the ‘omics’ era. Plant J. 2018, 96, 251–259. [Google Scholar] [CrossRef]
- Clemente-Moreno, M.J.; Omranian, N.; Saez, P.; Figueroa, C.M.; Del-Saz, N.; Elso, M.; Poblete, L.; Orf, I.; Cuadros-Inostroza, A.; Cavieres, L.; et al. Cytochrome respiration pathway and sulphur metabolism sustain stress tolerance to low temperature in the Antarctic species Colobanthus quitensis. New Phytol. 2020, 225, 754–768. [Google Scholar] [CrossRef]
- Clemente-Moreno, M.J.; Omranian, N.; Saez, P.L.; Figueroa, C.M.; Del-Saz, N.; Elso, M.; Poblete, L.; Orf, I.; Cuadros-Inostroza, A.; Cavieres, L.A.; et al. Low-temperature tolerance of the Antarctic species Deschampsia antarctica: A complex metabolic response associated with nutrient remobilization. Plant Cell Environ. 2020, 43, 1376–1393. [Google Scholar] [CrossRef] [PubMed]
- De Ollas, C.; Morillon, R.; Fotopoulos, V.; Puertolas, J.; Ollitrault, P.; Gomez-Cadenas, A.; Arbona, V. Facing Climate Change: Biotechnology of Iconic Mediterranean Woody Crops. Front. Plant Sci. 2019, 10, 427. [Google Scholar] [CrossRef] [PubMed]
- Conde, A.; Regalado, A.; Rodrigues, D.; Costa, J.M.; Blumwald, E.; Chaves, M.M.; Geros, H. Polyols in grape berry: Transport and metabolic adjustments as a physiological strategy for water-deficit stress tolerance in grapevine. J. Exp. Bot. 2015, 66, 889–906. [Google Scholar] [CrossRef] [Green Version]
- Savoi, S.; Wong, D.C.J.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 2016, 16, 67. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, U.; Batushansky, A.; Degu, A.; Rachmilevitch, S.; Fait, A. Metabolic and Physiological Responses of Shiraz and Cabernet Sauvignon (Vitis vinifera L.) to Near Optimal Temperatures of 25 and 35 degrees C. Int. J. Mol. Sci. 2015, 16, 24276–24294. [Google Scholar] [CrossRef] [Green Version]
- Ghan, R.; Van Sluyter, S.C.; Hochberg, U.; Degu, A.; Hopper, D.W.; Tillet, R.L.; Schlauch, K.A.; Haynes, P.A.; Fait, A.; Cramer, G.R. Five omic technologies are concordant in differentiating the biochemical characteristics of the berries of five grapevine (Vitis vinifera L.) cultivars. BMC Genom. 2015, 16, 946. [Google Scholar] [CrossRef] [Green Version]
- Pinasseau, L.; Vallverdu-Queralt, A.; Verbaere, A.; Roques, M.; Meudec, E.; Le Cunff, L.; Peros, J.P.; Ageorges, A.; Sommerer, N.; Boulet, J.C.; et al. Cultivar Diversity of Grape Skin Polyphenol Composition and Changes in Response to Drought Investigated by LC-MS Based Metabolomics. Front. Plant Sci. 2017, 8, 1826. [Google Scholar] [CrossRef] [Green Version]
- Degu, A.; Hochberg, U.; Wong, D.C.J.; Alberti, G.; Lazarovitch, N.; Peterlunger, E.; Castellarin, S.D.; Herrera, J.C.; Fait, A. Swift metabolite changes and leaf shedding are milestones in the acclimation process of grapevine under prolonged water stress. BMC Plant Biol. 2019, 19, 69. [Google Scholar] [CrossRef]
- Hochberg, U.; Degu, A.; Fait, A.; Rachmilevitch, S. Near isohydric grapevine cultivar displays higher photosynthetic efficiency and photorespiration rates under drought stress as compared with near anisohydric grapevine cultivar. Physiol. Plant. 2013, 147, 443–452. [Google Scholar] [CrossRef] [PubMed]
- El Aou-ouad, H.; Bota, J.; Obata, T.; Montero, R.; Fernie, A.R.; Medrano, H.; Pou, A.; Florez-Sarasa, I. Combined drought and virus infection trigger aspects of respiratory metabolism related to grapevine physiological responses. J. Plant Physiol. 2018, 231, 19–30. [Google Scholar] [CrossRef]
- Bota, J.; Stasyk, O.; Flexas, J.; Medrano, H. Effect of water stress on partitioning of C-14-labelled photosynthates in Vitis vinifera. Funct. Plant Biol. 2004, 31, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulias, J.; Flexas, J. Regulation of photosynthesis of C-3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- El Aou-ouad, H.; Pou, A.; Tomas, M.; Montero, R.; Ribas-Carbo, M.; Medrano, H.; Bota, J. Combined effect of virus infection and water stress on water flow and water economy in grapevines. Physiol. Plant. 2017, 160, 171–184. [Google Scholar] [CrossRef]
- Groom, P.K.; Lamont, B.B. Which common indices of sclerophylly best reflect differences in leaf structure? Ecoscience 1999, 6, 471–474. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic Composition of Plant Carbon Correlates with Water-Use Efficiency of Wheat Genotypes. Aust. J. Plant Physiol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef]
- Luedemann, A.; Strassburg, K.; Erban, A.; Kopka, J. TagFinder for the quantitative analysis of gas chromatography - mass spectrometry (GC-MS)-based metabolite profiling experiments. Bioinformatics 2008, 24, 732–737. [Google Scholar] [CrossRef]
- Fernie, A.R.; Aharoni, A.; Willmitzer, L.; Stitt, M.; Tohge, T.; Kopka, J.; Carroll, A.J.; Saito, K.; Fraser, P.D.; DeLuca, V. Recommendations for Reporting Metabolite Data. Plant Cell 2011, 23, 2477–2482. [Google Scholar] [CrossRef] [Green Version]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. MeV: MultiExperiment Viewer. In Biomedical Informatics for Cancer Research; Springer: Boston, MA, USA, 2010; pp. 267–277. [Google Scholar] [CrossRef]
- Florez-Sarasa, I.; Ribas-Carbo, M.; Del-Saz, N.F.; Schwahn, K.; Nikoloski, Z.; Fernie, A.R.; Flexas, J. Unravelling the invivo regulation and metabolic role of the alternative oxidase pathway in C-3 species under photoinhibitory conditions. New Phytol. 2016, 212, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Martorell, S.; Diaz-Espejo, A.; Tomas, M.; Pou, A.; El Aou-Ouad, H.; Escalona, J.M.; Vadell, J.; Ribas-Carbo, M.; Flexas, J.; Medrano, H. Differences in water-use-efficiency between two Vitis vinifera cultivars (Grenache and Tempranillo) explained by the combined response of stomata to hydraulic and chemical signals during water stress. Agric. Water Manag. 2015, 156, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Scoffoni, C.; Gago, J.; Sack, L. Leaf mesophyll conductance and leaf hydraulic conductance: An introduction to their measurement and coordination. J. Exp. Bot. 2013, 64, 3965–3981. [Google Scholar] [CrossRef]
- Escalona, J.M.; Flexas, J.; Medrano, H. Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines. Aust. J. Plant Physiol. 1999, 26, 421–433. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565. [Google Scholar]
- Rao, R.C.N.; Wright, G.C. Stability of the Relationship between Specific Leaf-Area and Carbon-Isotope Discrimination across Environments in Peanut. Crop Sci. 1994, 34, 98–103. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nesi, A.N.; Araujo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Degu, A.; Morcia, C.; Tumino, G.; Hochberg, U.; Toubiana, D.; Mattivi, F.; Schneider, A.; Bosca, P.; Cattivelli, L.; Terzi, V.; et al. Metabolite profiling elucidates communalities and differences in the polyphenol biosynthetic pathways of red and white Muscat genotypes. Plant Physiol. Biochem. 2015, 86, 24–33. [Google Scholar] [CrossRef]
- Massonnet, M.; Fasoli, M.; Tornielli, G.B.; Altieri, M.; Sandri, M.; Zuccolotto, P.; Paci, P.; Gardiman, M.; Zenoni, S.; Pezzotti, M. Ripening Transcriptomic Program in Red and White Grapevine Varieties Correlates with Berry Skin Anthocyanin Accumulation. Plant Physiol. 2017, 174, 2376–2396. [Google Scholar] [CrossRef] [Green Version]
- Rooy, S.S.B.; Salekdeh, G.H.; Ghabooli, M.; Gholami, M.; Karimi, R. Cold-induced physiological and biochemical responses of three grapevine cultivars differing in cold tolerance. Acta Physiol. Plant. 2017, 39, 264. [Google Scholar] [CrossRef]
- Zhang, Y.; Butelli, E.; Alseekh, S.; Tohge, T.; Rallapalli, G.; Luo, J.; Kawar, P.G.; Hill, L.; Santino, A.; Fernie, A.R.; et al. Multi-level engineering facilitates the production of phenylpropanoid compounds in tomato. Nat. Commun. 2015, 6, 8635. [Google Scholar] [CrossRef] [Green Version]
- Soubeyrand, E.; Colombie, S.; Beauvoit, B.; Dai, Z.W.; Cluzet, S.; Hilbert, G.; Renaud, C.; Maneta-Peyret, L.; Dieuaide-Noubhani, M.; Merillon, J.M.; et al. Constraint-Based Modeling Highlights Cell Energy, Redox Status and alpha-Ketoglutarate Availability as Metabolic Drivers for Anthocyanin Accumulation in Grape Cells Under Nitrogen Limitation. Front. Plant Sci. 2018, 9, 421. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Dudareva, N. The Shikimate Pathway and Aromatic Amino Acid Biosynthesis in Plants. Annu Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Clemente-Moreno, M.J.; Gago, J.; Daz-Vivancos, P.; Bernal, A.; Miedes, E.; Bresta, P.; Liakopoulos, G.; Fernie, A.R.; Hernndez, J.A.; Flexas, J. The apoplastic antioxidant system and altered cell wall dynamics influence mesophyll conductance and the rate of photosynthesis. Plant J. 2019, 99, 1031–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig-Oliver, M.; Nadal, M.; Clemente-Moreno, M.J.; Bota, J.; Flexas, J. Cell wall components regulate photosynthesis and leaf water relations of Vitis vinifera cv. Grenache acclimated to contrasting environmental conditions. J. Plant Physiol. 2020, 244, 153084. [Google Scholar] [CrossRef]
- Flamini, R.; Mattivi, F.; De Rosso, M.; Arapitsas, P.; Bavaresco, L. Advanced Knowledge of Three Important Classes of Grape Phenolics: Anthocyanins, Stilbenes and Flavonols. Int. J. Mol. Sci. 2013, 14, 19651–19669. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and Glutathione: The Heart of the Redox Hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Ribas-Carbo, M.; Taylor, N.L.; Giles, L.; Busquets, S.; Finnegan, P.M.; Day, D.A.; Lambers, H.; Medrano, H.; Berry, J.A.; Flexas, J. Effects of water stress on respiration in soybean leaves. Plant Physiol. 2005, 139, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Del-Saz, N.F.; Florez-Sarasa, I.; Clemente-Moreno, M.J.; Mhadhbi, H.; Flexas, J.; Fernie, A.R.; Ribas-Carbo, M. Salinity tolerance is related to cyanide-resistant alternative respiration in Medicago truncatula under sudden severe stress. Plant Cell Environ. 2016, 39, 2361–2369. [Google Scholar] [CrossRef]
- Sanchez-Guerrero, A.; del-Saz, N.F.; Florez-Sarasa, I.; Ribas-Carbo, M.; Fernier, A.R.; Jimenez, A.; Sevilla, F. Coordinated responses of mitochondrial antioxidative enzymes, respiratory pathways and metabolism in Arabidopsis thaliana thioredoxin trxo1 mutants under salinity. Environ. Exp. Bot. 2019, 162, 212–222. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Callet | Escursac | Merlot | Syrah | Giro Ros | Moscatel | ||
|---|---|---|---|---|---|---|---|
| AN | WW | 17.68 ± 0.86de | 17.55 ± 0.61de | 15.47 ± 1.01d | 18.28 ± 0.62e | 17.12 ± 0.62de | 16.9 ± 0.84de |
| WDS | 9.08 ± 2.03c | 7.85 ± 0.46bc | 5.14 ± 0.36a | 6.63 ± 0.68abc | 6.34 ± 0.09ab | 6.49 ± 0.59abc | |
| gs | WW | 0.32 ± 0.03c | 0.29 ± 0.02b | 0.22 ± 0.02b | 0.32 ± 0.02c | 0.26 ± 0.02bc | 0.28 ± 0.02bc |
| WDS | 0.1 ± 0.02a | 0.12 ± 0.02a | 0.06 ± 0a | 0.09 ± 0.02a | 0.06 ± 0a | 0.1 ± 0a | |
| Ci | WW | 262.26 ± 5.55bc | 256.78 ± 4.3bc | 240.95 ± 7.87abc | 258.9 ± 3.12bc | 248.1 ± 5.31bc | 255.87 ± 6.26bc |
| WDS | 233.09 ± 15.53ab | 260.99 ± 11.82bc | 246.38 ± 12.81abc | 246.65 ± 15.35abc | 218.8 ± 4.85a | 267.82 ± 6.63c | |
| E | WW | 4.68 ± 0.33c | 4.51 ± 0.14c | 3.82 ± 0.35c | 4.7 ± 0.18c | 4.23 ± 0.26c | 4.32 ± 0.24c |
| WDS | 2.64 ± 0.62b | 2.13 ± 0.29ab | 1.28 ± 0.09a | 1.64 ± 0.33a | 1.26 ± 0.03a | 1.71 ± 0.09a | |
| WUEi | WW | 57.32 ± 3.94a | 60.84 ± 2.53a | 73.07 ± 5.49ab | 58.72 ± 2.5a | 66.65 ± 3.95ab | 62.21 ± 4.14a |
| WDS | 84.43 ± 6.51bc | 70.31 ± 7.8ab | 82.5 ± 7.57bc | 80.85 ± 10.41b | 98.91 ± 3.06c | 67.85 ± 3.52ab | |
| δ13C | WW | −28.94 ± 0.42a | −28.35 ± 0.38ab | −27.31 ± 0.29bc | −28.31 ± 0.17ab | −28.45 ± 0.24ab | −27.1 ± 0.52bc |
| WDS | −27.22 ± 0.7bc | −27.18 ± 0.38bc | −26.46 ± 0.23c | −27.36 ± 0.39bc | −26.62 ± 0.41c | −26.82 ± 0.52c | |
| RWC | WW | 88.97 ± 0.5abc | 85.91 ± 1.11a | 90.33 ± 1.44abc | 87.14 ± 0.6ab | 87.05 ± 1.64ab | 90.59 ± 0.87bc |
| WDS | 92.92 ± 2.19c | 92.2 ± 1.01c | 91.27 ± 1.19bc | 89.92 ± 1.43abc | 87.66 ± 0.74ab | 91.38 ± 0.67bc | |
| ΨPD | WW | −0.25 ± 0.03e | −0.28 ± 0.02cde | −0.39 ± 0.04abcd | −0.35 ± 0.03bcde | −0.27 ± 0.03de | −0.26 ± 0.03de |
| WDS | −0.4 ± 0.03abc | −0.38 ± 0.04abcde | −0.45 ± 0.05ab | −0.48 ± 0.05a | −0.43 ± 0.01ab | −0.33 ± 0.05bcde | |
| ΨMid | WW | −1.22 ± 0.09abc | −1.26 ± 0.04ab | −1.24 ± 0.08abc | −1.3 ± 0.07ab | −1.12 ± 0.05abc | −1.07 ± 0.04bc |
| WDS | −1.12 ± 0.02abc | −1.18 ± 0.17abc | −1.31 ± 0.07ab | −1.39 ± 0.1a | −1.37 ± 0.15ab | −0.96 ± 0.03c | |
| Kplant | WW | 4.84 ± 0.21de | 4.66 ± 0.45de | 4.69 ± 0.6de | 5 ± 0.21de | 4.98 ± 0.19de | 5.41 ± 0.31e |
| WDS | 3.75 ± 1bcde | 3.89 ± 1.82cde | 1.6 ± 0.32ab | 2.07 ± 0.82abc | 1.39 ± 0.19a | 2.73 ± 0.2abcd | |
| LMA | WW | 90.4 ± 2.69bc | 62.3 ± 7a | 78.8 ± 3.22ab | 74.63 ± 5.47ab | 84.3 ± 1.14bc | 77.67 ± 6.41ab |
| WDS | 97.83 ± 3.88c | 82 ± 3.92bc | 76.75 ± 3.37ab | 76.5 ± 5.51ab | 82 ± 3.18bc | 78 ± 2.75ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Florez-Sarasa, I.; Clemente-Moreno, M.J.; Cifre, J.; Capó, M.; Llompart, M.; Fernie, A.R.; Bota, J. Differences in Metabolic and Physiological Responses between Local and Widespread Grapevine Cultivars under Water Deficit Stress. Agronomy 2020, 10, 1052. https://doi.org/10.3390/agronomy10071052
Florez-Sarasa I, Clemente-Moreno MJ, Cifre J, Capó M, Llompart M, Fernie AR, Bota J. Differences in Metabolic and Physiological Responses between Local and Widespread Grapevine Cultivars under Water Deficit Stress. Agronomy. 2020; 10(7):1052. https://doi.org/10.3390/agronomy10071052
Chicago/Turabian StyleFlorez-Sarasa, Igor, María José Clemente-Moreno, Josep Cifre, Miquel Capó, Miquel Llompart, Alisdair R. Fernie, and Josefina Bota. 2020. "Differences in Metabolic and Physiological Responses between Local and Widespread Grapevine Cultivars under Water Deficit Stress" Agronomy 10, no. 7: 1052. https://doi.org/10.3390/agronomy10071052
APA StyleFlorez-Sarasa, I., Clemente-Moreno, M. J., Cifre, J., Capó, M., Llompart, M., Fernie, A. R., & Bota, J. (2020). Differences in Metabolic and Physiological Responses between Local and Widespread Grapevine Cultivars under Water Deficit Stress. Agronomy, 10(7), 1052. https://doi.org/10.3390/agronomy10071052