Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions

 ,
, 
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Layout and Growth Conditions
2.2. Calculations of Water Regimes
2.3. Foliar Application and Sampling
2.4. Studied Parameters
2.4.1. Vegetative Growth
2.4.2. Leaf Relative Water Content (RWC)
2.4.3. Membranes Lipid Peroxidation
2.4.4. Proline and Soluble Sugars
2.4.5. Determination of Mineral Nutrients
2.4.6. Determination of Carotenoids, Ascorbic Acid and Total Soluble Phenols
2.4.7. Quantification of Antioxidant Enzymes
2.4.8. Determination of Yield Parameters
2.4.9. Statistical Analysis
3. Results
3.1. The Main Effects of the Irrigation Levels and K2SiO3 Foliar Applications
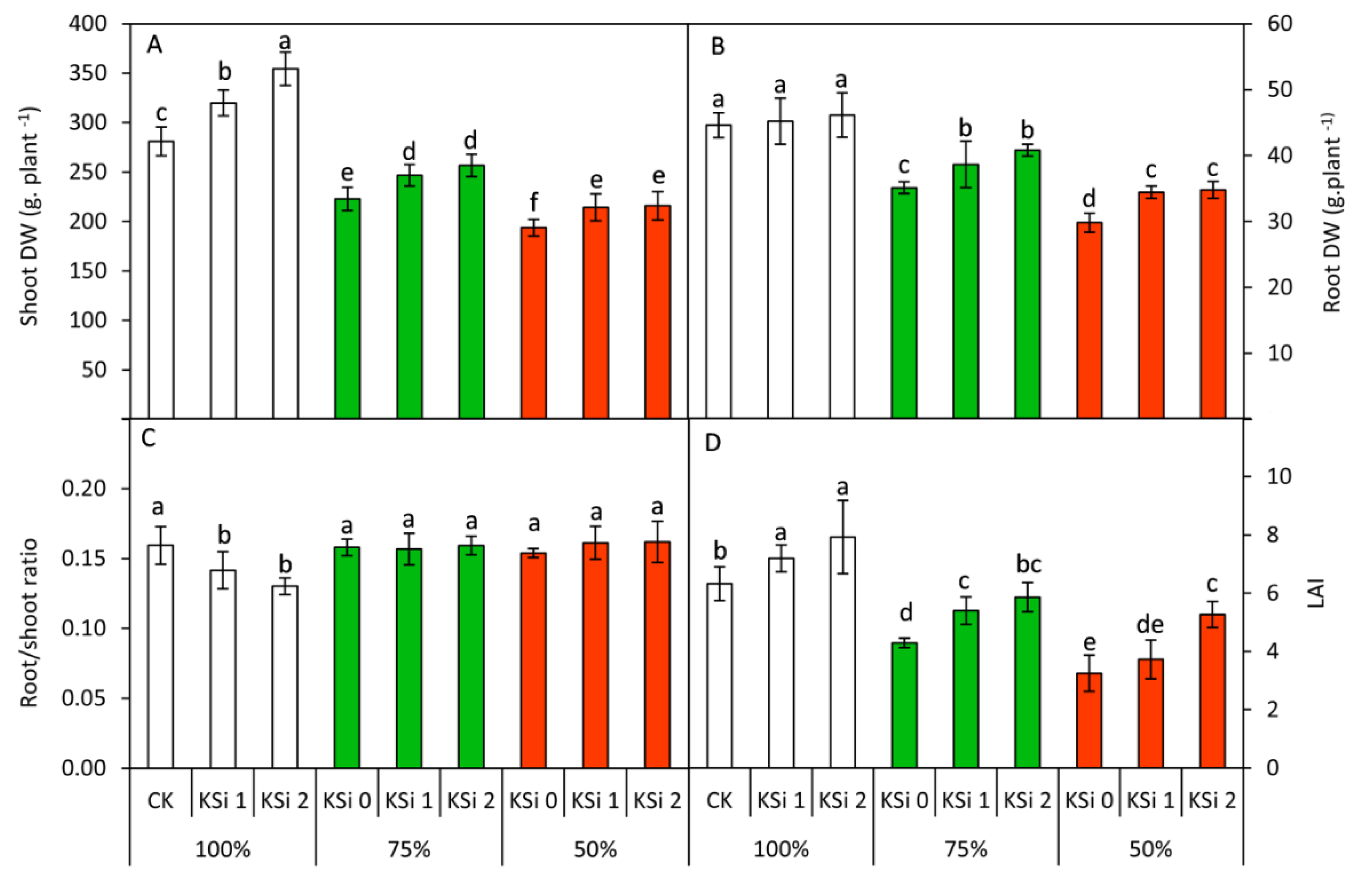
3.2. Changes in Plant Growth
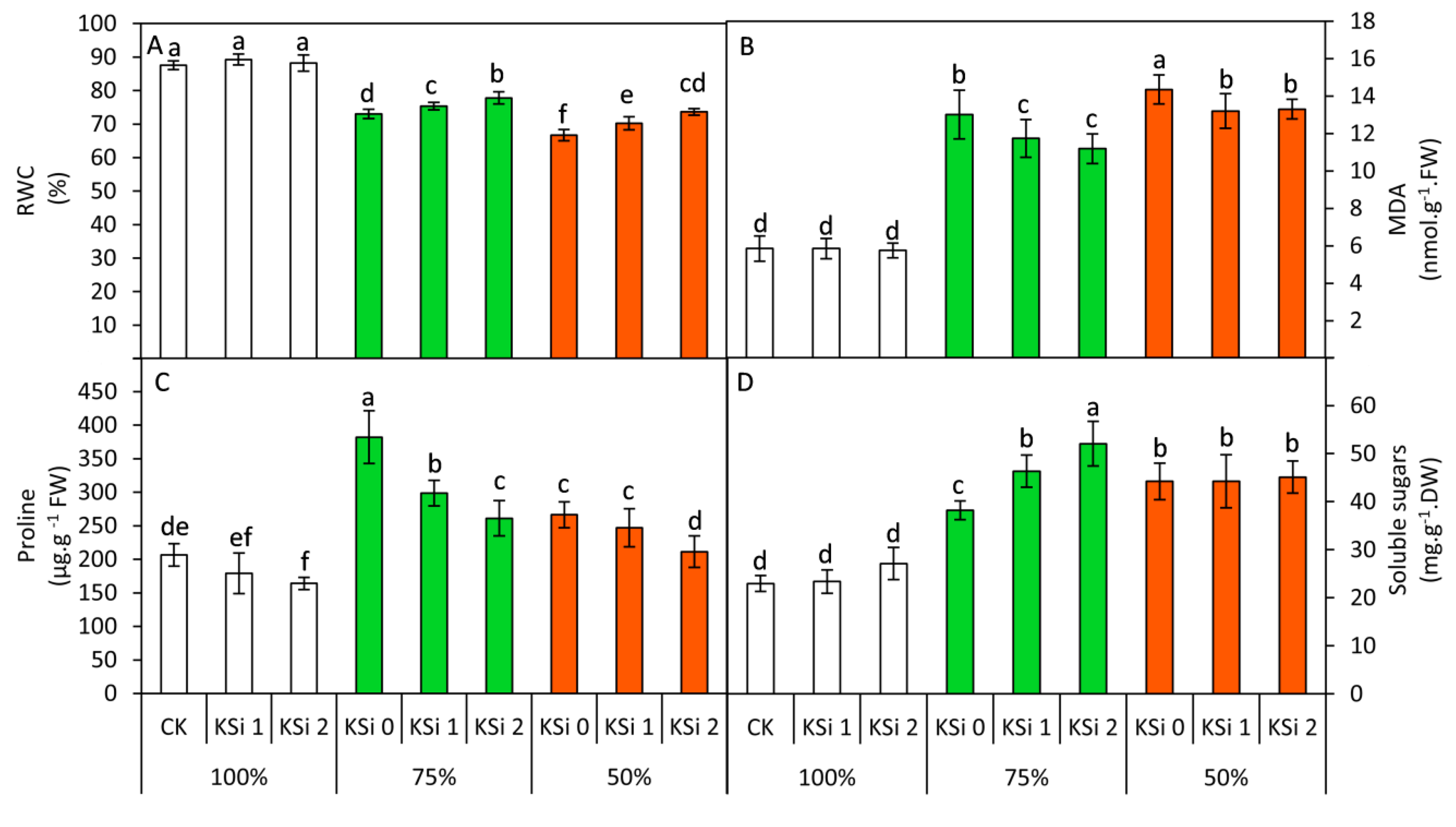
3.3. Changes in RWC, MDA, Proline and Soluble Sugars
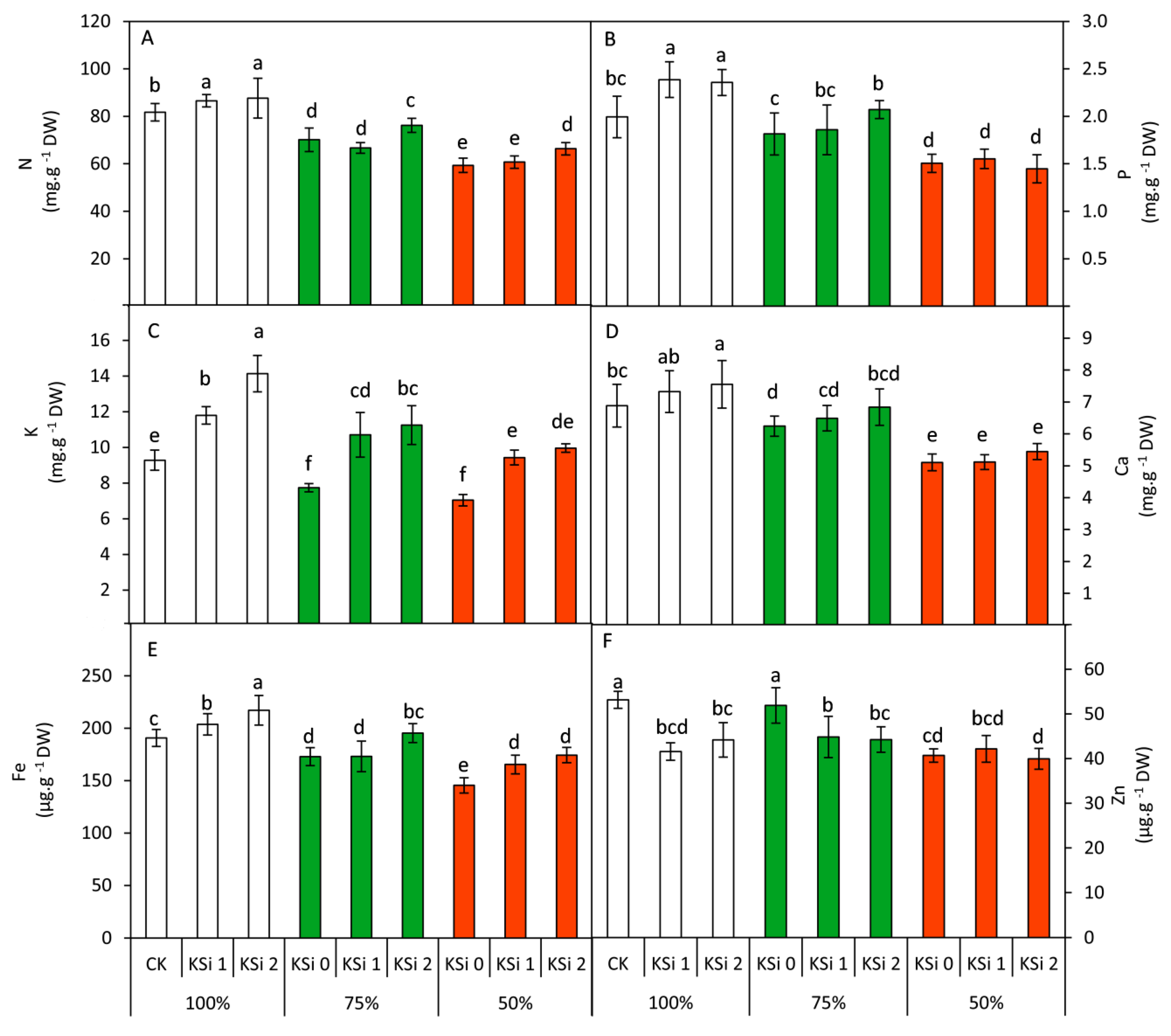
3.4. Changes in Mineral Nutrients
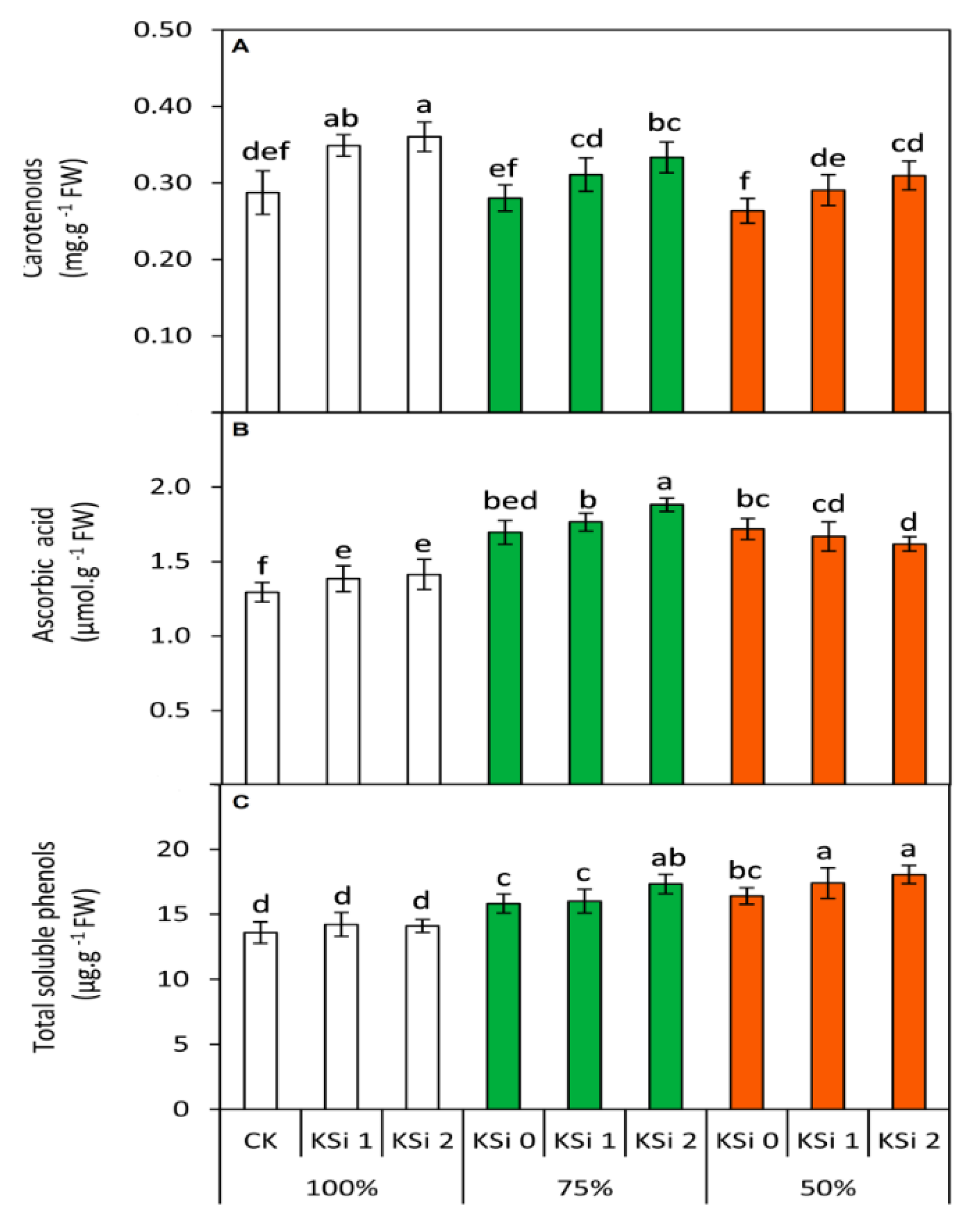
3.5. Changes in Nonenzymatic Antioxidants
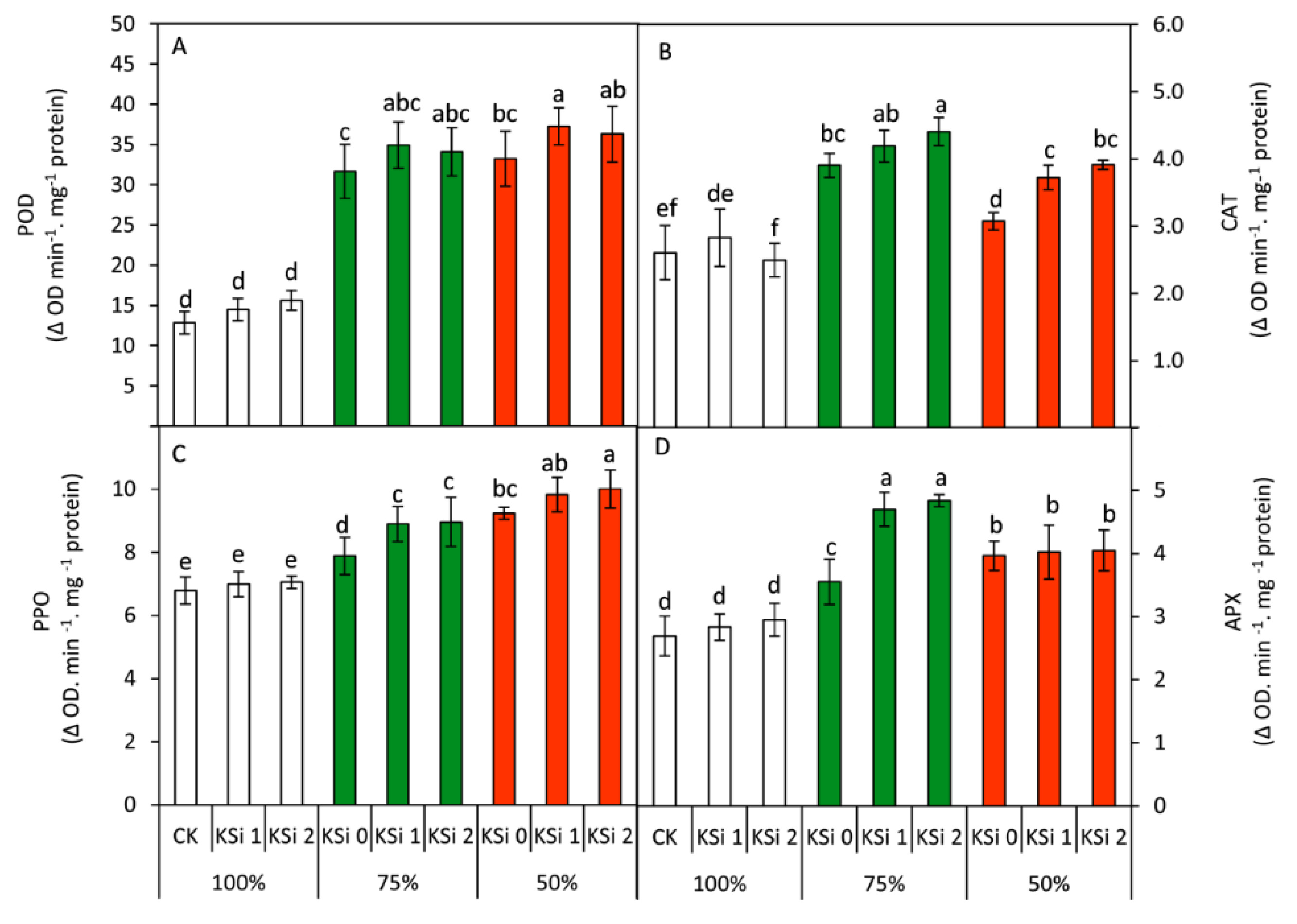
3.6. Changes in Antioxidant Enzymes
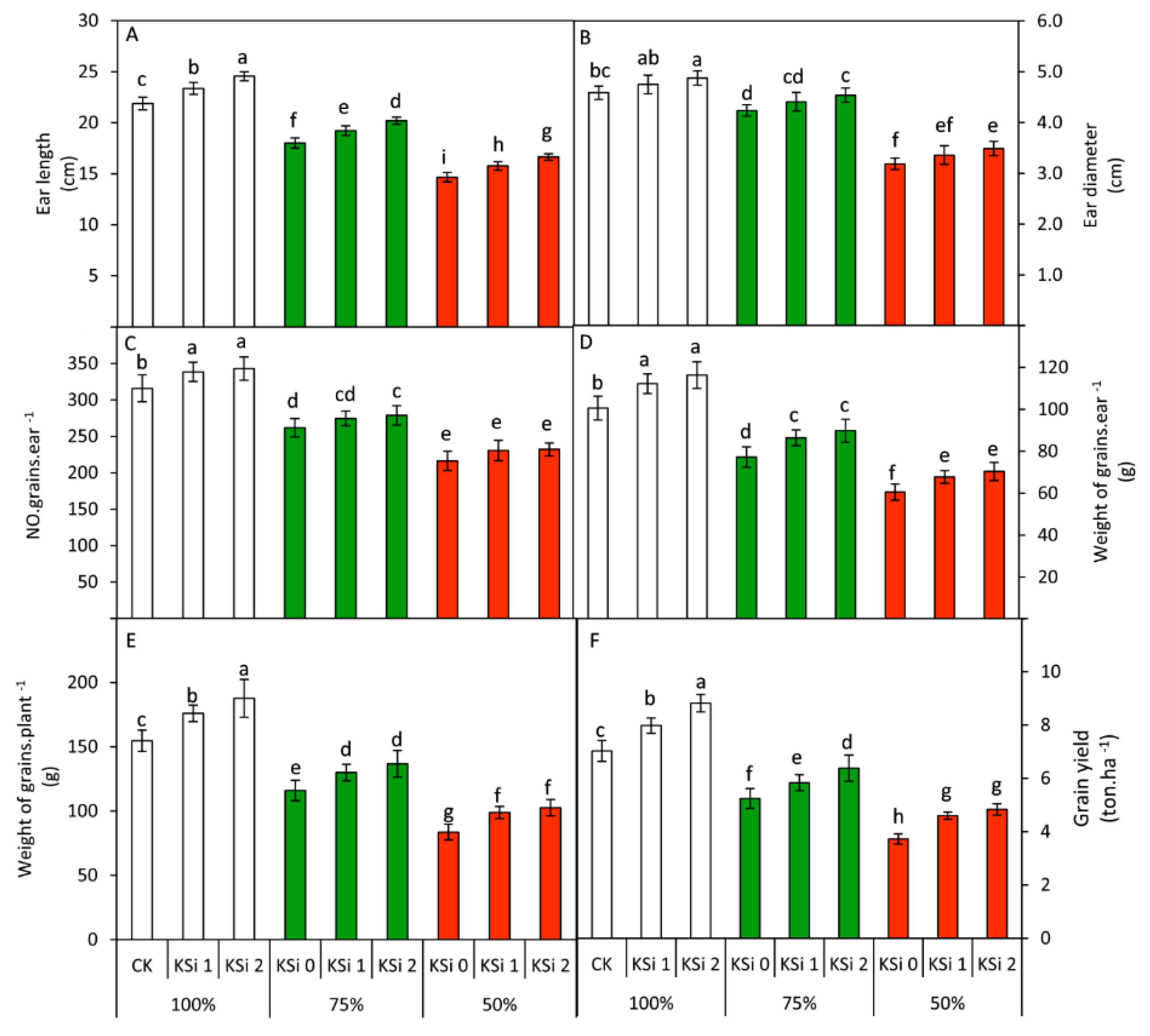
3.7. Changes in Yield Parameters
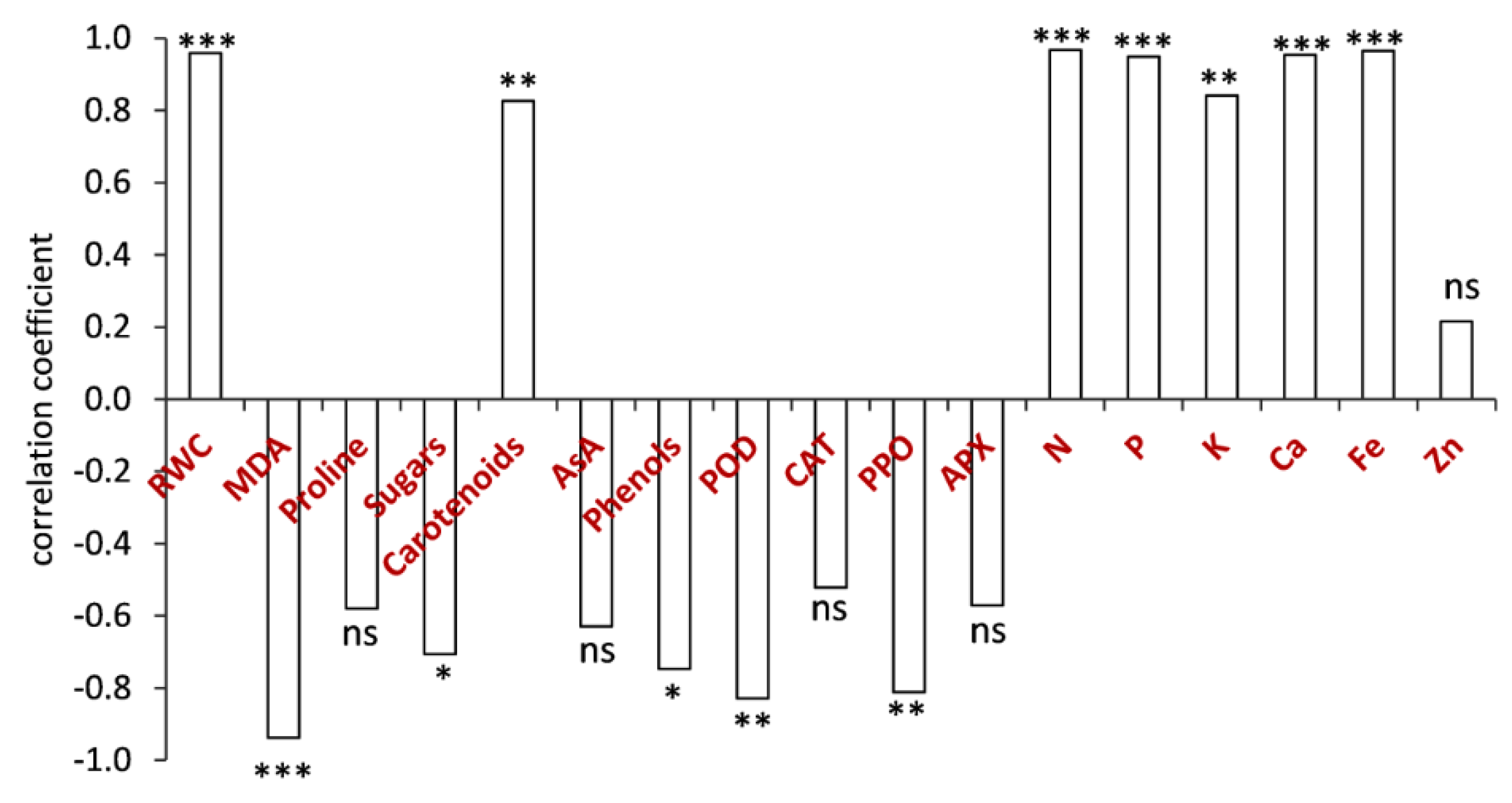
3.8. Relationships between Grain Yield and RWC, MDA, Osmolytes, Nutrients and Antioxidants
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gallart, F.; Solé, A.; Puigdefábregas, J.; Lázaro, R. Badland Systems in the Mediterranean; John Wiley & Sons, Ltd.: Chichester, UK, 2002; pp. 299–326. [Google Scholar]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkeilsh, A.; Awad, Y.M.; Soliman, M.H.; Abu-Elsaoud, A.; Abdelhamid, M.T.; El-Metwally, I.M. Exogenous application of β-sitosterol mediated growth and yield improvement in water-stressed wheat (Triticum aestivum) involves up-regulated antioxidant system. J. Plant Res. 2019, 132, 881–901. [Google Scholar] [CrossRef]
- Alhaithloul, H.A.; Soliman, M.H.; Ameta, K.L.; El-Esawi, M.A.; Elkelish, A. Changes in Ecophysiology, Osmolytes, and Secondary Metabolites of the Medicinal Plants of Mentha piperita and Catharanthus roseus Subjected to Drought and Heat Stress. Biomolecules 2020, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, N.; Ali, Q.; Ali, S.; Javed, M.T.; Zulqurnain Haider, M.; Perveen, R.; Shahid, M.R.; Rizwan, M.; Abdel-Daim, M.M.; Elkelish, A. Use of Nitric Oxide and Hydrogen Peroxide for Better Yield of Wheat (Triticum aestivum L.) under Water Deficit Conditions: Growth, Osmoregulation, and Antioxidative Defense Mechanism. Plants 2020, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.F.M.; Bondok, A.M.; Al-Senosy, N.K.; Younis, R.A. Stimulation Some of Defense Mechanisms in Tomato Plants under Water Deficit and Tobacco mosaic virus (TMV). World J. Agric. Sci. 2015, 11, 289–302. [Google Scholar]
- Badr, A.M.; Shabana, E.F.; Senousy, H.H.; Mohammad, H.Y. Anti-inflammatory and anti-cancer effects of β-carotene, extracted from Dunaliella bardawil by milking. J. Food Agric. Environ. 2014, 12, 24–31. [Google Scholar]
- Abd Ellatif, S.; El-Sheekh, M.M.; Senousy, H.H. Role of microalgal ligninolytic enzymes in industrial dye decolorization. Int. J. Phytoremediat. 2020, 1–12. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Dawood, M.A.O.; AlKahtane, A.A.; Abdeen, A.; Abdel-Latif, H.M.R.; Senousy, H.H.; Aleya, L.; Alkahtani, S. Spirulina platensis mediated the biochemical indices and antioxidative function of Nile tilapia (Oreochromis niloticus) intoxicated with aflatoxin B1. Toxicon 2020. [Google Scholar] [CrossRef]
- Senousy, H.H.; Abd Ellatif, S.; Ali, S. Assessment of the antioxidant and anticancer potential of different isolated strains of cyanobacteria and microalgae from soil and agriculture drain water. Environ. Sci. Pollut. Res. 2020, 27, 18463–18474. [Google Scholar] [CrossRef]
- Ahmad, I.; Ahmad, B.; Boote, K.; Hoogenboom, G. Adaptation strategies for maize production under climate change for semi-arid environments. Eur. J. Agron. 2020, 115, 126040. [Google Scholar] [CrossRef]
- Ranum, P.; Peña-Rosas, J.P.; Garcia-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. USA 2014, 1312, 105–112. [Google Scholar] [CrossRef]
- Dhugga, K.S. Maize biomass yield and composition for biofuels. Crop Sci. 2007, 47, 2211–2227. [Google Scholar] [CrossRef]
- Kleinmans, J.; Densley, R.; Hurley, T.; Williams, I. BRIEF COMMUNICATION: Feed value of maize silage in New Zealand—A review. In Proceedings of the New Zealand Society of Animal Production, Adelaide, Australia, 4–7 July 2016; pp. 100–102. [Google Scholar]
- Ostrander, B.M. Maize Starch for Industrial Applications. In Industrial Crops: Breeding for BioEnergy and Bioproducts; Cruz, V.M.V., Dierig, D.A., Eds.; Springer: New York, NY, USA, 2015; pp. 171–189. [Google Scholar] [CrossRef]
- de Araujo Rufino, C.; Fernandes-Vieira, J.; Martín-Gil, J.; Abreu Junior, J.D.S.; Tavares, L.C.; Fernandes-Correa, M.; Martín-Ramos, P. Water stress influence on the vegetative period yield components of different maize genotypes. Agronomy 2018, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Mi, N.; Cai, F.; Zhang, Y.; Ji, R.; Zhang, S.; Wang, Y. Differential responses of maize yield to drought at vegetative and reproductive stages. Plant Soil Environ. 2018, 64, 260–267. [Google Scholar]
- Martineau, E.; Domec, J.-C.; Bosc, A.; Dannoura, M.; Gibon, Y.; Bénard, C.; Jordan-Meille, L. The role of potassium on maize leaf carbon exportation under drought condition. Acta Physiol. Plant. 2017, 39, 219. [Google Scholar] [CrossRef]
- Ul-Allah, S.; Ijaz, M.; Nawaz, A.; Sattar, A.; Sher, A.; Naeem, M.; Shahzad, U.; Farooq, U.; Nawaz, F.; Mahmood, K. Potassium Application Improves Grain Yield and Alleviates Drought Susceptibility in Diverse Maize Hybrids. Plants 2020, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Du, Q.; Zhao, X.; Xia, L.; Jiang, C.; Wang, X.; Han, Y.; WANG, J.; YU, H.-Q. Effects of potassium deficiency on photosynthesis, chloroplast ultrastructure, ROS, and antioxidant activities in maize (Zea mays L.). J. Integr. Agric. 2019, 18, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Cooke, J.; Leishman, M.R. Consistent alleviation of abiotic stress with silicon addition: A meta-analysis. Funct. Ecol. 2016, 30, 1340–1357. [Google Scholar] [CrossRef]
- dos Santos, M.S.; Sanglard, L.M.P.V.; Martins, S.C.V.; Barbosa, M.L.; de Melo, D.C.; Gonzaga, W.F.; DaMatta, F.M. Silicon alleviates the impairments of iron toxicity on the rice photosynthetic performance via alterations in leaf diffusive conductance with minimal impacts on carbon metabolism. Plant Physiol. Biochem. 2019, 143, 275–285. [Google Scholar] [CrossRef]
- Snyder, G.H.; Matichenkov, V.V.; Datnoff, L.E. Silicon. In Handbook of Plant Nutrition; CRC Press: Boca Raton, FL, USA, 2007; pp. 551–568. [Google Scholar]
- Cao, B.-L.; Wang, L.; Gao, S.; Xia, J.; Xu, K. Silicon-mediated changes in radial hydraulic conductivity and cell wall stability are involved in silicon-induced drought resistance in tomato. Protoplasma 2017, 254, 2295–2304. [Google Scholar] [CrossRef]
- Eneji, A.E.; Inanaga, S.; Muranaka, S.; Li, J.; Hattori, T.; An, P.; Tsuji, W. Growth and nutrient use in four grasses under drought stress as mediated by silicon fertilizers. J. Plant Nutr. 2008, 31, 355–365. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Liu, P.; Wang, W.; Cao, D.; Deng, X.; Zhang, S. Silicon-mediated changes in polyamine and 1-aminocyclopropane-1-carboxylic acid are involved in silicon-induced drought resistance in Sorghum bicolor L. Plant Physiol. Biochem. 2014, 80, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Doorenbos, J. Guidelines for Predicting Crop Water Requirements; Food and Agriculture organization: Rome, Italy, 1975; Volume 24. [Google Scholar]
- Iqbal, A.; Hidayat, Z. Potassium management for improving growth and grain yield of maize (zea mays L.) under moisture stress condition. Sci. Rep. 2016, 6, 34627. [Google Scholar]
- Ünyayar, S.; Keleþ, Y.; Ünal, E. Proline and ABA levels in two sunflower genotypes subjected to water stress. Bulg. J. Plant Physiol. 2004, 30, 34–47. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Plummer, D.T. Practical Biochemistry; McGraw-Hill Book Company: New York, NY, USA, 1987; pp. 179–180. [Google Scholar]
- Cottenie, A.; Verloo, M.; Kiekens, L.; Velghe, G.; Camerlynck, R. Chemical Analysis of Plants and Soils; RUG. Laboratory of Analytical and Agrochemistry: Gent, Belgium, 1982; 63p. [Google Scholar]
- de Carvalho, L.M.J.; Gomes, P.B.; de Oliveira Godoy, R.L.; Pacheco, S.; do Monte, P.H.F.; de Carvalho, J.L.V.; Nutti, M.R.; Neves, A.C.L.; Vieira, A.C.R.A.; Ramos, S.R.R. Total carotenoid content, α-carotene and β-carotene, of landrace pumpkins (Cucurbita moschata Duch): A preliminary study. Food Res. Int. 2012, 47, 337–340. [Google Scholar] [CrossRef] [Green Version]
- A.O.A.C International. Official Methods of Analysis. Association of Official Analytical Chemists. Official Method 985.33. Vitamin C, (Reduced Ascorbic Acid) in Ready-to-Feed Milk Based Infant Formula 2, 6-Dichloroindophenol Titrimetric Method; A.O.A.C International: Washington, DC, USA, 1990; pp. 1108–1109. [Google Scholar]
- Skalindi, F.; Naczk, M. Food phenolics: Sources, chemistry, effects, applications. Trends Food Sci. Technol. 1995, 7, 235–277. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hammerschmidt, R.; Nuckles, E.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Cakmak, I.; Strbac, D.; Marschner, H. Activities of hydrogen peroxide-scavenging enzymes in germinating wheat seeds. J. Exp. Bot. 1993, 44, 127–132. [Google Scholar] [CrossRef]
- Oktay, M.; Küfreviolu, I.; Kocaçalişkan, I.; Şaklrolu, H. Polyphenoloxidase from Amasya apple. J. Food Sci. 1995, 60, 494–496. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Snedecor, G.; Cochran, W. Statistical Methods, 7th ed.; Lowa: Ames, IA, USA, 1982. [Google Scholar]
- Sacks, M.M.; Silk, W.K.; Burman, P. Effect of water stress on cortical cell division rates within the apical meristem of primary roots of maize. Plant Physiol. 1997, 114, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Kan, X.; Chen, J.; Hua, H.; Li, Y.; Ren, J.; Feng, K.; Liu, H.; Deng, D.; Yin, Z. Drought-induced changes in photosynthetic electron transport in maize probed by prompt fluorescence, delayed fluorescence, P700 and cyclic electron flow signals. Environ. Exp. Bot. 2019, 158, 51–62. [Google Scholar] [CrossRef]
- Ma, X.; He, Q.; Zhou, G. Sequence of Changes in Maize Responding to Soil Water Deficit and Related Critical Thresholds. Front. Plant Sci. 2018, 9, 511. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Inter. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Amin, M.; Ahmad, R.; Ali, A.; Hussain, I.; Mahmood, R.; Aslam, M.; Lee, D.J. Influence of silicon fertilization on maize performance under limited water supply. Silicon 2018, 10, 177–183. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2019, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Szepesi, Á.; Szőllősi, R. Mechanism of proline biosynthesis and role of proline metabolism enzymes under environmental stress in plants. In Plant Metabolites and Regulation Under Environmental Stress; Elsevier: Amsterdam, The Netherlands, 2018; pp. 337–353. [Google Scholar]
- Das, A.; Rushton, P.J.; Rohila, J.S. Metabolomic profiling of soybeans (Glycine max L.) reveals the importance of sugar and nitrogen metabolism under drought and heat stress. Plants 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-W.; Lee, S.-K.; Jeong, H.-J.; An, G.; Jeon, J.-S.; Jung, K.-H. Crosstalk between diurnal rhythm and water stress reveals an altered primary carbon flux into soluble sugars in drought-treated rice leaves. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hajiboland, R.; Cheraghvareh, L.; Poschenrieder, C. Improvement of drought tolerance in tobacco (Nicotiana rustica L.) plants by silicon. J. Plant Nutr. 2017, 40, 1661–1676. [Google Scholar] [CrossRef]
- Zahoor, R.; Zhao, W.; Abid, M.; Dong, H.; Zhou, Z. Potassium application regulates nitrogen metabolism and osmotic adjustment in cotton (Gossypium hirsutum L.) functional leaf under drought stress. J. Plant Physiol. 2017, 215, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Arve, L.; Torre, S.; Olsen, J.; Tanino, K. Stomatal responses to drought stress and air humidity. In Abiotic Stress in Plants-Mechanisms and Adaptations; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Bista, D.R.; Heckathorn, S.A.; Jayawardena, D.M.; Mishra, S.; Boldt, J.K. Effects of drought on nutrient uptake and the levels of nutrient-uptake proteins in roots of drought-sensitive and-tolerant grasses. Plants 2018, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloem, J.; de Ruiter, P.C.; Koopman, G.J.; Lebbink, G.; Brussaard, L. Microbial numbers and activity in dried and rewetted arable soil under integrated and conventional management. Soil Biol. Biochem. 1992, 24, 655–665. [Google Scholar] [CrossRef]
- Ge, T.-D.; Sun, N.-B.; Bai, L.-P.; Tong, C.-L.; Sui, F.-G. Effects of drought stress on phosphorus and potassium uptake dynamics in summer maize (zea mays) throughout the growth cycle. Acta Physiol. Plant. 2012, 34, 2179–2186. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, X.; Wang, Y.; Zhang, Y.; Qi, W. Responses to Potassium Application and Economic Optimum K Rate of Maize under Different Soil Indigenous K Supply. Sustainability 2018, 10, 2267. [Google Scholar] [CrossRef] [Green Version]
- Naeem, M.; Naeem, M.S.; Ahmad, R.; Ihsan, M.Z.; Ashraf, M.Y.; Hussain, Y.; Fahad, S. Foliar calcium spray confers drought stress tolerance in maize via modulation of plant growth, water relations, proline content and hydrogen peroxide activity. Arch. Agron. Soil Sci. 2018, 64, 116–131. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Lucena, J.J.; Hernández-Apaolaza, L. Effect of silicon addition on soybean (Glycine max) and cucumber (Cucumis sativus) plants grown under iron deficiency. Plant Physiol. Biochem. 2013, 70, 455–461. [Google Scholar] [CrossRef]
- Rinny, S.; Rout, G. Effect of silicon interaction with nutrients in rice. J. Exp. Biol. Agric. Sci. 2018, 6, 717–731. [Google Scholar]
- Carrasco-Gil, S.; Rodríguez-Menéndez, S.; Fernández, B.; Pereiro, R.; de la Fuente, V.; Hernandez-Apaolaza, L. Silicon induced Fe deficiency affects Fe, Mn, Cu and Zn distribution in rice (Oryza sativa L.) growth in calcareous conditions. Plant Physiol. Biochem. 2018, 125, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Apaolaza, L. Can silicon partially alleviate micronutrient deficiency in plants? A review. Planta 2014, 240, 447–458. [Google Scholar] [CrossRef]
- Rellán-Álvarez, R.; Giner-Martínez-Sierra, J.; Orduna, J.; Orera, I.; Rodríguez-Castrillón, J.Á.; García-Alonso, J.I.; Abadía, J.; Álvarez-Fernández, A. Identification of a tri-iron (III), tri-citrate complex in the xylem sap of iron-deficient tomato resupplied with iron: New insights into plant iron long-distance transport. Plant Cell Physiol. 2010, 51, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, H.-H.; Qiu, H.; Tian, T.; Zhan, S.-S.; Chaney, R.L.; Wang, S.-Z.; Tang, Y.-T.; Morel, J.-L.; Qiu, R.-L. Mitigation effects of silicon rich amendments on heavy metal accumulation in rice (Oryza sativa L.) planted on multi-metal contaminated acidic soil. Chemosphere 2011, 83, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- da Cunha, K.P.V.; do Nascimento, C.W.A. Silicon effects on metal tolerance and structural changes in maize (zea mays L.) grown on a cadmium and zinc enriched soil. Water Air Soil Pollut. 2009, 197, 323. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Panigrahi, R.; Mohapatra, P.K.; Ito, J.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity. Plant Sci. 2011, 180, 368–374. [Google Scholar] [CrossRef]
- Shen, X.; Zhou, Y.; Duan, L.; Li, Z.; Eneji, A.E.; Li, J. Silicon effects on photosynthesis and antioxidant parameters of soybean seedlings under drought and ultraviolet-B radiation. J. Plant Physiol. 2010, 167, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, R.A.; El Khawas, S.A.; Khafaga, H.S.; Abd El-Nabe, A.S.; Abd Elrady, A.S. Amelioration of Drought Stress on Physiological Performance of Pearl Millet (Pennisetum americanum) Plant Grown Under Saline Condition Using Potassium Humate and Silicon Source. Egypt. J. Exp. Biol. 2017, 13, 57–68. [Google Scholar] [CrossRef]
- de Camargo, M.; Bezerra, B.; Holanda, L.; Oliveira, A.; Vitti, A.; Silva, M. Silicon fertilization improves physiological responses in sugarcane cultivars grown under water deficit. J. Soil Sci. Plant Nutr. 2019, 19, 81–91. [Google Scholar] [CrossRef]
- Zhang, Y. Ascorbic Acid in Plants: Biosynthesis, Regulation and Enhancement; Springer Science & Business Media: New York, NY, USA; Heidelberg, Germany; Dordrecht, The Netherlands; London, UK, 2013; pp. 111–114. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Buet, A.; Grozeff, G.G.; Galatro, A.; Simontacchi, M. Ascorbate-glutathione cycle and abiotic stress tolerance in plants. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2017; pp. 177–200. [Google Scholar] [CrossRef]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef] [PubMed]
- Weidner, S.; Karolak, M.; Karamac, M.; Kosinska, A.; Amarowicz, R. Phenolic compounds and properties of antioxidants in grapevine roots [Vitis vinifera L.] under drought stress followed by recovery. Acta Soc. Bot. Pol. 2009, 78, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Malčovská, S.M.; Dučaiová, Z.; Maslaňáková, I.; Bačkor, M. Effect of silicon on growth, photosynthesis, oxidative status and phenolic compounds of maize (Zea mays L.) grown in cadmium excess. Water Air Soil Pollut. 2014, 225, 2056. [Google Scholar] [CrossRef]
- Shetty, R.; Fretté, X.; Jensen, B.; Shetty, N.P.; Jensen, J.D.; Jørgensen, H.J.L.; Newman, M.-A.; Christensen, L.P. Silicon-induced changes in antifungal phenolic acids, flavonoids, and key phenylpropanoid pathway genes during the interaction between miniature roses and the biotrophic pathogen Podosphaera pannosa. Plant Physiol. 2011, 157, 2194–2205. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Gao, H.; Zhao, B.; Dong, S.; Zhang, J.; Yang, J.; Wang, J.; Liu, P. Effects of drought stress on activity of photosystems in leaves of maize at grain filling stage. Acta Agron. Sin. 2009, 35, 1916–1922. [Google Scholar] [CrossRef]
- Aslam, M.; Zamir, M.; Afzal, I.; Yaseen, M.; Mubeen, M.; Shoaib, A. Drought stress, its effect on maize production and development of drought tolerance through potassium application. Cercetari Agronomice în Moldova 2013, 46, 154. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | pH | EC µS cm−1 | CaCO3% | Cation meq/L | Anion meq/L | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ca++ | Mg++ | Na+ | HCO3− | CL− | SO4−−2 | ||||
| 2018 | 7.84 | 0.41 | 2.87 | 5.52 | 0.38 | 1.03 | 1.59 | 1.20 | 1.74 |
| 2019 | 7.61 | 0.47 | 3.13 | 7.04 | 0.50 | 0.80 | 2.14 | 1.38 | 1.62 |
| N, P, K | N | P | K | Sand% | Silt% | Clay% | Soil texture | ||
| (ppm) | |||||||||
| 2018 | 2.88 | 6.38 | 1.17 | 91.95 | 4.81 | 3.24 | Sandy | ||
| 2019 | 2.03 | 6.22 | 0.91 | ||||||
| Date | Solar Radiation Dgt [MJ/m2] | Precipitation [mm] | Wind Speed [m/s] | Air Temperature [°C] | Relative Humidity [%] | |||
|---|---|---|---|---|---|---|---|---|
| Average | Sum | Average | Max | Average | Min | Max | Average | |
| 2018 | ||||||||
| May | 671.29 | 0.0 | 1.4 | 8.9 | 23.9 | 11.8 | 38.5 | 62.5 |
| June | 654.76 | 0.0 | 1.3 | 5.6 | 26.6 | 13.4 | 38.2 | 67.4 |
| July | 616.47 | 0.0 | 0.9 | 4.8 | 27.7 | 16.9 | 37.4 | 75.5 |
| August | 542.00 | 0.0 | 0.5 | 3.7 | 27.5 | 17.4 | 38.0 | 75.0 |
| September | 424.92 | 0.0 | 0.7 | 3.8 | 25.2 | 14.7 | 36.2 | 73.5 |
| 2019 | ||||||||
| May | 689.12 | 0.0 | 1.3 | 6.5 | 23.9 | 12.7 | 36.1 | 58.1 |
| June | 535.47 | 0.0 | 1.3 | 5.2 | 27.6 | 15.7 | 39.6 | 65.6 |
| July | 472.97 | 0.0 | 1.1 | 5.0 | 27.5 | 17.8 | 37.2 | 71.0 |
| August | 415.72 | 0.0 | 1.0 | 4.6 | 27.3 | 17.0 | 37.5 | 73.4 |
| September | 327.05 | 0.0 | 1.0 | 5.3 | 25.8 | 14.6 | 41.2 | 69.8 |
| Days | Date | Stage | KC* | Irrigation Level (m3·ha−1) | ||
|---|---|---|---|---|---|---|
| 100% | 75% | 50% | ||||
| 10 days | 17/5:26/5 | initial | 0.3 | 299.52 | 299.52 | 299.52 |
| 10 days | 27/5:5/6 | 0.6 | 694.08 | 694.08 | 694.08 | |
| Starting date of different irrigation regimes | ||||||
| 15 days | 6/6:20/6 | development | 0.9 | 953.28 | 714.96 | 476.64 |
| 20 days | 21/6:10/7 | 1.0 | 1114.56 | 835.92 | 557.28 | |
| 20 days | 11/7:30/7 | Mid-season | 1.2 | 1319.04 | 989.28 | 659.52 |
| 20 days | 31/7:19/8 | 1.0 | 1085.76 | 814.32 | 542.88 | |
| 10 days | 20/8:29/8 | Last season | 0.9 | 1097.28 | 822.96 | 548.64 |
| 11 days | 30/8:11/9 | Not irrigated before harvest | ||||
| Total amount (m3 ha−1) | ||||||
| 116 Days | 6563.52 | 5171.04 | 3778.56 | |||
| Variables | Irrigation Level | Foliar Application | ||||
|---|---|---|---|---|---|---|
| 100% | 75% | 50% | KSi 0 | KSi 1 | KSi 2 | |
| Shoot dry weight (g.plant−1) | 318.4 A | 242.1 B | 208.1 C | 232.6 C | 260.3 B | 275.7 A |
| Root dry weight (g.plant−1) | 45.32 A | 38.20 B | 33.01 C | 36.52 B | 39.43 A | 40.58 A |
| Root/shoot ratio | 0.144 B | 0.158 A | 0.159 A | 0.157 A | 0.153 A | 0.150 A |
| LAI | 7.15 A | 5.19 B | 4.08 C | 4.62 C | 5.44 B | 6.35 A |
| RWC (%) | 88.11 A | 73.13 B | 67.79 C | 75.52 B | 77.17 A | 76.34 AB |
| MDA (nmol.g−1 FW) | 5.82 C | 11.99 B | 13.62A | 11.08 A | 10.27 B | 10.09 B |
| Proline (µg.g−1 FW) | 183.3 C | 314.0 A | 241.6 B | 285.0 A | 241.5 B | 212.2C |
| Soluble sugars (mg.g−1 DW) | 24.50 B | 45.54 A | 44.53 A | 35.13 C | 37.99 B | 41.44 A |
| N (mg.g−1 DW) | 85.29 A | 71.00 B | 62.14 C | 70.39 B | 71.31 B | 76.72 A |
| P (mg.g−1 DW) | 2.24 A | 1.91 B | 1.50 C | 1.77 B | 1.93 A | 1.96 A |
| K (mg.g−1 DW) | 11.74 A | 9.90 B | 8.81 C | 8.02 C | 10.65 B | 11.79 A |
| Ca (mg.g−1 DW) | 7.25 A | 6.52 B | 5.22 C | 6.07 B | 6.31 AB | 6.61 A |
| Fe (µg.g−1 DW) | 203.9A | 180.5 B | 161.7 C | 169.7 C | 180.7 B | 195.6 A |
| Zn (µg.g−1 DW) | 46.3A | 47.0 A | 41.0 B | 48.6 A | 42.9 B | 42.8 B |
| Carotenoids (mg.g−1 FW) | 0.332 A | 0.308 B | 0.288 C | 0.277 C | 0.317 B | 0.334 A |
| Ascorbic acid (µmol.g−1 FW) | 1.36 C | 1.78 A | 1.67 B | 1.57 B | 1.61 AB | 1.64 A |
| Soluble phenols (µg.g−1 FW) | 13.97 C | 16.39 B | 17.29 A | 15.27 C | 15.87 B | 16.49A |
| POD (Δ O.D. min−1.mg protein) | 14.3 C | 33.6 B | 35.6 A | 25.92 B | 28.90 A | 28.67 A |
| CAT (Δ O.D. min−1.mg protein) | 2.64 C | 4.17 A | 3.57 B | 3.19 B | 3.58 A | 3.60 A |
| PPO (Δ O.D. min−1.mg protein) | 6.95 C | 8.59 B | 9.69 A | 7.97 B | 8.57 A | 8.67 A |
| APX (Δ O.D. min−1.mg protein) | 2.83 C | 4.36 A | 4.01 B | 3.40 B | 3.85 A | 3.94 A |
| Ear length (cm) | 23.3 A | 19.1 B | 15.7 C | 18.2 C | 19.4 B | 20.5 A |
| Ear diameter (cm) | 4.7 A | 4.4 B | 3.3 C | 4.00 C | 4.17 B | 4.30 A |
| Number of grains/ear | 332.4 A | 271.8 B | 226.3 C | 264.7 B | 281.2 A | 284.6 A |
| Weight of grains/ear (g) | 109.7 A | 84.5 B | 66.2 C | 79.5 C | 88.8 B | 92.2 A |
| Weight of grains/plant (g) | 172.8 A | 127.6 B | 95.0 C | 118.1 C | 135.0 B | 142.4 A |
| Grain yield (ton·ha−1) | 7.94 A | 5.82 B | 4.39 C | 5.33 C | 6.14 B | 6.68 A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, M.F.M.; Abd El-Samad, G.; Ashour, H.; El-Sawy, A.M.; Hikal, M.; Elkelish, A.; El-Gawad, H.A.; El-Yazied, A.A.; Hozzein, W.N.; Farag, R. Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions. Agronomy 2020, 10, 1212. https://doi.org/10.3390/agronomy10081212
Ibrahim MFM, Abd El-Samad G, Ashour H, El-Sawy AM, Hikal M, Elkelish A, El-Gawad HA, El-Yazied AA, Hozzein WN, Farag R. Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions. Agronomy. 2020; 10(8):1212. https://doi.org/10.3390/agronomy10081212
Chicago/Turabian StyleIbrahim, Mohamed F. M., Gomaa Abd El-Samad, Hatem Ashour, Ahmed M. El-Sawy, Mohamed Hikal, Amr Elkelish, Hany Abd El-Gawad, Ahmed Abou El-Yazied, Wael N. Hozzein, and Reham Farag. 2020. "Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions" Agronomy 10, no. 8: 1212. https://doi.org/10.3390/agronomy10081212
APA StyleIbrahim, M. F. M., Abd El-Samad, G., Ashour, H., El-Sawy, A. M., Hikal, M., Elkelish, A., El-Gawad, H. A., El-Yazied, A. A., Hozzein, W. N., & Farag, R. (2020). Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions. Agronomy, 10(8), 1212. https://doi.org/10.3390/agronomy10081212