Enhancing Drought Tolerance and Striga hermonthica Resistance in Maize Using Newly Derived Inbred Lines from the Wild Maize Relative, Zea diploperennis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Materials Used for the Study
2.2. Field Evaluations
2.3. Data Collection
2.4. Data Analysis
3. Results
3.1. Analysis of Variance across Multiple Environments
3.2. Performance of Hybrids Based on Multiple Trait Index across Environments
3.3. Correlation between Variables under Each Stress Environment
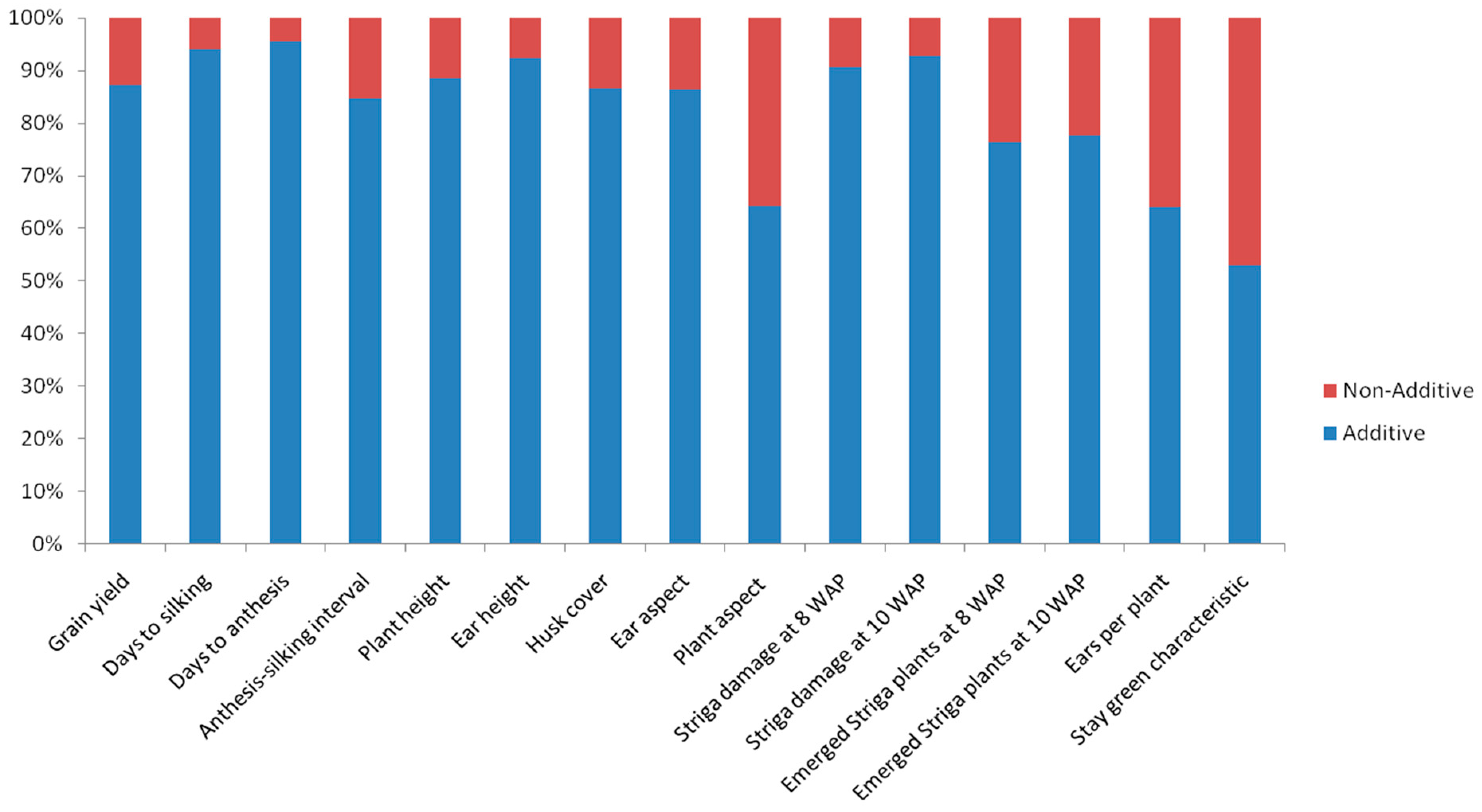
3.4. General and Specific Combining Abilities of Inbred Lines across Multiple Environments
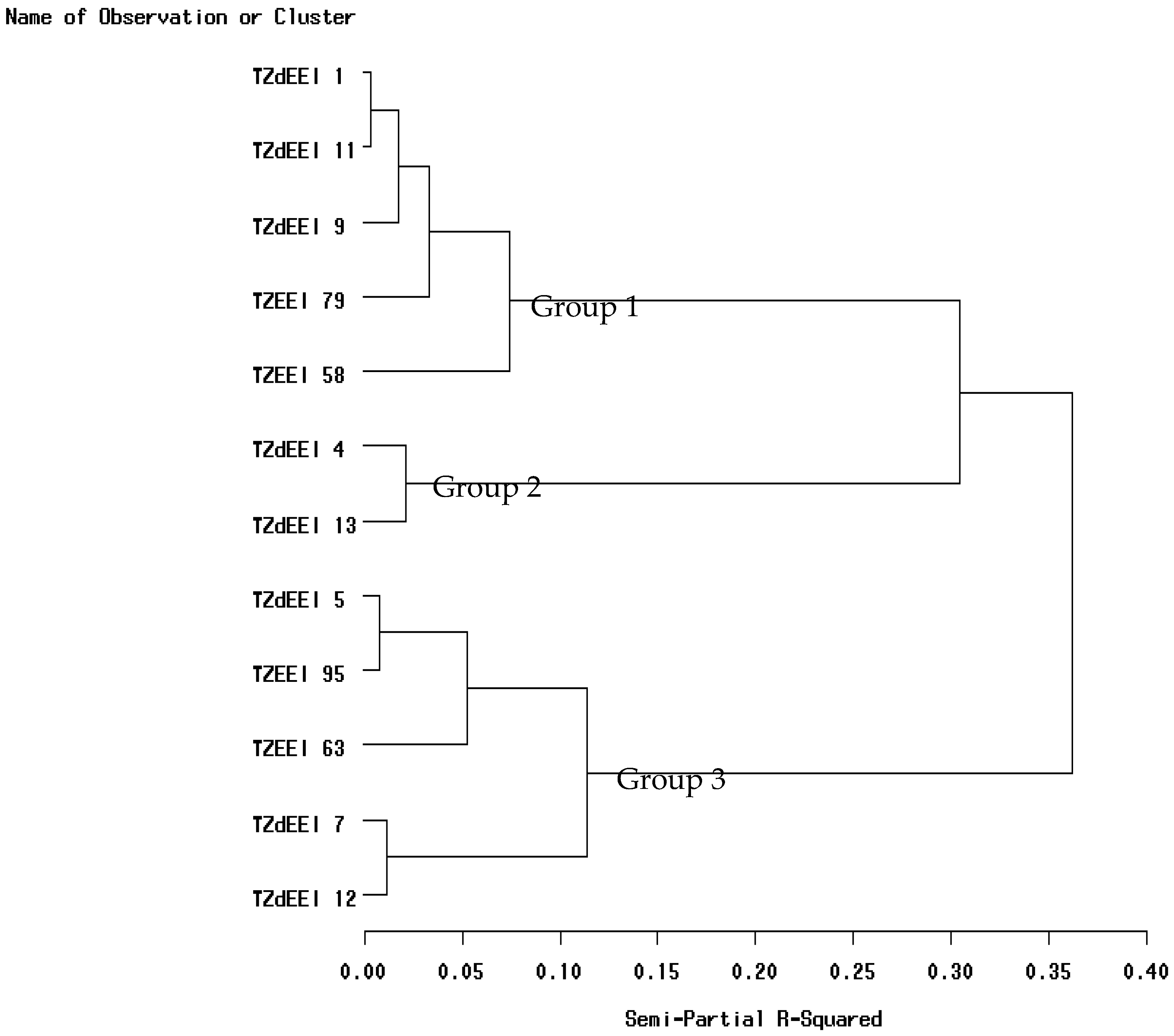
3.5. Classification of Inbred Lines into Heterotic Groups and Identification of Testers
3.6. Performance of Hybrids Based on GGE Biplot Analyses across Environments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byerlee, D.; Eicher, C. Africa’s Emerging Maize Revolution; Lynee Rienner Publishers: Boulder, CO, USA, 1971. [Google Scholar]
- Badu-Apraku, B.; Ewool, M.; Yallou, C.G. Registration of Striga-resistant tropical extra-early maize population. J. Plant Regist. 2010, 4, 60–66. [Google Scholar] [CrossRef]
- Kroschel, J. Analysis of the Striga problem: The first step towards future joint action. In Advances in Parasitic Weed Control at on Farm Level: Joint Action to Control Striga in Africa; Kroschel, J., Mercher-Quarshie, H., Sauerborn, J., Eds.; Margraf Publishers: Weikersheim, Germany, 1999; Volume 1, pp. 3–26. [Google Scholar]
- Haussmann, B.I.; Hess, D.E.; Welz, H.-G.; Geiger, H.H. Improved methodologies for breeding striga-resistant sorghums. Field Crop. Res. 2000, 66, 195–211. [Google Scholar] [CrossRef] [Green Version]
- Rodenburg, J.; Diagne, A.; Oikeh, S.; Futakuchi, K.; Kormawa, P.M.; Semon, M.; Akintayo, I.; Cissé, B.; Sié, M.; Narteh, L.; et al. Achievements and impact of NERICA on sustainable rice production in sub-Saharan Africa. Int. Rice Comm. Newsl. 2006, 55, 45–58. [Google Scholar]
- Badu-Apraku, B.; Akinwale, R. Cultivar evaluation and trait analysis of tropical early maturing maize under Striga-infested and Striga-free environments. Field Crop. Res. 2011, 121, 186–194. [Google Scholar] [CrossRef]
- DeVries, J.; Haussmann, B.I.G.; Hess, D.E.; Koyama, M.L.; Kling, J.; Melake-Berhan, A.; Geiger, A.A. The Inheritance of Striga Reactions in Maize. In Proceedings of the Workshop on Breeding for Striga Resistance in Cereals, IITA, Ibadan, Nigeria, 18–20 August 1999; Margraf Verlag: Weikersheim, Germany, 2000; pp. 73–84. [Google Scholar]
- Ejeta, G.; Butler, L.G.; Babiker, A.G. New approaches to the control of Striga. In Striga Research at Purdue University; Research Bulletin RB-991; Agricultural Experiment Station, Purdue University: West Lafayette, IN, USA, 1991. [Google Scholar]
- Rodenburg, J.; Bastiaans, L.; Weltzien, E.; Hess, D. How can field selection for Striga resistance and tolerance in sorghum be improved? Field Crop. Res. 2005, 93, 34–50. [Google Scholar] [CrossRef] [Green Version]
- NeSmith, D.S.; Ritchie, J.T. Short- and long-term responses of corn to a pre-anthesis soil water deficit. Agron. J. 1992, 84, 107–113. [Google Scholar] [CrossRef]
- Ribeiro, P.F.; Rodriguez, A.V.C. Emerging Advanced Technologies to Mitigate the Impact of Climate Change in Africa. Plants 2020, 9, 381. [Google Scholar] [CrossRef] [Green Version]
- Edmeades, G.O.; Bolaños, J.; Chapman, S.; Lafitte, H.; Bänziger, M. Selection improves drought tolerance in tropical maize populations: I. Gains in Biomass, grain yield, and harvest index. Crop Sci. 1999, 39, 1306–1315. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Oyekunle, M. Selection of extra-early maize inbreds under low N and drought at flowering and grain-filling for hybrid production. Maydica 2011, 56, 36–47. [Google Scholar]
- Edmeades, G.O.; Bänziger, M.; Schussler, J.R.; Campos, H. Improving Abiotic Stress Tolerance in Maize: A Random or Planned Process? In Proceedings of the Arnel R. Hallauer International Symposium on Plant Breeding, Mexico City, Mexico, 17–22 August 2003; Iowa State University Press: Iowa Ames, IA, USA, 2003. [Google Scholar]
- Akaogu, I.C.; Badu-Apraku, B.; Adetimirin, V.O. Combining ability and performance of extra-early maturing yellow maize inbreds in hybrid combinations under drought and rain-fed conditions. J. Agric. Sci. 2017, 155, 1520–1540. [Google Scholar] [CrossRef] [Green Version]
- Mammadov, J.; Buyyarapu, R.; Guttikonda, S.K.; Parliament, K.; Abdurakhmonov, I.Y.; Kumpatla, S.P. Wild relatives of maize, rice, cotton, and soybean: Treasure troves for tolerance to biotic and abiotic stresses. Front. Plant Sci. 2018, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Warburton, M.L.; Rauf, S.; Marek, L.; Hussain, M.; Ogunola, O.; Gonzalez, J.D.J.S. The Use of Crop Wild Relatives in Maize and Sunflower Breeding. Crop Sci. 2017, 57, 1227–1240. [Google Scholar] [CrossRef]
- Menkir, A.; Kling, J.; Badu-Apraku, B.; Ibikunle, O. Registration of 26 Tropical Maize Germplasm Lines with Resistance to Striga hermonthica. Crop Sci. 2006, 46, 1007–1009. [Google Scholar] [CrossRef]
- Yallou, C.G.; Menkir, A.; Adetimirin, V.O.; Kling, J.G. Combining ability of maize inbred lines containing genes from Zea diploperennis for resistance to Striga hermonthica (Del.) Benth. Plant Breed. 2009, 128, 143–148. [Google Scholar] [CrossRef]
- Amegbor, I.K.; Badu-Apraku, B.; Adu, G.; Adjebeng-Danquah, J.; Toyinbo, J. Combining Ability of Extra-Early Maize Inbreds Derived from a Cross between Maize and Zea diploperennis and Hybrid Performance under Contrasting Environments. Agronomy 2020, 10, 1069. [Google Scholar] [CrossRef]
- Lennon, J.R.; Krakowsky, M.; Goodman, M.; Flint-Garcia, S.; Balint-Kurti, P.J. Identification of Alleles Conferring Resistance to Gray Leaf Spot in Maize Derived from its Wild Progenitor Species Teosinte. Crop Sci. 2016, 56, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Meseka, S.; Fakorede, M.; Ajala, S.; Badu-Apraku, B.; Menkir, A. Introgression of Alleles from Maize Landraces to Improve Drought Tolerance in an Adapted Germplasm. J. Crop Improv. 2013, 27, 96–112. [Google Scholar] [CrossRef]
- Liu, Z.; Garcia, A.; McMullen, M.D.; Flint-Garcia, S.A. Genetic analysis of kernel traits in maize-teosinte introgression populations. G3 2016, 6, 2523–2530. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.-L.; Rong, T.; Song, Y.; Yang, J.; Pan, G.; Li, W.; Huang, Y.; Cao, M. Introgression of perennial teosinte genome into maize and identification of genomic in situ hybridization and microsatellite markers. Crop Sci. 2005, 45, 717–721. [Google Scholar] [CrossRef]
- Ayinde, T.B.; Fola, A.J.; Ibrahim, U. Economic Advantage of Hybrid Maize over Open-Pollinated Maize in Giwa Local Government Area of Kaduna State. Am. J. Exp. Agric. 2011, 1, 101–109. [Google Scholar] [CrossRef]
- Sprague, G.F.; Tatum, L.A. General versus specific combining ability in single crosses of corn. J. Am. Soc. Agron. 1942, 34, 923–932. [Google Scholar] [CrossRef]
- Abrha, S.W.; Habtamu, Z.Z.; Dagne, W.G. Line × tester analysis of maize inbred lines for grain yield and yield related traits. Asian J. Plant Sci. Res. 2013, 3, 12–19. [Google Scholar]
- Girma, C.H.; Sentayehu, A.; Berhanu, T.; Temesgen, M. Test cross performance and combining ability of maize (Zea mays L.) inbred lines at Bako, Western Ethiopia. GJSFR D Agric. Vet. 2015, 15, 2249–4626. [Google Scholar]
- Badu-Apraku, B.; Oyekunle, M.; Fakorede, M.A.B.; Vroh, I.; Akinwale, R.O.; Aderounmu, M. Combining ability, heterotic patterns and genetic diversity of extra-early yellow inbreds under contrasting environments. Euphytica 2013, 192, 413–433. [Google Scholar] [CrossRef]
- Griffing, B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J.; Miranda, J.B. Quantitative Genetics in Maize Breeding; Springer: New York, NY, USA, 1988. [Google Scholar]
- Badu-Apraku, B.; Fakorede, M.A.B.; Menkir, A.; Sanogo, D. Conduct and Management of Maize Field Trials; IITA: Ibadan, Nigeria, 2012; p. 59. [Google Scholar]
- Kim, S.K. Breeding maize for Striga tolerance and development of a field infestation technique. In Combating Striga in Africa, Proceedings of the International Workshop organized by IITA, ICRISAT, and IDRC, Ibadan, Nigeria, 22–24 August 1988; IITA: Ibadan, Nigeria, 1991. [Google Scholar]
- Kim, S.K.; Winslow, M.D. Progress in Breeding Maize for Striga Tolerance/Resistance at IITA. In Proceedings of the Fifth International Symposium on Parasitic Weeds, Nairobi, Kenya, 24–30 June 1991; Ransom, J.K., Ed.; CIMMYT: Nairobi, Kenya, 1991; pp. 494–499. [Google Scholar]
- Badu-Apraku, B.; Akinwale, R.; Franco, J.; Oyekunle, M. Assessment of reliability of secondary traits in selecting for improved grain yield in drought and low-nitrogen environments. Crop Sci. 2012, 52, 2050–2062. [Google Scholar] [CrossRef]
- SAS Institute. Statistical Analysis Software (SAS); Users’ Guide; SAS Inst.: Cary, NC, USA, 2011. [Google Scholar]
- Zhang, Y.; Kang, M.S.; Lamkey, K.R. DIALLEL-SAS05: A comprehensive program for Griffing’s and Gardner-Eberhart analyses. Agron. J. 2005, 97, 1097–1106. [Google Scholar] [CrossRef]
- Baker, R.J. Issues in diallel analysis. Crop Sci. 1978, 18, 533–536. [Google Scholar] [CrossRef]
- Hung, H.-Y.; Holland, J.B. Diallel analysis of resistance to fusarium ear rot and fumonisin contamination in maize. Crop Sci. 2012, 52, 2173–2181. [Google Scholar] [CrossRef] [Green Version]
- Pswarayi, A.; Vivek, B.S. Combining ability amongst CIMMYT’s early maturing maize (Zea mays L.) germplasm under stress and non-stress conditions and identification of testers. Euphytica 2008, 162, 353–362. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Annor, B.; Oyekunle, M.; Akinwale, R.; Fakorede, M.; Talabi, A.; Akaogu, I.; Melaku, G.; Fasanmade, Y. Grouping of early maturing quality protein maize inbreds based on SNP markers and combining ability under multiple environments. Field Crop. Res. 2015, 183, 169–183. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Hunt, L.; Sheng, Q.; Szlavnics, Z. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Yan, W. GGE Biplot-A windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Badu-Apraku, B.; Oyekunle, M.; Akinwale, R.O.; Fontem, L.A. Combining ability of early-maturing white maize inbreds under stress and nonstress environments. Agron. J. 2011, 103, 544–557. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Oyekunle, M.; Yallou, G.C.; Obeng-Antwi, K.; Haruna, A.; Usman, I.S.; Akinwale, R. Gains in grain yield of early maize cultivars developed during three breeding eras under multiple environments. Crop Sci. 2014, 55, 527–539. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Oyekunle, M. Genetic analysis of grain yield and other traits of extra-early yellow maize inbreds and hybrid performance under contrasting environments. Field Crop. Res. 2012, 129, 99–110. [Google Scholar] [CrossRef]
- Gethi, J.G.; Smith, M.E. Genetic responses of single crosses of Maize to Striga hermonthica (Del.) Benth. and Striga asiatica (L.) kuntze. Crop Sci. 2004, 44, 2068–2077. [Google Scholar] [CrossRef]
- Bolanos, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crop. Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
- Stewart, G.R.; Press, M.C.; Graves, J.D.; Nour, J.J.; Wylde, A. A physiological characterization of the host-parasite association between Sorghum bicolor and Striga hermonthica and its implication for Striga control. In Combating Striga in Africa, Proceedings of the International Workshop Organised by IITA, ICRISAT and IDRC, Ibadan, Nigeria, 22–24 August 1988; Kim, S.K., Ed.; IITA: Ibadan, Nigeria, 1991; pp. 48–54. [Google Scholar]
- Adetimirin, V.O.; Aken’Ova, M.E.; Kim, S.K. Effects of Striga hermonthica on yield components in maize. J. Agric. Sci. 2000, 135, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Adetimirin, V.O.; Thé, C.; Dossou, R. Yield losses in maize due to Striga hermonthica in West and Central Africa. Int. J. Pest Manag. 2002, 48, 211–217. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Menkir, A.; Kamara, A.Y.; Adam, A. Effects of drought screening methodology on genetic variances and covariances in Pool 16 DT maize population. J. Agric. Sci. 2004, 142, 445–452. [Google Scholar] [CrossRef]
- Akaogu, I.C.; Badu-Apraku, B.; Adetimirin, V.O.; Vroh-Bi, I.; Oyekunle, M.; Akinwale, R.O. Genetic diversity assessment of extra-early maturing yellow maize inbreds and hybrid performance inStriga-infested andStriga-free environments. J. Agric. Sci. 2012, 151, 519–537. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Gedil, M.; Talabi, A.O.; Annor, B.; Oyekunle, M.; Akinwale, R.O.; Fasanmade, T.Y.; Akaogu, I.C.; Aderounmu, M. Heterotic responses among crosses of IITA and CIMMYT early white maize inbred lines under multiple stress environments. Euphytica 2015, 206, 245–262. [Google Scholar] [CrossRef]
- Njeri, S.G.; Makumbi, D.; Warburton, M.L.; Diallo, A.; Jumbo, M.B.; Chemining’Wa, G. Genetic analysis of tropical quality protein maize (Zea mays L.) germplasm. Euphytica 2017, 213, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umar, U.U.; Ado, S.G.; Aba, D.A.; Bugaje, S.M. Estimates of combining ability and gene action in maize (Zea mays L.) under water stress and non-stress conditions. J. Biol. Agric. Healthc. 2014, 4, 25. [Google Scholar]
- Makumbi, D.; Betrán, J.F.; Bänziger, M.; Ribaut, J.-M. Combining ability, heterosis and genetic diversity in tropical maize (Zea mays L.) under stress and non-stress conditions. Euphytica 2011, 180, 143–162. [Google Scholar] [CrossRef]
- Oyetunde, O.A.; Badu-Apraku, B.; Ariyo, O.J.; Alake, C.O. Efficiencies of heterotic grouping methods for classifying early maturing maize inbred lines. Agronomy 2020, 10, 1198. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Optimal | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOV † | DF | YIELD | DS | ASI | PHT | EHT | PASP | EASP | EPP | ||||||
| Env | 1 | 1,506,994 ** | 6.8 ** | 1.6 ** | 776.9 ** | 332.8 ** | 0.38 ** | 0.31 ** | 0.02 ** | ||||||
| Repetition | 2 | 598,147 | 0.7 | 0.5 | 11.9 | 274.6 ** | 0.22 | 3.23 ** | 0.02 | ||||||
| Hybrid | 65 | 865,461 * | 2.2 ** | 0.9 * | 396.0 | 68.8 | 0.26 * | 0.24 * | 0.03 ** | ||||||
| Env*Hybrid | 65 | 841,744 * | 2.2 ** | 0.8 | 385.6 | 68.9 | 0.27 * | 0.23 ** | 0.03 ** | ||||||
| GCA | 11 | 2,998,626 ** | 28.5 ** | 3.9 ** | 2651.7 ** | 1450.4 ** | 1.03 ** | 0.46 ** | 0.04 ** | ||||||
| SCA | 54 | 1,203,143 ** | 2.5 ** | 1.2 ** | 395.1 | 105.1 ** | 0.26 * | 0.28 ** | 0.02 * | ||||||
| GCA*Env | 11 | 1,241,955 * | 4.4 ** | 2.1 ** | 634.7 * | 114.4 * | 0.72 ** | 0.42 ** | 0.07 ** | ||||||
| SCA*Env | 54 | 788,767 * | 1.7 * | 0.7 | 347.4 | 59.6 | 0.17 | 0.20 | 0.02 * | ||||||
| Error | 130 | 545,704 | 1.0 | 0.7 | 324.9 | 55..3 | 0.17 | 0.15 | 0.01 | ||||||
| Striga | |||||||||||||||
| SOV † | DF | YIELD | DS | ASI | PHT | EHT | SDR1 | SDR2 | ESP1 | ESP2 | EPP | ||||
| Env | 1 | 39,455,484 ** | 570.2 ** | 78.6 ** | 25,409.5 ** | 6264.4 ** | 0.09 | 1.83 | 3.63 ** | 4.23 ** | 0.62 ** | ||||
| Repetition | 2 | 784,070 | 6.4 | 0.1 | 1219.7 ** | 534.6 * | 2.43 | 0.52 | 0.43 ** | 0.41 ** | 0.05 | ||||
| Hybrid | 65 | 3,176,326 ** | 12.5 ** | 3.6 ** | 452.8 ** | 203.6 ** | 4.49 ** | 3.87 ** | 0.12 * | 0.09 * | 0.11 ** | ||||
| Env*Hybrid | 65 | 1 094,354 * | 5.6 ** | 2.6 * | 304.1 | 136.2 | 1.56 * | 1.06 ** | 0.08 | 0.06 | 0.03 * | ||||
| GCA | 11 | 13,626,819 ** | 52.6 ** | 9.2 ** | 1000.5 ** | 547.3 ** | 17.58 ** | 16.55 ** | 0.28 ** | 0.21 ** | 0.45 ** | ||||
| SCA | 54 | 1,047,522 * | 4.4 | 2.4 | 341.2 * | 133.6 | 1.82 * | 1.29 ** | 0.09 | 0.06 | 0.04 ** | ||||
| GCA*ENV | 11 | 1,345,046 | 8.5* | 3.1 | 433.3 * | 182.3 | 3.01 ** | 2.58 ** | 0.09 | 0.09 | 0.04 | ||||
| SCA*ENV | 54 | 1,043,287 | 5.4* | 2.5 | 277.8 | 126.8 | 1.26 | 0.75 | 0.07 | 0.05 | 0.03 | ||||
| Error | 130 | 726,465 | 3.2 | 1.8 | 227.0 | 113.6 | 1.10 | 0.63 | 0.08 | 0.06 | 0.02 | ||||
| Drought | |||||||||||||||
| SOV † | DF | YIELD | DS | DA | ASI | PHT | EHT | EASP | EPP | PASP | STGR | ||||
| Repetition | 1 | 19,449 | 3.1 | 2.0 | 0.1 | 737.0 | 185.9 | 0.03 | 0.00 | 1.14 | 1.32 | ||||
| Hybrid | 65 | 874,713 ** | 6.2 ** | 3.1 ** | 1.5 | 382.3 ** | 176.4 | 0.71 | 0.02 ** | 1.15 * | 1.38 * | ||||
| GCA | 11 | 2,165,504 ** | 9.0 ** | 17.4 ** | 2.7 * | 1004.4 ** | 535.3 ** | 1.63 ** | 0.02 * | 1.48 * | 1.51 * | ||||
| SCA | 54 | 611,774 | 1.9 * | 3.9 | 1.3 | 255.6 | 103.3 | 0.66 | 0.02 ** | 1.09 | 1.35 * | ||||
| Error | 65 | 424,342 | 1.3 | 3.0 | 1.1 | 303.2 | 118.6 | 0.58 | 0.01 | 0.72 | 0.77 | ||||
| Across | |||||||||||||||
| SOV † | DF | YIELD | DS | DA | ASI | PHT | EHT | HUSK | EASP | EPP | |||||
| ENV | 5 | 219,814,222 ** | 832.5 ** | 965.3 ** | 134.0 ** | 30,657.3 ** | 6062.6 ** | 73.50 ** | 266.12 ** | 1.57 ** | |||||
| Repetition | 6 | 618,319 | 13.1 ** | 8.6 ** | 1.0 | 1243.6 ** | 316.0 ** | 0.89 ** | 1.25 ** | 0.09 | |||||
| Hybrid | 65 | 42,095,210 ** | 26.8 ** | 22.2 ** | 4.0 ** | 1197.8 ** | 552.1 ** | 0.89 ** | 1.67 ** | 0.15 | |||||
| Hybrid*ENV | 325 | 1,242,600 ** | 4.5 ** | 3.3 ** | 1.5 ** | 333.9 | 106.7 | 0.48 ** | 0.59 ** | 0.12 | |||||
| GCA | 11 | 14,196,154 ** | 122.7 ** | 109.0 ** | 13.3 ** | 4741.3 ** | 2458.1 ** | 3.14 ** | 5.37 ** | 0.22 * | |||||
| SCA | 54 | 2,175,206 ** | 7.3 ** | 4.5 ** | 2.1 ** | 475.9 ** | 163.8 ** | 0.44 ** | 0.91 ** | 0.14 | |||||
| GCA*ENV | 55 | 2,590,061 ** | 9.5 ** | 7.2 ** | 2.0 ** | 434.8 * | 140.6 * | 1.46 ** | 1.62 ** | 0.18 ** | |||||
| SCA*ENV | 270 | 968,122 ** | 3.4 | 2.6 ** | 1.4 | 313.3 | 99.8 | 0.28 * | 0.38 | 0.11 | |||||
| Error | 390 | 754,137 | 2.9 | 1.9 | 1.2 | 300.4 | 97.6 | 0.23 | 0.35 | 0.11 | |||||
| YIELD | DS | DA | ASI | PHT | EHT | EASP | EPP | PASP | SDR1 | SDR2 | ESP1 | ESP2 | STGR | ||
| Heritability | 0.73 | 0.80 | 0.86 | 0.63 | 0.76 | 0.84 | 0.62 | 0.34 | 0.37 | 0.71 | 0.77 | 0.45 | 0.45 | 0.28 |
| PEDIGREE | Optimal | Striga | Drought | Across |
|---|---|---|---|---|
| TZdEEI 1 × TZdEEI 4 | 2664 | 2285 | 1634 | 2394 |
| TZdEEI 1 × TZdEEI 5 | 3864 | 2924 | 2661 | 3179 |
| TZdEEI 1 × TZdEEI 7 | 4888 | 4478 | 2577 | 4302 |
| TZdEEI 1 × TZdEEI 9 | 4659 | 2516 | 2834 | 3476 |
| TZdEEI 1 × TZdEEI 11 | 2698 | 1359 | 2614 | 2196 |
| TZdEEI 1 × TZdEEI 12 | 4866 | 3796 | 2262 | 3824 |
| TZdEEI 1 × TZdEEI 13 | 3419 | 1728 | 2026 | 2650 |
| TZdEEI 1 × TZEEI 58 | 4434 | 2958 | 3480 | 3468 |
| TZdEEI 1 × TZEEI 63 | 4241 | 2945 | 3046 | 3632 |
| TZdEEI 1 × TZEEI 79 | 4428 | 3250 | 2900 | 3433 |
| TZdEEI 1 × TZEEI 95 | 3909 | 2730 | 3014 | 3101 |
| TZdEEI 4 × TZdEEI 5 | 3698 | 2405 | 1504 | 2544 |
| TZdEEI 4 × TZdEEI 7 | 3679 | 1964 | 1854 | 2627 |
| TZdEEI 4 × TZdEEI 9 | 3405 | 1696 | 1928 | 2394 |
| TZdEEI 4 × TZdEEI 11 | 3128 | 1453 | 2498 | 2435 |
| TZdEEI 4 × TZdEEI 12 | 3817 | 1844 | 2099 | 2651 |
| TZdEEI 4 × TZdEEI 13 | 2636 | 644 | 1139 | 1564 |
| TZdEEI 4 × TZEEI 58 | 4253 | 1810 | 1892 | 2682 |
| TZdEEI 4 × TZEEI 63 | 4102 | 1432 | 1823 | 2581 |
| TZdEEI 4 × TZEEI 79 | 3233 | 1965 | 1020 | 2173 |
| TZdEEI 4 × TZEEI 95 | 4077 | 2252 | 2366 | 2931 |
| TZdEEI 5 × TZdEEI 7 | 3916 | 3269 | 2102 | 3201 |
| TZdEEI 5 × TZdEEI 9 | 4846 | 3599 | 2051 | 3940 |
| TZdEEI 5 × TZdEEI 11 | 4680 | 2866 | 3303 | 3834 |
| TZdEEI 5 × TZdEEI 12 | 3585 | 3114 | 1036 | 3122 |
| TZdEEI 5 × TZdEEI 13 | 3473 | 2336 | 1346 | 2637 |
| TZdEEI 5 × TZEEI 58 | 3979 | 3190 | 1933 | 3258 |
| TZdEEI 5 × TZEEI 63 | 4169 | 2842 | 1781 | 3133 |
| TZdEEI 5 × TZEEI 79 | 4386 | 2925 | 2602 | 3427 |
| TZdEEI 5 × TZEEI 95 | 3579 | 3385 | 1559 | 2983 |
| TZdEEI 7 × TZdEEI 9 | 4600 | 3937 | 2680 | 3815 |
| TZdEEI 7 × TZdEEI 11 | 4483 | 3255 | 3022 | 3793 |
| TZdEEI 7 × TZdEEI 12 | 4081 | 3619 | 3254 | 3694 |
| TZdEEI 7 × TZdEEI 13 | 4108 | 2382 | 2439 | 3097 |
| TZdEEI 7 × TZEEI 58 | 3980 | 4065 | 2320 | 3805 |
| TZdEEI 7 × TZEEI 63 | 5218 | 3551 | 3611 | 4177 |
| TZdEEI 7 × TZEEI 79 | 4387 | 4437 | 3463 | 4215 |
| TZdEEI 7 × TZEEI 95 | 4337 | 4040 | 2160 | 3750 |
| TZdEEI 9 × TZdEEI 11 | 4066 | 1965 | 2607 | 3000 |
| TZdEEI 9 × TZdEEI 12 | 5008 | 3516 | 3599 | 4033 |
| TZdEEI 9 × TZdEEI 13 | 4598 | 888 | 1517 | 2357 |
| TZdEEI 9 × TZEEI 58 | 4712 | 2878 | 2183 | 3405 |
| TZdEEI 9 × TZEEI 63 | 4051 | 2569 | 2407 | 3178 |
| TZdEEI 9 × TZEEI 79 | 4530 | 3747 | 2838 | 3717 |
| TZdEEI 9 × TZEEI 95 | 4831 | 2974 | 2325 | 3549 |
| TZdEEI 11 × TZdEEI 12 | 4323 | 3463 | 2867 | 3738 |
| TZdEEI 11 × TZdEEI 13 | 4450 | 1336 | 2152 | 2616 |
| TZdEEI 11 × TZEEI 58 | 5066 | 2527 | 3150 | 3798 |
| TZdEEI 11 × TZEEI 63 | 5043 | 2634 | 3017 | 3796 |
| TZdEEI 11 × TZEEI 79 | 3756 | 3620 | 2602 | 3429 |
| TZdEEI 11 × TZEEI 95 | 3358 | 2481 | 3092 | 3062 |
| TZdEEI 12 × TZdEEI 13 | 4742 | 1902 | 2864 | 3180 |
| TZdEEI 12 × TZEEI 58 | 4245 | 3446 | 3170 | 3681 |
| TZdEEI 12 × TZEEI 63 | 4349 | 2884 | 3883 | 3682 |
| TZdEEI 12 × TZEEI 79 | 3767 | 3355 | 3271 | 3612 |
| TZdEEI 12 × TZEEI 95 | 3673 | 3648 | 2874 | 3638 |
| TZdEEI 13 × TZEEI 58 | 4202 | 1300 | 2596 | 2567 |
| TZdEEI 13 × TZEEI 63 | 3974 | 2323 | 3121 | 3190 |
| TZdEEI 13 × TZEEI 79 | 3544 | 2469 | 2623 | 3029 |
| TZdEEI 13 × TZEEI 95 | 3769 | 2270 | 3149 | 3066 |
| TZEEI 58 × TZEEI 63 | 2674 | 1535 | 706 | 1533 |
| TZEEI 58 × TZEEI 79 | 3050 | 3574 | 2324 | 3063 |
| TZEEI 58 × TZEEI 95 | 3889 | 2767 | 2491 | 3399 |
| TZEEI 63 × TZEEI 79 | 4225 | 3608 | 1789 | 3445 |
| TZEEI 63 × TZEEI 95 | 3860 | 2306 | 2705 | 2888 |
| TZEEI 79 × TZEEI 95 | 3495 | 2719 | 2819 | 3237 |
| Check 1 | 3247 | 2837 | 1278 | 2859 |
| Check 2 | 3882 | 3202 | 2224 | 3233 |
| Check 3 | 3445 | 2556 | 1904 | 3052 |
| Check 4 | 3164 | 2507 | 2032 | 2629 |
| Mean | 4013 | 2729 | 2429 | 3183 |
| Hybrid | Yield (kg ha−1) | EPP | ASI (Days) | EASP | STGR | SDR1 | SDR2 | ESP1 | ESP2 | MI | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ST | NS | ACR | ST | NS | ST | NS | ST | NS | ST | ||||||
| TZdEEI 7 × TZEEI 79 | 4112 | 4928 | 4438 | 1.00 | 0.94 | 1.75 | 1.14 | 4.5 | 2.7 | 3.3 | 2.7 | 3.8 | 26.7 | 28.6 | 13.76 |
| TZdEEI 9 × TZEEI 79 | 3445 | 5163 | 4132 | 0.90 | 0.89 | 1.48 | 0.39 | 4.4 | 2.7 | 3.5 | 2.8 | 3.9 | 31.0 | 34.8 | 10.64 |
| TZdEEI 7 × TZdEEI 12 | 3497 | 4757 | 4001 | 1.02 | 0.88 | 0.99 | 0.46 | 4.7 | 3.1 | 4.0 | 2.6 | 3.9 | 38.3 | 40.3 | 10.21 |
| TZdEEI 1 × TZdEEI 7 | 3844 | 4991 | 4303 | 1.09 | 1.05 | 1.79 | 0.86 | 4.5 | 3.0 | 6.0 | 2.9 | 3.8 | 45.3 | 48.8 | 9.76 |
| TZdEEI 7 × TZdEEI 9 | 3518 | 5047 | 4130 | 0.96 | 0.89 | 0.73 | 0.71 | 4.7 | 3.1 | 3.5 | 3.3 | 4.5 | 46.2 | 53.2 | 9.68 |
| TZdEEI 9 × TZdEEI 12 | 3544 | 4328 | 3857 | 1.00 | 1.12 | 0.71 | 1.07 | 4.7 | 3.0 | 4.0 | 4.0 | 4.9 | 26.2 | 33.3 | 9.26 |
| TZdEEI 11 × TZEEI 79 | 3281 | 5274 | 4078 | 0.89 | 0.97 | 1.85 | 1.37 | 4.2 | 2.6 | 3.8 | 3.3 | 3.9 | 24.8 | 30.4 | 9.18 |
| TZdEEI 1 × TZEEI 79 | 3134 | 5078 | 3912 | 0.90 | 0.74 | 1.82 | 1.38 | 4.5 | 2.7 | 2.7 | 4.3 | 4.5 | 28.5 | 38.0 | 8.66 |
| TZdEEI 12 × TZEEI 95 | 3390 | 5602 | 4275 | 0.95 | 0.75 | 1.37 | 0.91 | 4.5 | 2.9 | 3.6 | 3.8 | 4.8 | 30.0 | 38.6 | 8.57 |
| TZdEEI 12 × TZEEI 79 | 3327 | 5038 | 4011 | 0.91 | 0.94 | 1.62 | 0.96 | 4.8 | 3.1 | 3.7 | 3.6 | 4.1 | 42.8 | 42.3 | 8.24 |
| TZdEEI 58 × TZEEI 79 | 3158 | 4360 | 3638 | 0.85 | 0.88 | 1.80 | 1.65 | 4.9 | 3.1 | 3.4 | 2.5 | 4.2 | 36.1 | 42.9 | 8.19 |
| TZdEEI 1 × TZdEEI 12 | 3285 | 4633 | 3824 | 1.01 | 0.95 | 1.58 | 0.51 | 4.8 | 3.0 | 3.2 | 4.2 | 4.3 | 26.5 | 32.5 | 7.98 |
| TZdEEI 5 × TZEEI 79 | 2818 | 5402 | 3852 | 0.84 | 0.89 | 0.62 | 1.21 | 4.8 | 2.3 | 3.4 | 4.8 | 5.3 | 29.7 | 37.1 | 6.49 |
| TZdEEI 7 × TZEEI 63 | 3571 | 5142 | 4199 | 0.88 | 0.92 | 1.84 | 0.92 | 4.7 | 2.6 | 3.5 | 3.9 | 4.6 | 52.2 | 58.0 | 6.44 |
| TZdEEI 7 × TZEEI 95 | 3413 | 4330 | 3780 | 0.97 | 0.99 | 1.90 | 1.21 | 5.5 | 3.2 | 4.1 | 3.4 | 4.8 | 31.4 | 36.0 | 5.93 |
| Check 2 † | 2876 | 4391 | 3482 | 0.89 | 0.92 | 1.91 | 1.79 | 4.6 | 2.7 | 4.5 | 4.0 | 4.8 | 21.1 | 22.6 | 5.30 |
| Check 1 | 2316 | 4819 | 3318 | 0.85 | 0.84 | 2.64 | 1.99 | 5.5 | 2.9 | 4.5 | 3.6 | 4.8 | 27.6 | 34.1 | 0.65 |
| Check 3 | 2339 | 3513 | 2808 | 0.84 | 0.94 | 2.74 | 1.42 | 5.5 | 3.3 | 5.6 | 4.6 | 5.7 | 3.8 | 6.3 | −2.40 |
| Check 4 | 2349 | 4114 | 3055 | 0.80 | 1.07 | 2.20 | 1.21 | 5.6 | 3.2 | 4.8 | 5.0 | 5.9 | 38.2 | 36.6 | −3.00 |
| TZdEEI 5 × TZdEEI 13 | 2006 | 4161 | 2868 | 0.77 | 0.93 | 3.42 | 1.88 | 5.7 | 3.2 | 3.0 | 5.6 | 6.2 | 44.6 | 51.8 | −7.26 |
| TZdEEI 1 × TZdEEI 13 | 1827 | 4668 | 2964 | 0.79 | 0.90 | 2.73 | 1.17 | 5.7 | 3.0 | 4.7 | 6.2 | 6.2 | 44.8 | 49.1 | −7.39 |
| TZdEEI 4 × TZdEEI 11 | 1801 | 2456 | 2063 | 0.75 | 0.90 | 2.07 | 0.60 | 5.7 | 3.6 | 3.2 | 5.8 | 6.5 | 48.1 | 52.3 | −8.98 |
| TZdEEI 13 × TZEEI 58 | 1732 | 4589 | 2875 | 0.55 | 0.95 | 1.47 | 1.64 | 6.1 | 2.9 | 3.2 | 6.4 | 7.2 | 47.5 | 53.8 | −9.58 |
| TZdEEI 4 × TZEEI 58 | 1837 | 4604 | 2944 | 0.61 | 1.00 | 3.81 | 1.36 | 6.0 | 3.2 | 5.3 | 5.7 | 6.5 | 28.5 | 31.5 | −10.53 |
| TZdEEI 4 × TZEEI 63 | 1562 | 4055 | 2559 | 0.68 | 0.93 | 3.10 | 2.12 | 6.2 | 3.0 | 4.8 | 5.9 | 6.7 | 33.3 | 37.4 | −11.37 |
| TZdEEI 11 × TZdEEI 13 | 1608 | 3857 | 2508 | 0.68 | 0.89 | 3.27 | 1.81 | 6.1 | 2.9 | 3.3 | 6.9 | 7.4 | 45.2 | 49.8 | −11.84 |
| TZdEEI 9 × TZdEEI 13 | 1097 | 4694 | 2536 | 0.60 | 1.02 | 3.86 | 0.62 | 6.6 | 3.1 | 3.6 | 6.7 | 7.1 | 76.6 | 84.9 | −16.38 |
| TZdEEI 4 × TZdEEI 13 | 808 | 4186 | 2159 | 0.61 | 1.04 | 3.58 | 0.84 | 6.8 | 3.1 | 4.1 | 6.9 | 7.8 | 27.7 | 35.4 | −17.80 |
| TZdEEI 58 × TZEEI 63 | 1247 | 3557 | 2171 | 0.52 | 0.88 | 4.46 | 2.58 | 6.3 | 3.3 | 5.3 | 5.7 | 6.7 | 53.3 | 59.4 | −17.98 |
| Mean | 2629 | 4546 | 3396 | 0.85 | 0.98 | 2.21 | 1.20 | 5.3 | 3.0 | 4.0 | 4.6 | 5.4 | 40.6 | 45.5 | |
| LSD | 758 | 1216 | 663 | 0.15 | 0.75 | 1.39 | 1.13 | 0.74 | 0.49 | 1.64 | 1.21 | 0.97 | 26.91 | 28.79 | |
| S/N | Parent | YIELD ‡ (kg ha−1) | SDR1 | SDR2 | ESP1 | ESP2 | STGR | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Optimal | Striga | Drought | Across | |||||||
| 1 | TZdEEI 1 | 21 | 102 | −1 | −46 | 0.09 | −0.13 | 0.57 | 0.42 | 0.08 |
| 2 | TZdEEI 4 | −585 ** | −1119 ** | −689 ** | −860 ** | 0.69 ** | 1.02 ** | −10.15 ** | −10.83 * | 0.32 |
| 3 | TZdEEI 5 | 4 | 279 ** | −521 ** | −11 | 0.01 | −0.13 | 7.30 | 7.72 | −0.03 |
| 4 | TZdEEI 7 | 425 ** | 767 ** | 271 | 571 ** | −0.79 ** | −0.73 ** | 13.45 ** | 12.69 ** | 0.07 |
| 5 | TZdEEI 9 | 363 * | 103 | −83 | 63 | −0.31 | −0.16 | 4.77 | 4.42 | −0.43 * |
| 6 | TZdEEI 11 | 101 | −164 | 390 ** | −166 | 0.44 ** | 0.12 | −3.33 | −3.21 | −0.12 |
| 7 | TZdEEI 12 | 150 | 602 ** | 338 * | 334 * | −0.59 ** | −0.61 ** | −5.23 | −6.01 | 0.57 ** |
| 8 | TZdEEI 13 | −256 | −1026 ** | −43 | −424 ** | 1.39 ** | 1.24 ** | 2.05 | 2.69 | −0.38 * |
| 9 | TZEEI 58 | −29 | −40 | −114 | 38 | 0.09 | 0.04 | 8.32 * | 8.09 | 0.07 |
| 10 | TZEEI 63 | 123 | −173 | 133 | −37 | 0.16 | 0.34 ** | 5.70 | 6.87 | 0.07 |
| 11 | TZEEI 79 | −210 | 572 ** | 82 | 313 * | −1.06 ** | −0.93 ** | −11.93 ** | −11.58 ** | −0.08 |
| 12 | TZEEI 95 | −106 | 97 ** | 237 | 226 | −0.11 | −0.06 | −11.50 ** | −11.26 ** | −0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaibu, A.S.; Badu-Apraku, B.; Ayo-Vaughan, M.A. Enhancing Drought Tolerance and Striga hermonthica Resistance in Maize Using Newly Derived Inbred Lines from the Wild Maize Relative, Zea diploperennis. Agronomy 2021, 11, 177. https://doi.org/10.3390/agronomy11010177
Shaibu AS, Badu-Apraku B, Ayo-Vaughan MA. Enhancing Drought Tolerance and Striga hermonthica Resistance in Maize Using Newly Derived Inbred Lines from the Wild Maize Relative, Zea diploperennis. Agronomy. 2021; 11(1):177. https://doi.org/10.3390/agronomy11010177
Chicago/Turabian StyleShaibu, Abdulwahab S., Baffour Badu-Apraku, and Monininuola A. Ayo-Vaughan. 2021. "Enhancing Drought Tolerance and Striga hermonthica Resistance in Maize Using Newly Derived Inbred Lines from the Wild Maize Relative, Zea diploperennis" Agronomy 11, no. 1: 177. https://doi.org/10.3390/agronomy11010177
APA StyleShaibu, A. S., Badu-Apraku, B., & Ayo-Vaughan, M. A. (2021). Enhancing Drought Tolerance and Striga hermonthica Resistance in Maize Using Newly Derived Inbred Lines from the Wild Maize Relative, Zea diploperennis. Agronomy, 11(1), 177. https://doi.org/10.3390/agronomy11010177