Early-Stage Phenotyping of Root Traits Provides Insights into the Drought Tolerance Level of Soybean Cultivars


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.1.1. Controlled Conditions
2.1.2. Field Conditions
2.2. Sampling and Data Collection in Controlled Conditions
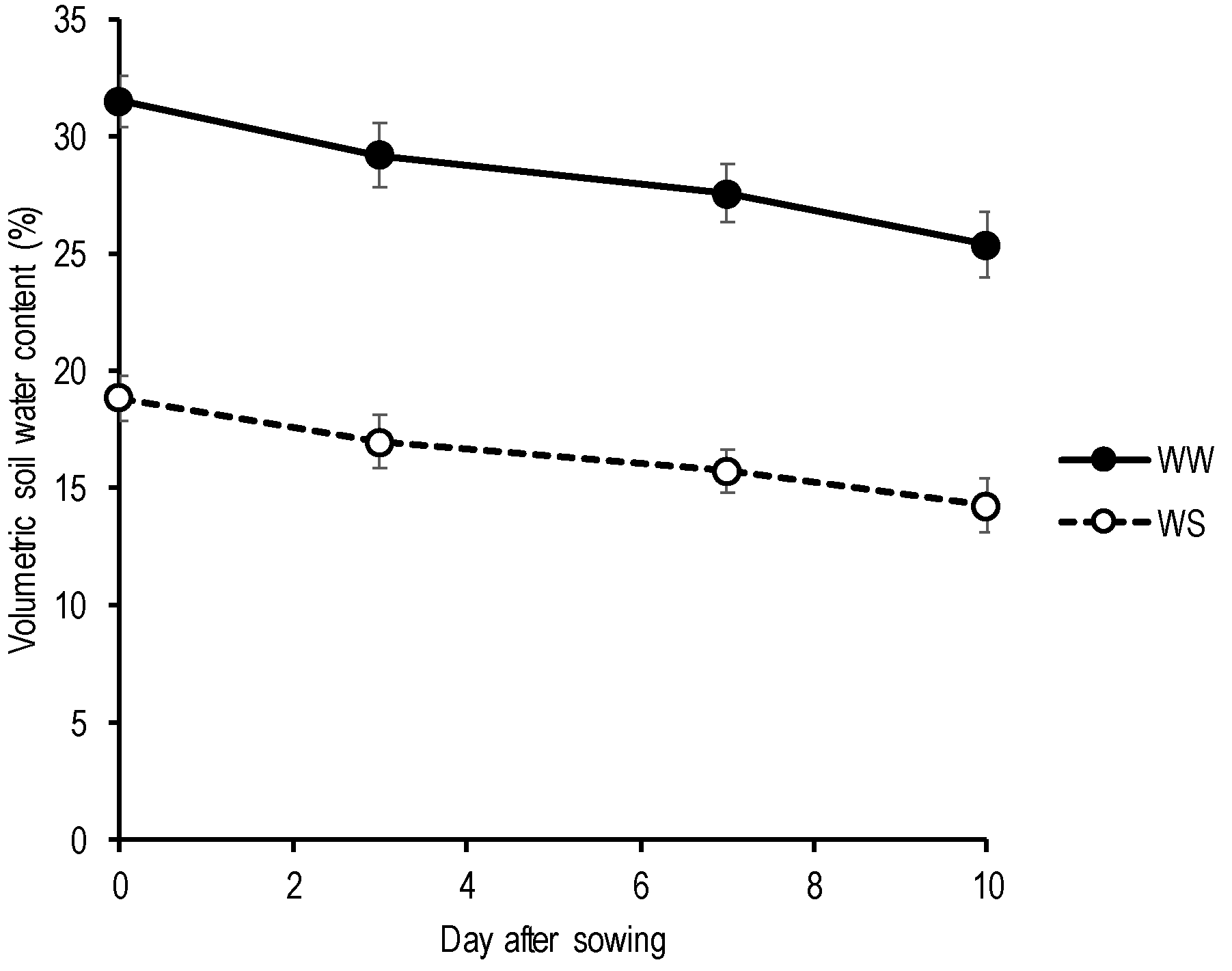
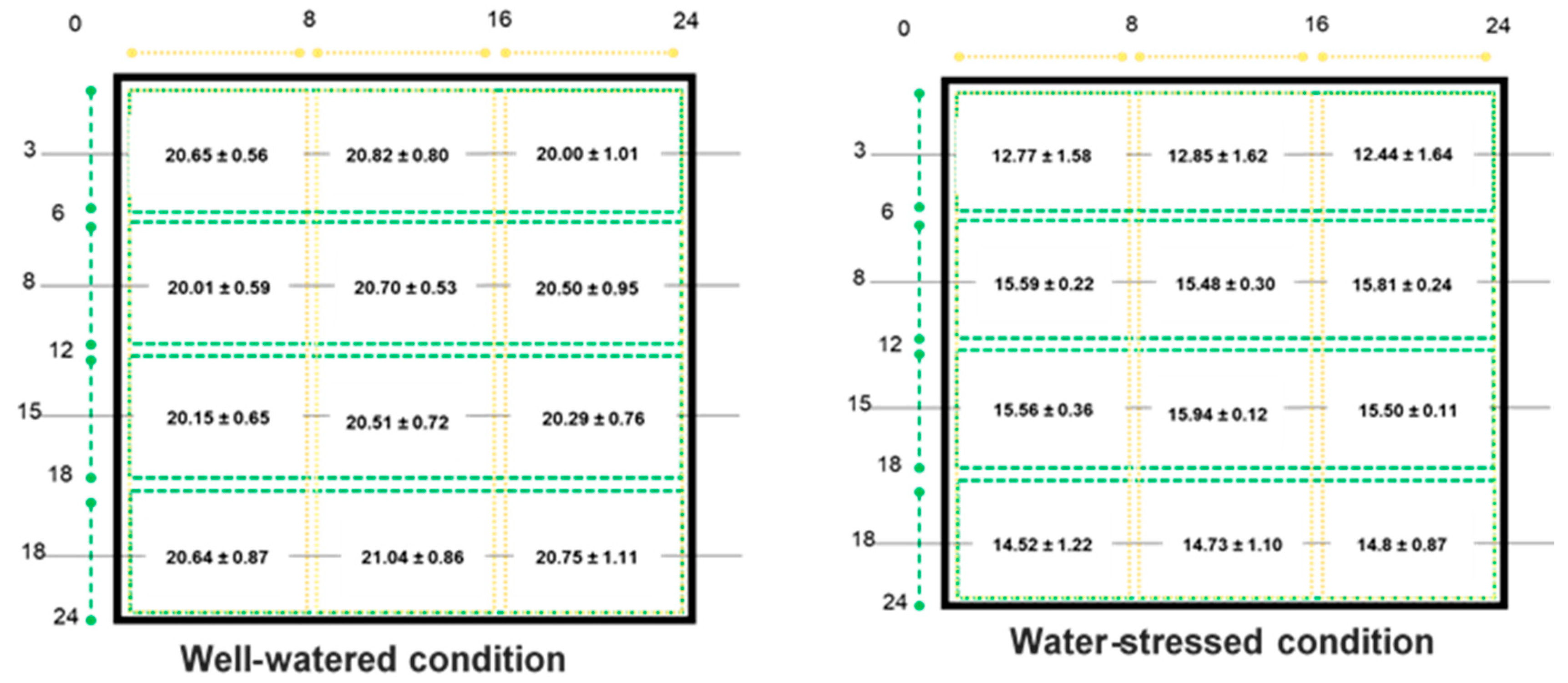
2.2.1. Soil Moisture Status
2.2.2. Shoot and Root Morpho-Physiological Traits
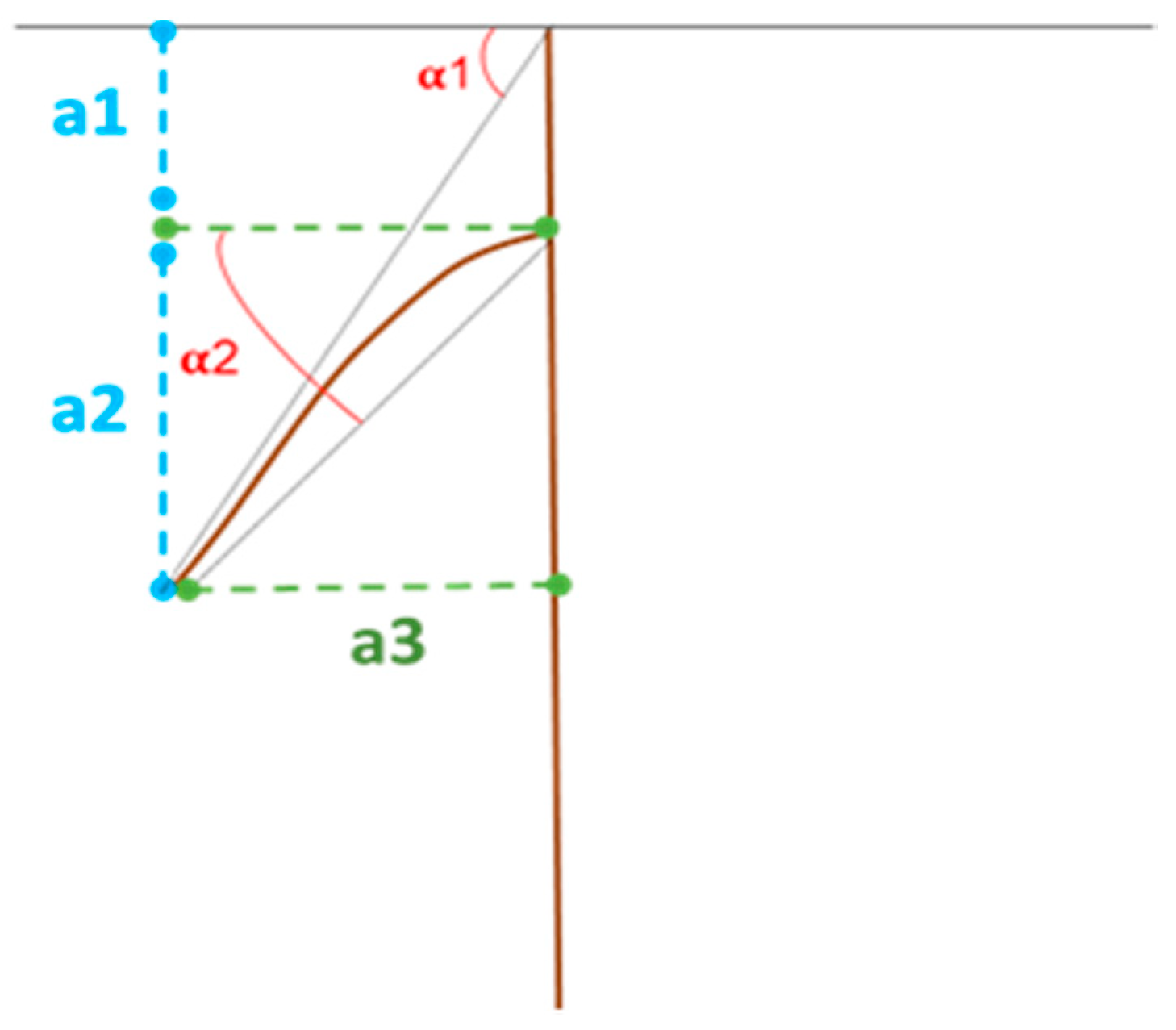
2.2.3. Root Architectural Traits
- α1: the angle between the longest lateral root and the horizontal line of soil (as average on both sides);
- α2: angle between the longest lateral root and the horizontal line from the point of insertion of this root on taproot (as averaged on both sides);
- a1: vertical distance from soil to longest lateral root insertion on taproot;
- a2: vertical distance between the end of a1 and the tip of the longest lateral root;
- a3: horizontal distance between the tip of the longest lateral root and the taproot.
2.3. Sampling and Data Collection under Field Conditions
- With: CC0: canopy cover when emergence has occurred;
- CGC: canopy growth coefficient;
- t: time from emergence stage (in days).
2.4. Statistical Analyses
3. Results
3.1. Difference among Cultivars for Shoot and Root Traits under Controlled Conditions
3.2. Impact of Water-Stressed Conditions on Shoot and Root Traits under Controlled Conditions
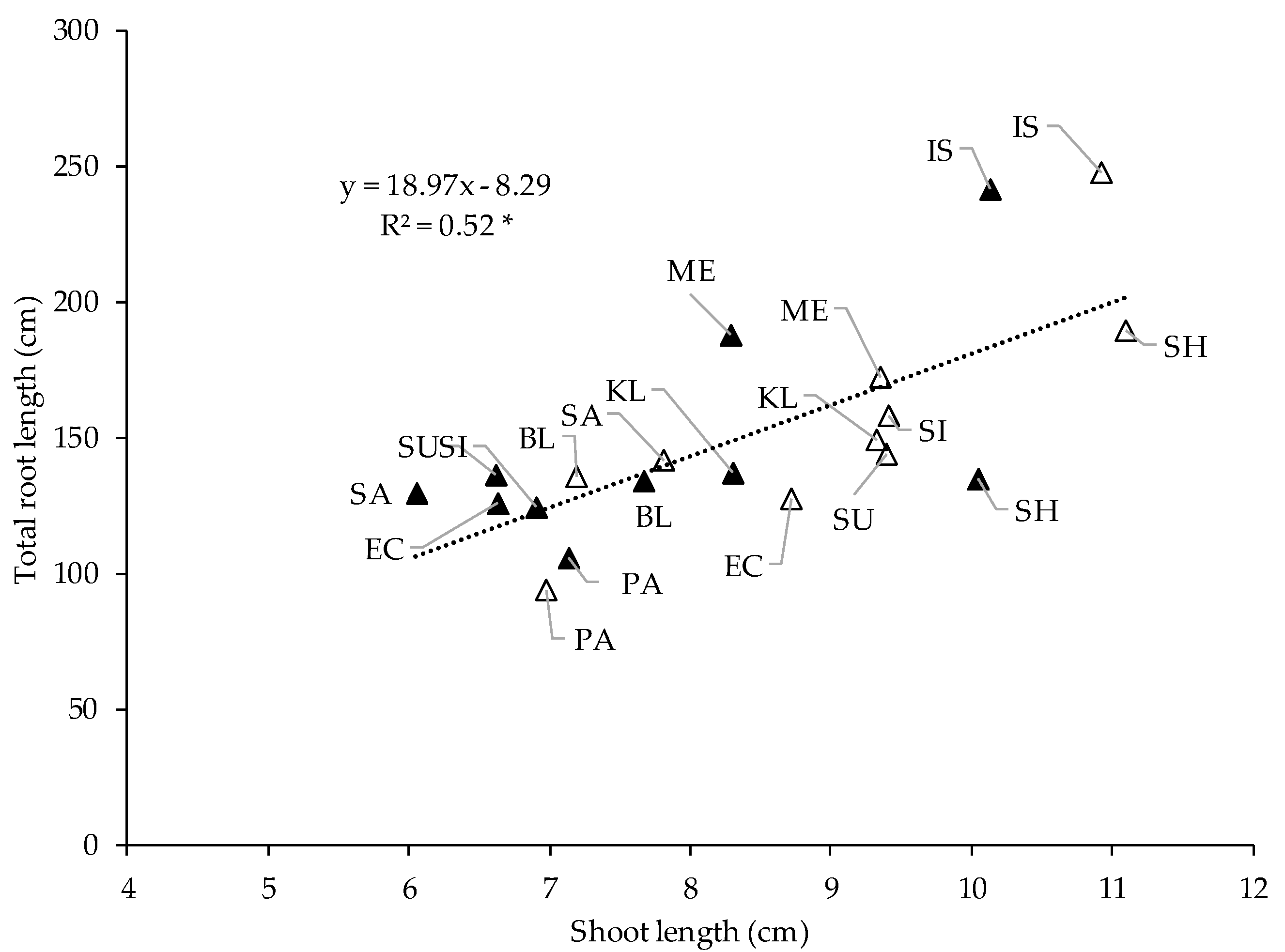
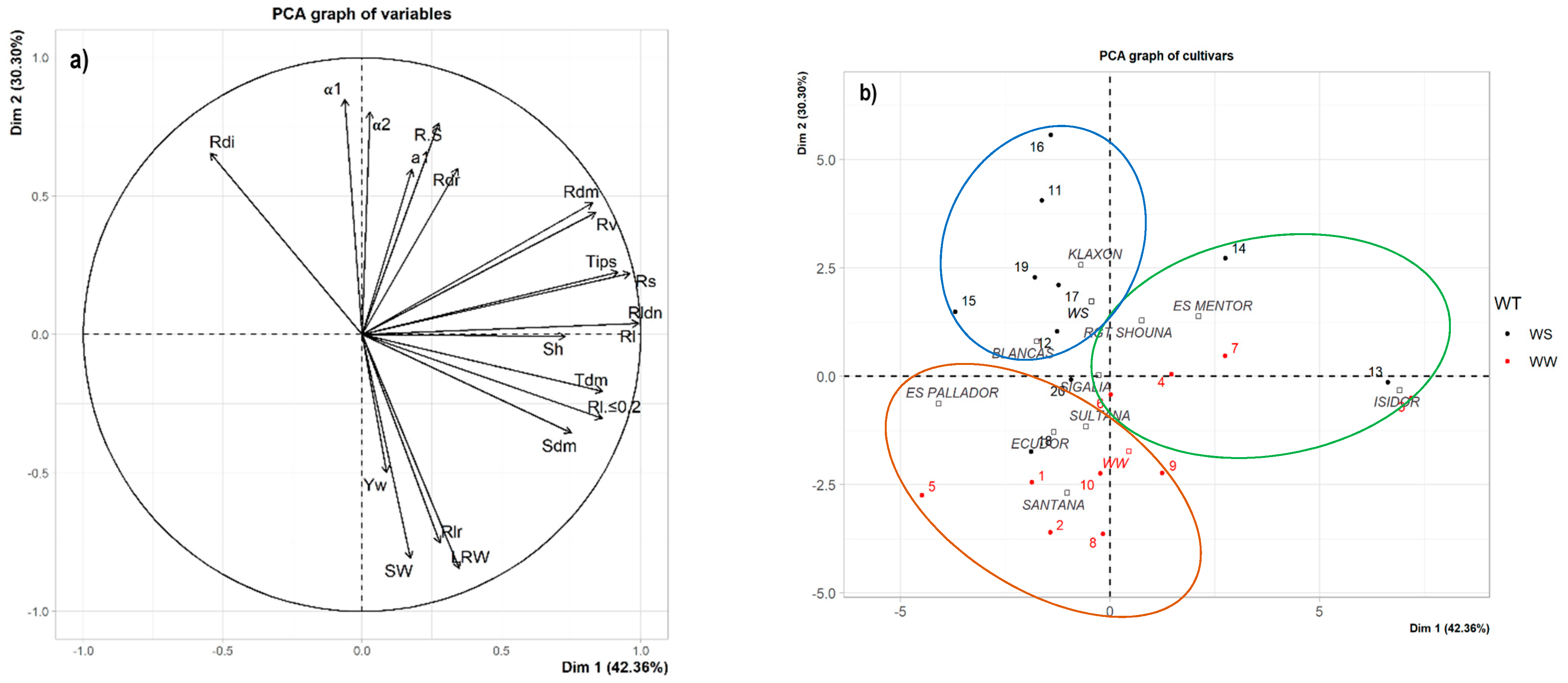
3.3. Relationships between Studied Traits during the Early Growth under Controlled Conditions
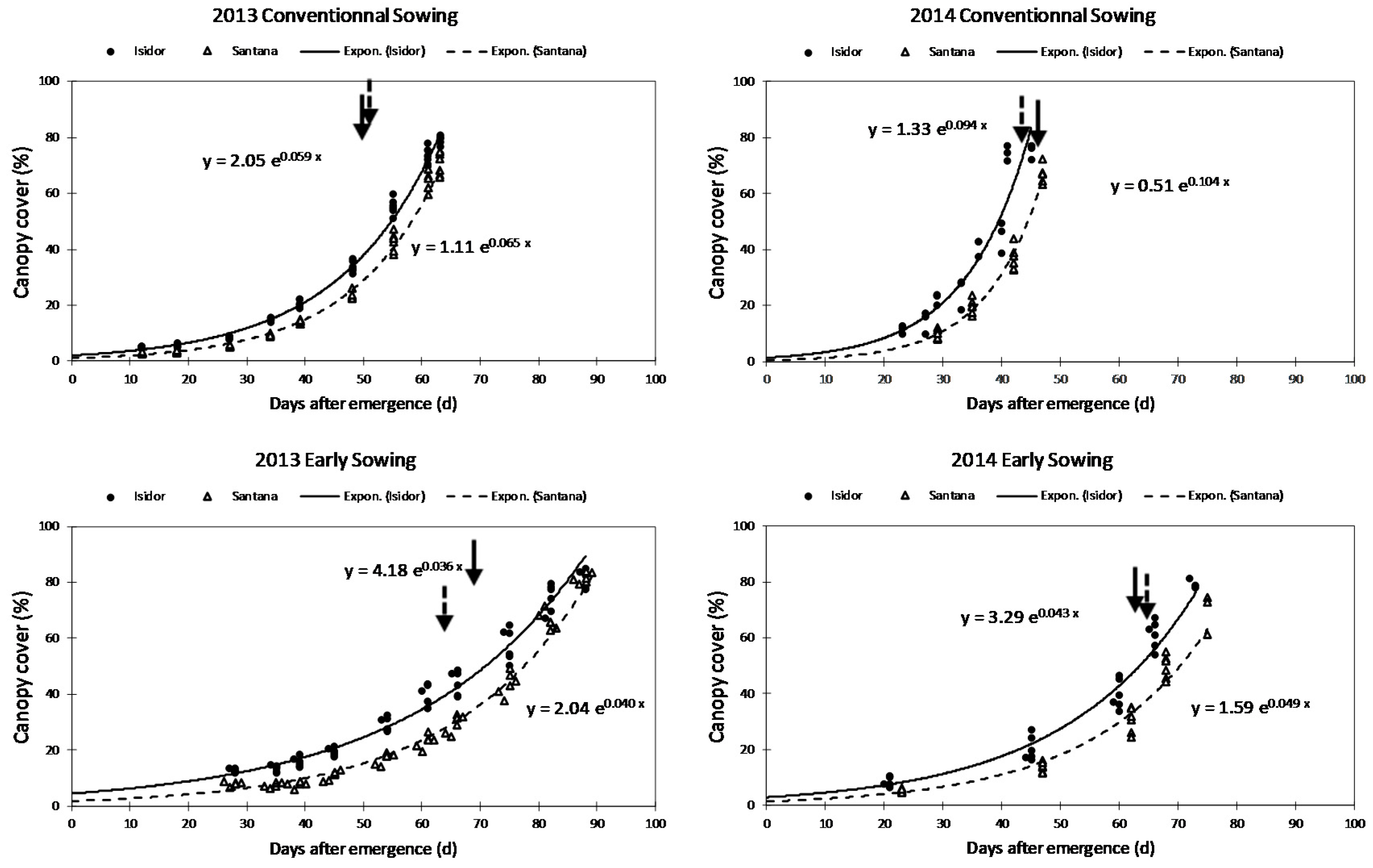
3.4. Comparison of Two Contrasted Cultivars for Early Growth under Field Conditions
4. Discussion
4.1. Shoot and Root Growth for Soybean Cultivars under Well-Watered Conditions
4.2. Impact of Water-Stressed Conditions on Shoot and Root Growth
4.3. Correlations among Root and Shoot Traits
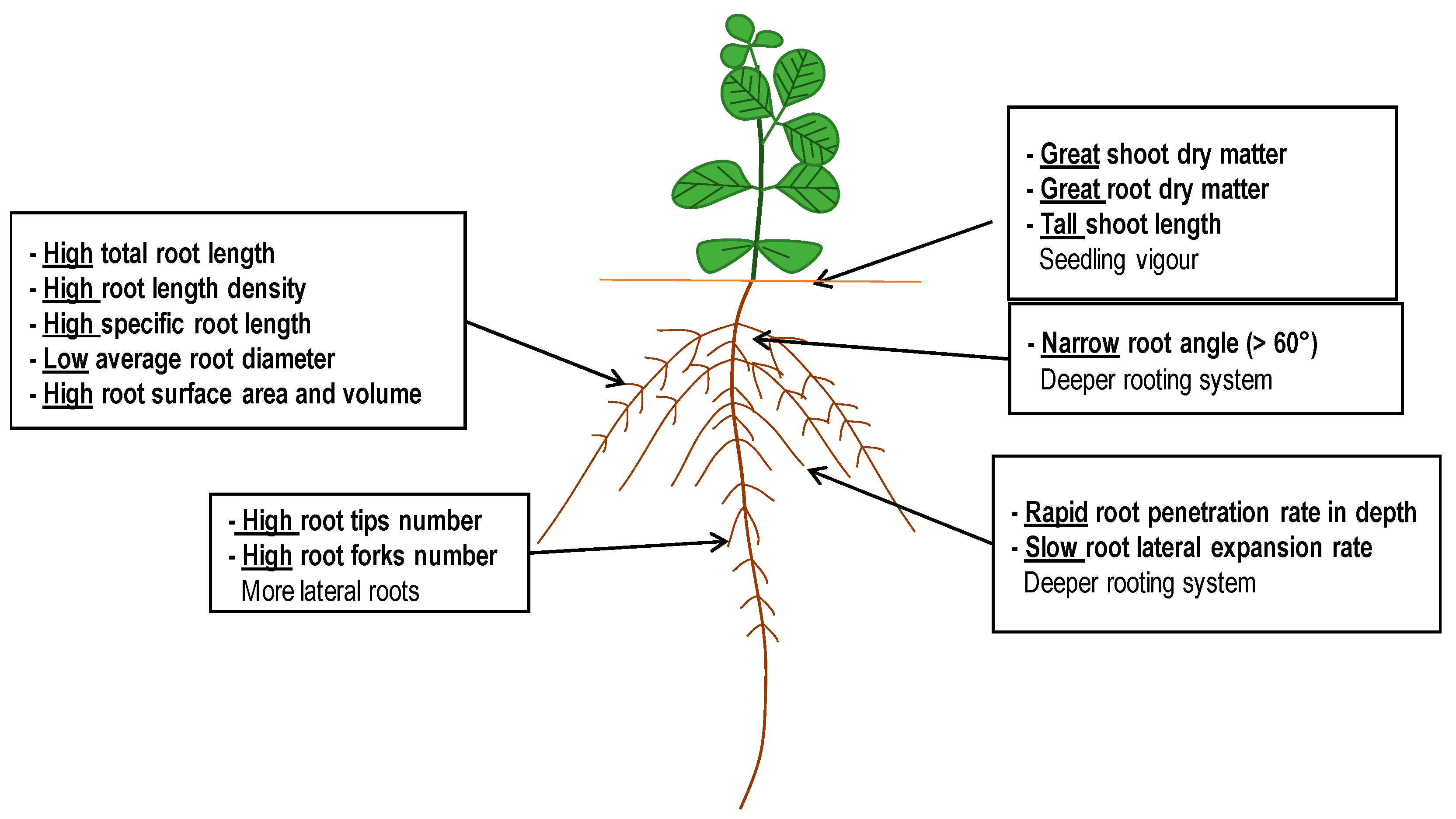
4.4. Soybean Ideotypes during Early Growth under Water-Stressed Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Set 1 | Set 2 | |||
|---|---|---|---|---|
| WW | WS | WW | WS | |
| Blancas | 4 | 4 | 4 | 4 |
| Ecudor | 3 | 3 | 5 | 5 |
| ES Mentor | 3 | 3 | 5 | 5 |
| ES Pallador | 3 | 3 | 5 | 5 |
| Isidor | 4 | 4 | 5 | 4 |
| Klaxon | 4 | 4 | 5 | 5 |
| RGT Shouna | 4 | 4 | 5 | 5 |
| Santana | 4 | 4 | 4 | 4 |
| Sigalia | 4 | 3 | 4 | 5 |
| Sultana | 3 | 4 | 5 | 4 |
| Type of traits | Variable |
|---|---|
| Morpho-physiological traits (shoot or root) | Average root diameter (mm) |
| Length of roots with diameter ≤ 0.2 mm (cm) | |
| Length of roots with diameter between 0.2 and 0.4 mm (cm) | |
| Length of roots with diameter between 0.4 and 0.6 mm (cm) | |
| Length of roots with diameter between 0.6 and 0.8 mm (cm) | |
| Length of roots with diameter between 0.8 and 1 mm (cm) | |
| Length of roots with diameter between 1 and 2 mm (cm) | |
| Length of roots with diameter between 2 and 3 mm (cm) | |
| Root dry matter (mg) | |
| Root length density (cm cm−3) | |
| Root surface area (cm2) | |
| Root: shoot ratio (mg/mg) | |
| Root tips number | |
| Root volume (cm3) | |
| Shoot length (cm) | |
| Shoot dry matter (mg) | |
| Specific root length (cm mg−1) | |
| Specific root surface (cm2 mg−1) | |
| Total dry matter (mg) | |
| Total root length (cm) | |
| Plant water potential (MPa) | |
| Architectural traits (root) | Distance from soil to longest lateral root insertion on taproot (cm) |
| Root angle 1 (°) | |
| Root angle 2 (°) | |
| Root lateral expansion rate (cm day−1) | |
| Root penetration rate in depth (cm day−1) |

| Cultivars | Root Lateral Expansion Rate (cm day−1) | Root Penetration Rate in Depth (cm day−1) | Distance from Soil to Longest Lateral Root Insertion on Taproot (cm) | Root Angle 2 (°) |
| Well-watered conditions | ||||
| Blancas | 1.65 ± 0.15 | 2.91 ± 0.40 | 1.28 ± 0.61 | 35.35 ± 3.71 |
| Ecudor | 1.97± 0.30 | 3.20 ± 0.48 | 1.35 ± 0.74 | 33.18 ± 2.93 |
| ES Mentor | 1.74 ± 0.31 | 3.66 ± 0.42 | 2.59 ± 0.65 | 37.52 ± 5.24 |
| ES Pallador | 1.50 ± 0.29 | 3.50 ± 0.48 | 2.58 ± 0.54 | 36.06 ± 4.12 |
| Isidor | 1.94 ± 0.33 | 4.04 ± 0.41 | 1.87 ± 0.55 | 43.01 ± 2.86 |
| Klaxon | 2.00 ± 0.29 | 3.87 ± 0.34 | 1.23 ± 0.37 | 44.21 ± 4.46 |
| RGT Shouna | 1.74 ± 0.21 | 3.19 ± 0.47 | 2.20 ± 0.92 | 46.37 ± 4.90 |
| Santana | 2.08 ± 0.30 | 3.52 ± 0.32 | 1.43 ± 0.36 | 32.04 ± 4.27 |
| Sigalia | 1.95 ± 0.22 | 3.87 ± 0.37 | 1.69 ± 0.50 | 42.24 ± 4.49 |
| Sultana | 1.44 ± 0.23 | 3.79 ± 0.51 | 2.09 ± 1.04 | 46.74 ± 5.31 |
| Significance | NS | NS | NS | NS |
| Water-stressed conditions | ||||
| Blancas | 1.09 ± 0.19 | 4.29 ± 0.30 | 3.33 ± 1.15 | 48.69 ± 5.30 |
| Ecudor | 1.44 ± 0.23 | 3.67 ± 0.48 | 4.03 ± 1.20 | 40.49 ± 6.36 |
| ES Mentor | 1.21 ± 0.21 | 4.00 ± 0.40 | 5.22 ± 1.55 | 45.97 ± 4.04 |
| ES Pallador | 1.39 ± 0.33 | 3.83 ± 0.24 | 2.64 ± 0.78 | 47.53 ± 4.59 |
| Isidor | 1.39 ± 0.23 | 4.84 ± 0.22 | 4.49 ± 1.22 | 41.48 ± 2.72 |
| Klaxon | 0.81 ± 0.16 | 4.67 ± 0.31 | 4.19 ± 1.49 | 51.90 ± 6.53 |
| RGT Shouna | 1.17 ± 0.18 | 3.79 ± 0.32 | 1.86 ± 0.84 | 47.65 ± 7.56 |
| Santana | 1.45 ± 0.26 | 3.66 ± 0.48 | 1.94 ± 0.71 | 43.23 ± 6.38 |
| Sigalia | 1.25 ± 0.20 | 3.97 ± 0.29 | 2.32 ± 0.66 | 49.08 ± 5.03 |
| Sultana | 1.06 ± 0.22 | 4.13 ± 0.34 | 4.45 ± 1.02 | 40.31 ± 3.90 |
| Significance | NS | NS | NS | NS |
| Root Dry Matter (mg) | Root: Shoot Ratio | Shoot Dry Matter (mg) | Total Dry Matter (mg) | Plant Water Potential (MPa) | Average Root Diameter (mm) | Length of Roots with Diameter ≤ 0.2 mm (cm) | Root Length Density (cm cm−3) | Root Surface Area (cm2) | Root Tips Number | Root Volume (cm3) | Shoot Length (cm) | Specific Root Length (cm mg−1) | Specific Root Surface (cm2 mg−1) | Total Root Length (cm) | Distance from Soil to Longest Lateral Root Insertion on Tap Root (cm) | Root Angle 1 (°) | Root Angle 2 (°) | Root Lateral Expansion Rate (cm day−1) | Root Penetration Rate in Depth (cm day−1) | Thousand Seed Weight (g) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root dry matter (mg) | 1 | ||||||||||||||||||||
| Root: shoot ratio | 0.83 *** | 1 | |||||||||||||||||||
| Shoot dry matter (mg) | 0.50 | −0.07 | 1 | ||||||||||||||||||
| Total dry matter (mg) | 0.69 * | 0.17 | 0.97 *** | 1 | |||||||||||||||||
| Plant water potential (MPa) | −0.21 | 0.05 | −0.41 | −0.4 | 1 | ||||||||||||||||
| Average root diameter (mm) | −0.04 | 0.47 | −0.77 ** | −0.65 * | 0.04 | 1 | |||||||||||||||
| Length of roots with diameter ≤ 0.2 mm (cm) | 0.89 *** | 0.56 | 0.70 * | 0.83 *** | −0.16 | −0.4 | 1 | ||||||||||||||
| Root length density (cm cm−3) | 0.95 *** | 0.66 * | 0.67 * | 0.84 *** | −0.24 | −0.27 | 0.95 *** | 1 | |||||||||||||
| Root surface area (cm2) | 0.97 *** | 0.74 ** | 0.60 * | 0.77 ** | −0.27 | −0.11 | 0.91 *** | 0.99 *** | 1 | ||||||||||||
| Root tips number | 0.89 *** | 0.62 * | 0.67 * | 0.81 *** | −0.23 | −0.23 | 0.93 *** | 0.95 *** | 0.95 *** | 1 | |||||||||||
| Root volume (cm3) | 0.97 *** | 0.82 ** | 0.47 | 0.66 * | −0.28 | 0.08 | 0.84 *** | 0.93 *** | 0.98 *** | 0.91 *** | 1 | ||||||||||
| Shoot length (cm) | 0.79 ** | 0.61 * | 0.53 | 0.66 * | −0.28 | 0.04 | 0.71 ** | 0.84 *** | 0.89 *** | 0.85 *** | 0.91 *** | 1 | |||||||||
| Specific root length (cm mg−1) | −0.57 | −0.75 ** | 0.19 | 0.00 | 0.11 | −0.53 | −0.20 | −0.31 | −0.41 | −0.23 | −0.51 | −0.23 | 1 | ||||||||
| Specific root surface (cm2 mg−1) | −0.67 * | −0.67 * | −0.09 | −0.26 | 0.01 | −0.11 | −0.45 | −0.47 | −0.49 | −0.36 | −0.52 | −0.16 | 0.88 *** | 1 | |||||||
| Total root length (cm) | 0.95 *** | 0.66 * | 0.69 * | 0.84 *** | −0.24 | −0.27 | 0.95 *** | 1.00 | 0.99 *** | 0.95 *** | 0.93 *** | 0.84 *** | −0.31 | −0.47 | 1 | ||||||
| Distance from soil to longest lateral root insertion on tap root (cm) | 0.06 | 0.03 | 0.00 | 0.02 | 0.24 | 0.03 | −0.07 | 0.06 | 0.07 | −0.05 | 0.07 | 0.16 | −0.09 | −0.04 | 0.06 | 1 | |||||
| Root angle 1 (°) | 0.18 | 0.12 | 0.11 | 0.15 | 0.00 | 0.09 | 0.09 | 0.25 | 0.27 | 0.18 | 0.28 | 0.44 | −0.04 | 0.08 | 0.25 | 0.84 *** | 1 | ||||
| Root angle 2 (°) | 0.39 | 0.29 | 0.30 | 0.36 | −0.31 | 0.13 | 0.31 | 0.49 | 0.53 | 0.50 | 0.54 | 0.74 ** | −0.03 | 0.14 | 0.49 | 0.22 | 0.68 * | 1 | |||
| Root lateral expansion rate (cm day−1) | 0.36 | 0.21 | 0.33 | 0.37 | −0.45 | −0.07 | 0.44 | 0.32 | 0.34 | 0.45 | 0.36 | 0.22 | −0.20 | −0.26 | 0.32 | −0.61 * | −0.57 | −0.23 | 1 | ||
| Root penetration rate in depth (cm day−1) | 0.28 | −0.19 | 0.73 ** | 0.68 * | −0.73 ** | −0.43 | 0.39 | 0.42 | 0.39 | 0.38 | 0.32 | 0.43 | 0.10 | 0.03 | 0.42 | 0.17 | 0.41 | 0.47 | 0.22 | 1 | |
| Thousand Seed Weight (g) | 0.34 | −0.18 | 0.88 *** | 0.83 *** | −0.23 | −0.84 *** | 0.59 * | 0.53 | 0.42 | 0.47 | 0.28 | 0.34 | 0.36 | 0.03 | 0.53 | −0.01 | −0.08 | −0.02 | 0.29 | 0.51 | 1 |
| Root Dry Matter (mg) | Root: Shoot Ratio | Shoot Dry Matter (mg) | Total Dry Matter (mg) | Plant Water Potential (MPa) | Average Root Diameter (mm) | Length of Roots with Diameter ≤ 0.2 mm (cm) | Root Length Density (cm cm−3) | Root Surface Area (cm2) | Root Tips Number | Root Volume (cm3) | Shoot Length (cm) | Specific Root Length (cm mg−1) | Specific Root Surface (cm2 mg−1) | Total Root Length (cm) | Distance from Soil to Longest Lateral Root Insertion on tap root (cm) | Root Angle 1 (°) | Root Angle 2 (°) | Root Lateral Expansion Rate (cm day−1) | Root Penetration Rate in Depth (cm day−1) | Thousand Seed Weight (g) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root dry matter (mg) | 1 | ||||||||||||||||||||
| Root: shoot ratio | 0.63 * | 1 | |||||||||||||||||||
| Shoot dry matter (mg) | 0.28 | −0.55 | 1 | ||||||||||||||||||
| Total dry matter (mg) | 0.45 | −0.39 | 0.98 *** | 1 | |||||||||||||||||
| Plant water potential (MPa) | −0.01 | −0.22 | 0.24 | 0.23 | 1 | ||||||||||||||||
| Average root diameter (mm) | −0.21 | 0.39 | −0.69 * | −0.69 * | −0.66 * | 1 | |||||||||||||||
| Length of roots with diameter ≤ 0.2 mm (cm) | 0.31 | −0.28 | 0.71 ** | 0.72 ** | 0.64 * | −0.88 *** | 1 | ||||||||||||||
| Root length density (cm cm−3) | 0.78 ** | 0.09 | 0.71 ** | 0.80 ** | 0.39 | −0.69 * | 0.74 ** | 1 | |||||||||||||
| Root surface area (cm2) | 0.90 *** | 0.29 | 0.60 * | 0.72 ** | 0.24 | −0.49 | 0.58 * | 0.97 *** | 1 | ||||||||||||
| Root tips number | 0.85 *** | 0.25 | 0.59 * | 0.71 ** | 0.35 | −0.64 * | 0.70 * | 0.94 *** | 0.94 *** | 1 | |||||||||||
| Root volume (cm3) | 0.95 *** | 0.53 | 0.37 | 0.52 | −0.01 | −0.12 | 0.30 | 0.79 ** | 0.92 *** | 0.80 ** | 1 | ||||||||||
| Shoot length (cm) | 0.74 ** | 0.54 | 0.09 | 0.23 | 0.10 | −0.25 | 0.26 | 0.65 * | 0.69 ** | 0.63 * | 0.66 * | 1 | |||||||||
| Specific root length (cm mg−1) | 0.05 | −0.62 * | 0.82 ** | 0.77 ** | 0.60 * | −0.87 *** | 0.87 ** | 0.64 * | 0.45 | 0.50 | 0.11 | 0.10 | 1 | ||||||||
| Specific root surface (cm2 mg−1) | −0.06 | −0.70 * | 0.80 ** | 0.73 ** | 0.52 | −0.69 * | 0.71 ** | 0.55 | 0.38 | 0.36 | 0.08 | 0.00 | 0.95 *** | 1 | |||||||
| Total root length (cm) | 0.78 ** | 0.09 | 0.71 ** | 0.80 ** | 0.39 | −0.69 * | 0.74 ** | 1.00 | 0.97 *** | 0.94 *** | 0.79 ** | 0.65 * | 0.64 * | 0.55 | 1 | ||||||
| Distance from soil to longest lateral root insertion on tap root (cm) | 0.60 * | 0.26 | 0.34 | 0.43 | 0.26 | −0.35 | 0.53 | 0.58 * | 0.60 * | 0.68 ** | 0.57 | 0.16 | 0.22 | 0.09 | 0.58 * | 1 | |||||
| Root angle 1 (°) | 0.37 | 0.58 * | −0.32 | −0.23 | −0.38 | 0.39 | −0.37 | −0.06 | 0.09 | 0.06 | 0.31 | 0.00 | −0.60 * | −0.64 * | −0.06 | 0.55 | 1 | ||||
| Root angle 2 (°) | 0.17 | 0.61 * | −0.62 * | −0.55 | −0.64 * | 0.70 * | −0.84 *** | −0.32 | −0.13 | −0.26 | 0.13 | 0.19 | −0.80 ** | −0.71 * | −0.32 | −0.29 | 0.53 | 1 | |||
| Root lateral expansion rate (cm day−1) | −0.27 | −0.61* | 0.47 | 0.39 | 0.27 | −0.20 | 0.34 | 0.11 | 0.02 | 0.03 | −0.09 | −0.20 | 0.56 | 0.69 * | 0.11 | −0.27 | −0.81 ** | −0.57 | 1 | ||
| Root penetration rate in depth (cm day−1) | 0.65 * | 0.36 | 0.26 | 0.36 | 0.02 | −0.19 | 0.23 | 0.64 * | 0.69 * | 0.50 | 0.68 * | 0.53 | 0.14 | 0.11 | 0.64 * | 0.53 | 0.48 | 0.17 | −0.49 | 1 | |
| Thousand Seed Weight (g) | 0.20 | −0.59 * | 0.97 *** | 0.94 *** | 0.31 | −0.69 * | 0.76 ** | 0.63 * | 0.51 | 0.52 | 0.29 | −0.03 | 0.83 *** | 0.80 ** | 0.63 * | 0.35 | -0.36 | −0.74 ** | 0.52 | 0.19 | 1 |
References
- European Commission. Report Form the Commission to the Council and the European Parliament on the Development of Plant Proteins in the European Union; European Commission: Brussels, Belgium, 2018; pp. 1–15. [Google Scholar]
- Labalette, F.; Bourrel, C.; Jouffret, P.; Lecomte, V.; Quinsac, A.; Ledoux, S. Panorama et Futur de La Filière Du Soja Français. Oléagineux Corps Gras Lipides 2010, 17, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Specht, J.E.; Hume, D.J.; Kumudini, S.V. Soybean Yield Potential-A Genetic and Physiological Perspective Yield ImProvement in Retrospect. Crop Sci. 1999, 39, 1560–1570. [Google Scholar] [CrossRef]
- Meckel, L.; Egli, D.B.; Phillips, R.E.; Radcliffe, D.; Leggett, J.E. Effect of Moisture Stress on Seed Growth in Soybeans1. Agron. J. 1984, 76, 647–650. [Google Scholar] [CrossRef]
- Maury, P.; Andrianasolo, F.N.; Alric, F.; Berger, M.; Beugniet, G.; Chambert, C.; Champolivier, L.; Doumenc, A.; Estragnat, A.; Gras, A.; et al. Le Semis Très Précoce: Une Stratégie Agronomique Pour Améliorer Les Performances Du Soja En France? OCL Oilseeds Fats 2015, 22. [Google Scholar] [CrossRef]
- Blum, A. Effective Use of Water (EUW) and Not Water-Use Efficiency (WUE) Is the Target of Crop Yield Improvement under Drought Stress. Field Crop. Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K. Are Differences in Root Growth of Nitrogen Catch Crops Important for Their Ability to Reduce Soil Nitrate-N Content, and How Can This Be Measured? Plant Soil 2001, 230, 185–195. [Google Scholar] [CrossRef]
- Hunt, J.R.; Lilley, J.M.; Trevaskis, B.; Flohr, B.M.; Peake, A.; Fletcher, A.; Zwart, A.B.; Gobbett, D.; Kirkegaard, J.A. Early Sowing Systems Can Boost Australian Wheat Yields despite Recent Climate Change. Nat. Clim. Chang. 2019, 9, 244–247. [Google Scholar] [CrossRef]
- Lynch, J. Root Architecture and Plant Productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.C.; Wright, G.C.; Siddique, K.H.M. Adaptation of Grain Legumes (Pulses) to Water-Limited Environments. Adv. Agron. 2001, 71, 193–231. [Google Scholar]
- Battisti, R.; Sentelhas, P.C. Improvement of Soybean Resilience to Drought through Deep Root System in Brazil. Agron. J. 2017, 109, 1612–1622. [Google Scholar] [CrossRef]
- Han, E.; Dresbøll, D.B.; Thorup-Kristensen, K. Core-Labelling Technique (CLT): A Novel Combination of the Ingrowth-Core Method and Tracer Technique for Deep Root Study. Plant Methods 2020, 16, 84. [Google Scholar] [CrossRef] [PubMed]
- Svane, S.F.; Jensen, C.S.; Thorup-Kristensen, K. Construction of a Large-Scale Semi-Field Facility to Study Genotypic Differences in Deep Root Growth and Resources Acquisition. Plant Methods 2019, 15, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Ingram, P.A.; Benfey, P.N.; Elich, T. From Lab to Field, New Approaches to Phenotyping Root System Architecture. Curr. Opin. Plant Biol. 2011, 14, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Boiffin, J.; Durr, C.; Fleury, A.; Maillet, I.; Marin-Laflèche, A. Analysis of the Variability of Sugar Beet (Beta Vulgaris L) Growth during the Early Stages. I. Influence of Various Conditions on Crop Establishment. Agronomie 2007, 12, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Fayaud, B.; Coste, F.; Corre-Hellou, G.; Gardarin, A.; Dürr, C. Modelling Early Growth under Different Sowing Conditions: A Tool to Predict Variations in Intercrop Early Stages. Eur. J. Agron. 2014, 52, 180–190. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Constantin, J.; Schoving, C.; Maury, P.; Debaeke, P.; Aubertot, J.N.; Dürr, C. Analysis of Soybean Germination, Emergence, and Prediction of a Possible Northward Establishment of the Crop under Climate Change. Eur. J. Agron. 2020, 113. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Aubertot, J.-N.; Champolivier, L.; Debaeke, P.; Maury, P. Combining Experimental and Modeling Approaches to Understand Genotype x Sowing Date x Environment Interaction Effects on Emergence Rates and Grain Yield of Soybean. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Andersen, M.K.; Hauggaard-Nielsen, H.; Weiner, J.; Jensen, E.S. Competitive Dynamics in Two- and Three-Component Intercrops. J. Appl. Ecol. 2007, 44, 1556. [Google Scholar] [CrossRef]
- Dayoub, E.; Naudin, C.; Piva, G.; Shirtliffe, S.J.; Fustec, J.; Corre-Hellou, G. Traits Affecting Early Season Nitrogen Uptake in Nine Legume Species. Heliyon 2017, 3, e00244. [Google Scholar] [CrossRef] [Green Version]
- Manavalan, L.P.; Guttikonda, S.K.; Phan Tran, L.S.; Nguyen, H.T. Physiological and Molecular Approaches to Improve Drought Resistance in Soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [Green Version]
- Thu, N.B.A.; Nguyen, Q.T.; Hoang, X.L.T.; Thao, N.P.; Tran, L.-S.P. Evaluation of Drought Tolerance of the Vietnamese Soybean Cultivars Provides Potential Resources for Soybean Production and Genetic Engineering. Biomed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirasawa, T.; Tanaka, K.; Miyamoto, D.; Takei, M.; Ishihara, K. Effects of Pre-Flowering Soil Moisture Deficits on Dry Matter Production and Ecophysiological Characteristics in Soybean Plants under Drought Conditions during Grain Filling. Jpn. J. Crop Sci. 1994, 63, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Carton, N.; Naudin, C.; Piva, G.; Baccar, R.; Corre-Hellou, G. Differences for Traits Associated with Early N Acquisition in a Grain Legume and Early Complementarity in Grain Legume-Triticale Mixtures. AoB Plants 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.L.; Dunbabin, V.M.; Diggle, A.J.; Siddique, K.H.M.; Rengel, Z. Assessing Variability in Root Traits of Wild Lupinus Angustifolius Germplasm: Basis for Modelling Root System Structure. Plant Soil 2012, 354, 141–155. [Google Scholar] [CrossRef]
- McPhee, K. Variation for Seedling Root Architecture in the Core Collection of Pea Germplasm. Crop Sci. 2005, 45, 1758–1763. [Google Scholar] [CrossRef]
- Lynch, J.; van Beem, J.J. Growth and Architecture of Seedling Roots of Common Bean Genotypes. Crop Sci. 1993, 33, 1253–1257. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Guttikonda, S.K.; Nguyen, V.T.; Shannon, J.G.; Nguyen, H.T. Evaluation of Diverse Soybean Germplasm for Root Growth and Architecture. Plant Soil 2010, 330, 503–514. [Google Scholar] [CrossRef]
- Matsuo, N.; Takahashi, M.; Fukami, K.; Tasaka, K.; Tsuchiya, S. Root Growth of Two Soybean [ Glycine Max (L.) Merr.] Cultivars Grown under Different Groundwater Level Conditions. Plant Prod. Sci. 2013, 16, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Boote, K.J. Improving Soybean Cultivars to Climate Change an Variability—Chaptitre 17; John Willey and Sons: Chichester, West Sussex, UK, 2011. [Google Scholar]
- Martre, P.; Quilot-Turion, B.; Luquet, D.; Memmah, M.-M.O.-S.; Chenu, K.; Debaeke, P. Chapter 14—Model-Assisted Phenotyping and Ideotype Design; Sadras, V.O., Calderini, D.F.B.T.-C.P., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 349–373. [Google Scholar]
- Lynch, J.P. Root Phenes for Enhanced Soil Exploration and Phosphorus Acquisition: Tools for Future Crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Brown, K.M. Topsoil Foraging—An Architectural Adaptation of Plants to Low Phosphorus Availability. Plant Soil 2001, 237, 225–237. [Google Scholar] [CrossRef]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; et al. Plant and Microbial Strategies to Improve the Phosphorus Efficiency of Agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Wang, X.; Yan, X.; Liao, H. Genetic Improvement for Phosphorus Efficiency in Soybean: A Radical Approach. Ann. Bot. 2010, 106, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Schoving, C.; Stöckle, C.O.; Colombet, C.; Champolivier, L.; Debaeke, P.; Maury, P. Combining Simple Phenotyping and Photothermal Algorithm for the Prediction of Soybean Phenology: Application to a Range of Common Cultivars Grown in Europe. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhm, W.; Maduakor, H.; Taylor, H.M. Comparison of Five Methods for Characterizing Soybean Rooting Density and Development1. Agron. J. 1977, 69, 415. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E. Stages of Soybean Development; Special Report 80; Iowa State University, Agricultural and Home Economics Experiment Station: Ames, IA, USA, 1977. [Google Scholar]
- Terres Inovia. myVar-Web Links. Available online: http://www.myvar.fr/ (accessed on 16 December 2020).
- Ao, J.; Fu, J.; Tian, J.; Yan, X.; Liao, H. Genetic Variability for Root Morph-Architecture Traits and Root Growth Dynamics as Related to Phosphorus Efficiency in Soybean. Funct. Plant Biol. 2010, 37, 304–312. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. New Roots for Agriculture: Exploiting the Root Phenome. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1598–1604. [Google Scholar] [CrossRef]
- ImageJ Free Software. Available online: https://imagej.nih.gov/ij/ (accessed on 18 December 2020).
- Steduto, P.; Hsiao, T.C.; Raes, D.; Fereres, E. AquaCrop—The FAO Crop Model to Simulate Yield Response to Water: I. Concepts and Underlying Principles. Agron. J. 2009, 101, 426–437. [Google Scholar] [CrossRef] [Green Version]
- R Core. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Lê, S.; Josse, J.; Rennes, A.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. JSS J. Stat. Softw. 2008, 25, 1–18. [Google Scholar]
- Fisher, R.A.; Yates, F. Statistical Tables for Biological, Agricultural Aad Medical Research; Oliver and Boyd: London, UK, 1938; p. 90. [Google Scholar]
- Fried, H.G.; Narayanan, S.; Fallen, B. Characterization of a Soybean (Glycine Max, L. Merr.) Germplasm Collection for Root Traits. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Torrion, J.A.; Setiyono, T.D.; Cassman, K.G.; Ferguson, R.B.; Irmak, S.; Specht, J.E. Soybean Root Development Relative to Vegetative and Reproductive Phenology. Agron. J. 2012, 104, 1702–1709. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, T.C.; Taylor, H.M.; Shibles, R.M. Taproot-Elongation Rates of Soybean Cultivars in the Glasshouse and Their Relation to Field Rooting Depth1. Crop Sci. 1984, 24, 916–920. [Google Scholar] [CrossRef]
- Fenta, B.; Barlow, K.; Burridge, J.; Lynch, J.; Beebe, S.; Foyer, C.; Kunert, K. Field Phenotyping of Soybean Roots for Drought Stress Tolerance. Agronomy 2014, 4, 418–435. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J. Characterization of Root Architecture in an Applied Core Collection for Phosphorus Efficiency of Soybean Germplasm. Chinese Sci. Bull. 2004, 49, 1611–1620. [Google Scholar] [CrossRef]
- Christopher, J.; Christopher, M.; Jennings, R.; Jones, S.; Fletcher, S.; Borrell, A.; Manschadi, A.M.; Jordan, D.; Mace, E.; Hammer, G. QTL for Root Angle and Number in a Population Developed from Bread Wheats (Triticum Aestivum) with Contrasting Adaptation to Water-Limited Environments. Theor. Appl. Genet. 2013, 126, 1563–1574. [Google Scholar] [CrossRef] [PubMed]
- Mace, E.S.; Singh, V.; van Oosterom, E.J.; Hammer, G.L.; Hunt, C.H.; Jordan, D.R. QTL for Nodal Root Angle in Sorghum (Sorghum Bicolor, L. Moench) Co-Locate with QTL for Traits Associated with Drought Adaptation. Theor. Appl. Genet. 2012, 124, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of Root System Architecture by DEEPER ROOTING 1 Increases Rice Yield under Drought Conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef]
- Joshi, D.C.; Singh, V.; Hunt, C.; Mace, E.; van Oosterom, E.; Sulman, R.; Jordan, D.; Hammer, G. Development of a Phenotyping Platform for High Throughput Screening of Nodal Root Angle in Sorghum. Plant Methods 2017, 13, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; van Oosterom, E.J.; Jordan, D.R.; Hunt, C.H.; Hammer, G.L. Genetic Variability and Control of Nodal Root Angle in Sorghum. Crop Sci. 2011, 51. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Hammer, G.L.; Christopher, J.T.; DeVoil, P. Genotypic Variation in Seedling Root Architectural Traits and Implications for Drought Adaptation in Wheat (Triticum Aestivum, L.). Plant Soil 2008, 303, 115–129. [Google Scholar] [CrossRef]
- Nakamoto, T.; Oyanagi, A. The Direction of Growth of Seminal Roots of Triticum Aestivum, L. and Experimental Modification Thereof. Ann. Bot. 1994, 73, 363–367. [Google Scholar] [CrossRef]
- Singh, V.; van Oosterom, E.J.; Jordan, D.R.; Messina, C.D.; Cooper, M.; Hammer, G.L. Morphological and Architectural Development of Root Systems in Sorghum and Maize. Plant Soil 2010, 333, 287–299. [Google Scholar] [CrossRef]
- Xiong, R.; Liu, S.; Considine, M.J.; Siddique, K.H.M.; Lam, H.M.; Chen, Y. Root System Architecture, Physiological and Transcriptional Traits of Soybean (Glycine Max, L.) in Response to Water Deficit: A Review. Physiol. Plant. 2020, 1–14. [Google Scholar] [CrossRef]
- Guerrero-Campo, J.; Fitter, A.H. Relationships between Root Characteristics and Seed Size in Two Contrasting Floras. Acta Oecologica 2001, 22. [Google Scholar] [CrossRef]
- Strock, C.F.; Burridge, J.; Massas, A.S.F.; Beaver, J.; Beebe, S.; Camilo, S.A.; Fourie, D.; Jochua, C.; Miguel, M.; Miklas, P.N.; et al. Seedling Root Architecture and Its Relationship with Seed Yield across Diverse Environments in Phaseolus Vulgaris. F. Crop. Res. 2019, 237, 53–64. [Google Scholar] [CrossRef]
- Koester, R.P.; Nohl, B.M.; Diers, B.W.; Ainsworth, E.A. Has Photosynthetic Capacity Increased with 80years of Soybean Breeding? An Examination of Historical Soybean Cultivars. Plant. Cell Environ. 2016, 39. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, Cheap and Deep: An Ideotype to Optimize Water and N Acquisition by Maize Root Systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Lersten, N.R.; Carlson, J.B. Vegetative Morphology. In Soybeans: Improvement, Production, and Uses; Agronomy Monographs Series; American Society of Agronomy, Inc.: Madison, WI, USA, 2004; pp. 15–57. [Google Scholar]
- Brisson, N.; Gary, C.; Justes, E.; Roche, R.; Mary, B.; Ripoche, D.; Zimmer, D.; Sierra, J.; Bertuzzi, P.; Burger, P.; et al. An Overview of the Crop Model STICS. Eur. J. Agron. 2003, 18. [Google Scholar] [CrossRef]






| Cultivars 1 | Maturity Group | Breeding Company | Year and Country of Registration | Thousand Seed Weight (g) 2 | Shoot Length Classification | Growth Type |
|---|---|---|---|---|---|---|
| Blancas | II | Caussade Semences | 2007-Italy | 177.7 | Medium | Indeterminate |
| Ecudor | II | Euralis Semences | 2006-France | 213.5 | Tall | Indeterminate |
| ES Mentor | 0 | Euralis Semences | 2009-France | 204.6 | Short | Semi-determinate |
| ES Pallador | I | Euralis Semences | 2015-France | 174.7 | Medium | Indeterminate |
| Isidor | I | Euralis Semences | 2004-France | 248.3 | Short | Semi-determinate |
| Klaxon | 000(0) | RAGT Semences | 2005-France | 167.6 | Short | Indeterminate |
| RGT Shouna | 0 | RAGT Semences | 2014-France | 170.4 | Tall | Indeterminate |
| Santana | I/II | RAGT Semences | 2007-France | 208.6 | Short | Indeterminate |
| Sigalia | 0 | RAGT Semences | 2008-France | 213.8 | Tall | Semi-determinate to indeterminate |
| Sultana | 0 | RAGT Semences | 2009-France | 214.5 | Short | Semi-determinate |
| Cultivars | Shoot Length (cm plant−1) | Shoot Dry Matter (mg plant−1) | Root Dry Matter (mg plant−1) | Root:Shoot Ratio | Total Dry Matter (mg plant−1) | Total Root Length (cm plant−1) | Root Surface Area (cm2) | Root Volume (cm3) | Root Tips Number | Average Root Diameter (mm) | Root Angle 1 (°) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Well-watered conditions | |||||||||||
| Blancas | 7.20 ± 0.63 a | 128.18 ± 4.37 c | 22.54 ± 3.31 abc | 0.18 ± 0.03 ab | 150.71 ± 4.30 c | 136.05 ± 18.95 bc | 21.87 ± 2.43 bc | 0.29 ± 0.03 ab | 119.88 ± 17.03 bc | 0.53 ± 0.04 | 44.31 ± 3.83 a |
| Ecudor | 8.71 ± 0.76 a | 138.11 ± 16.37 bc | 20.08 ± 4.23 bc | 0.16 ± 0.03 ab | 158.19 ± 16.43 bc | 128.02 ± 18.34 bc | 21.89 ± 3.49 bc | 0.30 ± 0.06 ab | 118.50 ± 19.17 bc | 0.54 ± 0.03 | 42.91 ± 2.60 a |
| ES Mentor | 9.35 ± 0.91 a | 152.59 ± 4.03 abc | 29.96 ± 4.58 ab | 0.20 ± 0.03 ab | 182.55 ± 7.06 abc | 172.21 ± 26.39 abc | 28.71 ± 4.14 abc | 0.39 ± 0.06 ab | 136.75 ± 21.49 abc | 0.54 ± 0.03 | 54.47 ± 4.28 a |
| ES Pallador | 6.98 ± 1.09 a | 122.23 ± 10.32 c | 13.78 ± 2.86 c | 0.12 ± 0.03 b | 136.00 ± 10.02 c | 93.93 ± 15.89 c | 15.79 ± 3.05 c | 0.22 ± 0.05 b | 86.00 ± 15.02 c | 0.54 ± 0.03 | 57.06 ± 4.11 a |
| Isidor | 10.92 ± 1.08 a | 184.72 ± 4.74 a | 36.68 ± 3.63 a | 0.20 ± 0.02 ab | 221.40 ± 6.07 a | 247.57 ± 27.56 a | 38.14 ± 3.82 a | 0.47 ± 0.05 a | 204.33 ± 18.43 a | 0.50 ± 0.02 | 53.45 ± 3.06 a |
| Klaxon | 9.32 ± 0.56 a | 136.67 ± 6.72 bc | 25.04 ± 2.68 abc | 0.18 ± 0.02 ab | 161.71 ± 8.00 bc | 149.24 ± 11.64 abc | 26.05 ± 2.01 abc | 0.37 ± 0.04 ab | 134.33 ± 12.66 abc | 0.56 ± 0.03 | 50.12 ± 3.79 a |
| RGT Shouna | 11.10 ± 0.54 a | 133.46 ± 7.25 ab | 31.03 ± 4.24 ab | 0.24 ± 0.03 a | 164.49 ± 8.79 ab | 189.46 ± 29.74 ab | 32.44 ± 4.55 ab | 0.45 ± 0.06 a | 177.75 ± 28.23 ab | 0.56 ± 0.03 | 57.42 ± 2.83 a |
| Santana | 7.81 ± 1.51 a | 156.21 ± 4.57 abc | 22.06 ± 2.52 abc | 0.14 ± 0.02 ab | 178.28 ± 5.13 abc | 141.65 ± 12.72 abc | 22.47 ± 1.88 bc | 0.29 ± 0.03 ab | 137.88 ± 17.12 abc | 0.51 ± 0.03 | 44.83 ± 2.63 a |
| Sigalia | 9.41 ± 0.94 a | 169.02 ± 5.01 bc | 22.40 ± 2.17 abc | 0.14 ± 0.02 ab | 191.42 ± 4.35 bc | 158.63 ± 13.86 abc | 25.82 ± 2.17 abc | 0.34 ± 0.03 ab | 159.22 ± 11.20 abc | 0.52 ± 0.01 | 51.37 ± 3.87 a |
| Sultana | 9.40 ± 0.90 a | 162.31 ± 4.42 abc | 19.36 ± 3.02 bc | 0.12 ± 0.02 b | 181.68 ± 4.10 abc | 144.24 ± 21.85 bc | 22.90 ± 3.43 bc | 0.29 ± 0.04 ab | 123.75 ± 18.31 abc | 0.51 ± 0.02 | 56.46 ± 3.12 a |
| Significance | * | *** | *** | * | *** | ** | *** | ** | ** | NS | * |
| Water-stressed conditions | |||||||||||
| Blancas | 7.68 ± 0.84 a | 115.83 ± 5.13 bc | 27.26 ± 2.46 a | 0.24 ± 0.02 a | 143.09 ± 5.44 c | 134.34 ± 14.98 b | 25.30 ± 2.85 ab | 0.38 ± 0.05 ab | 134.63 ± 17.00 abc | 0.60 ± 0.02 a | 66.58 ± 4.49 |
| Ecudor | 6.63 ± 0.97 a | 143.41 ± 8.22 abc | 24.01 ± 5.04 a | 0.18 ± 0.05 a | 167.43 ± 4.81 bc | 125.97 ± 22.77 b | 22.68 ± 4.20 b | 0.34 ± 0.07 ab | 137.75 ± 18.87 abc | 0.58 ± 0.05 ab | 62.53 ± 4.41 |
| ES Mentor | 8.29 ± 0.77 a | 149.75 ± 3.61 abc | 31.90 ± 3.62 a | 0.22 ± 0.03 a | 181.65 ± 3.10 b | 188.38 ± 18.91 ab | 31.27 ± 3.16 ab | 0.42 ± 0.05 ab | 214.75 ± 25.24 ab | 0.53 ± 0.01 ab | 67.01 ± 3.48 |
| ES Pallador | 7.14 ± 1.03 a | 122.15 ± 4.38 bc | 20.04 ± 3.13 a | 0.17 ± 0.03 a | 142.19 ± 3.79 c | 106.17 ± 15.70 b | 20.02 ± 2.88 b | 0.30 ± 0.04 ab | 103.75 ± 15.00 c | 0.61 ± 0.03 a | 63.15 ± 3.00 |
| Isidor | 10.14 ± 1.06 a | 186.99 ± 7.82 a | 32.26 ± 3.45 a | 0.18 ± 0.02 a | 219.25 ± 7.58 a | 241.94 ± 20.22 a | 37.15 ± 3.01 a | 0.46 ± 0.04 a | 217.63 ± 14.75 a | 0.49 ± 0.02 b | 61.07 ± 3.96 |
| Klaxon | 8.30 ± 0.64 a | 116.56 ± 4.42 c | 27.93 ± 2.60 a | 0.24 ± 0.02 a | 144.49 ± 6.17c | 137.19 ± 18.29 b | 25.08 ± 2.95 ab | 0.37 ± 0.04 ab | 140.78 ± 16.55 abc | 0.60 ± 0.02 ab | 71.39 ± 2.98 |
| RGT Shouna | 10.05 ± 0.85 a | 118.56 ± 4.00 bc | 25.91 ± 3.20 a | 0.22 ± 0.03 a | 144.48 ± 5.10 c | 135.55 ± 17.55 b | 23.33 ± 3.00 b | 0.32 ± 0.04 ab | 144.38 ± 16.30 abc | 0.55 ± 0.02 ab | 60.30 ± 5.06 |
| Santana | 6.05 ± 1.00 a | 146.25 ± 11.89 abc | 18.88 ± 3.21 a | 0.13 ± 0.03 a | 165.13 ± 12.82 bc | 130.00 ± 23.28 b | 20.63 ± 3.11 b | 0.27 ± 0.04 b | 124.25 ± 19.74 c | 0.53 ± 0.03 ab | 57.72 ± 2.98 |
| Sigalia | 6.91 ± 1.12 a | 157.36 ± 5.64 ab | 24.57 ± 3.34 a | 0.16 ± 0.03 a | 181.92 ± 5.05 b | 124.63 ± 13.85 b | 23.14 ± 2.59 b | 0.34 ± 0.04 ab | 122.44 ± 07.83 bc | 0.59 ± 0.02 ab | 65.03 ± 2.67 |
| Sultana | 6.61 ± 0.80 a | 149.54 ± 6.68 abc | 21.26 ± 2.87 a | 0.15 ± 0.02 a | 170.80 ± 4.90 b | 136.51 ± 16.95 b | 21.64 ± 2.69 b | 0.28 ± 0.04 ab | 136.75 ± 15.14 abc | 0.51 ± 0.02 ab | 65.66 ± 3.16 |
| Significance | * | *** | * | * | *** | *** | ** | * | *** | ** | NS |
| Well-Watered Conditions | Water-Stressed Conditions | Effect | ||||||
|---|---|---|---|---|---|---|---|---|
| Traits | Variable | Mean | Cultivar Range | Mean | Cultivar Range | C | WT | C*WT |
| Morpho-physiological traits (shoot or root) | Shoot length (cm) | 9.02 | 6.98–10.92 | 7.78 | 6.05–10.14 | *** | ** | NS |
| Shoot dry matter (mg) | 148.35 | 122.23–184.72 | 140.64 | 115.83–186.99 | *** | * | NS | |
| Root dry matter (mg) | 24.29 | 13.78–36.68 | 25.4 | 18.88–32.26 | *** | NS | NS | |
| Root: shoot ratio | 0.17 | 0.12–0.24 | 0.19 | 0.13–0.24 | *** | NS | NS | |
| Total dry matter (mg) | 172.64 | 136–221.4 | 166.04 | 142.19–219.25 | *** | * | NS | |
| Average root diameter (mm) | 0.53 | 0.50–0.56 | 0.56 | 0.49–0.61 | * | * | NS | |
| Specific root surface (cm2 mg−1) | 1.11 | 0.97–1.21 | 1.01 | 0.89–1.20 | NS | ** | NS | |
| Specific root length (cm mg−1) | 6.86 | 5.91–7.52 | 5.97 | 4.89–7.93 | NS | ** | NS | |
| Total root length (cm) | 156.1 | 93.93–247.57 | 146.07 | 106.17–241.94 | *** | NS | NS | |
| Root surface area (cm2) | 25.61 | 15.79–38.14 | 25.02 | 20.02–37.15 | *** | NS | NS | |
| Root volume (cm3) | 0.34 | 0.22–0.47 | 0.35 | 0.27–0.46 | *** | NS | NS | |
| Root tips number | 139.84 | 86–204.33 | 147.71 | 103.75–217.63 | *** | NS | NS | |
| Root length density (cm cm−3) | 0.1 | 0.06–0.16 | 0.1 | 0.07–0.16 | *** | NS | NS | |
| Length of roots with diameter ≤ 0.2 mm (cm) | 27.16 | 14.37–49.75 | 21.35 | 12.09–35.30 | *** | ** | NS | |
| Length of roots with diameter between 0.2 and 0.4 mm (cm) | 55.37 | 37.49–95.87 | 54.95 | 33.04–116.40 | *** | NS | NS | |
| Length of roots with diameter between 0.4 and 0.6 mm (cm) | 32.39 | 12.38–45.01 | 27.21 | 10.72–41.80 | *** | * | NS | |
| Length of roots with diameter between 0.6 and 0.8 mm (cm) | 19.82 | 10.29–31.27 | 18 | 11.54–25.98 | * | NS | NS | |
| Length of roots with diameter between 0.8 and 1 mm (cm) | 12.52 | 7.68–18.09 | 14.35 | 10.37–22.02 | NS | NS | NS | |
| Length of roots with diameter between 1 and 2 mm (cm) | 7.83 | 5.77–10.91 | 9.46 | 6.90–14.32 | ** | * | NS | |
| Length of roots with diameter between 2 and 3 mm (cm) | 0.82 | 0.58–1.18 | 0.58 | 0.25–0.76 | NS | ** | NS | |
| Plant water potential (MPa) | −0.34 | −0.55–−0.18 | −0.41 | −0.65–−0.25 | NS | NS | NS | |
| Architectural traits (root) | Root lateral expansion rate (cm day−1) | 1.80 | 1.44–2.08 | 1.23 | 0.81–1.45 | NS | *** | NS |
| Root penetration rate in depth (cm day−1) | 3.55 | 2.91–4.04 | 4.09 | 3.66–4.84 | NS | ** | NS | |
| Root angle 1 (°) | 51.24 | 42.91–57.42 | 64.04 | 57.72–71.39 | NS | *** | NS | |
| Root angle 2 (°) | 39.67 | 32.04–46.74 | 45.63 | 40.31–51.90 | NS | ** | NS | |
| Distance from soil to longest lateral root insertion on taproot (cm) | 1.83 | 1.23–2.59 | 3.45 | 1.86–5.22 | NS | *** | NS | |
| Year | Sowing Date | Cultivar | CC0 (%) | CGC | Shoot Dry Matter at R1 Stage (g m−2) | Number of Days from Emergence to R1 Stage | Grain Yield (q ha−1) |
|---|---|---|---|---|---|---|---|
| 2013 | Conventional sowing | Isidor | 2.05 ± 0.08 b | 0.059 ± 0.001 b | 146.07 ± 5.91 b | 59.00 ± 0.00 c | 45.27 ± 1.41 ab |
| Santana | 1.11 ± 0.05 c | 0.065 ± 0.001 b | 119.19 ± 4.81 cde | 58.50 ± 0.22 c | 49.64 ± 1.32 a | ||
| Early sowing | Isidor | 4.18 ± 0.18 a | 0.036 ± 0.001 e | 140.68 ± 6.00 bc | 75.00 ± 0.93 a | 47.67 ± 1.36 ab | |
| Santana | 2.04 ± 0.09 b | 0.040 ± 0.001 de | 106.79 ± 4.45 de | 76.00 ± 0.77 a | 50.31 ± 0.59 a | ||
| 2014 | Conventional sowing | Isidor | 1.33 ± 0.22 c | 0.094 ± 0.005 a | 202.83 ± 18.54 a | 47.33 ± 1.20 d | 45.69 ± 1.49 ab |
| Santana | 0.51 ± 0.09 d | 0.104 ± 0.004 a | 136.38 ± 05.11 bcd | 46.67± 0.95 d | 50.92 ± 1.03 a | ||
| Early sowing | Isidor | 3.29 ± 0.45 a | 0.043 ± 0.005 d | 89.00 ± 2.43 e | 63.33 ± 0.33 b | 43.36 ± 0.58 b | |
| Santana | 1.59 ± 0.11 bc | 0.049 ± 0.001 c | 120.38 ± 8.31 bcde | 63.50 ± 0.22 b | 48.08 ± 1.26 ab | ||
| Effects | Cultivar | *** | *** | *** | NS | *** | |
| Year | *** | *** | NS | *** | NS | ||
| Sowing date | *** | *** | *** | *** | NS | ||
| Cultivar * Year | NS | NS | NS | NS | NS | ||
| Cultivar * Sowing date | *** | *** | ** | ** | NS | ||
| Year * Sowing date | NS | NS | *** | *** | * | ||
| Cultivar * Year * Sowing date | NS | NS | *** | *** | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dayoub, E.; Lamichhane, J.R.; Schoving, C.; Debaeke, P.; Maury, P. Early-Stage Phenotyping of Root Traits Provides Insights into the Drought Tolerance Level of Soybean Cultivars. Agronomy 2021, 11, 188. https://doi.org/10.3390/agronomy11010188
Dayoub E, Lamichhane JR, Schoving C, Debaeke P, Maury P. Early-Stage Phenotyping of Root Traits Provides Insights into the Drought Tolerance Level of Soybean Cultivars. Agronomy. 2021; 11(1):188. https://doi.org/10.3390/agronomy11010188
Chicago/Turabian StyleDayoub, Elana, Jay Ram Lamichhane, Céline Schoving, Philippe Debaeke, and Pierre Maury. 2021. "Early-Stage Phenotyping of Root Traits Provides Insights into the Drought Tolerance Level of Soybean Cultivars" Agronomy 11, no. 1: 188. https://doi.org/10.3390/agronomy11010188
APA StyleDayoub, E., Lamichhane, J. R., Schoving, C., Debaeke, P., & Maury, P. (2021). Early-Stage Phenotyping of Root Traits Provides Insights into the Drought Tolerance Level of Soybean Cultivars. Agronomy, 11(1), 188. https://doi.org/10.3390/agronomy11010188