In Vitro and Cryobiotechnology Approaches to Safeguard Lupinus rivularis Douglas ex Lindl., an Endangered Plant in Canada



Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Germination In Vitro and Multiple Shoot Induction
2.2. Shoot Multiplication In Vitro
2.3. In Vitro Rooting and Acclimatization
2.4. Seed Cryopreservation and Its Effect on Plant Development
2.5. Cryopreservation of Apical Shoot Tips
2.6. Statistical Analysis
3. Results
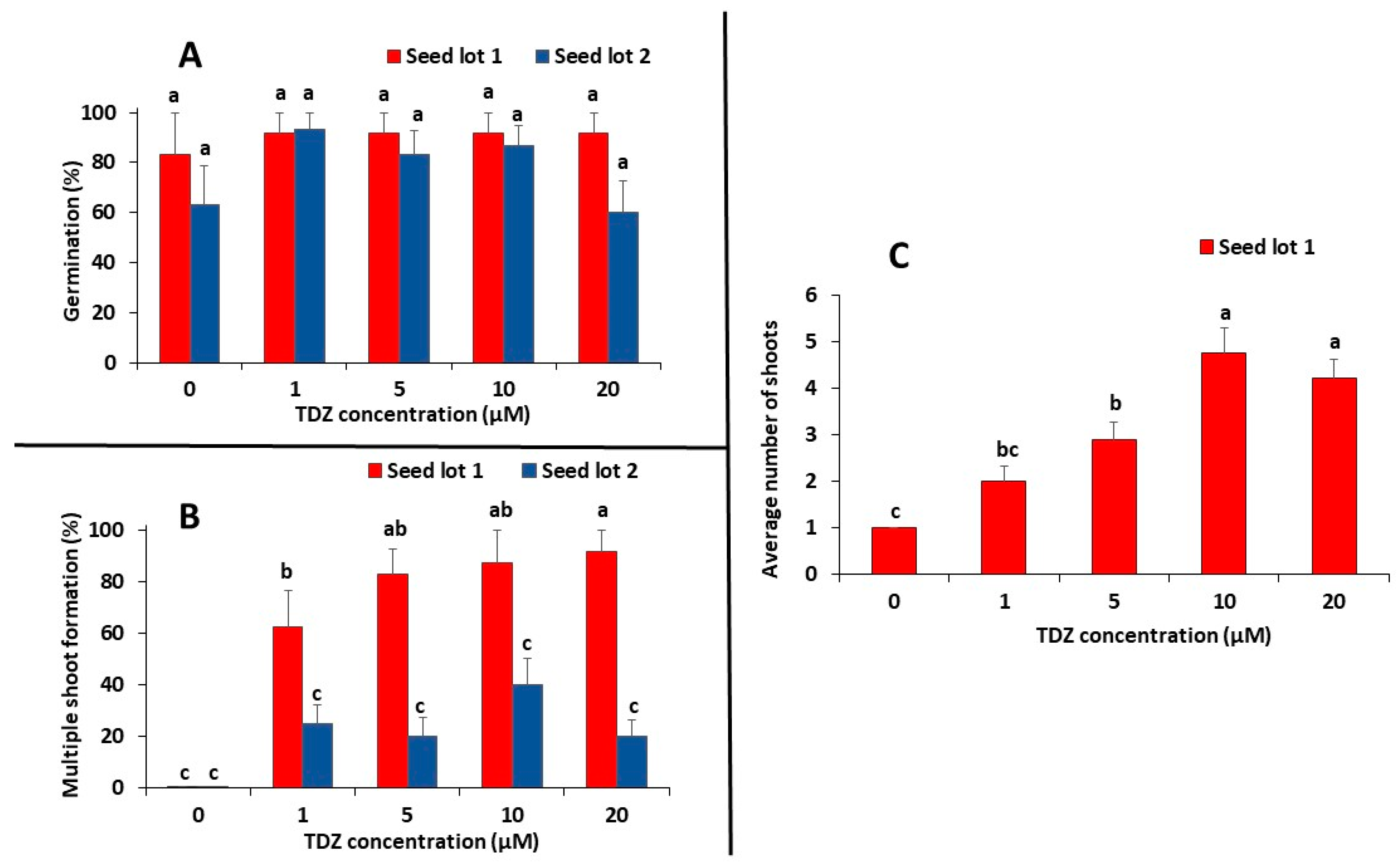
3.1. Seed Germination in Vitro and Multiple Shoot Induction
3.2. Shoot Multiplication In Vitro
3.3. In Vitro Rooting and Acclimatization
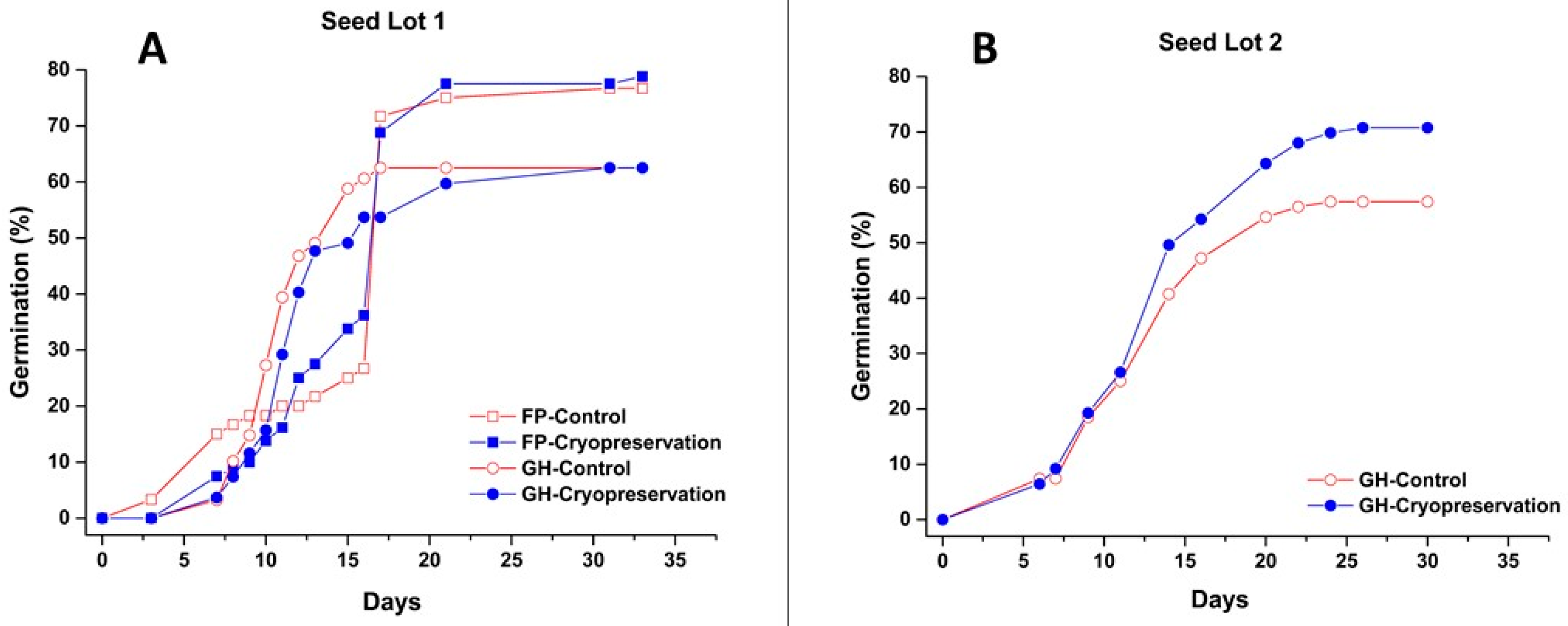
3.4. Seed Cryopreservation and Its Effect on Plant Development
3.5. Cryopreservation of Apical Shoot Tips
4. Discussion
4.1. Seed Germination, Shoot Multiplication and Plantlet Development
4.2. Seed Cryopreservation and Its Effect on Plant Development
4.3. Cryopreservation of Apical Shoot Tips
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riggins, R.; Sholars, T. Lupinus. In The Jepson Manual: Higher Plants of California; University of California Press: Berkeley, CA, USA, 1993; pp. 622–636. [Google Scholar]
- COSEWIC. COSEWIC Assessment and Status Report on the Streambank Lupine Lupinus rivularis in Canada; COSEWIC: Ottawa, ON, Canada, 2002. [Google Scholar]
- Streambank Lupine Recovery Team. Recovery Plan for Streambank Lupine (Lupinus rivularis) in British Columbia; Government of Canada: Victoria, BC, Canada, 2014.
- Environment and Climate Change Canada. In Recovery Strategy for the Streambank Lupine (Lupinus rivularis) in Canada; Environment and Climate Change Canada: Ottawa, ON, Canada, 2017.
- Halmagyi, A.; Coste, A.; Jarda, L.; Butiuc-Keul, A.; Holobiuc, I.; Cristea, V. A safeguard measure of endemic and endangered plant species: Cryostorage of Dianthus taxa. Biodivers. Conserv. 2020, 29, 3445–3460. [Google Scholar] [CrossRef]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.-C. Cryobiotechnologies: Tools for expanding long-term ex situ conservation to all plant species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- González-Benito, M.E.; Martin, C. In vitro preservation of Spanish biodiversity. In Vitr. Cell. Dev. Biol. Plant. 2010, 47, 46–54. [Google Scholar] [CrossRef]
- Pence, V. In Vitro methods and cryopreservation: Tools for endangered exceptional Sspecies preservation and restoration. Acta Hortic. 2014, 73–79. [Google Scholar] [CrossRef]
- Salama, A.; Shukla, M.R.; Popova, E.; Fisk, N.S.; Jones, M.P.; Saxena, P.K. In vitro propagation and reintroduction of golden paintbrush (Castilleja levisecta), a critically imperiled plant species. Can. J. Plant Sci. 2018, 98, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Sheikholeslami, B.; Shukla, M.; Turi, C.; Harpur, C.; Saxena, P.K. Saving threatened plant species: Reintroduction of Hill’s thistle (Cirsium hillii (Canby) Fernald) to its natural habitat. PLoS ONE 2020, 15, e0231741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, H.W. Cryopreservation of desiccation-tolerant seeds. Cryopreserv. Free. Protoc. Methods Mol. Biol. 2007, 368, 185–201. [Google Scholar] [CrossRef]
- Uchendu, E.E.; Shukla, M.R.; Reed, B.M.; Saxena, P.K. Melatonin enhances the recovery of cryopreserved shoot tips of American elm (Ulmus americana L.). J. Pineal Res. 2013, 55, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Pence, V.C.; Philpott, M.; Culley, T.M.; Plair, B.; Yorke, S.R.; Lindsey, K.; Vanhove, A.-C.; Ballesteros, D. Survival and genetic stability of shoot tips of Hedeoma todsenii R.S.Irving after long-term cryostorage. In Vitr. Cell. Dev. Biol. Plant. 2017, 53, 328–338. [Google Scholar] [CrossRef]
- Salama, A.; Popova, E.; Jones, M.P.; Shukla, M.R.; Fisk, N.S.; Saxena, P.K. Cryopreservation of the critically endangered golden paintbrush (Castilleja levisecta Greenm.): From nature to cryobank to nature. In Vitr. Cell. Dev. Biol. Plant. 2018, 54, 69–78. [Google Scholar] [CrossRef]
- Ciringer, T.; Martín, C.; Šajna, N.; Kaligarič, M.; Ambrožič-Dolinšek, J. Cryopreservation of an endangered Hladnikia pastinacifolia Rchb. by shoot tip encapsulation-dehydration and encapsulation-vitrification. In Vitr. Cell. Dev. Biol. Plant. 2018, 54, 565–575. [Google Scholar] [CrossRef]
- Kew 2019. Royal Botanic Gardens Kew Seed Information Database (SID). Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 18 September 2020).
- Darris, D.; Young-Mathews, A. Plant Fact Sheet for Riverbank Lupine (Lupinus rivularis); USDA NRCs Plant Materials Center: Corvallis, OR, USA, 2012.
- Costa, J.; Ricardo, C.P. Tunicamycin and swainsonine stimulate Lupinus albus L. root growth in vitro. Plant Sci. 1994, 101, 137–142. [Google Scholar] [CrossRef]
- Borek, S.; Kalemba, E.M.; Pukacka, S.; Pietrowska-Borek, M.; Stawiński, S.; Ratajczak, L. Nitrate simultaneously enhances lipid and protein accumulation in developing yellow lupin cotyledons cultured in vitro, but not under field conditions. J. Plant Physiol. 2017, 216, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Pniewski, T.; Kapusta, J.; Legocki, A.B. In vitro micropropagation of four lupin species. Acta Physiol. Plant. 2002, 24, 417–424. [Google Scholar] [CrossRef]
- Warren, R.; Vanderwal, J.; Price, J.C.; Welbergen, A.J.; Atkinson, I.; Ramirezvillegas, J.; Osborn, T.J.; Jarvis, A.; Shoo, L.P.; Williams, S.; et al. Quantifying the benefit of early climate change mitigation in avoiding biodiversity loss. Nat. Clim. Chang. 2013, 3, 678–682. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gitelson, A.; Buschmann, C.; Lichtenthaler, H.K. The chlorophyll fluorescence ratio F735/F700 as an accurate measure of the chlorophyll content in plants. Remote. Sens. Environ. 1999, 69, 296–302. [Google Scholar] [CrossRef]
- Popova, E.; Shukla, M.; Kim, H.H.; Saxena, P.K. Plant cryopreservation for biotechnology and breeding. Adv. Plant Breed. Strateg. Breed. Biotechnol. Mol. Tools 2015, 63–93. [Google Scholar] [CrossRef]
- Kim, H.-H.; Lee, Y.-G.; Shin, D.-J.; Ko, H.-C.; Gwag, J.-G.; Cho, E.-G.; Engelmann, F. Development of alternative plant vitrification solutions in droplet-vitrification procedures. CryoLetters 2009, 30, 320–334. [Google Scholar] [CrossRef]
- Knecht, K.T.; Nguyen, H.; Auker, A.D.; Kinder, D.H. Effects of extracts of lupine seed on blood glucose levels in glucose resistant mice. J. Herb. Pharmacother. 2006, 6, 89–104. [Google Scholar] [CrossRef]
- Kinder, D.H.; Knecht, K.T. Lupine (Lupinus caudatus L., Lupinus albus L.) Seeds: History of use, use as an antihyperglycemic medicinal, and use as a food. Nuts Seeds Health Dis. Prev. 2011, 711–716. [Google Scholar] [CrossRef]
- Elmer, W.H.; Yang, H.A.; Sweetingham, M.W. Characterization of Colletotrichum gloeosporioides isolates from ornamental lupines in connecticut. Plant Dis. 2001, 85, 216–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didur, I.M.; Prokopchuk, V.M.; Pantsyreva, H.V. Investigation of biomorphological and decorative characteristics of ornamental species of the genus Lupinus L. Ukr. J. Ecol. 2019, 9, 287–290. [Google Scholar] [CrossRef]
- Murthy, B.N.S.; Murch, S.J.; Saxena, P.K. Thidiazuron: A potent regulator of in vitro plant morphogenesis. In Vitr. Cell. Dev. Biol. Plant. 1998, 34, 267–275. [Google Scholar] [CrossRef]
- Singh, R.; Jat, R.S.; Sahoo, P.D.; Srinivasan, R. Thidiazuron induced multiple shoot formation in chickpea (Cicer arietinum L). J. Plant Biochem. Biotechnol. 2002, 11, 129–131. [Google Scholar] [CrossRef]
- Yucesan, B.; Turker, A.U.; Gurel, E. TDZ-induced high frequency plant regeneration through multiple shoot formation in witloof chicory (Cichorium intybus L.). Plant Cell Tissue Organ Cult. 2007, 91, 243–250. [Google Scholar] [CrossRef]
- Chhabra, G.; Chaudhary, D.; Varma, M.; Sainger, M.; Jaiwal, P.K. TDZ-induced direct shoot organogenesis and somatic embryogenesis on cotyledonary node explants of lentil (Lens culinaris Medik). Physiol. Mol. Biol. Plants 2008, 14, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, P.; Murch, S.J.; Sullivan, J.A.; Saxena, P.K. Development of an efficient protocol for high frequency in vitro regeneration of a horticultural plant Primulina tamiana (B.L. Burtt) Mich. Möller & A. Webber. Can. J. Plant Sci. 2014, 94, 1281–1287. [Google Scholar] [CrossRef] [Green Version]
- Malik, K.; Saxena, P. Thidiazuron induces high-frequency shoot regeneration in intact seedlings of pea (Pisum sativum), chickpea (Cicer arietinum) and lentil (Lens culinaris). Funct. Plant Biol. 1992, 19, 731. [Google Scholar] [CrossRef]
- Murthy, B.N.S.; Murch, S.J.; Saxena, P.K. Thidiazuron-induced somatic embryogenesis in intact seedlings of peanut (Arachis hypogaea): Endogenous growth regulator levels and significance of cotyledons. Physiol. Plant. 1995, 94, 268–276. [Google Scholar] [CrossRef]
- Ye, G.; McNeil, D.L.; Conner, A.J.; Hill, G.D. Multiple shoot formation in lentil (Lens culinaris) seeds. N. Z. J. Crop. Hortic. Sci. 2010, 30, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Monthony, A.S.; Baethke, K.; Erland, L.A.E.; Murch, S.J. Tools for conservation of Balsamorhiza deltoidea and Balsamorhiza sagittata: Karrikin and thidiazuron-induced growth. In Vitr. Cell. Dev. Biol. Plant. 2020, 56, 398–406. [Google Scholar] [CrossRef]
- Mundhara, R.; Rashid, A. Recalcitrant grain legume Vigna radiata, mung bean, made to regenerate on change of hormonal and cultural conditions. Plant Cell Tissue Organ Cult. 2006, 85, 265–270. [Google Scholar] [CrossRef]
- Zhihui, S.; Tzitzikas, M.; Raemakers, C.J.J.M.; Zhengqiang, M.; Visser, R. Effect of TDZ on plant regeneration from mature seeds in pea (Pisum sativum). In Vitr. Cell. Dev. Biol. Plant. 2009, 45, 776–782. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Naidoo, Y.; Da Silva, J.A.T. Thidiazuron-induced abnormalities in plant tissue cultures. Plant Cell Rep. 2018, 37, 1451–1470. [Google Scholar] [CrossRef]
- Surma, M.; Adamski, T.; Święcicki, W.; Barzyk, P.; Kaczmarek, Z.; Kuczyńska, A.; Krystkowiak, K.; Mikołajczak, K.; Ogrodowicz, P. Preliminary results of in vitro culture of pea and lupin embryos for the reduction of generation cycles in single seed descent technique. Acta. Soc. Bot. Pol. 2013, 82, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Daws, M.I.; Pritchard, H.W. The development and limits of freezing tolerance in Acer pseudoplatanus fruits across Europe is dependent on provenance. CryoLetters 2008, 29, 189–198. [Google Scholar]
- Ramírez-González, G.; Rodríguez-de, J.L.L.O.; Arreola-Ávila, J.G.; Álvarez-Moctezuma, J.G. Morphogenic responses of three explants of Lupinus montanus (H.B.K.) cultured in vitro. Rev. Chapingo Ser. Cienc. For. Ambient. 2015, 17–27. [Google Scholar] [CrossRef]
- Puddephat, I.; Alderson, P.; Wright, N. Influence of explant source, plant growth regulators and culture environment on culture initiation and establishment of Quercus robur L. in vitro. J. Exp. Bot. 1997, 48, 951–962. [Google Scholar] [CrossRef] [Green Version]
- Shekafandeh, A.; Khosh-Khui, M. Effects of bud position and culture medium on shoot proliferation from nodal culture of two mature guava cultivars. Asian J. Plant Sci. 2008, 7, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Hand, C.R.; Wada, N.; Stockwell, V.; Reed, B.M. Node position influences viability and contamination in hazelnut shoot explants. In Vitr. Cell. Dev. Biol. Plant. 2016, 52, 580–589. [Google Scholar] [CrossRef]
- Shukla, M.R.; Jones, A.M.P.; Sullivan, J.A.; Liu, C.; Gosling, S.; Saxena, P.K. In vitro conservation of American elm (Ulmus americana): Potential role of auxin metabolism in sustained plant proliferation. Can. J. For. Res. 2012, 42, 686–697. [Google Scholar] [CrossRef]
- Walters, C.; Wheeler, L.; Stanwood, P.C. Longevity of cryogenically stored seeds. Cryobiology 2004, 48, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, H.W.; Nadarajan, J. Cryopreservation of orthodox (desiccation tolerant) Seeds. Plant Cryopreserv. A Pract. Guid. 2008. [Google Scholar] [CrossRef]
- Pence, V.C. Cryopreservation of seeds of Ohio native plants and related species. Seed Sci. Technol. 1991, 19, 235–251. [Google Scholar]
- Tikhonova, V.L. Long-term storage of seeds. Russ. J. Plant Physiol. 1999, 46, 400–408. [Google Scholar]
- Kholina, A.B.; Voronkova, N.M. Conserving the gene pool of Far Eastern plants by means of seed cryopreservation. Izv. Akad. Nauk. Seriia Boil. 2008, 35, 262–269. [Google Scholar] [CrossRef]
- Popova, E.V.; Kim, D.H.; Han, S.H.; Moltchanova, E.; Pritchard, H.W.; Hong, Y.P. Systematic overestimation of Salicaceae seed survival using radicle emergence in response to drying and storage: Implications for ex situ seed banking. Acta Physiol. Plant. 2013, 35, 3015–3025. [Google Scholar] [CrossRef]
- Gonzalez-Benito, M.E.; Iriondo, J.M.; Pérez-García, F. Seed cryopreservation: An alternative method for the conservation of Spanish endemics. Seed Sci. Technol. 1998, 26, 257–262. [Google Scholar]
- Decruse, S.W.; Seeni, S.; Pushpangadan, P. Effects of cryopreservation on seed germination of selected rare medicinal plants of India. Seed Sci. Technol. 1999, 27, 501–505. [Google Scholar]
- Tarré, E.; Pires, B.B.M.; Guimarães, A.P.M.; Carneiro, L.A.; Forzza, R.C.; Mansur, E. Germinability after desiccation, storage and cryopreservation of seeds from endemic Encholirium Mart. ex Schult. & Schult. f. and Dyckia Schult. & Schult. f. species (Bromeliaceae). Acta Bot. Bras. 2007, 21, 777–783. [Google Scholar] [CrossRef]
- Pérez-García, F. Effect of cryopreservation, gibberellic acid and mechanical scarification on the seed germination of eight endemic species from the Canary Islands. Seed Sci. Technol. 2008, 36, 237–242. [Google Scholar] [CrossRef]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic plant species conservation: Biotechnological approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, E.V.; Nikishina, T.V.; Kolomeitseva, G.L.; Popov, A.S. The Effect of seed cryopreservation on the development of protocorms by the hybrid orchid Bratonia. Russ. J. Plant Physiol. 2003, 50, 672–677. [Google Scholar] [CrossRef]
- Nikishina, T.V.; Popova, E.V.; Vakhrameeva, M.G.; Varlygina, T.I.; Kolomeitseva, G.L.; Burov, A.V.; Popovich, E.A.; Shirokov, A.I.; Shumilov, V.Y.; Popov, A.S. Cryopreservation of seeds and protocorms of rare temperate orchids. Russ. J. Plant Physiol. 2007, 54, 121–127. [Google Scholar] [CrossRef]
- Zevallos, B.; Cejas, I.; Rodríguez, R.C.; Yabor, L.; Aragón, C.; González, J. Biochemical characterization of Ecuadorian wild Solanum lycopersicum Mill. plants produced from non-cryopreserved and cryopreserved seeds. CryoLetters 2016, 37, 413–421. [Google Scholar]
- Whiteley, S.E.; Bunn, E.; Menon, A.; Mancera, R.L.; Turner, S.R. Ex situ conservation of the endangered species Androcalva perlaria (Malvaceae) by micropropagation and cryopreservation. Plant Cell Tissue Organ Cult. 2016, 125, 341–352. [Google Scholar] [CrossRef]
- Streczynski, R.; Clark, H.; Whelehan, L.M.; Ang, S.-T.; Hardstaff, L.K.; Funnekotter, B.; Bunn, E.; Offord, C.A.; Sommerville, K.D.; Mancera, R.L. Current issues in plant cryopreservation and importance for ex situ conservation of threatened Australian native species. Aust. J. Bot. 2019, 67, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sakai, A.; Engelmann, F. Vitrification, encapsulation-vitrification and droplet-vitrification: A review. CryoLetters 2007, 28, 151–172. [Google Scholar]
- Wang, M.-R.; Lambardi, M.; Engelmann, F.; Pathirana, R.; Panis, B.; Volk, G.M.; Wang, Q.-C. Advances in cryopreservation of in vitro-derived propagules: Technologies and explant sources. Plant Cell Tissue Organ Cult. 2020, 1–14. [Google Scholar] [CrossRef]
- Kim, H.-H.; Lee, Y.G.; Park, S.U.; Lee, S.C.; Baek, H.J.; Cho, E.G.; Engelmann, F. Development of alternative loading solutions in droplet-vitrification procedures. CryoLetters 2009, 30, 291–299. [Google Scholar] [PubMed]
- Rathwell, R.; Popova, E.; Shukla, M.R.; Saxena, P.K. Development of cryopreservation methods for cherry birch (Betula lenta L.), an endangered tree species in Canada. Can. J. For. Res. 2016, 46, 1284–1292. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.H.; Popova, E.; Lee, H.; Park, S.U.; Ku, J.; Kang, J.H.; Kim, H.H. Cryopreservation of endangered wild Species, Aster altaicus var. uchiyamae Kitam, using droplet-vitrification procedure. CryoLetters 2019, 40, 113–122. [Google Scholar] [PubMed]
- Yang, X.; Popova, E.; Shukla, M.R.; Saxena, P.K. Root cryopreservation to biobank medicinal plants: A case study for Hypericum perforatum L. In Vitr. Cell. Dev. Biol. Plant. 2019, 55, 392–402. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BA Concentration (µM) | Explant Type | Explant Response (%) | Shoot Height (mm) | Avg. nos. of Shoot/Explant |
|---|---|---|---|---|
| 0 | Tip | 100 | 18.80 abc | 1.0 de |
| Mid | 30 | 1.34 e | 0.3 e | |
| Base | 60 | 8.61 de | 1.2 bcde | |
| 2 | Tip | 100 | 20.94 ab | 2.2 abcd |
| Mid | 60 | 4.74 de | 1.1 cde | |
| Base | 80 | 7.97 de | 1.5 abcde | |
| 5 | Tip | 100 | 24.59 a | 2.7 ab |
| Mid | 100 | 12.36 bcd | 1.8 abcde | |
| Base | 90 | 13.34 bcd | 2.0 abcd | |
| 10 | Tip | 100 | 19.66 ab | 2.6 abd |
| Mid | 90 | 9.69 cde | 2.0 abcd | |
| Base | 100 | 14.07 bcd | 2.8 a | |
| 20 | Tip | 100 | 18.66 abc | 2.1 abcd |
| Mid | 80 | 9.74 cde | 1.6 abcde | |
| Base | 70 | 10.02 cde | 1.0 e |
| Seed Lot | Treatment | Number of Leaves | Number of Branches | Length of the Longest Branch (cm) | Number of Flower Stalks | Chlorophyll Content (mg/m2) |
|---|---|---|---|---|---|---|
| Lot 1 | Control | 5.8 ± 0.32 b | 5.9 ± 0.4 a | 21.1 ± 1.4 a | 1.4 ± 0.2 a | 454.2 ± 7.7 a |
| Cryopreserved | 5.8 ± 0.26 b | 6.0 ± 0.4 a | 18.9 ±1.4 a | 1.6 ± 0.2 a | 470.4 ± 10.9 a | |
| Lot 2 | Control | 8.0 ± 0.5 a | 5.6 ± 0.7 a | 18.4 ± 2.5 a | 1.0 ± 0.0 a | 451.9 ±11.2 a |
| Cryopreserved | 8.1 ± 0.8 a | 3.0 ± 1.5 b | 24.7 ± 5.0 a | 1.5 ± 0.7 a | 462.5 ±9.4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popova, E.V.; Shukla, M.R.; McIntosh, T.; Saxena, P.K. In Vitro and Cryobiotechnology Approaches to Safeguard Lupinus rivularis Douglas ex Lindl., an Endangered Plant in Canada. Agronomy 2021, 11, 37. https://doi.org/10.3390/agronomy11010037
Popova EV, Shukla MR, McIntosh T, Saxena PK. In Vitro and Cryobiotechnology Approaches to Safeguard Lupinus rivularis Douglas ex Lindl., an Endangered Plant in Canada. Agronomy. 2021; 11(1):37. https://doi.org/10.3390/agronomy11010037
Chicago/Turabian StylePopova, Elena V., Mukund R. Shukla, Terry McIntosh, and Praveen K. Saxena. 2021. "In Vitro and Cryobiotechnology Approaches to Safeguard Lupinus rivularis Douglas ex Lindl., an Endangered Plant in Canada" Agronomy 11, no. 1: 37. https://doi.org/10.3390/agronomy11010037
APA StylePopova, E. V., Shukla, M. R., McIntosh, T., & Saxena, P. K. (2021). In Vitro and Cryobiotechnology Approaches to Safeguard Lupinus rivularis Douglas ex Lindl., an Endangered Plant in Canada. Agronomy, 11(1), 37. https://doi.org/10.3390/agronomy11010037