1. Introduction
Gooseberry (
Ribes grossularia L.) is a small fruit crop producing valuable fruits, which is constantly gaining importance. Gooseberry fruits are a rich source of bioactive compounds such as vitamins, especially vitamin C, mineral salts, organic acids, dietary fiber, and antioxidant polyphenols, which are considered beneficial to human health [
1,
2,
3]. Gooseberries are grown on large commercial plantations in some European countries with favorable agroclimatic conditions, including Poland, which is the second largest producer of these fruits in Europe [
3].
Gooseberries are propagated vegetatively by cuttings or layering, but the rooting efficiency is below 50% [
4]. There are several scientific reports on gooseberry micropropagation [
5,
6,
7,
8,
9,
10,
11]. In vitro propagation of this species is difficult due to the appearance of shoot necrosis during initiation and multiplication stages, lack of shoot elongation, leaf yellowing, and hyperhydration. For the rapid multiplication of selected gooseberry clones, an effective method of micropropagation using
meta-topolin (m-T) instead of benzylaminopurine (BAP) was developed [
12]. This method can significantly support the production of virus-free planting material and accelerate the introduction of new cultivars to the market.
The successful application of in vitro techniques in plant multiplication depends on, among other things, maintaining genetic stability of the genotype. In plant tissue cultures, especially those maintained for a longer time, somaclonal variation (SV) is a common problem [
13,
14]. SV, either of genetic or epigenetic nature, appearing among clones propagated in in vitro, results either from pre-existing genetic variation within explants or is induced during micropropagation [
15]. The character of SV includes, among other things, changes in the number and structure of chromosomes, changes in nucleotide sequences and the number of gene copies, activation of transposons, and alterations in DNA methylation pattern [
13,
14,
16].
SV can be identified by analyzing morphological, biochemical, physiological, and genetic characteristics of plants. Methods based on molecular markers more precisely detect the variability than other techniques, and they are easy in application and guarantee high repeatability of results. Molecular markers such as random amplified polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), inter simple sequence repeat (ISSR), or other microsatellite markers are successfully used as tools to study not only somaclonal variability, but also to identify genotypes, study germplasm genetic diversity, and identify putative DNA markers of valuable traits that might be used in marker-assisted selection in many fruit plant species [
4,
13,
17,
18,
19], including the genus
Ribes [
20,
21,
22,
23,
24,
25,
26,
27].
There are few reports available concerning gooseberry DNA markers. Lanham and Brennan [
21] used RAPD, ISSR, and AFLP markers to estimate the genetic diversity within this species. Unique profiles for each of the 12 tested gooseberry genotypes were generated only by AFLP. The other techniques did not allow the distinguishing of genotypes. Molecular markers based on RAPD and ISSR techniques were applied to characterize gooseberry genotypes collected in the germplasm collection of the Research Institute of Horticulture in Skierniewice, Poland, used in breeding programs [
17]. The results confirmed the usefulness of both techniques in estimating the genetic distance between the 12 gooseberry genotypes tested; however, the ISSR analysis was assessed as being more precise and repetitive.
To date, no scientific reports on SV generated by in vitro gooseberry cultures have been published. In our research, we decided to use the AFLP technique, as it was considered to be the most effective in detecting DNA polymorphism in gooseberry [
21], and is widely used in other species [
28,
29,
30,
31]. In order to increase the reliability of the analysis, a combination of two or more markers was used [
32,
33,
34]. In our research, besides AFLP, ISSR markers were applied. ISSR markers were also used to compare the level of genetic diversity within in vitro-derived and vegetative seedlings.
4. Discussion
In vitro plant propagation is one of the most important methods of vegetative reproduction used in horticultural production. The main advantage of micropropagation is the possibility of obtaining, in a short time, a large number of true-to-type, high-quality, and pathogen-free plants. Micropropagation of fruit plants is successfully used for the production of elite nursery material, in breeding work and in the mass production of seedlings of species difficult to reproduce in a traditional way [
40,
41,
42,
43].
It has been demonstrated, in many scientific reports, that plants produced in in vitro tissue cultures show greater juvenility and productivity. Berry plants obtained from in vitro cultures are characterized by increased vegetative growth and more intense rhizome and fruit formation than plants obtained by standard multiplication methods. This phenomenon has been observed in strawberry [
44,
45], raspberry [
46], blueberry [
47,
48,
49], cranberry [
50], and lingonberry [
51].
There are no scientific reports on the growth of in vitro-derived gooseberry plants under field conditions except for one publication by Wainwright and Flegmann [
7]. The authors found that in vitro-derived gooseberry plants cv. Invicta developed more shoots at the base of the bushes and had more thorns compared to plants propagated by the standard method. In our field experiment, for three studied cultivars, the plants’ growth vigor and fruit yield were greater in in vitro-derived plants than in plants propagated from softwood cuttings. In the case of one cultivar, Resika, most of these parameters were higher in plants propagated by the standard method. This phenomenon can be explained by the influence of plant growth regulators, mainly cytokinins, used during in vitro propagation; their residues may increase plant vigor and yield in ex vitro conditions [
45,
52].
Many scientific reports indicate changes in the structure, shape, and density of the stomata in the leaves of plants grown in in vitro cultures compared to those grown in a greenhouse or in the field. Noe and Bonini [
53] reported that leaves of
Vaccinium corymbosum collected from in vitro shoots had a higher number of smaller stomata (361 mm
−2) compared to leaves taken from plants grown in the field (241 mm
−2). Brainerd and Fuchigami [
54] made similar observations on apple trees and Blanke and Belcher [
55] and Capellades et al. [
56] on rose. Joshi et al. [
57] observed that the leaves of in vitro seedlings showed twice the frequency of stomata compared to traditionally propagated seedlings of
Wrightia tomentosa. The opposite results were found for the plum cv. Pixi in a study by Brainerd and co-workers [
58]. In our field experiment, a higher frequency of stomata on leaves of MP plants was demonstrated for all studied gooseberry cultivars. Stomatal morphology and structure are highly affected by environmental conditions, such as relative humidity, and light level. In in vitro conditions, these two parameters (high humidity and low light levels) differ significantly from those ex vitro [
55,
59]. The high humidity in the culture vessel causes the transpiration in in vitro conditions to be low [
54]. The water imbalance exhibited by micropropagated plants can result in deformation and malfunctioning of the stomata [
55,
59,
60].
Somaclonal variability appearing in plant tissue cultures depends on many factors, such as the type of explant, the culture conditions, including the composition of the propagation medium, especially the type and concentration of growth regulators and culture age [
13,
14,
15,
16]. Of the many factors, genotype is considered to be the most important determinant of variability [
14,
16]. We did not observe phenotypic variability among the obtained MP plants of all the gooseberry cultivars tested. In order to assess the influence of the applied in vitro propagation method on the appearance of SV among progeny plants, we examined variability at the DNA level. In our research, the level of genetic variability in micropropagated gooseberry plants varied depending on the cultivar. Using both marker systems, the lowest genetic variability was detected in Hinnonmaki Rot and the highest in Invicta. Mohamed and co-workers [
61] assessed the genetic stability of in vitro-derived plants of several strawberry cultivars using RAPD markers. The study also revealed that the frequency of genetic variation depends on the cultivar studied; plants of Tudla, depending on the culture conditions, showed polymorphism within 2.86–11.77%, while the variability in the cultivar Festival ranged from 10.35 to 30.77%.
The age of the culture is another factor considered to have a strong impact on inducing variability in in vitro cultures [
13,
14]. The MP plants used for the research were obtained from stabilized two-year-old shoot cultures, subcultured every 6 weeks. In these plants, the level of genetic variation was low and ranged from 0.35% to 2.12% (AFLP markers) and from 0% to 8.69% (ISSR markers). Nookaraju and Agrawal [
62] demonstrated clonal homogeneity of six-month-old tissue culture raised plants of grapevine cv. Crimson Seedless compared to donor plants using ISSR and simple sequence repeat (SSR) markers. In a study by Khan [
63], the variability of in vitro shoots of blackcurrant cv. Ben Sark increased significantly when plants were kept in cultures for a long time. As assessed with the use of RAPD markers, micropropagated shoots showed no genetic variation up to the fourteenth subculture, the first polymorphic products were detected in the fifteenth subculture, and the final genetic diversity detected in the study was 13.4% (after 16 subcultures). The same analysis was performed for micropropagated raspberry cv. Autumn Bliss. Genetic variability was already detected in the fifth subculture and the maximum variation observed was 26% [
63]. In some plant species, genetic variability can reach a very high level, especially when the conditions of the culture are favorable for this phenomenon. In plants derived from four-year pineapple in vitro cultures, the analysis, with the use of RAPD markers, showed as much as 95.9% variability [
64]. On the other hand, Borsai and co-workers [
65], using sequence-related amplified polymorphism (SRAP) and RAPD markers, did not detect any polymorphism in plants of blackberry cv. Loch Ness and Chester Thornless propagated for both 3 and 11 subcultures. Similarly, no genetic variability was observed by Negi and Saxena [
66] using ISSR markers in
Bambusa balcooa plants multiplied up to 33 passages or by Singh and co-workers [
67] in
Dendrocalamus asper in vitro raised shoots up to the thirtieth passage, checked with the use of RAPD, ISSR, SSR, and AFLP markers.
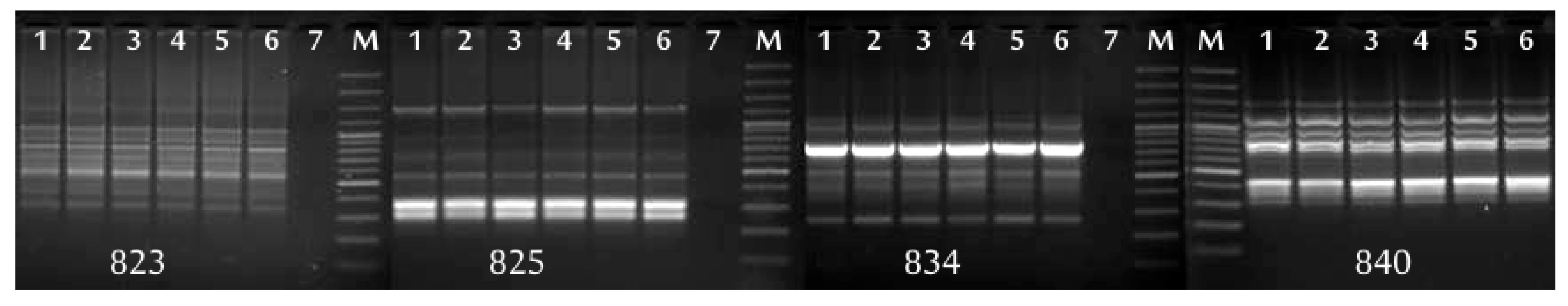
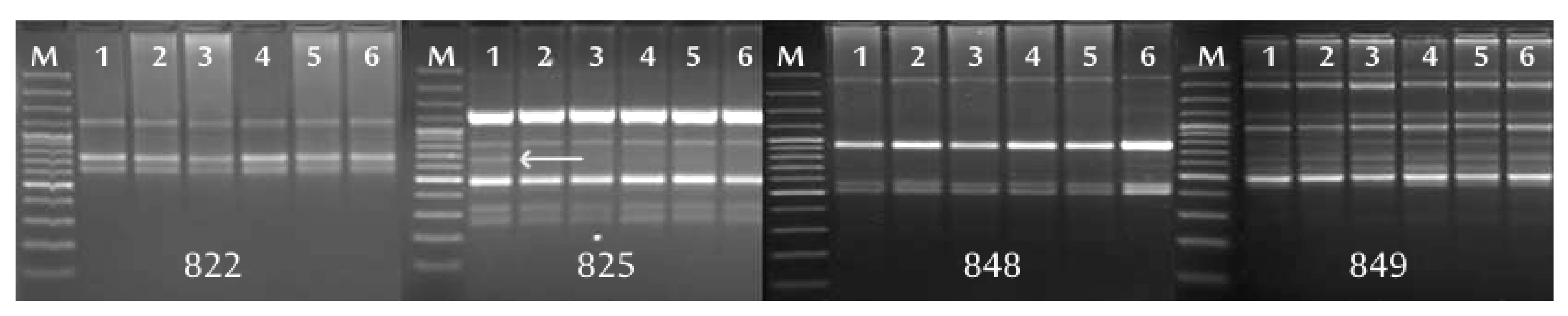
In research using molecular markers, the quality of the DNA preparations used in the analyses is of great importance. The isolation of genomic DNA from plants of the genus
Ribes is most often carried out by the method according to Doyle and Doyle [
68], based on an extraction buffer containing CTAB (hexadecyltrimethylammonium bromide) [
17,
26] or the method developed by Milligan [
20,
21,
22,
24,
69]. Compared to the technique using commercial DNA isolation kits, these methods are longer and more labor-intensive. All commercial kits used in our study enabled to isolate DNA from small amounts of plant material and to obtain preparations of genomic DNA with sufficient DNA content and purity for AFLP and ISSR analyses. In our study, AFLP markers detected genetic variability in MP plants of all gooseberry genotypes, while ISSR markers only did so in Captivator and Invicta. In order to assess the genetic variability of berry plants, the RAPD and ISSR markers are most frequently used. In several research papers, the use of AFLP, SSR, and SRAP markers was also reported [
17,
19,
21,
25,
61,
62,
63,
65,
70,
71,
72]. AFLP analysis proved to be more reliable and sensitive than other methods not only in our research, but also in the studies of Lanham and Brennan [
21] and Costa and co-workers [
31]. The AFLP marker system is believed to be a very sensitive and reproducible method [
13,
30,
31]; no prior knowledge of DNA sequences is required and it can check large fragments of the genome, but it also has disadvantages—it is time-consuming, technically demanding and expensive. In terms of the facility and speed of performing the analysis, microsatellite markers are more advantageous than AFLP. In our research, ISSR analysis proved to be less reliable than the AFLP. Presumably, the reason is that only five primers were used to analyze each gooseberry cultivar; therefore, a small number of products for analysis was obtained.
The results obtained showed that the method of in vitro propagation of four gooseberry cultivars used in our research is suitable for obtaining plants with high genetic fidelity. It supports the opinion that variability is very rare when the technique of shoot regeneration from lateral buds is used in micropropagation and is consistent with the results obtained by Saker et al. [
73] and Bhatia et al. [
74] for date palm and gerbera.






{kind=link}
{kind=link}
{kind=link}