1. Introduction
Cassava (
Manihot esculenta), a native crop to tropical America, is called a 21st century crop [
1] because it has the ability to adapt to poor environmental conditions, such as semi-arid land, acidic soil, and low soil fertility. It has been widely used for staple foods, livestock feeds, processed foods, and starch production, mainly in tropical Asia and Africa [
1]. Cassava tubers have a higher starch content than other tuber crops [
2], and it was reported that the conversion efficiency of solar energy per unit land area to carbohydrates under favorable conditions exceeds that of maize and rice [
3,
4]. The global cassava production increased by 3.9 times between 1961 and 2018. By region, it increased by 5.4 times in Africa and 4.5 times in Asia. Compared to other major crops, cassava has the second highest growth rate after corn (5.6 times the growth rate during the same period). The harvest area of cassava expanded 2.6 times worldwide during the same period, with the largest increase rate in Africa (3.4 times), which has expanded rapidly since 2011, while the increase rate in other regions was relatively small (1.2 to 1.7 times). On the other hand, its yield increased by 1.5 times worldwide during the same period. By region, it increased by 2.7 times in Asia, followed by 1.6 times in Africa. From these trends in cassava production over the past half century, it can be seen that production has increased due to area expansion in Africa and increased yields in Asia. Especially in Asia, it is evident that the yield increased sharply after 1995 [
5]. The increase in cassava production has been supported by the development and dissemination of regional adaptable varieties centered on Centro Internacional de Agricultura Tropical (CIAT), the International Institute of Tropical Agriculture (IITA), and agricultural research institutes in each country. With the expansion of cassava production in recent years, cultivation cases have been reported in marginal regions utilizing the poor environmental resistance of cassava. Some examples have been reported, such as an investigation of the productivity of initial-maturing varieties using the fallow period after paddy rice cultivation in the tropical region of Asia [
6], a productivity comparison in the high-altitude areas of tropical and subtropical regions [
7,
8], and the development of a resistant variety to low temperature stress that grows in the subtropical inland region during winter [
9]. On the other hand, among the photosynthesis-related traits that form the basis of substance production, the maximum rate to receive the light energy projected on the crop canopy is as low as about 64% compared to soybean (90%), and the plant shape has been improved through breeding. The conversion rate from photosynthetic active radiation (PAR) to the biomass of cassava is around 1.4%, lower than that of soybean, which is 2.9~4.3%. Furthermore, El-Sharkwy and De Tafur [
10,
11] pointed out that no improvement in photosynthesis rate was observed between native and cultivars, and these results suggest that the previous breeding traits did not include photosynthesis-related traits.
Cassava cultivation in Japan is carried out only by small-holder farmers in the Kagoshima Prefecture island area and Okinawa Prefecture located in the southern part of the country. There are not many studies from the viewpoint of agronomics other than Minami et al. [
12] and Minoda et al. [
13]. Minami et al. cultivated cassava introduced from Brazil for three years in Kagoshima, which has an eight-month frost-free period from April to November, and reported that 20 t ha
−1 of raw potato yield, comparable to the yield of sweet potatoes cultivated in the same region, could be obtained. In Kagoshima Prefecture, sweet potato production is the highest in Japan and it is supplied to the starch, alcohol, and brewing industries. However, in recent years, the invasion of new pests and diseases that are thought to be related to rising temperatures has become a problem. [
10]. As such, it has become important to search for new agricultural resources for regional industrial promotion as part of the climate change mitigation measures. At present, although the cultivation techniques and knowledge of cassava cultivation in Kagoshima are scarce, the fact that a certain yield has been obtained is expected to contribute to the expansion of cassava cultivation and production areas. On the other hand, there are many differences between the results of experiments conducted in the tropics and the results of Minami et al. [
12]. For example, the leaf area index (LAI) was generally reported to reach its maximum at 4–5 MAP (months after planting) [
14,
15], but according to Minami et al. [
12], the maximum LAI was measured 6 MAP. There was also big gap between their report, in which the three years average of harvest index was 0.3, and another report, in which it ranged between 0.46 and 0.71 including varietal difference [
12,
16]. Minami et al. [
12] cite the effects of low sunshine and low temperature on this growth delay. The frost-free period in Kagoshima is about eight months, but it includes the period below the low temperature limit of cassava growth, which is said to be an average daily temperature of 18 to 20 °C [
7]. In addition, the effects of heavy rainfall that occurs about 2 to 3 MAP and the accompanying low sunshine are also factors to consider. Therefore, it is necessary to clarify the growth characteristics of cassava in Kagoshima, which is a subtropical island region. In this study, among the photosynthesis-related traits that are the basis of the dry matter production of cassava, we focused on the canopy formation, the gas exchange rate of individual leaves, and the distribution of photosynthesis products. Therefore, this experiment was conducted to clarify the effect of the change in the community structure under different nitrogen fertilization conditions on the gas exchange rate, productive structure, dry matter production, and dry matter distribution (DMD). It was assumed that fertilization at the planting stage increases the gas exchange rate and promotes the developments of leaves and stems, resulting in the accelerated establishment of a plant community. The increase of the total dry weight and root DMD is expected to contribute the increase of the root dry weight. On the other hand, we hypothesize that although fertilization at a later growth stage increases the photosynthesis rate, root DMD does not change since the dry matter will be distributed to the leaves, stems, and roots. Therefore, it is thought that only an increase of the total dry weight contributes an increase in the tuberous root dry weight.
4. Discussion
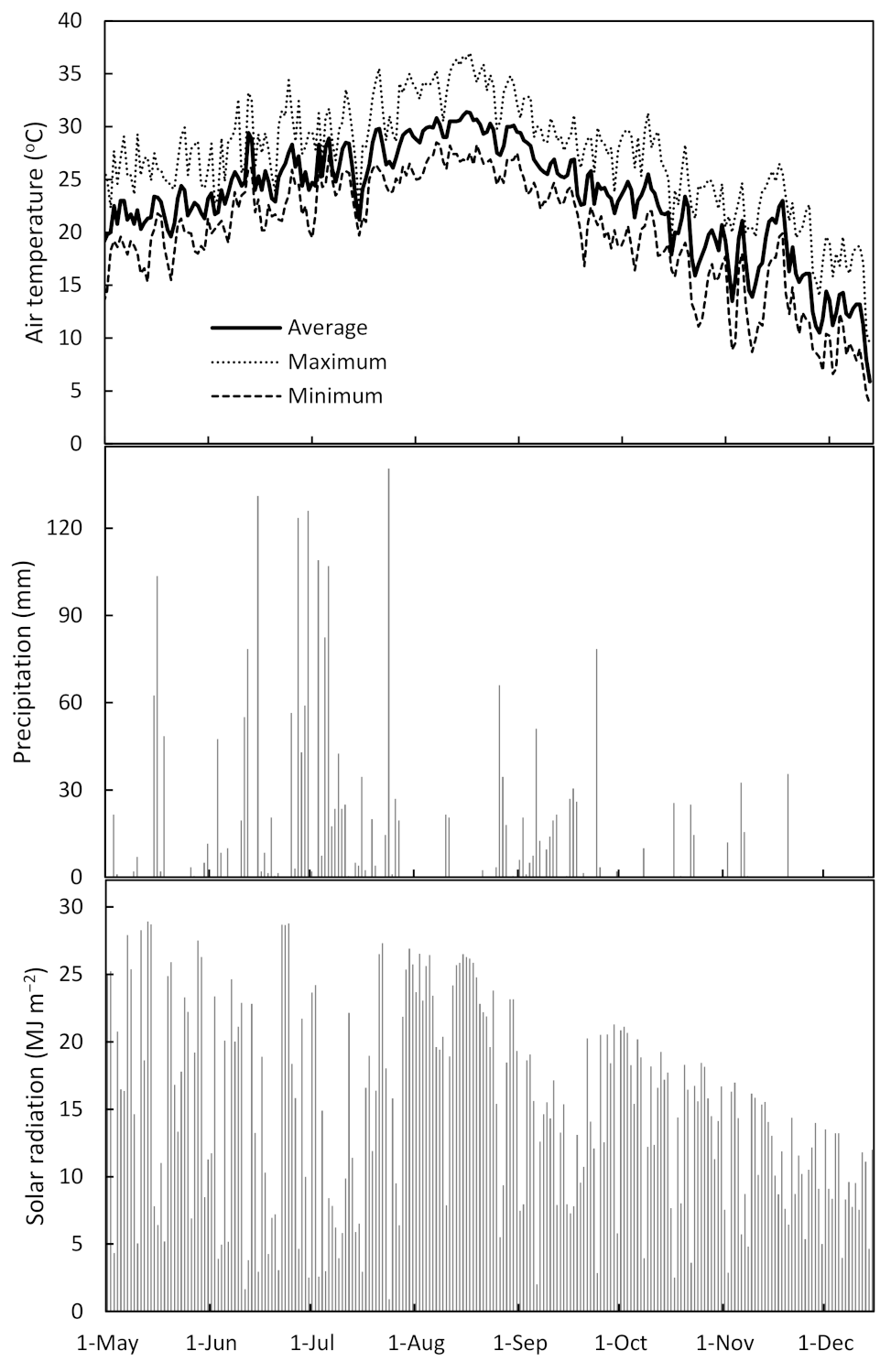
First, the effects of the weather conditions in 2020 on the result of this study were examined. The total temperature and total solar radiation were at the same levels as the average year, but the total precipitation was higher than that at the average year due to the effect of the rainy season in June and July. However, it is assumed that the rhizosphere of cassava was not submerged because the cuttings were transplanted on top of the ridges. The total solar radiation in July was 76.3% of the average year, because of the low insolation during the rainy season. Although there was significantly higher gs at the initial fertilization treatment on July, it is probable that the effect of fertilization on gs was limited, since the stomata does not open under low solar radiation. However, the total dry weight of the initial fertilization treatment was significantly higher than that of the control, so it is considered that the effects of the low insolation and high precipitation in July on the conclusions of this study are not strong. Since stable climatic conditions continued after September, except for the typhoon, the effect of the latter fertilization treatment is expected to be the same as the average year conditions.
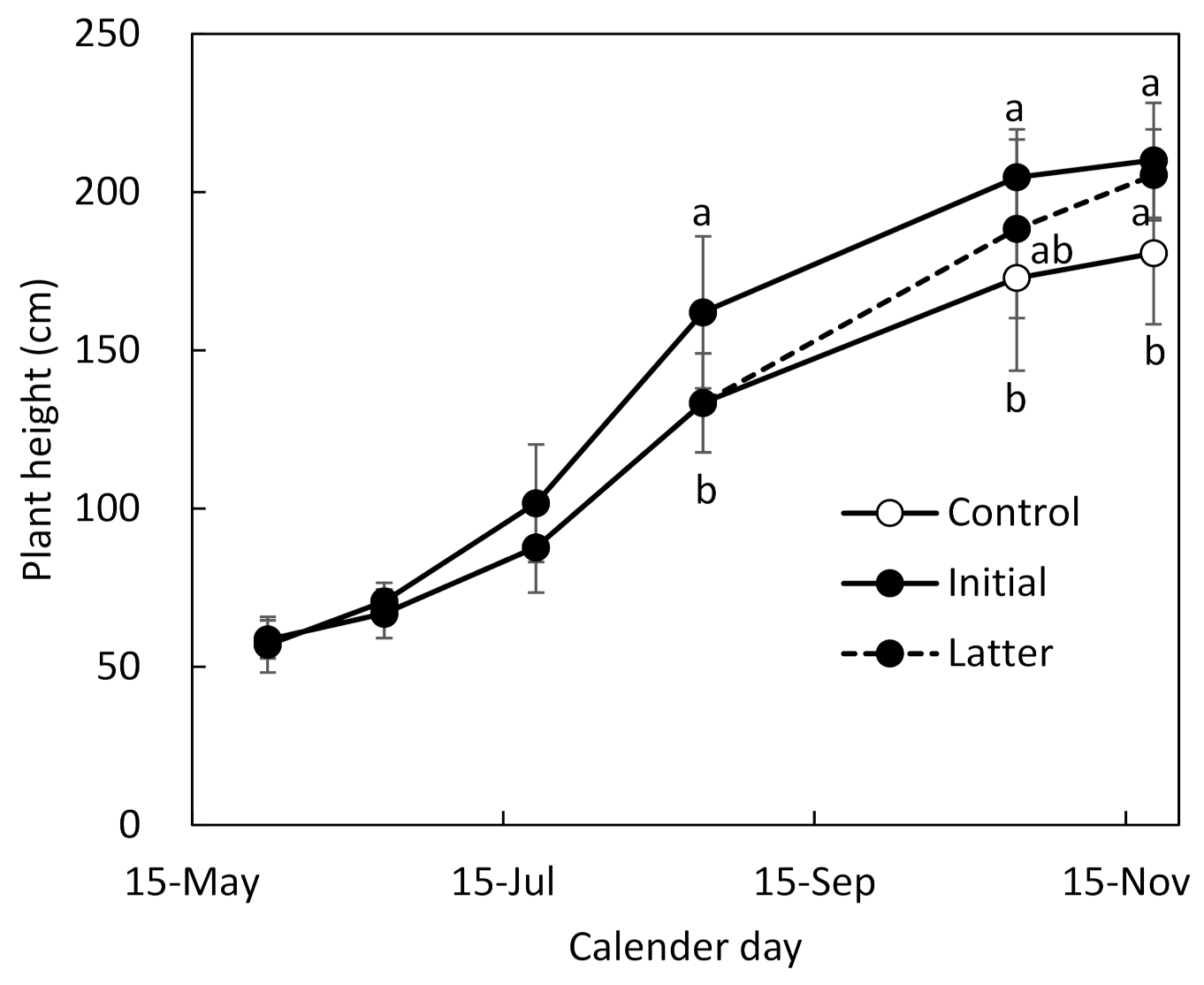
In this study, cassava of tropical origin was cultivated in Kagoshima, which belongs to the temperate zone, under different fertilization conditions, and its growth characteristics were investigated from the viewpoint of individual leaf gas exchange rate, plant community structure, and dry matter production and distribution. The initial fertilization promoted an increase in plant height from the initial growth stage, resulting in early canopy closure. The increase of plant height on latter fertilization continued in October and November, when the plant height increases of the control and initial fertilization declined. These results indicated that fertilization at different growth stages can affect canopy development.
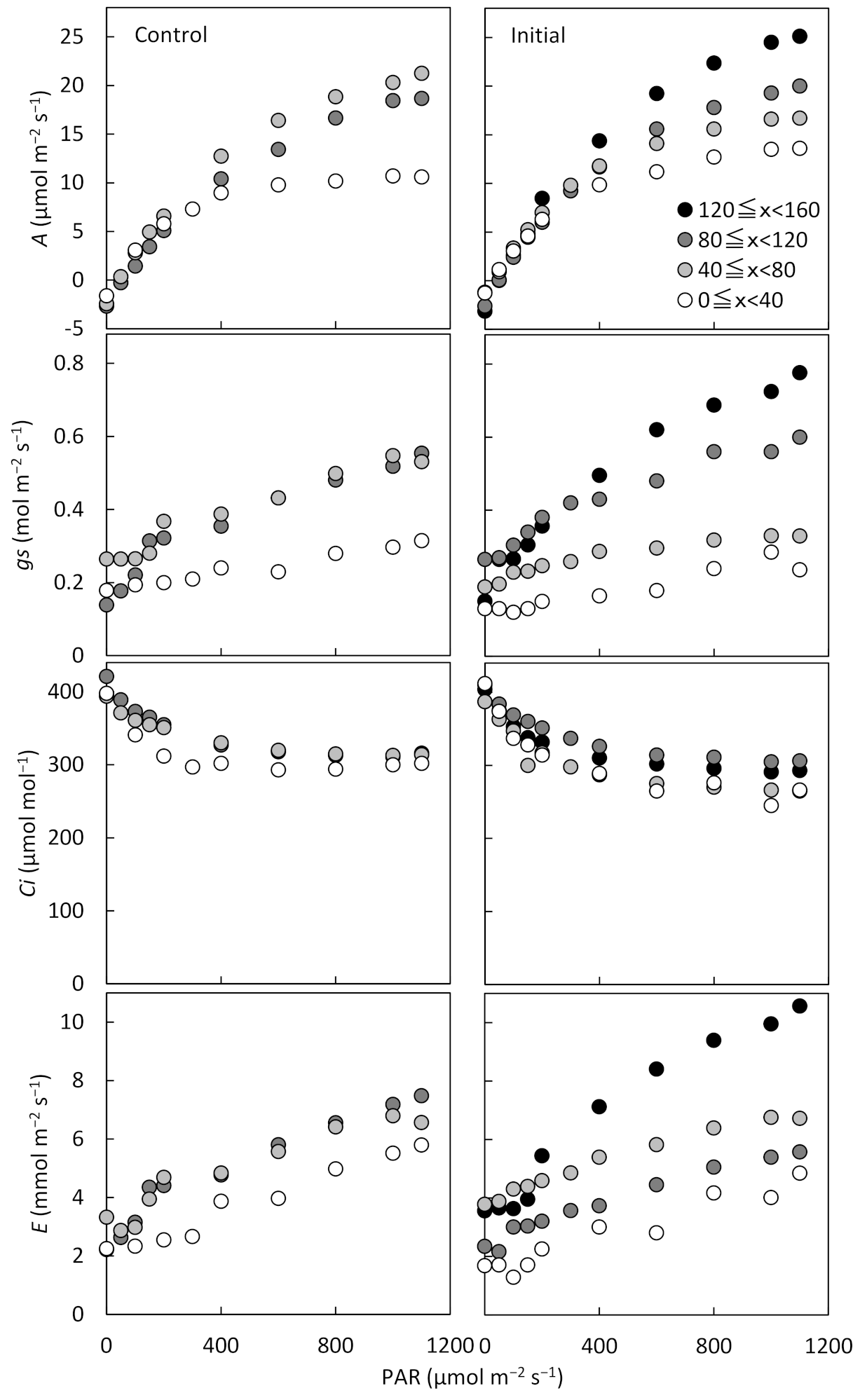
The response of leaves in the upper layer, which has high photosynthetic activity, to light intensity was compared with previous reports. While the
Amax in this study was 21.3–25.1 μmol m
−2 s
−1, Santanoo et al. [
18] and Mahakosee et al. [
19] reported it to be approximately 30 μmol m
−2 s
−1. PAR at a light saturation of
A was also reported to be about 1500 μmol m
−2 s
−1, higher than the 1200 μmol m
−2 s
−1 reported in this study. One of the reasons for this is that Kagoshima, where this experiment was conducted, is at a higher latitude and receives less solar radiation than Khon Kaen, Thailand, where Santanoo et al. [
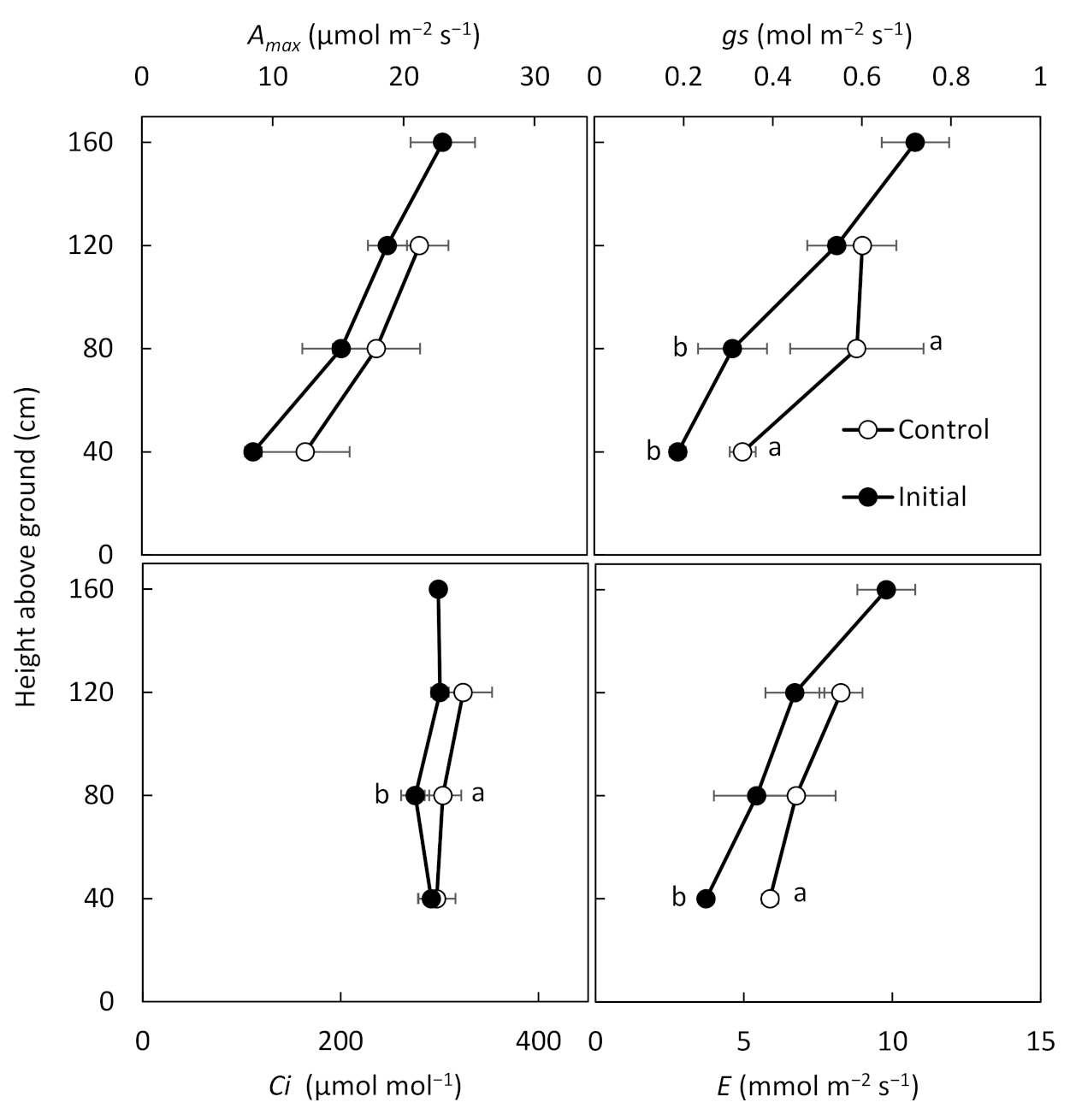
18] conducted their experiments. The result of light curve on different canopy height levels showed high
Amax on the upper layer and photosynthesis was conducted under high solar radiation. On the other hand, although the lower leaves had a low
Amax, they showed the same level of
A as the upper leaves, when the PAR was less than 400 μmol m
−2 s
−1. This demonstrates that they adapted to the low solar radiation environment inside the crop community. This result is also consistent with the report by Santanoo et al. [
18], which divided the canopy into six layers and measured
A in four varieties of cassava. A comparison of the photosynthetic parameters during the light saturated condition in August (
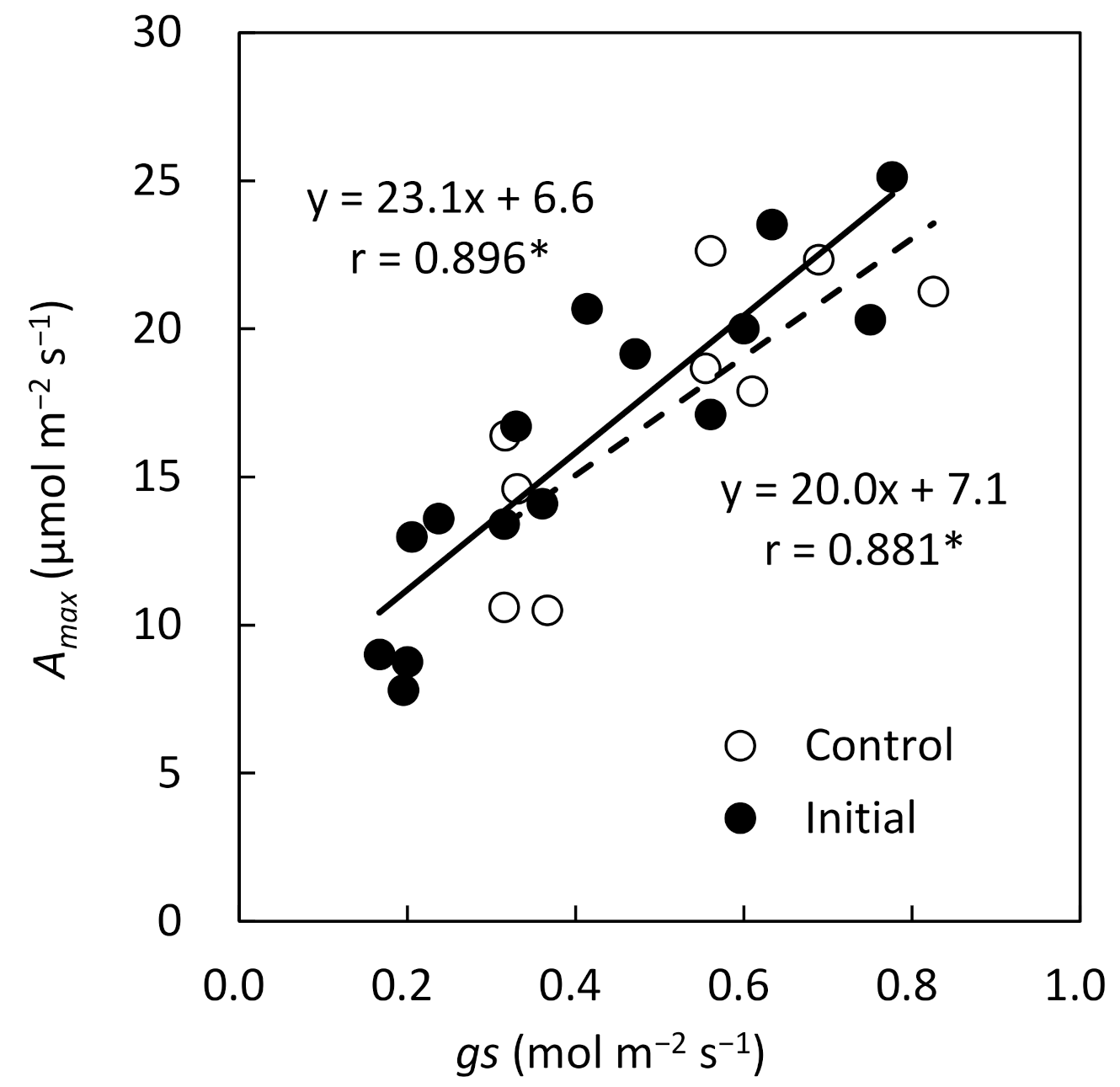
Figure 3) showed that there were no significant differences at each height level between treatments. This was the result of adaptation to the degraded light environment, as the initial fertilization promoted increased plant height and new leaf expansion, causing self-shading. In addition, there were significant correlations between gs and
A at initial fertilization and the control, and it was suggested that the
A can be estimated using
gs as an index even under different fertilization conditions (
Figure 4). The effect of initial fertilization on
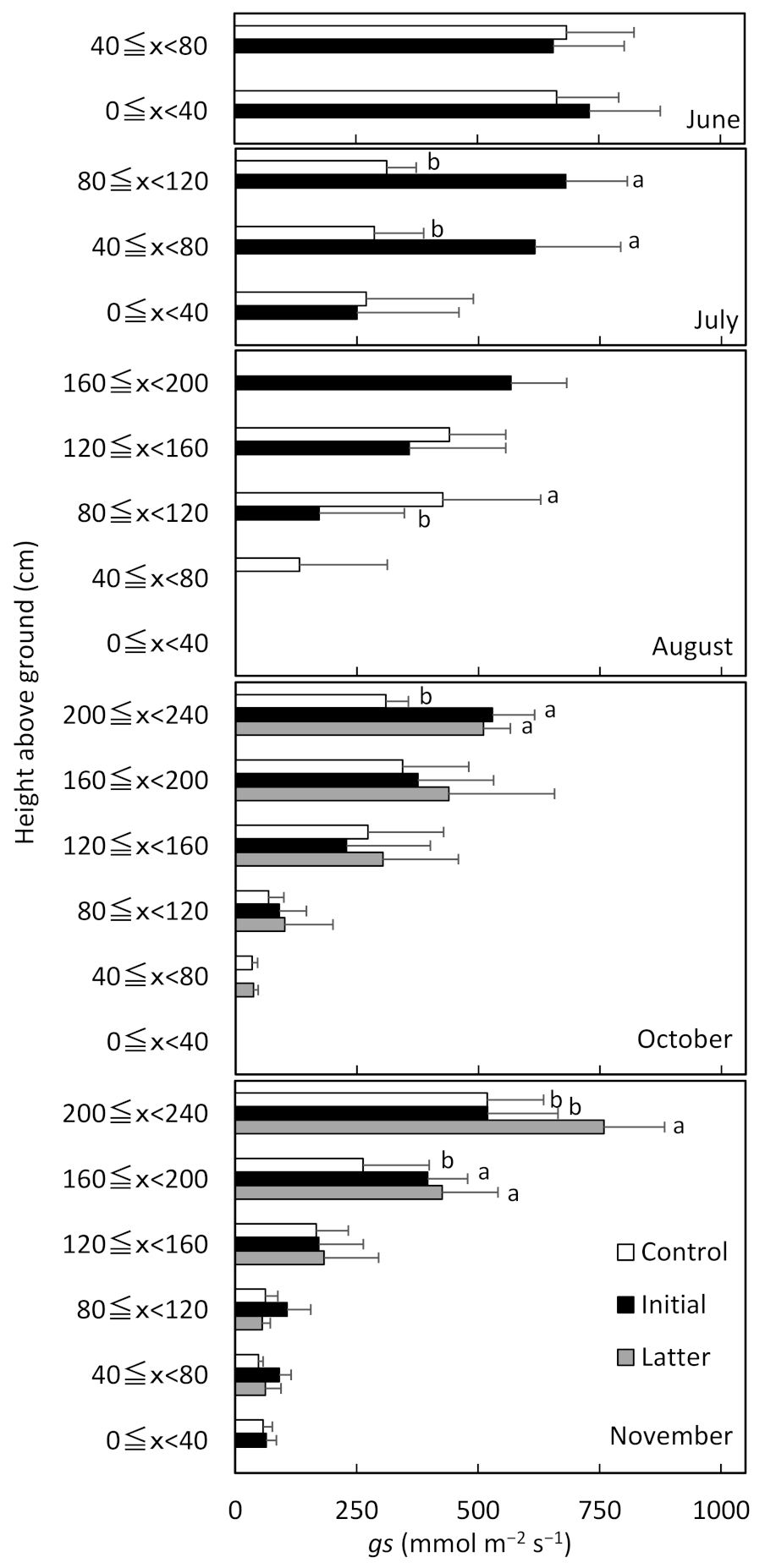
gs was apparent in July and August, especially in the top canopy layer exposed to high solar radiation. There were significant differences in gs in October and November between the latter fertilization treatment and the other treatments. These results make it clear that fertilization in the initial and latter growth stage of cassava improves photosynthetic activity on the upper layer of the crop community.
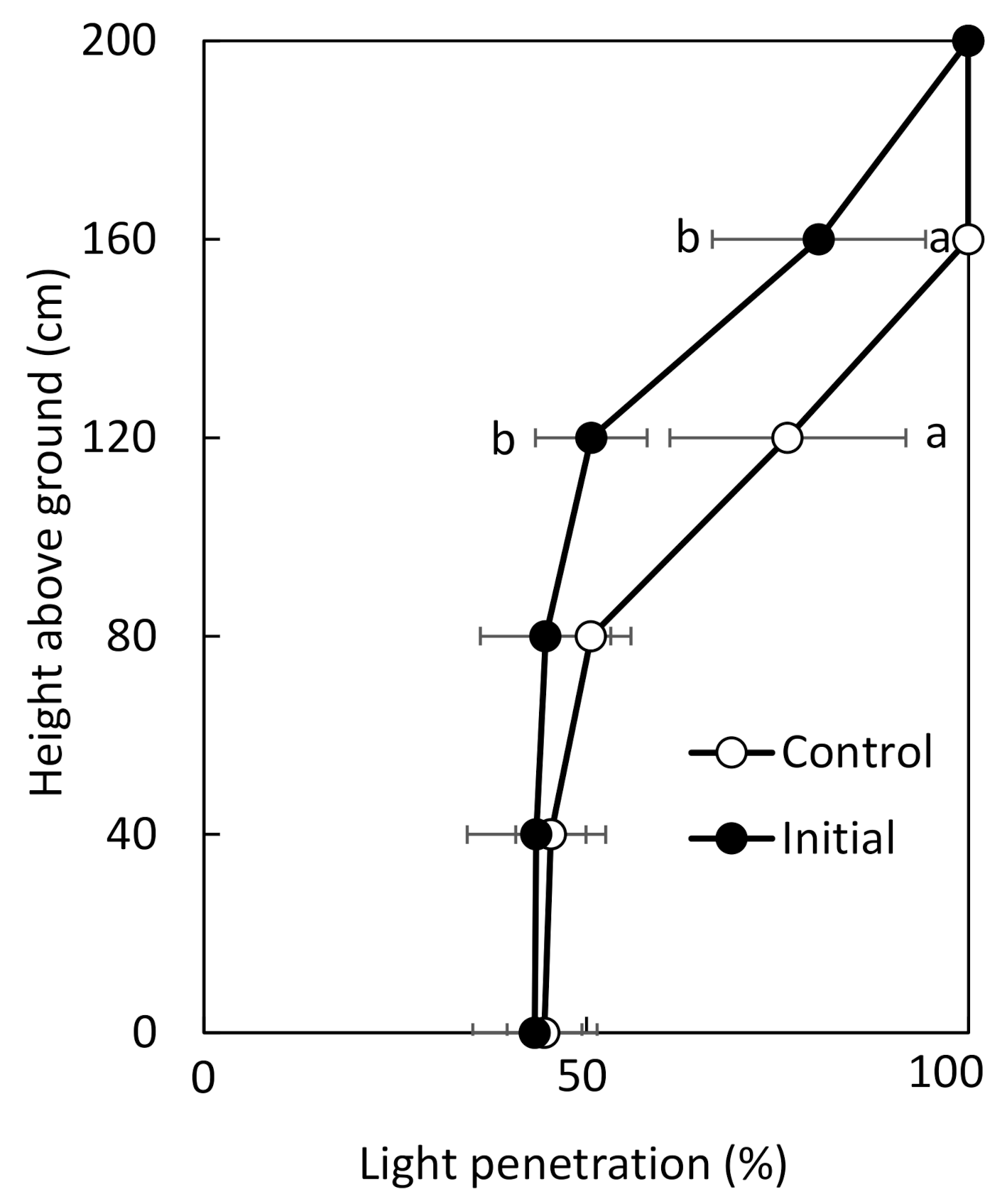
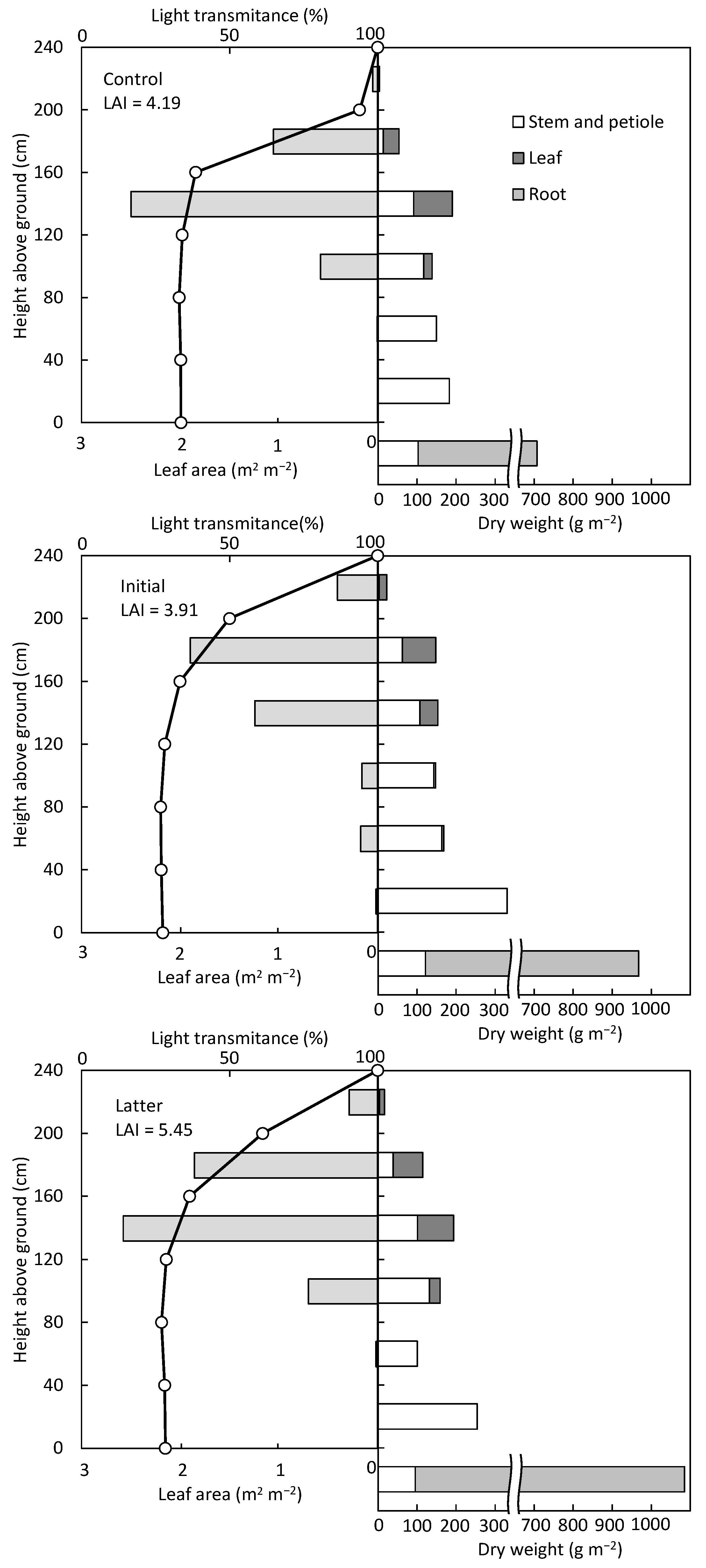
According to the measurements of light transmittance in the crop community in August, the height of the top layer was different between the control and initial fertilization treatments (
Figure 7); light transmittance dropped sharply to 40% below the second layer and did not change between the third layer and ground surface. However, there were differences in the layer heights and light transmittance decreased rapidly among all treatments in November. A sharp decrease in light transmittance of about 50% was observed in the second layer of the control and in the first layer of the initial fertilization plots. It was shown that only the leaves in the upper part of the canopy had sufficient sunlight. On the other hand, in the latter fertilization plots, light transmittance decreased by 38.8% and 24.6% at the first and second layers, respectively, indicating that the light reached the inside of the canopy. However, the maximum leaf area per layer was distributed in the third layer in the control and latter fertilization plots and in the second layer in the initial fertilization plot, below the layer where the transmittance was sharply reduced. Several research works have reported that LAI maximizes the yield by 2.5 to 3.5 times [
20,
21,
22]. The LAI of this study was between 3.91 (initial fertilization) and 5.45 (latter fertilization). This suggested that excess leaves may have grown in the community. In this study, the decrease in light transmission occurred in the first and second layers, which was common in both the August and November measurements. However, according to Santanoo et al. [
18], the decrease in light transmittance occurred in lower layers at 6 MAP than at 3 MAP, and the minimum light transmittance was smaller at 6 MAP, suggesting that the community was still growing after 3 MAP. This may be because the planting density of Santanoo et al. [
18] was lower than that of this study, and thus it may have taken longer to close the community.
A comparison of total dry matter weights revealed that both initial and latter fertilization had a positive effect on dry matter production and that the application time influenced DMD. The root DMD of the control and initial fertilization treatment were almost the same, 46.3 and 46.4%, respectively, and that of latter fertilization treatment was slightly higher at 53.8% (
Table 1). It is known that root DMD, an important factor related to yield, is greatly influenced by the cultivar, cultivation period, and environment. There are reports of root DMD of 30.1% to 31.6% [
23] and 49% to 56% [
24] for cultivation periods of around 180 days, so the root DMD in this study is not lower than that of other reports for similar cultivation periods. Minami et al. [
12] also conducted cassava cultivation at Kagoshima with almost the same cultivation period as this research for three years. However, the root DMD was lower than it was in this study, about 30%. Lian and Cock [
25] stated that although the roots and the above-ground part grow at the same time, the dry matter is preferentially distributed on the above-ground part until canopy development finishes. It is purported that the period of preferential distribution to the roots was short, and the harvest index remained low in Kagoshima, where the growing period is limited. The plant density of Minami et al. [
12] was 1.23 to 1.30 hill m
−2, lower than in this study, which imitated Tokunoshima’s plant density, which is a cassava producing area in Kagoshima. As a result, we believe that the root DMD became low because it took a long time for the development of the plant community. Our first hypothesis was that the early enlargement of tuberous root leads high root DMD. However, the root DMD of initial fertilization treatment did not change but rather the stem DMD showed the higher tendency. These results suggest that although the initiation of tuberous formation was promoted by initial fertilization treatment, the dry matter accumulation in the stem before it was large and they were maintained until harvest. But in latter fertilization treatment, due to not only the increase of gs in October and November, but also the distribution of photosynthetic products focusing on root by fertilization in August, when the plant community was developed, a high DMD was observed. On the other hand, there is a report of 60–84% after ten months of planting [
10], suggesting that further improvement is possible if the cultivation period at Kagoshima can be extended. The DMR of tuberous roots was significantly higher at the initial fertilization treatment (32.5%) than in the other two treatments (26.5, 27.4%). It is suggested that starch accumulation may have started earlier at the initial fertilization treatment where the plant height increased faster, and the canopy was closed earlier. Since the dry matter weight and DMD of the stems are significantly higher at the initial fertilization treatment, fertilization at the initial growth stage, when starch accumulation has not started, increases the dry matter production, but the effect of the increasing yield may be limited due to increasing the distribution to the stem. The effects of growing period, cultivar, management, and precipitation during the growing period on root DMR have been reported [
26,
27], but most of the reports discussed the results of samples around 12 MAP. Variety is one of the important factors affecting root DMR, which has been reported to be a minimum of 27.49–31.9% and a maximum of 41.19–41.7% [
11,
28,
29]. The highest dry matter rate of root was 32.5% at initial fertilization in this study, and was low compared to other reports. Lian and Cock [
25] stated that tuberous root enlargement occurs after the development of above-ground parts. In this study, since the canopy was not fully developed until August, the actual tuberous root enlargement period may have been even shorter. In particular, the control and latter fertilization treatments, which lacked fertilizer in the early growth stage, delayed community development and the initiation of tuber hypertrophy, resulting in a decrease of tuber DMR (
Table 1). In addition, the latter fertilization resulted in significantly higher fresh yield than the other treatments due to the tuber enlargement caused by vigorous photosynthesis after fertilization, but the DMD of tuberous root was probably lower because there was not enough time to fill the enlarged sink capacity.
The summary of the mechanisms by which each fertilization treatment increased the root dry weight is as follows. Both the initial and latter fertilization increased total dry weight by promoting the gas exchange rate on the upper layer of the plant community, but there was no difference in the utilization of solar radiation associated with the change in productive structure in any treatments. The increase in root dry matter weight was achieved not only by increasing the gas exchange rate, but also by increasing root DMR due to promoting tuber formation in the initial fertilization. In case of the latter fertilization treatment, a high gas exchange rate and increasing DMD to the root contributed to the high root dry weight.
Although there are not many reports investigating cassava production in Kagoshima, the tuberous root dry weight of this study was 694 g–1087 g m
−2 (6.9–10.8 t ha
−1), larger than the data reported by Minami et al. [
12], whose yield ranged 2.30~5.24 t ha
−1. An important difference in the cultivation management of these studies was the plant density, which was almost twice as high in our study as in Minami et al.’s study [
12]. This suggests that the high planting density may have caused early canopy closure and enlargement of the tuberous root. It is difficult to compare yields at different regions because cassava yield is easily affected by cultivation period [
10,
30], genetic background [
31], and cultivation methods [
27]. However, we selected studies with a cultivation period of about six months and compared them with the yield data of this study. Howeler and Cadavid [
15] and Connor et al. [
23] compared and evaluated the yield of two cassava cultivars, M Mex59 and M Col22, and their yield ranged 2.0–3.0 t ha
−1 and 4.0–6.2 t ha
−1, respectively. Hammer et al. [
30], who tracked the yield with four different planting periods, reported a yield of 7.0–13.0 t ha
−1, if the low temperature period was not encountered immediately after planting. Lenis et al. [
31] grew 1350 plants of cassava clone which had different two type of leaf retention ability and reported that the clone group with leaf retention ability (6.95–9.16 t ha
−1) had a significantly larger yield than the clone group without it (5.10–7.09 t ha
−1) in both March and May. As for the dry matter production in this study, it was clarified that the productivity of cassava within 6 MAP in Kagoshima is comparable to that in the tropical region. On the other hand, it is known that tuberous root dry matter continues to increase after six months and it increased by 2–3 times at 10–20 MAP [
15,
23,
30]. Irikura et al. [
7] reported that the temperature limit of cassava growth was 18–20 °C daily mean temperature and observed that cassava growth stopped at Kagoshima after mid-November, when the daily mean temperature dipped below 15 °C [
12]. Thus, Minami et al. [
12] concluded the period in which cassava can grow in Kagoshima is around eight months, from May to November. Although the period that cassava cannot grow is one of the limiting factors for the consideration to introduce various cultivation methods into Kagoshima, Singh et al. [
32] conducted a cultivation experiment in Northern India, which has low temperatures in the winter and hot weather in the summer and reportedly obtained 7.5–12.3 t ha
−1 yield in terms of dry weight in the eight-month cultivation period. Although the cultivation results, techniques, and knowledge accumulated in Kagoshima are insufficient, the fact that a certain yield was obtained in that study suggests that there is much room for yield improvement by accumulating experience and introducing new techniques and knowledge into cassava production. As the cassava cultivation area expands, it is estimated that the use of the edge area in which environment is not favorable for cultivating cassava will increase. All regions have their own problems; therefore, the accumulation of knowledge and cultivation examples in these regions, including in this study, is expected to contribute to maximizing the yield in similar areas.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}