1. Introduction
Strawberry (
Fragaria × ananassa Duch.) is the most economically important soft fruit in the world. The global production of strawberries has risen almost 30% between 2009 and 2019, making it the second-largest berry fruit, after grape (
Vitis vinifera L.), in terms of production [
1]. Strawberries are known for their characteristic aroma and taste. They are also an excellent source of vitamin C and have considerable amounts of vitamin B9. The antioxidative properties of strawberry fruits are reported to be beneficial for blood sugar and heart health [
2,
3,
4].
In recent years, greenhouse strawberry production around the globe has gained importance over traditional soil culture. The advantages of greenhouse production over traditional cultivation include better cultivation practices, improved pest and disease management, and more efficient nutrient and irrigation use resulting in higher yield. In addition to these advantages, the short cultivation period and suitability for small family farms are the key reasons for the interest in soilless strawberry production [
5]. Peat predominates as a commercial substrate for greenhouse strawberry production. However, due to extensive utilization, peat resources are at the edge of depletion [
6,
7,
8]. Considering its non-renewability, increasing cost, future availability, and environmental sustainability, growers around the globe need a high-quality, renewable, and sustainable substitute [
8]. At the same time, many reports also stress the need for finding potential peat-free growing media [
9,
10,
11].
Increasing environmental concerns and need for effective utilisation and disposal of agro-waste to achieve sustainability has encouraged investigations into utilizing accumulating potential agro-industrial residues in horticulture. Some studies have reported the influence of agro-waste-based substrates as a substitute for peat in strawberry production [
12,
13,
14,
15,
16]. Among many agro-wastes, the possibility of utilizing spent mushroom substrate (SMS) as a potential substitute can be beneficial. This agro-waste has a high organic composition, is readily available, and the cost is negligible [
17,
18,
19,
20]. SMS is the residual material left over after commercial mushroom production. The substrate obtained immediately after mushroom production is fresh-SMS (F-SMS) or SMS, and the same material after further decomposition or weathering from 3–24 months is called weathered-SMS (W-SMS) or spent mushroom compost (SMC) [
21]. Mushroom production can be considered as a non-sustainable agriculture activity due to the accumulation of an enormous amount of SMS and its limited re-use. Global mushroom production has passed ten million tons [
1]. Approximately five kilograms of SMS is generated for each kilogram of mushrooms produced [
18,
22,
23]. On this account, the annual SMS generated from the global mushroom industry is estimated to be more than 50 million tons. SMSs generated from mushroom enterprises in large quantities are often burnt, discarded, or simply thrown away, which is neither economical nor environmentally safe [
24]. Increasing mushroom production in recent years and the expected increase in the future will lead to a significant accumulation of SMS. The generated amount of SMS can pose potential environmental threats if not properly disposed of or effectively utilised.
Researchers over decades reported that SMC can be utilised for agricultural and horticultural purposes [
25], as a soil conditioner [
26,
27,
28], as nursery media [
7,
29,
30], and as a soilless growing medium [
31,
32]. On the contrary, fresh SMS is recommended only for use after further decomposition/weathering process ranging from 3–24 months [
21,
29,
33]. It has been reported that even after 24 months of passive weathering, SMS can still release a significant amount of soluble solids [
21], and the leachates from SMS would significantly increase the salt content of underlying soil and groundwater [
21,
34]. Fresh SMS has high EC due to excess accumulation of salts during mushroom cultivation and unfavourable pH, which are the major limiting factors for its immediate use and hence using fresh SMS is not recommended [
21,
35,
36]. The unfavourable pH and EC of growing media may negatively influence overall plant development [
37,
38] and alter photosynthetic processes due to stress [
39,
40,
41,
42].
In recent years, various techniques have been developed to indicate stress processes in plants. The most promising and reliable results have been obtained in processes that analyse photosynthesis, which is closely associated with plant performance and yield [
43,
44,
45]. The decline in photosynthetic activity, either directly or indirectly due to various abiotic stress factors may largely influence the overall performance and yield of the plant [
44,
46]. To study the impact of different stress factors on photosynthesis, chlorophyll
a fluorescence has become a popular approach [
44,
47,
48], including salt stress [
46] and nutrient deficiencies [
49]. The stress-induced changes on the Photosystem II (PSII) electron acceptor are well reflected in the values of integrative OJIP-test parameters, known as Performance Indices (PIs). The OJIP-test has been successfully used to understand the influence of several stress factors in plants [
44,
50,
51].
The F
v/F
0 value defines the maximum quantum efficiency of PSII photochemistry and is the most common indicator of chlorophyll fluorescence transient on plant leaves under stress conditions [
52]. F
v/F
m indicates the maximum quantum yield of PSII photochemistry, whereas PI(
abs) is a photosynthetic parameter that takes several different phenomena related to PSII photochemistry into consideration [
53]. Vegetation indices (VIs) are radiative transfer-based methods, with some mathematical combination or transformation of spectral bands that accentuate the spectral properties of plants. The expressed values of different VIs, including the Normalized Difference Vegetation Index (NDVI), Photochemical Reflectance Index (PRI), and Modified Chlorophyll Absorption in Reflectance Index (MCARI) help to analyse crop growth, vigour, and several other vegetation properties including biomass and chlorophyll content [
54]. Vegetation indices are also reported to be a reliable indicator of plant health [
55,
56] and abiotic stress in plants [
57,
58].
The scientific information supporting the utilisation of SMC (weathered SMS) is evident. However, the process of further decomposition can be considered laborious, time-consuming and the leachates released during this process can alter underlying soil and water chemistry, which may lead to various environmental hazards. Hence, this study was aimed to come up with an easy, effective, immediate, and cheap utilisation of fresh SMS, which may be environmentally and economically beneficial.
Strawberry greenhouse production is progressively gaining importance around the globe [
13,
59,
60]. However, the yield and overall performance of strawberries largely depends on the substrate [
12,
13,
14,
15,
16,
59,
60]. Considering the growing popularity of soilless strawberry cultivation, which relies mainly on peat, and the increasing demand for peat-reduced media, the development of a peat alternative is of great importance.
The present investigation was designed to study the possible utilization of fresh SMS as a renewable, easily available, comparatively cheap and sustainable alternative to non-renewable, relatively expensive, and non-sustainable peat in soilless strawberry production. In the present study, fresh SMS was obtained after the cultivation of white button mushroom—Agaricus bisporus (J.E. Lange), shiitake—Lentinus edodes (Berk.) and oyster mushroom—Pleurotus ostreatus (Jacq.). The obtained SMSs were used as a peat substitute in 15 and 25% (v/v) for soilless strawberry cv. ‘Honeoye’ cultivation in an unheated plastic tunnel. The investigation was carried out with three objectives: to evaluate the suitability of fresh SMS as a peat substitute, to study the strawberry growth and yield performances on SMS based substrates, and to determine selected abiotic stress responses and vegetation indices.
4. Discussion
The high value of EC is one of the most important factors limiting the use of SMS as a growing medium [
34,
70,
71]. Many researchers have reported that EC values of
Agaricus-SMS,
Lentinus-SMS, and
Pleurotus-SMS range between 0.58–10.70 mS∙cm
−1 [
71,
72,
73], 1.96 mS∙cm
−1, and 0.89–4.01 mS∙cm
−1 [
74], respectively. However, these values may vary as the composition of SMS greatly depends on the mushroom species cultivated [
75]. The optimum EC for soilless strawberry production is reported to be 1.4–2.5 [
76,
77], whereas the EC of 2.5 mS∙cm
−1 is considered to be an upper limit [
77,
78]. Strawberries are a highly salt-sensitive crop [
79,
80]. A high salt concentration of the substrate can negatively affect overall crop performance [
40,
81,
82,
83]. Salinity stress in strawberries both directly or indirectly affects chlorophylls and carotenoids, which may lead to low productivity [
84].
A high level of EC restricts the use of fresh SMS. In the present study, none of the substrate EC values before the experiment were reported to be saline, as the values ranged between 0.11–1.28 mS∙cm
−1 and nearly optimum EC for strawberry soilless culture was achieved in A-15 and A-25 with A-SMS (
Table 6). The chemical analysis of substrates after the experiment revealed that the EC values increased among all tested substrates (
Table 6). Substrates supplemented with A-SMS in 15 and 25% showed higher EC values of 3.20 and 3.69 mS∙cm
−1, respectively. Bryla and Scagel [
80] reported that to achieve optimum growth of strawberry cv. ‘Honeoye’, the EC of the growing media should be maintained at ≤1.3 mS∙cm
−1 during the early stage, and at ≤3.4 mS∙cm
−1 once the plants have matured.
The unfavourable value of pH is another factor that limits the usage of SMS as a growing medium. It has been reported that strawberries can perform well in acidic soil pHs of 4.6–6.5 [
85,
86]. In the present study, the pH values of 100% A-SMS, L-SMS, and P-SMS were outside of this optimal pH range (
Table 5). Optimum pH levels were found in L-15 and L-25 with L-SMS when compared to other substrates.
The EC and pH values of all substrates studied in the present investigation were lowered to nearly peat values when fresh A-SMS, L-SMS, and P-SMS were mixed with peat in 15 and 25% (
v/
v) (
Table 6). These results are in agreement with Eudoxie and Alexander [
36], who concluded that mixing SMS with peat (50:50%) nearly neutralised the limiting nature of the SMS.
Results of the study showed that strawberry plants grown in A-25 and L-25 had the highest shoot dry weight, root dry weight, total plant dry weight, and leaf area when compared to peat. In previous studies, promising results with agro-waste-based substrates for strawberry cultivation were reported. In the study by Altieri et al. [
12], olive mill waste mixture (OMWM) was found to be an effective and cheap alternative to peat. Kuisma et al. [
14] reported using ground reed canary grass (
Phalaris arundinacea L.) as a peat or coir substitute (50%). Depardieu et al. [
16] revealed that a peat–sawdust mixture and aged bark were found to be promising alternative substrates to coconut coir dust for strawberry cultivation. However, others reported peat- and coconut-coir-based substrates were found to be the best substrates for strawberries [
13,
87,
88,
89,
90].
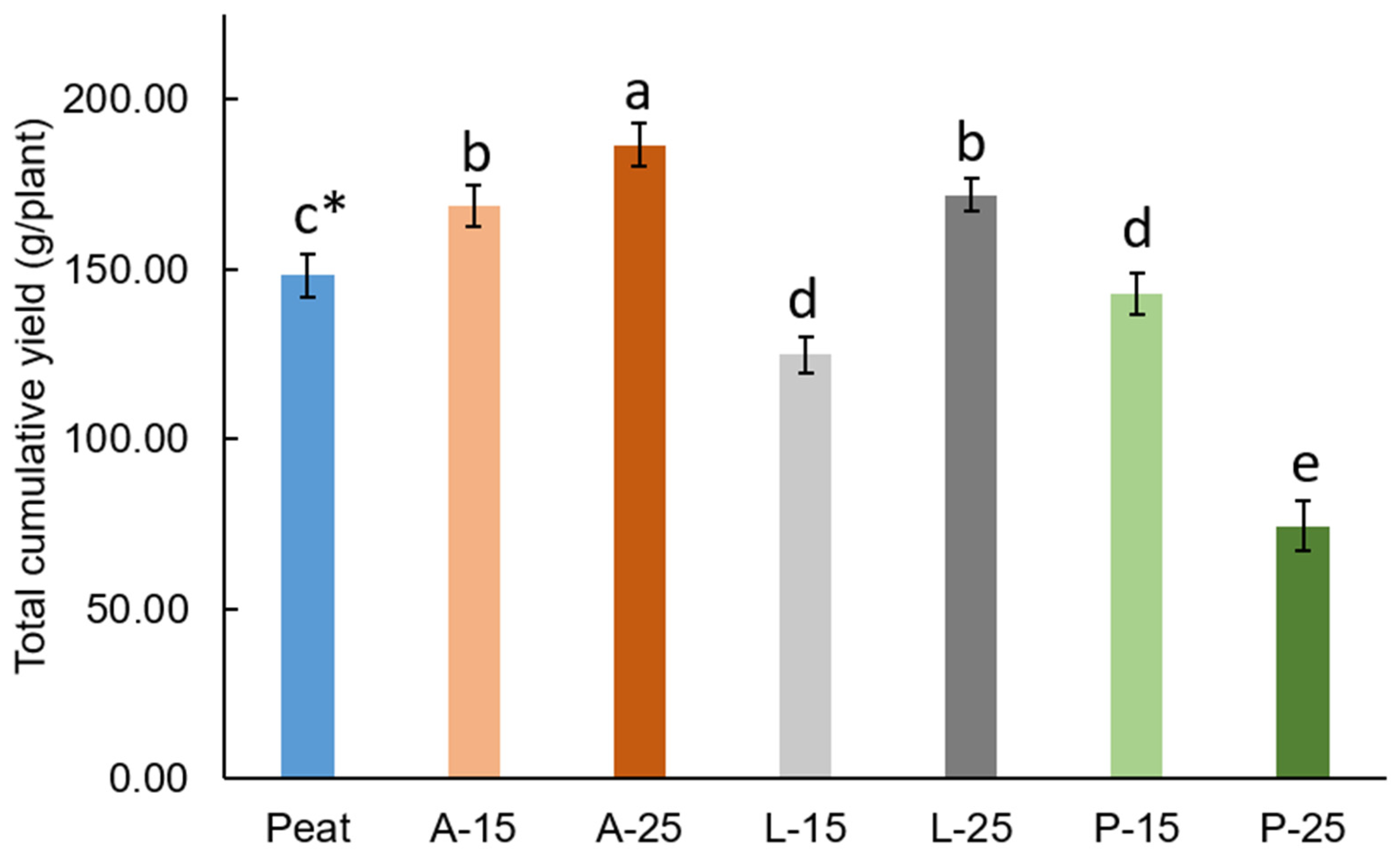
The plants grown in A-SMS (A-15 and A-25) had significantly higher total cumulative and marketable yield than plants grown in peat (
Figure 1 and
Table 7). The initial nutrient concentration of A-SMS, especially macronutrients including N, P, K, as well as secondary elements like Ca and Mg, was observed to be higher when compared to other substrates, which may have contributed to higher yield and better morphological parameters. As reported by Medina et al. [
35] and Benito et al. [
91], SMS has been proven to increase the nutrient status of growing media. The amount of N-NH
4 in the growing media, which is easily available for plant uptake, can also benefit plant growth and yield [
92].
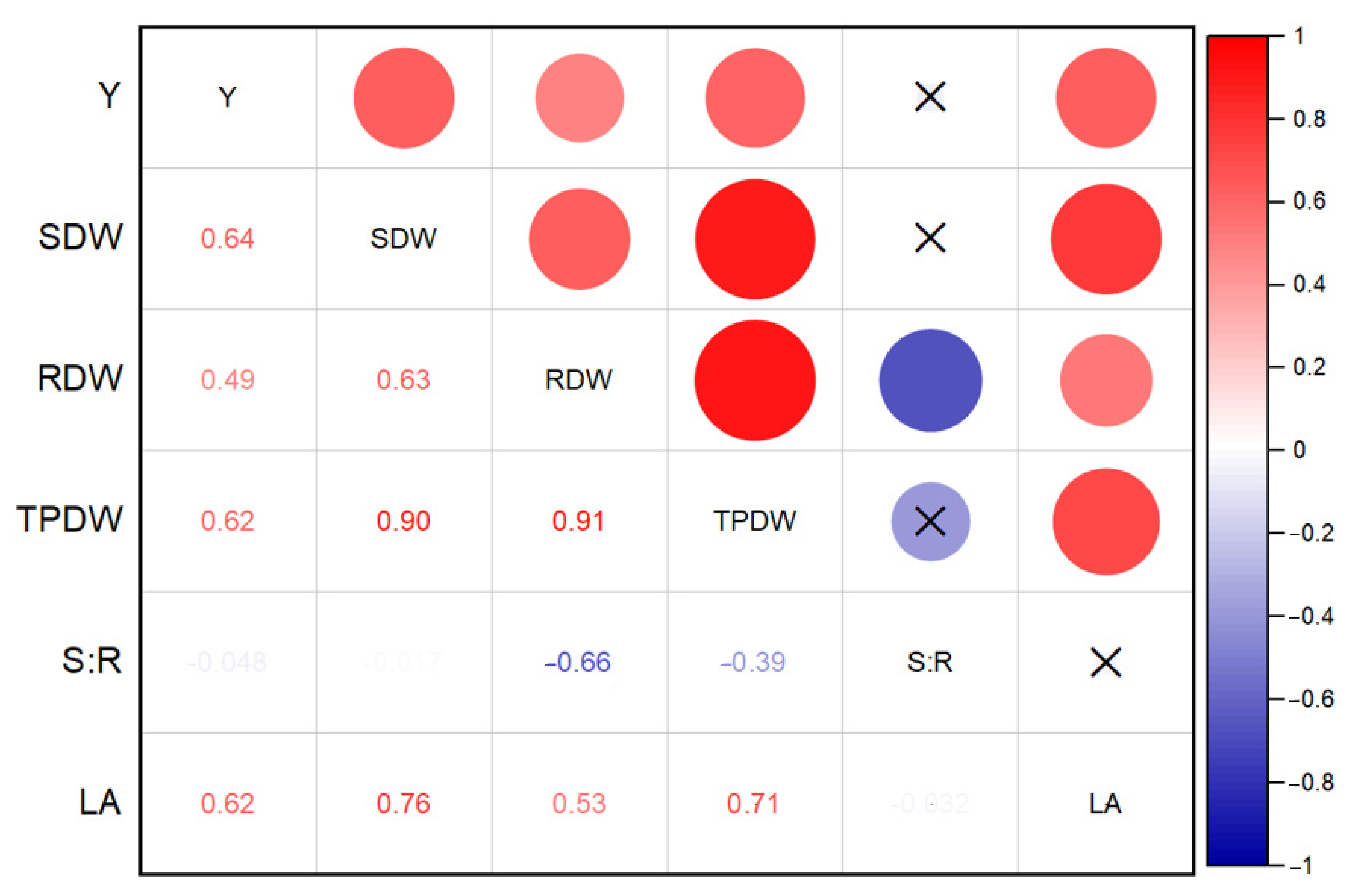
Based on Cohen [
93], the correlation coefficient values obtained in the present study suggested a strong relationship. The observed ‘
r’ values among marketable yield and tested morphological parameters except for root to shoot ratio are positively correlated (0.445–0.552). Correlation values greater than 0.50 indicates that the relationship is strong. The observed correlation results are in agreement with Adak et al. [
94], who also reported a positive correlation between marketable yield to root dry weight and marketable yield to shoot dry weights.
Fluorescence is a highly sensitive photosynthetic plant retraction analysis that can detect any change in the overall bioenergy status of a plant [
95]. Unfavourable pH and EC values can significantly influence strawberry growth and overall performance, which can be explained by measured PIs and VIs. Alkalinity stress due to high pH may destroy the photosynthetic activity of plants [
34,
42]. Similar results were reported under salinity stress conditions [
41,
96]. Alkalinity stress lowered the maximal quantum yield of PSII photochemistry (F
v/F
m) in strawberry cv. ‘Camarosa’ [
37,
42]. Chlorophyll
a fluorescence parameters might be a useful indicator for diagnosing the occurrence of salt and alkaline stress in strawberries [
42,
96].
Among several photosynthetic parameters measured in the study, F
v/F
0, F
v/F
m, and PI(
abs) were selected to be presented in this article due to their proven sensitivity in identifying different abiotic stress responses in plants [
51,
52,
97,
98,
99,
100]. Angelini et al. [
98] reported that the maximum photochemical yield of PSII (F
v/F
m) is a reliable indicator of the photochemical activity of the photosynthetic apparatus. For the majority of plants at the stage of full development and under optimal growing conditions, the value of F
v/F
m was found to be around 0.83. Significant differences in F
v/F
m values (0.77–0.81) among three substrates studied in soilless strawberry production [
88]. In wild strawberries (
Fragaria vesca L.), substrate salinity caused a significant decrease in the F
v/F
m (0.71–0.74) and F
v/F
0 (2.61–2.90) values [
93]. Shamsabad et al. [
42] observed that the F
v/F
m (0.39–0.82), F
v/F
0 (0.37–4.49), and PI
abs (0.22–14.6) parameters declined among all six cultivars with increasing sodium bicarbonate (alkalinity).
In turn, the large range of observed values among these studies and a decrease in these parameters indicated that the plant exposed to stress factors can alter PSII functions. The structural damage of thylakoids and decrease in efficiency of water fission reaction may lead to slow relaxing quenching processes and a reduction in the efficiency of electron transport [
101,
102,
103]. Changes in the content of photosynthetic pigments are also dependent on the tolerance of the plants to the salinity of the substrate, i.e., their genotype [
104,
105]. However, strawberry cv. ‘Honeoye’ used in the study is a salt-sensitive cultivar [
80].
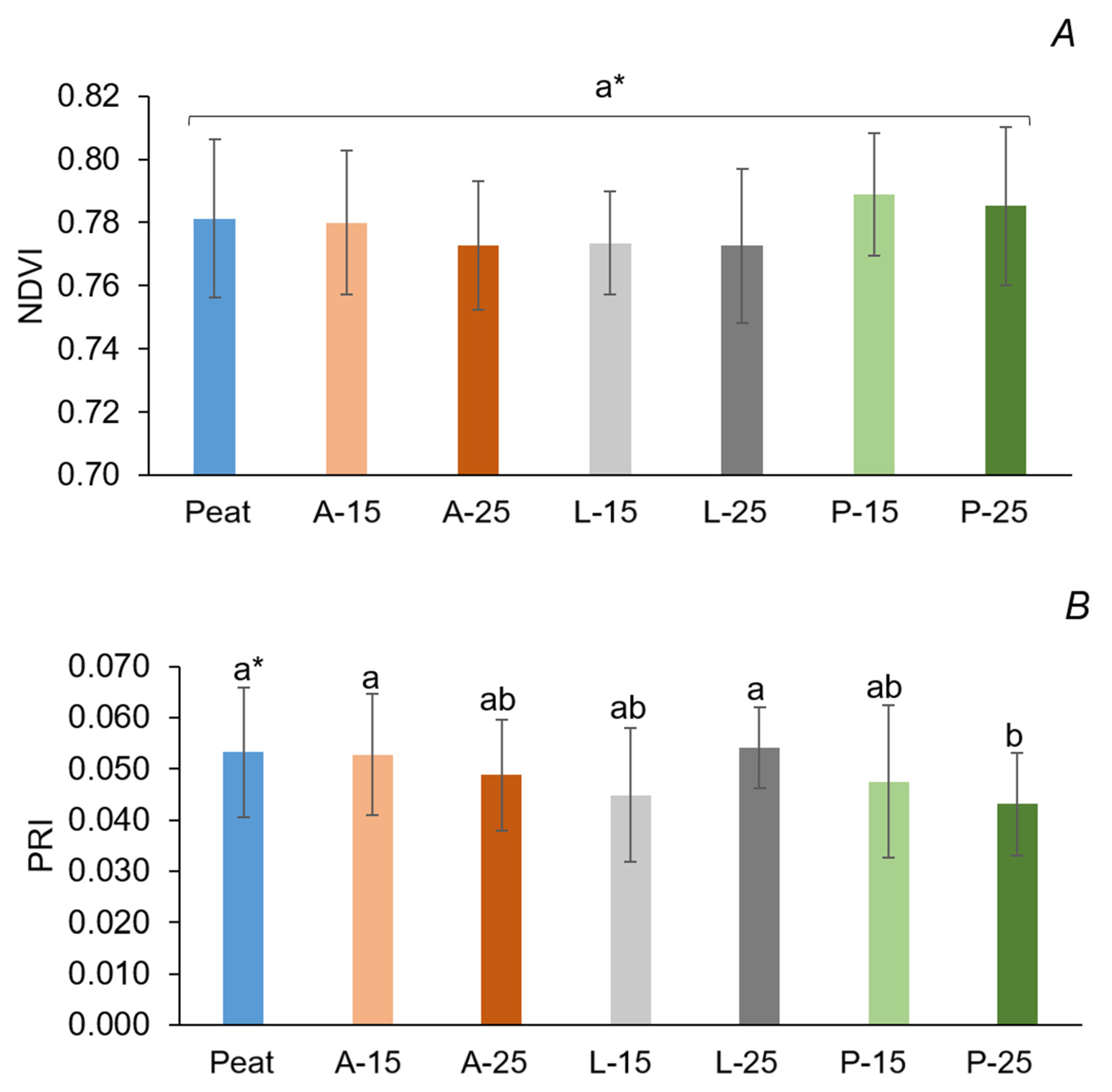
In the present investigation, the F
v/F
m values (0.82–0.83) were found to be in the optimum range [
98,
106]. The F
v/F
0 and PI
(abs) values were observed to be in the range of 4.52–4.93 and 2.91–3.60, respectively. The observed values suggest that the plants were not under stress and the photosynthetic activity did not differ among plants grown in substrates with A-SMS, L-SMS, and P-SMS substitutes when compared to standard peat. This is also supported by NDVI values (0.77–0.79), which were found to be in the range indicating normal vegetation [
107]. NDVI is a spectral vegetative index widely used in the determination of plant N status and can further explain the difference in yield [
108,
109,
110]. The strawberry nitrogen content of leaves and marketable yield were found to be positively correlated to the NDVI values, as reported by Li et al. [
107] and España-Boquera et al. [
111].
PRI is a quantitative measure of reflectance change at 531 nm, which indicates changes in the state of xanthophyll cycles and is strongly related to photosynthetic light use efficiency [
112]. The differences in PRI values among substrates (0.043–0.054) in the present study were probably due to the involvement of multiple processes with separate time constants affecting reflectance and fluorescence to different degrees [
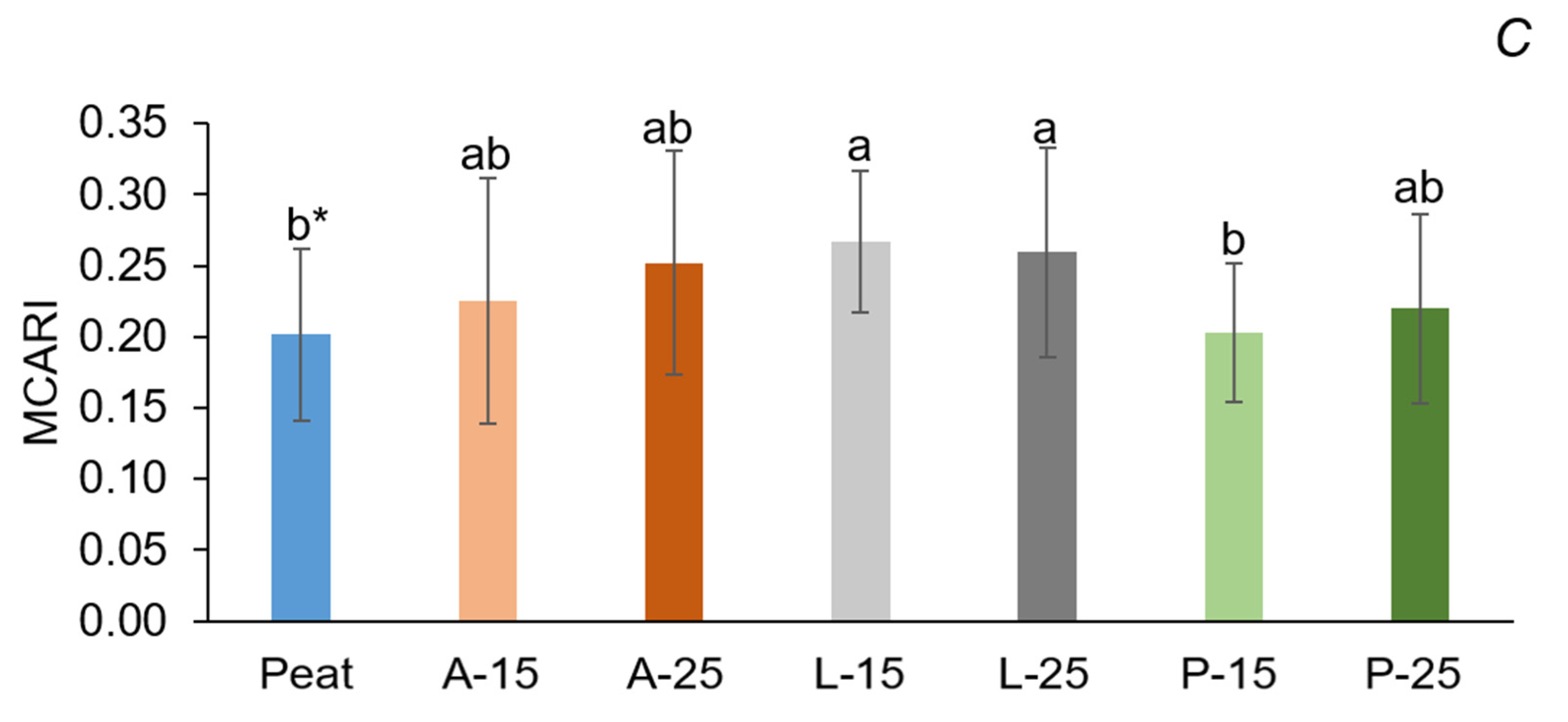
113]. Significant differences among substrates concerning MCARI (0.20–0.27) were probably due to differences in leaf chlorophyll concentrations [
68]. The numerical difference among PIs and VIs in the present study indicates that the data points are spread out over a small range of values. These values further demonstrate that the plants were not influenced by stress due to varying pH and EC levels of substrates.
Several reports have demonstrated that the type of growing media used in soilless strawberry production influences yield parameters [
12,
13,
14,
94,
114,
115,
116,
117]. The obtained results in the present investigation also suggest that substrates largely influence marketable and total yield, as well as morphological parameters of strawberries.
The results of the present investigation are in line with Atikmen et al. [
33], who reported that fresh mushroom substrate can be used as a substitute for peat primarily at 12.5% and 25% in greenhouse chrysanthemum production. In contrast, many reports recommend using SMS only after a weathering process in lower concentrations (5–50%) for better plant growth [
29,
36,
118,
119]. Some authors even claim that weathering alone is not enough to reduce salinity to a satisfactory level, recommending further leaching of weathered SMS for short times (up to 20 days) to remove excess salt (EC) before using it as a soilless substitute in crop production [
120,
121,
122].









{kind=link}
{kind=link}
{kind=link}
{kind=link}