
Genome-Wide Association Mapping of Freezing Tolerance Loci in Canola (Brassica napus L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Phenotype Canola for Freezing Tolerance
2.3. Genome-Wide Association Mapping of Winter Hardiness
2.4. Selection of Candidate Genes
3. Results
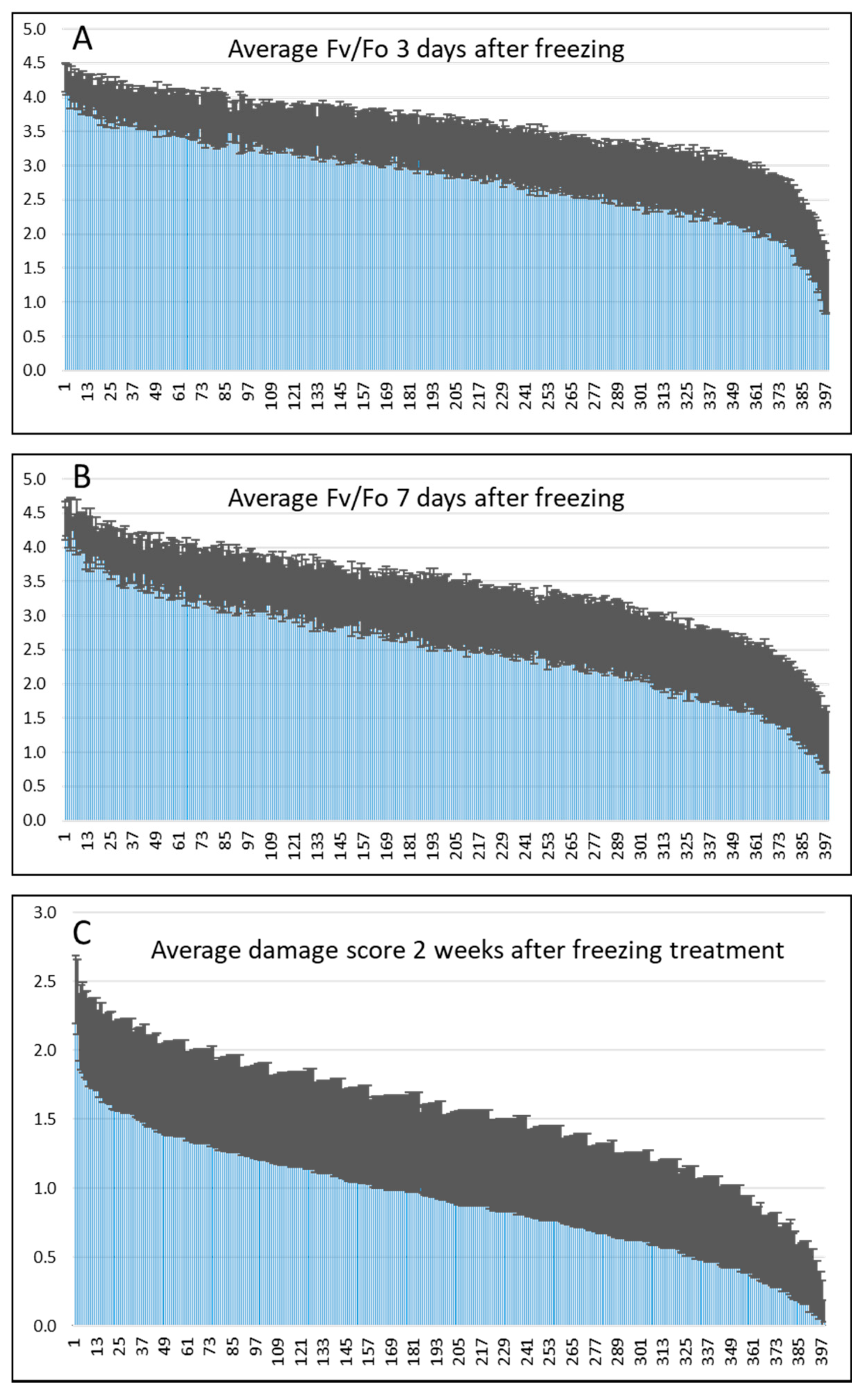
3.1. Phenotyping Canola Accessions Freezing Treatments
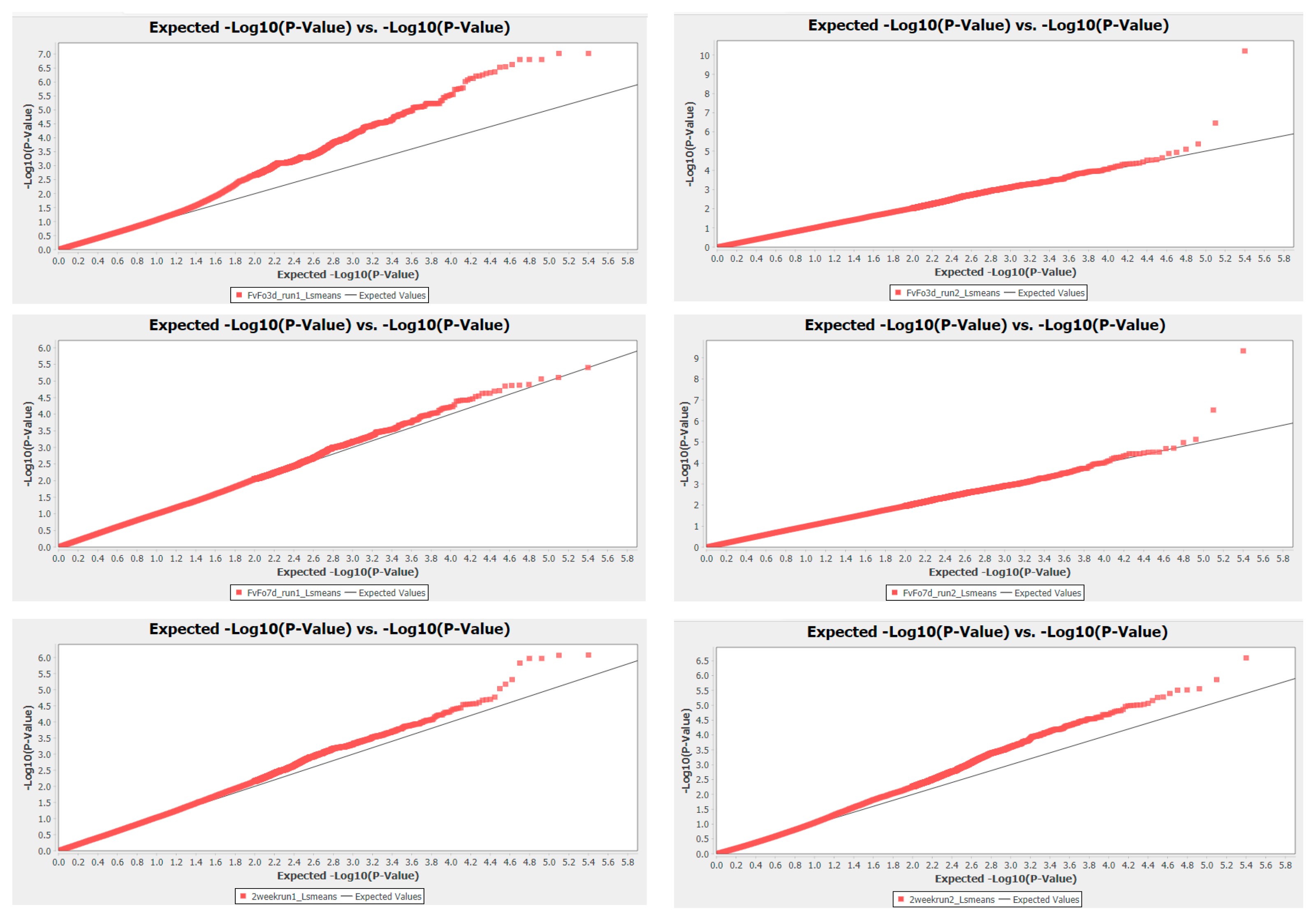
3.2. Assessing Model Fit
3.3. Loci Associated with Freezing Tolerance Based on GWAS
3.4. Underlying Candidate Genes Associated with Freezing Tolerance
4. Discussion
4.1. Comparison to Other Association Mapping Studies Identify Additional Regions of Interest
4.2. Phenotypic Measures of Visual Damage and Photosynthetic Damage are Correlated
4.3. Candidate Genes Associated with Freezing Tolerance
4.4. Other Genes of Interest
4.5. Photosynthetic and Light Signaling Processes Are Correlated with Cold Tolerance
4.6. CBF and Freezing Tolerance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bushong, J.; Lofton, J.; Sanders, H.; Stamm, M.; Arnall, B.; Ciampitti, I.; Damicone, J.; DeVuyst, E.; Epplin, F.; Giles, K.; et al. Great Plains Canola Production Handbook; MF-2734; Kansas Agricultural Experiment Station and Cooperative Extension Service: Manhattan, KS, USA, 2018. [Google Scholar]
- Stamm, M.; Dooley, S.; Aiken, R.; Angadi, S.; Begna, S.; Baldwin, B.; Morrison, J.; Bell, J.; Blair, M.; Braden, I.; et al. National Winter Canola Variety Trial; Contribution No. 19-252-S Kansas Agricultural Experiment Station; Kansas State University: Manhattan, KS, USA, 2019; pp. 1–35. [Google Scholar]
- Gusta, L.V.; Wisniewski, M. Understanding plant cold hardiness: An opinion. Physiol. Plant. 2013, 147, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Rife, C.L.; Zeinali, H. Cold tolerance in oilseed rape over varying acclimation durations. Crop Sci. 2003, 43, 96–100. [Google Scholar] [CrossRef]
- Miura, K.; Furumoto, T. Cold signaling and cold response in plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hincha, D.K.; Zuther, E. Introduction: Plant cold acclimation and freezing tolerance. Methods Mol. Biol. 2014, 1166, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Narusaka, M.; Abe, H.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Carninci, P.; Hayashizaki, Y.; Shinozaki, K. Monitoring the expression pattern of the 1300 Arabidopsis genes under drought and cold stresses by using full length cDNA microarrays. Plant Cell 2001, 13, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef] [Green Version]
- Kreps, J.A.; Wu, Y.; Chang, H.S.; Zhu, T.; Wang, X.; Harper, J.F. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol. 2002, 130, 2129–2141. [Google Scholar] [CrossRef] [Green Version]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fujita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold, and high-salinity stresses using a full-length cDNA microarray. Plant J. 2002, 31, 279–292. [Google Scholar] [CrossRef]
- Lee, B.H.; Henderson, D.A.; Zhu, J.K. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhang, Z.; Xie, S.; Si, T.; Li, Y.; Zhu, J.K. Mutational evidence for the critical role of CBF transcription factors in cold acclimation in Arabidopsis. Plant Physiol. 2016, 171, 2744–2759. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dang, P.; Liu, L.; He, C. Cold acclimation by the CBF–COR pathway in a changing climate: Lessons from Arabidopsis thaliana. Plant Cell Rep. 2019, 38, 511–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Gene regulation during cold acclimation in plants. Physiol. Plant. 2006, 126, 52–61. [Google Scholar] [CrossRef]
- Park, S.; Lee, C.M.; Doherty, C.J.; Gilmour, S.J.; Kim, Y.; Thomashow, M.F. Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network. Plant J. 2015, 82, 193–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, S.G.; Cook, D.; Thomashow, M.F. Low temperature induction of Arabidopsis CBF1, 2, and 3 is gated by the circadian clock. Plant Physiol. 2005, 137, 961–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieniawska, Z.; Espinoza, C.; Schlereth, A.; Sulpice, R.; Hincha, D.K.; Hannah, M.A. Disruption of the Arabidopsis circadian clock is responsible for extensive variation in the cold-responsive transcriptome. Plant Physiol. 2008, 147, 263–279. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, C.; Degenkolbe, T.; Caldana, C.; Zuther, E.; Leisse, A.; Willmitzer, L.; Hincha, D.K.; Hannah, M.A. Interaction with diurnal and circadian regulation results in dynamic metabolic and transcriptional changes during cold acclimation in Arabidopsis. PLoS ONE 2010, 5, e14101. [Google Scholar] [CrossRef]
- Gray, G.R.; Chauvin, L.P.; Sarhan, F.; Huner, N.P.A. Cold acclimation and freezing tolerance: A complex interaction of light and temperature. Plant Physiol. 1997, 114, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Wanner, L.A.; Junttila, O. Cold-induced freezing tolerance in Arabidopsis. Plant Physiol. 1999, 120, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Cessna, S.; Demmig-Adams, B.; Adams, W.W., III. Exploring photosynthesis and plant stress using inexpensive chlorophyll fluorometers. J. Nat. Resour. Life Sci. Educ. 2010, 39, 22–30. [Google Scholar] [CrossRef]
- Gesch, R.W.; Heilman, J.L. Responses of photosynthesis and phosphorylation of the light-harvesting complex of PSII to chilling temperature in ecologically divergent cultivars of rice. Environ. Exp. Bot. 1999, 41, 257–266. [Google Scholar] [CrossRef]
- Opti-Sciences. Os30p + Chlorophyll Fluorometer. Available online: http://www.surechem.com.my/upload/products/doc/doc_1839_181004023717.pdf (accessed on 29 December 2020).
- Gusta, L.V.; Wisniewski, M.; Nesbitt, N.T.; Gusta, M.L. The effect of water, sugars, and proteins on the pattern of ice nucleation and propagation in acclimated and non-acclimated canola leaves. Plant Physiol. 2004, 135, 1642–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananga, A.O.; Cebert, E.; Ochieng, J.W.; Kumar, S.; Kambiranda, K.; Vasanthaiah, H.; Tsolova, V.; Senwo, Z.; Konan, K.; Anike, F.; et al. Prospects for Transgenic and Molecular Breeding for Cold Tolerance in Canola (Brassica napus L.). In Oilseeds; Akpan, U.G., Ed.; InTech: Rijeka, Crotia, 2012; pp. 1–32. Available online: http://www.intechopen.com/books/oilseeds/prospects-for-transgenic-and-molecular-breeding-for-cold-tolerance-in-canola-brassica-napus-l- (accessed on 26 January 2021).
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Feng, Q.; Qian, Q.; Zhao, Q.; Wang, L.; Wang, A.; Guan, J.; Fan, D.; Weng, Q.; Huang, T.; et al. High-throughput genotyping by whole-genome resequencing. Genome Res. 2009, 19, 1068–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 years of GWAS discovery: Biology, function, and translation. Am. J. Hum. Genet. 2017, 101, 5–22. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Torkamaneh, D.; Boyle, B.; Belzile, F. Efficient genome wide genotyping strategies and data integration in crop plants. Theor. Appl. Genet. 2018, 131, 499–511. [Google Scholar] [CrossRef]
- Bus, A.; Körber, N.; Snowdon, R.J.; Stich, B. Patterns of molecular variation in a species-wide germplasm set of Brassica napus. Theor. Appl. Genet. 2011, 123, 1413–1423. [Google Scholar] [CrossRef]
- Hatzig, S.V.; Frisch, M.; Breuer, F.; Nesi, N.; Ducournau, S.; Wagner, M.H.; Leckband, G.; Abbadi, A.; Snowdon, R.J. Genome-wide association mapping unravels the genetic control of seed germination and vigor in Brassica napus. Front. Plant Sci. 2015, 6, 221. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Mamidi, S.; Del Rio, L.; Ross, A.; Kadir, M.; Rahaman, M.; Arifuzzaman, M. Association mapping in Brassica napus (L.) accessions identifies a major QTL for blackleg disease resistance on chromosome A01. Mol. Breed. 2016, 36, 90. [Google Scholar] [CrossRef]
- Xu, L.; Hu, K.; Zhang, Z.; Guan, C.; Chen, S.; Hua, W.; Li, J.; Wen, J.; Yi, B.; Shen, J.; et al. Genome-wide association study reveals the genetic architecture of flowering time in rapeseed (Brassica napus L.). DNA Res. 2016, 23, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiessl, S.; Huettel, B.; Kuehn, D.; Reinhard, R.; Snowdon, R.J. Post-polyploidisation morphotype diversification associates with gene copy number variation. Sci. Rep. 2017, 7, 41845. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhou, C.; Zheng, W.; Mason, A.S.; Fan, S.; Wu, C.; Fu, D.; Huang, Y. Genome-wide SNP markers based on SLAF-seq uncover breeding traces in rapeseed (Brassica napus L.). Front. Plant Sci. 2017, 8, 648–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrucke, D.F.; Mamidi, S.; Rahman, M. Genome-wide association study for frost tolerance in canola (Brassica napus L.) under field conditions. J. Plant Biochem. Biotechnol. 2019, 28, 211–222. [Google Scholar] [CrossRef]
- Horvath, D.P.; Stamm, M.; Talukder, Z.I.; Fiedler, J.; Horvath, A.P.; Horvath, G.A.; Chao, W.S.; Anderson, J.V. A new diversity panel for winter rapeseed (Brassica napus L.) genome wide association studies. Agronomy 2020, 10, 2006. [Google Scholar] [CrossRef]
- Anderson, J.V.; Horvath, D.P.; Doğramacı, M.; Dorn, K.M.; Chao, W.S.; Watkin, E.E.; Hernandez, A.G.; Marks, M.D.; Gesch, R. Expression of FLOWERING LOCUS C and a frame shift mutation of this gene on Chromosome 20 differentiate a summer- and winter-annual biotype of Camelina sativa. Plant Direct 2018, 2, e00060. [Google Scholar] [CrossRef] [Green Version]
- Horvath, D.; Anderson, J.V.; Chao, W.S.; Zheng, P.; Buchwaldt, M.; Parkin, I.A.P.; Dorn, K. Genes associated with chloroplasts and hormone-signaling, and transcription factors other than CBFs are associated with differential survival after low temperature treatments of Camelina sativa biotypes. PLoS ONE 2019, 14, e0217692. [Google Scholar] [CrossRef]
- Mamidi, S.; Chikara, S.; Goos, R.J.; Hyten, D.L.; Annam, D. Genome-wide association analysis identifies candidate genes associated with iron deficiency chlorosis in soybean. Plant Genome 2011, 4, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Horvath, D.P.; Zhang, J.; Chao, W.S.; Mandal, A.; Rahman, M.; Anderson, J.V. Genome wide association studies and transcriptome changes during acclimation and deacclimation in divergent canola varieties. Int. J. Mol. Sci. 2020, 21, 9148. [Google Scholar] [CrossRef]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef]
- Cho, S.K.; Ryu, M.Y.; Kim, J.H.; Hong, J.S.; Oh, T.R.; Kim, W.T.; Yang, S.W. RING E3 ligases: Key regulatory elements are involved in abiotic stress responses in plants. BMB Rep. 2017, 50, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Peralta, D.A.; Araya, A.; Gomez-Casati, D.F.; Busi, M.V. Over-expression of SINAL7 increases biomass and drought tolerance, and also delays senescence in Arabidopsis. J. Biotechnol. 2018, 283, 11–21. [Google Scholar] [CrossRef]
- Lewandowska-Sabat, A.M.; Winge, P.; Fjellheim, S.; Dørum, G.; Bones, A.M.; Rognli, O.A. Genome wide transcriptional profiling of acclimation to photoperiod in high-latitude accessions of Arabidopsis thaliana. Plant Sci. 2012, 185–186, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Schenkman, J.B.; Jansson, I. The many roles of cytochrome b5. Pharmacol. Ther. 2003, 97, 139–152. [Google Scholar] [CrossRef]
- Gomès, E.; Jakobsen, M.K.; Axelsen, K.B.; Geisler, M.; Palmgren, M.G. Chilling tolerance in Arabidopsis involves ALA1, a member of a new family of putative aminophospholipid translocases. Plant Cell 2000, 12, 2441–2453. [Google Scholar] [CrossRef]
- Liu, T.; Guo, S.; Lian, Z.; Chen, F.; Yang, Y.; Chen, T.; Ling, X.; Liu, A.; Wang, R.; Zhang, B. A P4-ATPase gene GbPATP of cotton confers chilling tolerance in plants. Plant Cell Physiol. 2015, 56, 549–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belles-Boix, E.; Hamant, O.; Witiak, S.M.; Morin, H.; Traas, J.; Pautot, V. KNAT6: An Arabidopsis homeobox gene involved in meristem activity and organ separation. Plant Cell 2006, 18, 1900–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dao, T.T.; Linthorst, H.J.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Upadhyay, M.K.; Srivastava, A.K.; Abdelrahman, M.; Suprasanna, P.; Tran, L.P. Cellular and subcellular phosphate transport machinery in plants. Int. J. Mol. Sci. 2018, 19, 1914. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Zhao, N.; Qin, M.; Xu, A. Mapping of quantitative trait loci related to cold resistance in Brassica napus L. J. Plant Physiol. 2018, 231, 147–154. [Google Scholar] [CrossRef]
- Fiebelkorn, D.; Horvath, D.; Rahman, M. Genome-wide association study for electrolyte leakage in rapeseed/canola (Brassica napus L.). Mol. Breed. 2018, 38, 129. [Google Scholar] [CrossRef]
- Wrucke, D.F.; Talukder, Z.I.; Rahman, M. Genome-wide association study for frost tolerance in rapeseed/canola (Brassica napus) under simulating freezing conditions. Plant Breed. 2020, 139, 356–367. [Google Scholar] [CrossRef]
- Ishitani, M.; Xiong, L.; Lee, H.; Stevenson, B.; Zhu, J.K. HOS1, a genetic locus involved in cold-responsive gene expression in Arabidopsis. Plant Cell 1998, 10, 1151–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, J.Y.; Kim, W.T. Arabidopsis RING E3 ubiquitin ligase AtATL80 is negatively involved in phosphate mobilization and cold stress response in sufficient phosphate growth conditions. Biochem. Biophys. Res. Commun. 2015, 463, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Nylander, M.; Svensson, J.; Palva, E.T.; Welin, B. Stress-induced accumulation and tissue specific localization of dehydrins in Arabidopsis thaliana. Plant Mol. Biol. 2001, 45, 263–279. [Google Scholar] [CrossRef]
- Danyluk, J.D.; Houde, M.; Rassart, E.; Sarhan, F. Differential expression of a gene encoding an acidic dehydrin in chilling sensitive and freezing tolerant gramineae species. FEBS Lett. 1994, 344, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, Y.; Uemura, M. Mass spectrometric approach for identifying putative plasma membrane proteins of Arabidopsis leaves associated with cold acclimation. Plant J. 2003, 36, 141–154. [Google Scholar] [CrossRef]
- Puhakainen, T.; Hess, M.W.; Mäkelä, P.; Svensson, J.; Heino, P.; Palva, E.T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Mol. Biol. 2004, 54, 743–753. [Google Scholar] [CrossRef]
- Svensson, J.; Ismail, A.M.; Palva, E.T.; Close, T.J. Dehydrins. In Sensing, Signaling and Cell Adaptation; Storey, K.B., Storey, J.M., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2002; pp. 155–171. [Google Scholar]
- Desikan, R.; Hancock, J.T.; Neill, S.J. Oxidative stress signaling. In Plant Responses to Abiotic Stress; Hirt, H., Shinozaki, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 120–150. [Google Scholar] [CrossRef]
- Yamazaki, J.-Y.; Ohashi, A.; Hashimoto, Y.; Negishi, E.; Kumagai, S.; Kubo, T.; Oikawa, T.; Maruta, E.; Kamimura, Y. Effects of high light and low temperature during harsh winter on needle photodamage of Abies mariesii growing at the forest limit on Mt. Norikura in Central Japan. Plant Sci. 2003, 165, 257–264. [Google Scholar] [CrossRef]
- Baek, K.H.; Skinner, D.Z. Alteration of antioxidant enzyme gene expression during cold acclimation of near-isogenic wheat lines. Plant Sci. 2003, 165, 1221–1227. [Google Scholar] [CrossRef]
- Yong, Z.; Hao-Ru, T.; Ya, L. Variation in antioxidant enzyme activities of two strawberry cultivars with short-term low temperature stress. World J. Agric. Sci. 2008, 4, 458–462. [Google Scholar]
- Valizadeh-Kamran, R.; Toorchia, M.; Mogadama, M.; Mohammadic, H.; Pessaraklid, M. Effects of freeze and cold stress on certain physiological and biochemical traits in sensitive and tolerant barley (Hordeum vulgare) genotypes. J. Plant Nutr. 2018, 41, 102–111. [Google Scholar] [CrossRef]
- Hwang, Y.T.; Pelitire, S.M.; Henderson, M.P.; Andrews, D.W.; Dyer, J.M.; Mullen, R.T. Novel targeting signals mediate the sorting of different isoforms of the tail-anchored membrane protein cytochrome b5 to either endoplasmic reticulum or mitochondria. Plant Cell 2004, 16, 3002–3019. [Google Scholar] [CrossRef] [Green Version]
- Nagano, M.; Ihara-Ohori, Y.; Imai, H.; Inada, N.; Fujimoto, M.; Tsutsumi, N.; Uchimiya, H.; Kawai-Yamada, M. Functional association of cell death suppressor, Arabidopsis Bax inhibitor-1, with fatty acid 2-hydroxylation through cytochrome b5. Plant J. 2009, 58, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Tran, L.S.; Neelakandan, A.K.; Nguyen, H.T. Higher plant cytochrome b5 polypeptides modulate fatty acid desaturation. PLoS ONE 2012, 7, e31370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Thelen, J.J. ACYL-LIPID DESATURASE2 is required for chilling and freezing tolerance in Arabidopsis. Plant Cell 2013, 25, 1430–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.R.; Hincha, D.K. Flavonoids are determinants of freezing tolerance and cold acclimation in Arabidopsis thaliana. Sci. Rep. 2016, 6, 34027. [Google Scholar] [CrossRef]
- Zhu, W.; Miao, Q.; Sun, D.; Yang, G.; Wu, C.; Huang, J.; Zheng, C. The mitochondrial phosphate transporters modulate plant responses to salt stress via affecting ATP and gibberellin metabolism in Arabidopsis thaliana. PLoS ONE 2012, 7, e43530. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhu, W.; Dai, S.; Gai, S.; Zheng, G.; Zheng, C. The involvement of mitochondrial phosphate transporter in accelerating bud dormancy release during chilling treatment of tree peony (Paeonia suffruticosa). Planta 2008, 228, 545–552. [Google Scholar] [CrossRef]
- De Croos, J.N.A.; McNally, J.D.; Palmieri, F.; Storey, K.B. Upregulation of the mitochondrial phosphate carrier during freezing in the wood frog rana sylvatica: Potential roles of transporters in freeze tolerance. J. Bioenergy Biomembr. 2004, 36, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Kakani, A.; Peng, Z. ARR5 and ARR6 mediate tissue specific cross-talk between auxin and cytokinin in Arabidopsis. Am. J. Plant Sci. 2011, 2, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Sung, D.Y.; Kaplan, F.; Lee, K.J.; Guy, C.L. Acquired tolerance to temperature extremes. Trends Plant Sci. 2003, 8, 179–187. [Google Scholar] [CrossRef]
- Huner, N.P.A.; Öquist, G.; Hurry, V.M.; Krol, M.; Falk, S.; Griffith, M. Photosynthesis, photoinhibition and low temperature acclimation in cold tolerant plants. Photosynth. Res. 1993, 37, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Hurry, V.M.; Huner, N.P.A. Effects of cold-hardening on sensitivity of winter and spring wheat leaves to short-term photoinhibition and recovery of photosynthesis. Plant Physiol. 1992, 100, 1283–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Öquist, G.; Hurry, V.M.; Huner, N.P.A. Low-temperature effects on photosynthesis and correlation with freezing tolerance in spring and winter cultivars of wheat and rye. Plant Physiol. 1993, 101, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Karki, A.; Horvath, D.P.; Sutton, F. Induction of DREB2A pathway with repression of E2F, Jasmonic acid biosynthetic and photosynthesis pathways in cold acclimation specific freeze resistant wheat crown. Funct. Integr. Genom. 2013, 13, 57–65. [Google Scholar] [CrossRef]
- Liu, D.; Horvath, D.; Li, P.; Liu, W. RNA sequencing characterizes transcriptomes differences in cold response between northern and southern Alternanthera philoxeroides and highlight adaptations associated with northward expansion. Front. Plant Sci. 2019, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Hudson, M.E.; Quail, P.H. Identification of promoter motifs involved in the network of phytochrome A-regulated gene expression by combined analysis of genomic sequence and microarray data. Plant Physiol. 2003, 133, 1605–1616. [Google Scholar] [CrossRef] [Green Version]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998, 280, 104–106. [Google Scholar] [CrossRef] [Green Version]
- Savitch, L.V.; Allard, G.; Seki, M.; Robert, L.S.; Tinker, N.A.; Hüner, N.P.A.; Shinozaki, K.; Singh, J. The effect of overexpression of two Brassica CBF/DREB1-like transcription factors on photosynthetic capacity and freezing tolerance in Brassica napus. Plant Cell Physiol. 2005, 46, 1525–1539. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, Y.; Randall, S.K. Functionality of soybean CBF/DREB1 transcription factors. Plant Sci. 2016, 246, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvallo, M.A.; Pino, M.T.; Jeknic, Z.; Zou, C.; Doherty, C.J.; Shiu, S.H.; Chen, T.H.H.; Thomashow, M.F. A comparison of the low temperature transcriptomes and CBF regulons of three plant species that differ in freezing tolerance: Solanum commersonii, Solanum tuberosum, and Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 3807–3819. [Google Scholar] [CrossRef]
- Gehan, M.A.; Park, S.; Gilmour, S.J.; An, C.; Lee, C.M.; Thomashow, M.F. Natural variation in the C-repeat binding factor cold response pathway correlates with local adaptation of Arabidopsis ecotypes. Plant J. 2015, 84, 682–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wang, X.; Wang, H.; Xin, H.; Yang, X.; Yan, J.; Li, J.; Tran, L.-S.P.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; et al. Genome-wide analysis of ZmDREB genes and their association with natural variation in drought tolerance at seedling stage of Zea mays L. PLoS Genet. 2013, 9, e1003790. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Correlation of Visual Damage Scores at 2 Weeks Post Freezing with 3- and 7-Day Fv/Fo Data | |||
|---|---|---|---|
| Frz Avg | 3 Day Fv/Fo Avg | 7 Day Fv/Fo Avg | |
| Frz Avg | 1 | ||
| 3 day Fv/Fo Avg | 0.5975 | 1 | |
| 7 day Fv/Fo Avg | 0.7208 | 0.8409 | 1 |
| Model | MSD 2week_run1_Lsmeans | MSD 2week_run2_Lsmeans | MSD FvFo3d_run1_Lsmeans | MSD FvFo3d_run2_Lsmeans | MSD FvFo7d_run1_LSmeans | MSD FvFo7d_run2_LSmeans | Average |
|---|---|---|---|---|---|---|---|
| MLM Naïve kinship | 2.36 × 10−5 | 3.10 × 10−4 | 7.98 × 10−5 | 1.02 × 10−5 | 1.18 × 10−5 | 2.71 × 10−6 | 7.30 × 10−5 |
| MLM PC17 kinship | 4.40 × 10−5 | 3.72 × 10−4 | 1.24 × 10−5 | 1.54 × 10−5 | 1.56 × 10−5 | 2.92 × 10−6 | 7.71 × 10−5 |
| MLM PC1 kinship | 4.48 × 10−4 | 4.33 × 10−4 | 8.47 × 10−5 | 1.23 × 10−5 | 1.03 × 10−5 | 2.46 × 10−6 | 1.65 × 10−4 |
| GLM PC17 population | 1.38 × 10−4 | 1.51 × 10−4 | 1.32 × 10−4 | 8.13 × 10−4 | 2.57 × 10−5 | 4.80 × 10−4 | 2.90 × 10−4 |
| MLM PC17 kinship population | 3.03 × 10−5 | 2.26 × 10−4 | 7.19 × 10−4 | 1.70 × 10−5 | 4.90 × 10−4 | 6.02 × 10−4 | 3.47 × 10−4 |
| GLM PC17 | 3.25 × 10−4 | 3.10 × 10−4 | 1.76 × 10−4 | 8.75 × 10−4 | 3.01 × 10−4 | 5.09 × 10−4 | 4.16 × 10−4 |
| MLM Naïve kinship population | 1.08 × 10−3 | 1.19 × 10−4 | 5.90 × 10−4 | 2.49 × 10−3 | 1.78 × 10−3 | 2.09 × 10−3 | 1.36 × 10−3 |
| GLM Naïve population | 1.41 × 10−3 | 1.11 × 10−4 | 1.24 × 10−3 | 3.19 × 10−3 | 1.22 × 10−3 | 2.02 × 10−3 | 1.53 × 10−3 |
| GLM PC1 population | 1.48 × 10−3 | 1.09 × 10−4 | 1.21 × 10−3 | 3.20 × 10−3 | 1.24 × 10−3 | 2.3 × 10−3 | 1.54 × 10−3 |
| GLM PC1 | 2.54 × 10−3 | 1.47 × 10−4 | 1.75 × 10−3 | 3.26 × 10−3 | 2.96 × 10−3 | 2.23 × 10−3 | 2.15 × 10−3 |
| MLM PC1 kinship population | 6.24 × 10−4 | 2.83 × 10−4 | 1.23 × 10−3 | 5.16 × 10−3 | 1.13 × 10−3 | 4.77 × 10−3 | 2.20 × 10−3 |
| GLM Naïve | 1.59 × 10−2 | 1.10 × 10−3 | 3.78 × 10−3 | 3.45 × 10−3 | 2.47 × 10−3 | 2.18 × 10−3 | 4.82 × 10−3 |
| MLM PC1 | MLM PC17 | MLM Naïv | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 Week Damage | 3 Day Fv/Fo | 7 Day Fv/Fo | 2 Week Damage | 3 Day Fv/Fo | 7 Day Fv/Fo | 2 Week Damage | 3 Day Fv/Fo | 7 Day Fv/Fo | |||||||||||
| Chromosom | Mark | Exp 1 | Exp 2 | Exp 1 | Exp 2 | Exp 1 | Exp 2 | Exp 1 | Exp 2 | Exp 1 | Exp 2 | Exp 1 | Exp 2 | Exp 1 | Exp 2 | Exp 1 | Exp 2 | Exp 1 | Exp 2 |
| A04_RANDOM | S1_656249548 | 1.53 × 10−4 | 1.20 × 10−4 | 1.01 × 10−4 | 2.97 × 10−3 | 9.39 × 10−6 | 1.05 × 10−3 | 2.82 × 10−3 | 1.14 × 10−3 | 9.42 × 10−4 | 6.70 × 10−3 | 5.77 × 10−5 | 1.05 × 10−3 | 8.04 × 10−4 | 3.24 × 10−4 | 1.13 × 10−4 | 3.04 × 10−3 | 8.67 × 10−6 | 1.07 × 10−3 |
| A07 | S1_155086644 | 8.06 × 10−3 | 6.93 × 10−5 | 9.51 × 10−3 | 1.00 × 10−3 | 1.15 × 10−4 | 1.19 × 10−3 | 8.94 × 10−4 | 1.42 × 10−4 | 8.86 × 10−3 | 2.78 × 10−3 | 6.15 × 10−5 | 1.19 × 10−3 | 1.20 × 10−3 | 7.78 × 10−5 | 9.88 × 10−3 | 1.04 × 10−3 | 1.15 × 10−4 | 1.23 × 10−3 |
| A09 | S1_209198432 | 5.74 × 10−2 | 2.50 × 10−3 | 2.09 × 10−3 | 3.16 × 10−4 | 2.51 × 10−3 | 3.94 × 10−4 | 9.03 × 10−2 | 6.76 × 10−3 | 2.39 × 10−3 | 8.64 × 10−4 | 1.42 × 10−3 | 3.94 × 10−4 | 8.12 × 10−2 | 4.21 × 10−3 | 2.17 × 10−3 | 3.23 × 10−4 | 2.53 × 10−3 | 3.98 × 10−4 |
| ANN_RANDOM | S1_739997351 | 1.45 × 10−2 | 2.86 × 10−1 | 1.24 × 10−1 | 1.94 × 10−3 | 1.44 × 10−3 | 1.56 × 10−3 | 1.55 × 10−2 | 3.45 × 10−1 | 1.56 × 10−1 | 1.62 × 10−3 | 7.17 × 10−4 | 1.56 × 10−3 | 1.01 × 10−2 | 2.86 × 10−1 | 1.19 × 10−1 | 1.83 × 10−3 | 1.42 × 10−3 | 1.47 × 10−3 |
| ANN_RANDOM | S1_738423768 | 4.77 × 10−2 | 7.73 × 10−3 | 9.94 × 10−3 | 9.17 × 10−3 | 2.19 × 10−3 | 2.45 × 10−3 | 1.08 × 10−1 | 1.37 × 10−2 | 1.11 × 10−2 | 2.38 × 10−3 | 1.08 × 10−3 | 2.45 × 10−3 | 4.43 × 10−2 | 1.32 × 10−2 | 1.09 × 10−2 | 3.75 × 10−3 | 1.82 × 10−3 | 4.86 × 10−4 |
| C02 | S1_316742524 | 2.05 × 10−3 | 1.60 × 10−3 | 2.49 × 10−1 | 1.45 × 10−1 | 1.64 × 10−1 | 9.41 × 10−2 | 6.35 × 10−4 | 2.87 × 10−3 | 2.50 × 10−1 | 1.74 × 10−1 | 1.50 × 10−1 | 9.41 × 10−2 | 6.03 × 10−4 | 6.85 × 10−4 | 2.59 × 10−1 | 1.35 × 10−1 | 1.64 × 10−1 | 8.81 × 10−2 |
| C03 | S1_359625016 | 1.97 × 10−1 | 1.55 × 10−2 | 2.10 × 10−2 | 3.33 × 10−3 | 1.44 × 10−3 | 2.60 × 10−3 | 2.10 × 10−1 | 2.96 × 10−2 | 2.45 × 10−2 | 1.09 × 10−2 | 2.07 × 10−3 | 2.60 × 10−3 | 2.40 × 10−1 | 1.81 × 10−2 | 2.20 × 10−2 | 3.45 × 10−3 | 1.46 × 10−3 | 2.69 × 10−3 |
| C03 | S1_376295769 | 2.37 × 10−3 | 1.69 × 10−4 | 3.09 × 10−1 | 3.23 × 10−3 | 4.36 × 10−1 | 1.36 × 10−3 | 7.00 × 10−4 | 7.86 × 10−4 | 3.80 × 10−1 | 7.00 × 10−3 | 4.13 × 10−1 | 1.36 × 10−3 | 6.28 × 10−4 | 2.05 × 10−4 | 3.14 × 10−1 | 3.30 × 10−3 | 4.31 × 10−1 | 1.38 × 10−3 |
| C07 | S1_528342773 | 5.40 × 10−2 | 1.11 × 10−2 | 1.04 × 10−2 | 3.63 × 10−3 | 1.81 × 10−3 | 4.74 × 10−4 | 4.10 × 10−2 | 2.55 × 10−2 | 9.65 × 10−3 | 5.54 × 10−4 | 1.01 × 10−3 | 4.74 × 10−4 | 1.06 × 10−1 | 1.54 × 10−2 | 9.41 × 10−3 | 9.52 × 10−3 | 2.22 × 10−3 | 2.52 × 10−3 |
| C07 | S1_515244307 | 1.59 × 10−3 | 4.79 × 10−4 | 6.92 × 10−1 | 2.58 × 10−2 | 3.90 × 10−1 | 4.00 × 10−3 | 4.89 × 10−4 | 1.66 × 10−4 | 6.57 × 10−1 | 4.67 × 10−3 | 3.91 × 10−1 | 4.00 × 10−3 | 4.62 × 10−4 | 6.20 × 10−4 | 6.82 × 10−1 | 2.61 × 10−2 | 3.85 × 10−1 | 4.06 × 10−3 |
| C08 | S1_567184601 | 7.64 × 10−3 | 4.51 × 10−2 | 4.29 × 10−1 | 2.65 × 10−3 | 6.63 × 10−4 | 2.37 × 10−3 | 8.74 × 10−4 | 7.76 × 10−2 | 3.86 × 10−1 | 7.53 × 10−3 | 5.02 × 10−4 | 2.37 × 10−3 | 1.17 × 10−3 | 4.19 × 10−2 | 4.32 × 10−1 | 2.59 × 10−3 | 6.56 × 10−4 | 2.31 × 10−3 |
| C09 | S1_641455172 | 1.05 × 10−1 | 8.96 × 10−1 | 1.22 × 10−4 | 1.49 × 10−3 | 6.94 × 10−3 | 1.34 × 10−2 | 3.97 × 10−1 | 8.76 × 10−1 | 2.35 × 10−3 | 4.82 × 10−3 | 1.99 × 10−2 | 1.34 × 10−2 | 4.23 × 10−1 | 9.65 × 10−1 | 1.29 × 10−4 | 1.86 × 1003 | 6.53 × 10−3 | 1.55 × 10−2 |
| C09 | S1_641455176 | 1.05 × 10−1 | 8.96 × 10−1 | 1.22 × 10−4 | 1.49 × 10−3 | 6.94 × 10−3 | 1.34 × 10−2 | 3.97 × 10−1 | 8.76 × 10−1 | 2.35 × 10−3 | 4.82 × 10−3 | 1.99 × 10−2 | 1.34 × 10−2 | 4.23 × 10−1 | 9.65 × 10−1 | 1.29 × 10−4 | 1.86 × 10−3 | 6.53 × 10−3 | 1.55 × 10−2 |
| Marker | Chromosome | Position | ROI Based on Local LD-Decay Rate | Canola Gene Model | AGI Orthologue | Names |
|---|---|---|---|---|---|---|
| S1_656249548 | A04_random | 522883 | 521883-523883 | BnaA04g27970D | AT5G37890 | RING E3 LIGASE (SINAL7) |
| S1_155086644 | A07 | 10641317 | 10625317-10657317 | BnaA07g11360D | AT1G20630 | CATALASE 1 (CAT1) |
| S1_155086644 | A07 | 10641317 | 10625317-10657317 | BnaA07g11370D | AT1G20620 | CATALASE 3 (CAT3) |
| S1_155086644 | A07 | 10641317 | 10625317-10657317 | BnaA07g11440D | AT1G20470 | SMALL AUXIN UPREGULATED RNA 60 (SAUR60) |
| S1_155086644 | A07 | 10641317 | 10625317-10657317 | BnaA07g11450D | AT1G20450 | LOW TEMPERATURE INDUCED 29 (LTI29) |
| S1_209198432 | A09 | 21784443 | 21771443-21797443 | BnaA09g28870D | AT1G26340 | CYTOCHROME B5-A (CB5-A) |
| S1_209198432 | A09 | 21784443 | 21771443-21797443 | BnaA09g28880D | AT1G26370 | ROOT INITIATION DEFECTIVE 1 (RID1) |
| S1_209198432 | A09 | 21784443 | 21771443-21797443 | BnaA09g28910D | AT1G26440 | UREIDE PERMEASE 5 (UPS5) |
| S1_209198432 | A09 | 21784443 | 21771443-21797443 | BnaA09g28950D | AT1G26480 | GENERAL REGULATORY FACTOR 12 (GRF12) |
| S1_209198432 | A09 | 21784443 | 21771443-21797443 | BnaA09g28990D | AT1G26560 | BETA GLUCOSIDASE 40 (BGLU40) |
| S1_209198432 | A09 | 21784443 | 21771443-21797443 | BnaA09g29000D | AT1G26570 | UDP-GLUCOSE DEHYDROGENASE 1 (UGD1) |
| S1_739997351 | ANN-RANDOM | 27349546 | 27348546-27350546 | BnaAnng23820D | AT1G08640 | CHLOROPLAST J-LIKE DOMAIN 1, CJD1 |
| S1_738423768 | ANN_RANDOM | 25775963 | 25774963-25776963 | BnaAnng22920D | AT2G34420 | PHOTOSYSTEM II LIGHT HARVESTING COMPLEX B1B2 (LHB1B2) |
| S1_738423768 | ANN_RANDOM | 25775963 | 25774963-25776963 | BnaAnng22950D | AT5G18460 | Carboxyl-terminal peptidase |
| S1_316742524 | C02 | 39235351 | 39234351-39236351 | BnaC02g36140D | AT2G19890 | Hypothetical protein |
| S1_359625016 | C03 | 35895939 | 35877939-35913939 | BnaC03g51340D | AT5G62920 | RESPONSE REGULATOR 6 (ARR6) |
| S1_376295769 | C03 | 52566692 | 52543692-52589692 | BnaC03g63190D | AT4G19420 | PECTIN ACETYLESTERASE 8 (PAE8) |
| S1_528342773 | C07 | 14698486 | 14397486-14999486 | BnaC07g09400D | AT1G29930 | CHLOROPHYLL A/B PROTEIN 140 (CAB140) |
| S1_567184601 | C08 | 8769737 | 8770737-8768737 | BnaC08g06270D | AT1G23310 | ALANINE-2-OXOGLUTARATE AMINOTRANSFERASE 1 (AOAT1) |
| S1_567184601 | C08 | 8769737 | 8770737-8768737 | BnaC08g06310D | AT1G23360 | MENG |
| S1_567184601 | C08 | 8769737 | 8770737-8768737 | BnaC08g06320D | AT1G23380 | KNOTTED1-LIKE HOMEOBOX GENE 6 (KNAT6) |
| S1_641455172 | C09 | 44563121 | 44562121-44564121 | BnaC09g43140D | AT5G14040 | MITOCHONDRIAL PHOSPHATE TRANSPORTER (MPT) |
| S1_641455172 | C09 | 44563121 | 44562121-44564121 | BnaC09g43220D | AT5G13970 | Midasin-like protein |
| S1_641455172 | C09 | 44563121 | 44562121-44564121 | BnaC09g43230D | AT5G13950 | Nuclear factor kappa-B-binding protein |
| S1_641455172 | C09 | 44563121 | 44562121-44564121 | BnaC09g43250D | AT5G13930 | CHALCONE SYNTHASE (CHS) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chao, W.S.; Horvath, D.P.; Stamm, M.J.; Anderson, J.V. Genome-Wide Association Mapping of Freezing Tolerance Loci in Canola (Brassica napus L.). Agronomy 2021, 11, 233. https://doi.org/10.3390/agronomy11020233
Chao WS, Horvath DP, Stamm MJ, Anderson JV. Genome-Wide Association Mapping of Freezing Tolerance Loci in Canola (Brassica napus L.). Agronomy. 2021; 11(2):233. https://doi.org/10.3390/agronomy11020233
Chicago/Turabian StyleChao, Wun S., David P. Horvath, Michael J. Stamm, and James V. Anderson. 2021. "Genome-Wide Association Mapping of Freezing Tolerance Loci in Canola (Brassica napus L.)" Agronomy 11, no. 2: 233. https://doi.org/10.3390/agronomy11020233
APA StyleChao, W. S., Horvath, D. P., Stamm, M. J., & Anderson, J. V. (2021). Genome-Wide Association Mapping of Freezing Tolerance Loci in Canola (Brassica napus L.). Agronomy, 11(2), 233. https://doi.org/10.3390/agronomy11020233