
Genome-Wide Identification of the NHX Gene Family in Punica granatum L. and Their Expressional Patterns under Salt Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Sequence Analysis of PgNHXs
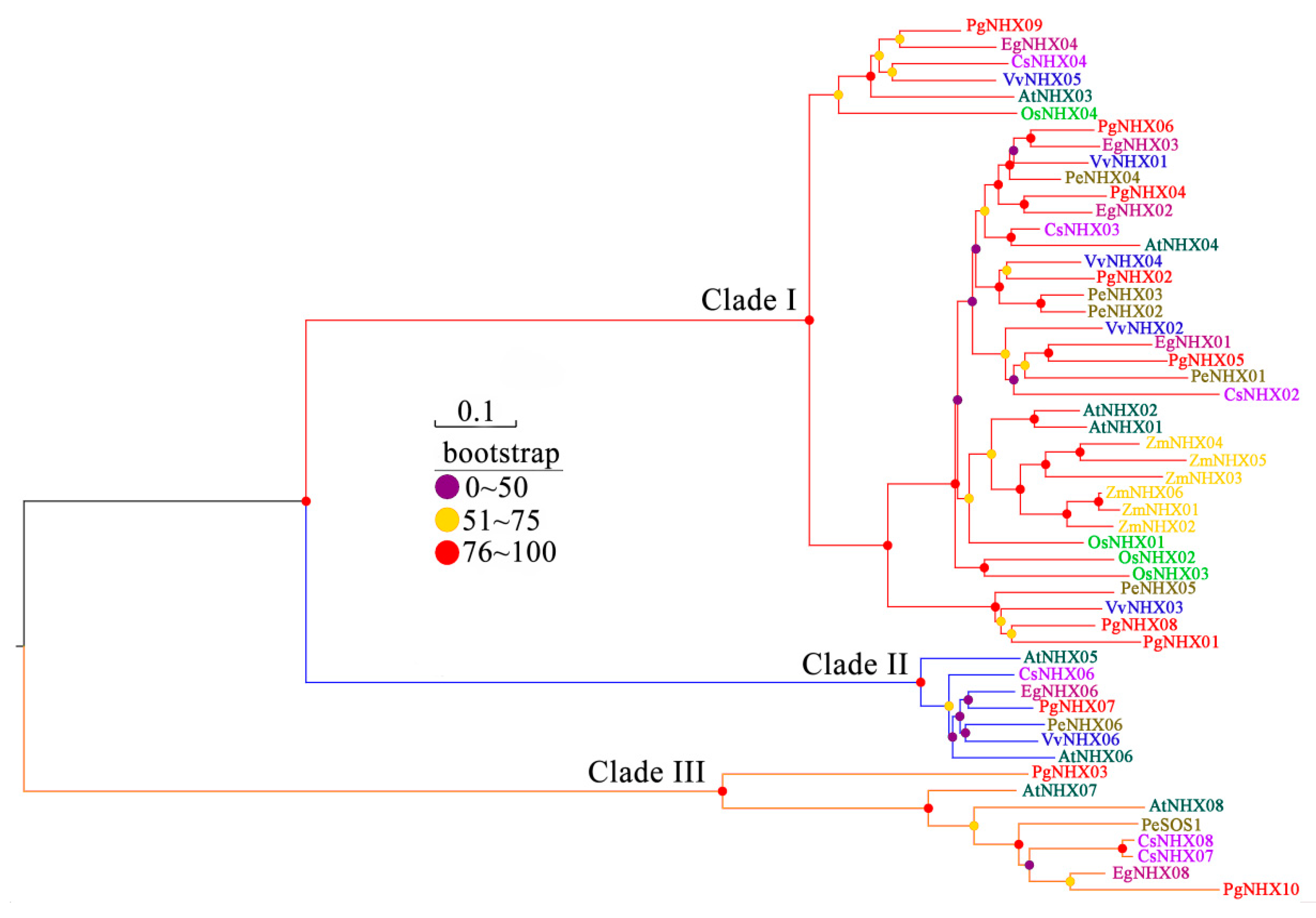
2.2. Phylogenetic Analysis
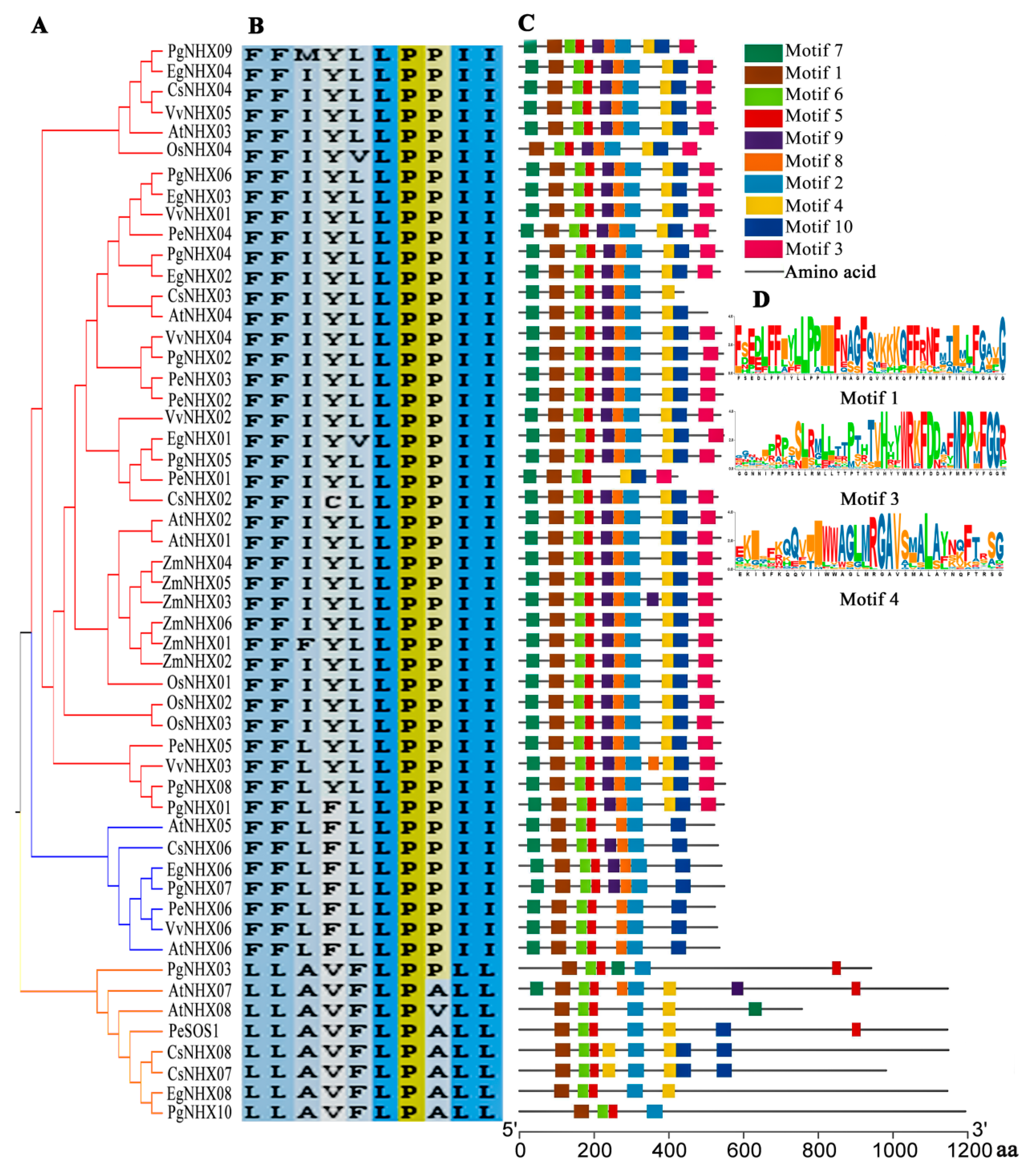
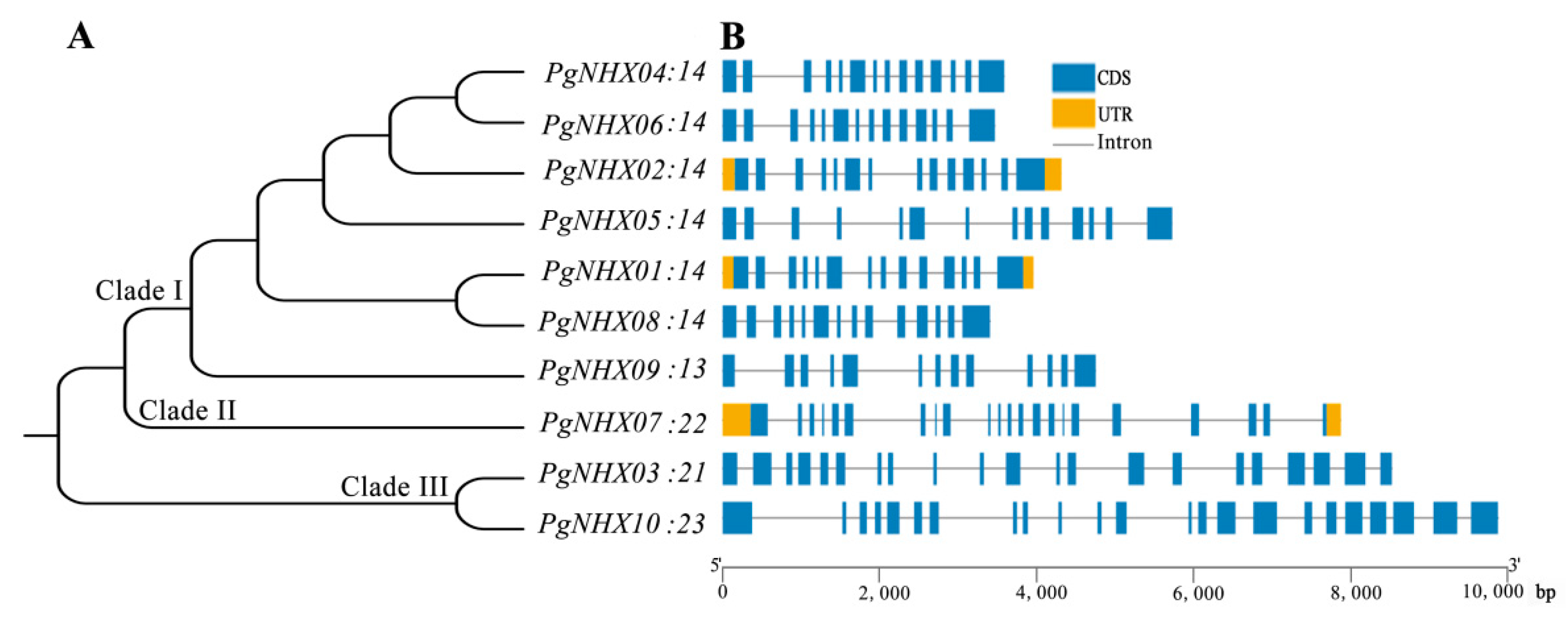
2.3. Motif Identification and Gene Structure Analysis
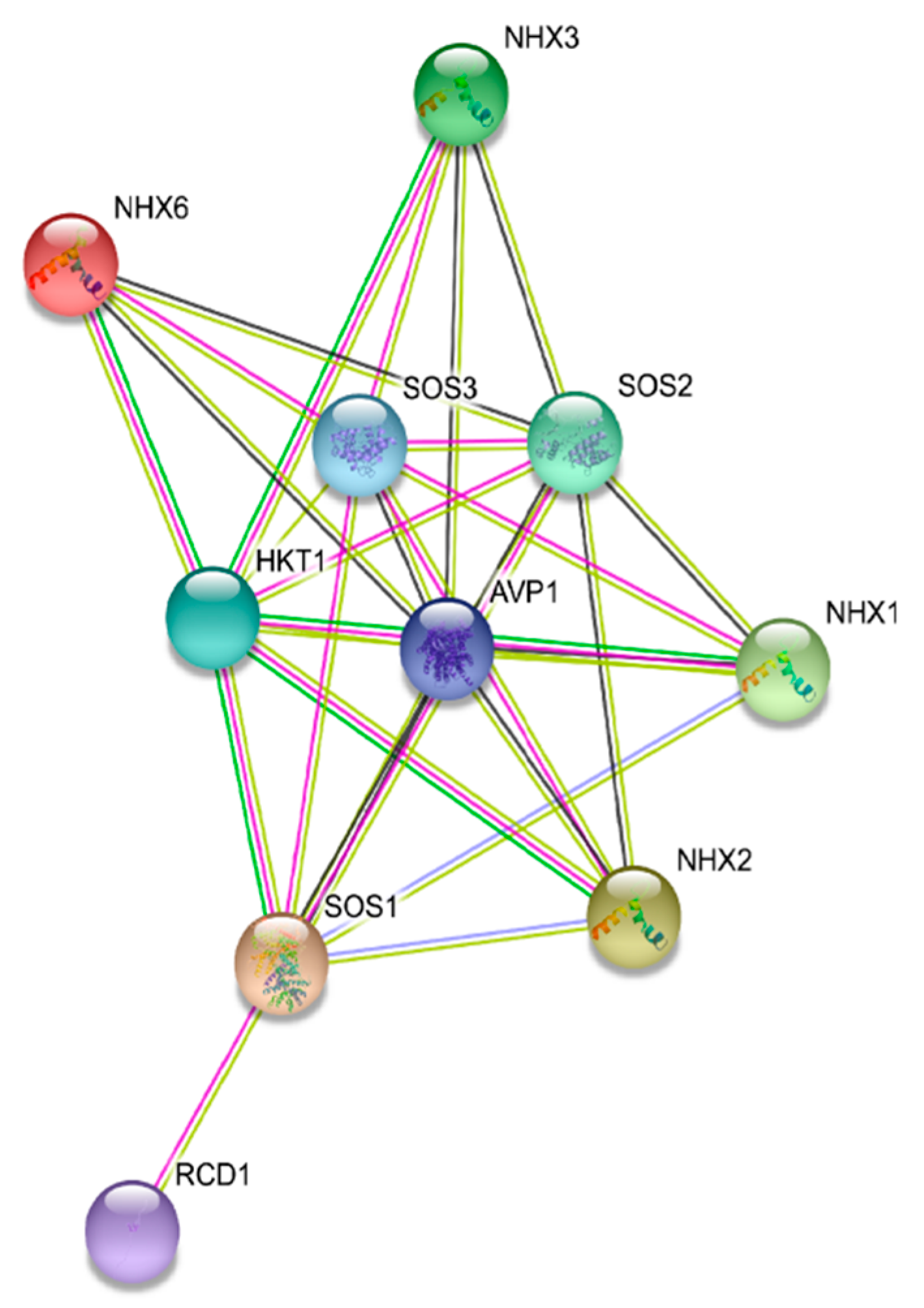
2.4. Protein Tertiary Structure and Protein Interaction Network Analysis
2.5. Cis-Acting Elements Located in NHX Gene Promoters
2.6. Plant Material, Treatment, and qRT-PCR Analysis
3. Results
3.1. Identification and Sequence Analysis of PgNHXs
3.2. Phylogenetic Analysis
3.3. Motif Identification and Gene Structure of PgNHXs
3.4. Protein Tertiary Structure and Protein Interaction Network Analysis
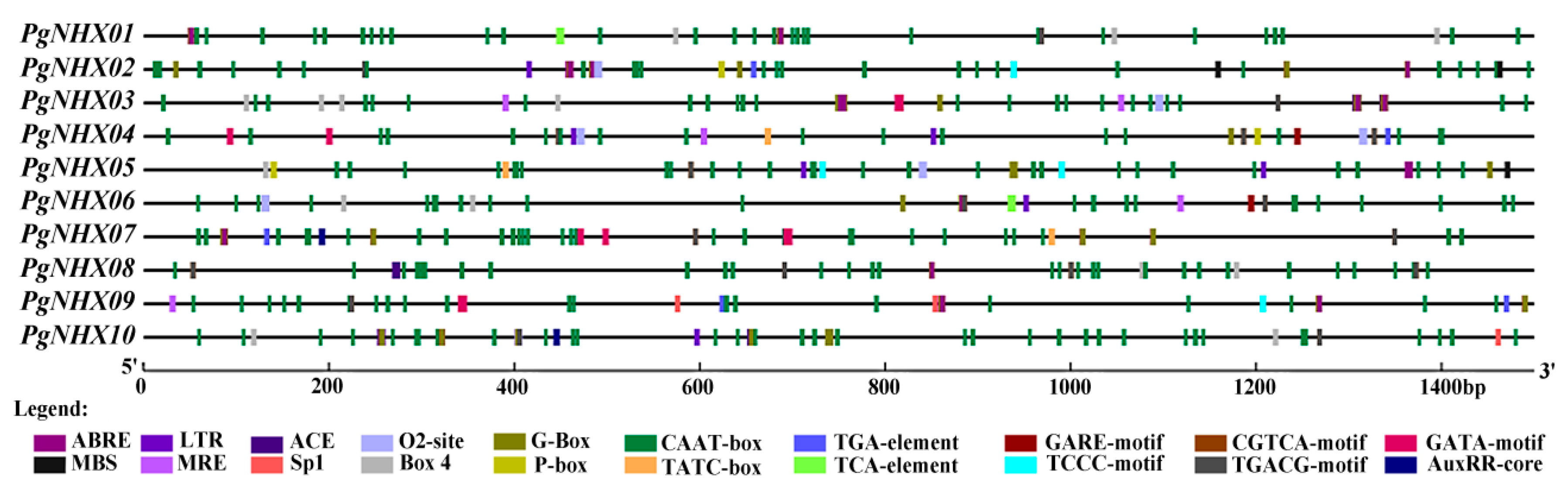
3.5. Cis-Acting Elements Located in Promoters of PgNHXs
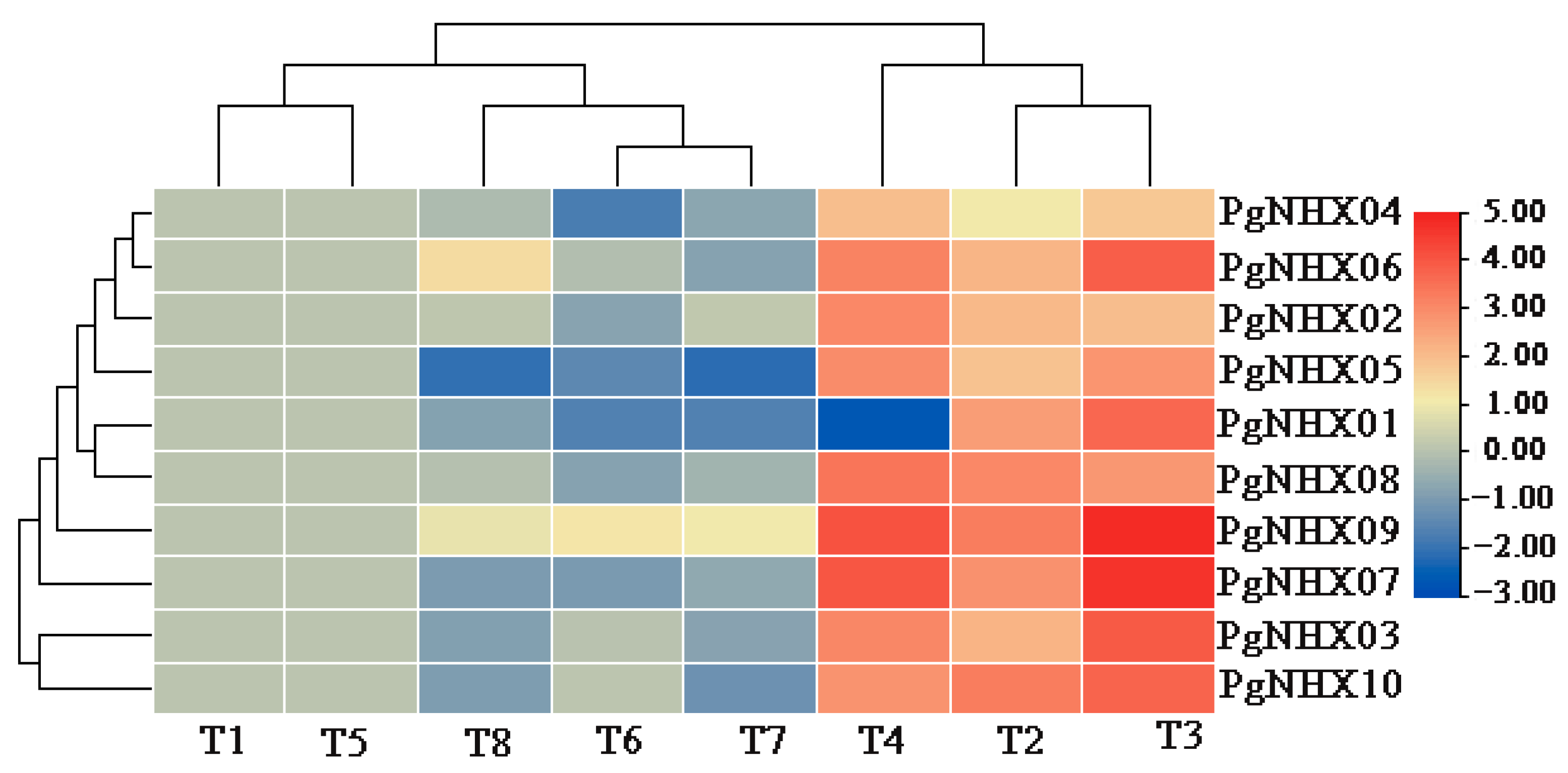
3.6. qRT-PCR Analysis of PgNHXs under Salt Stress
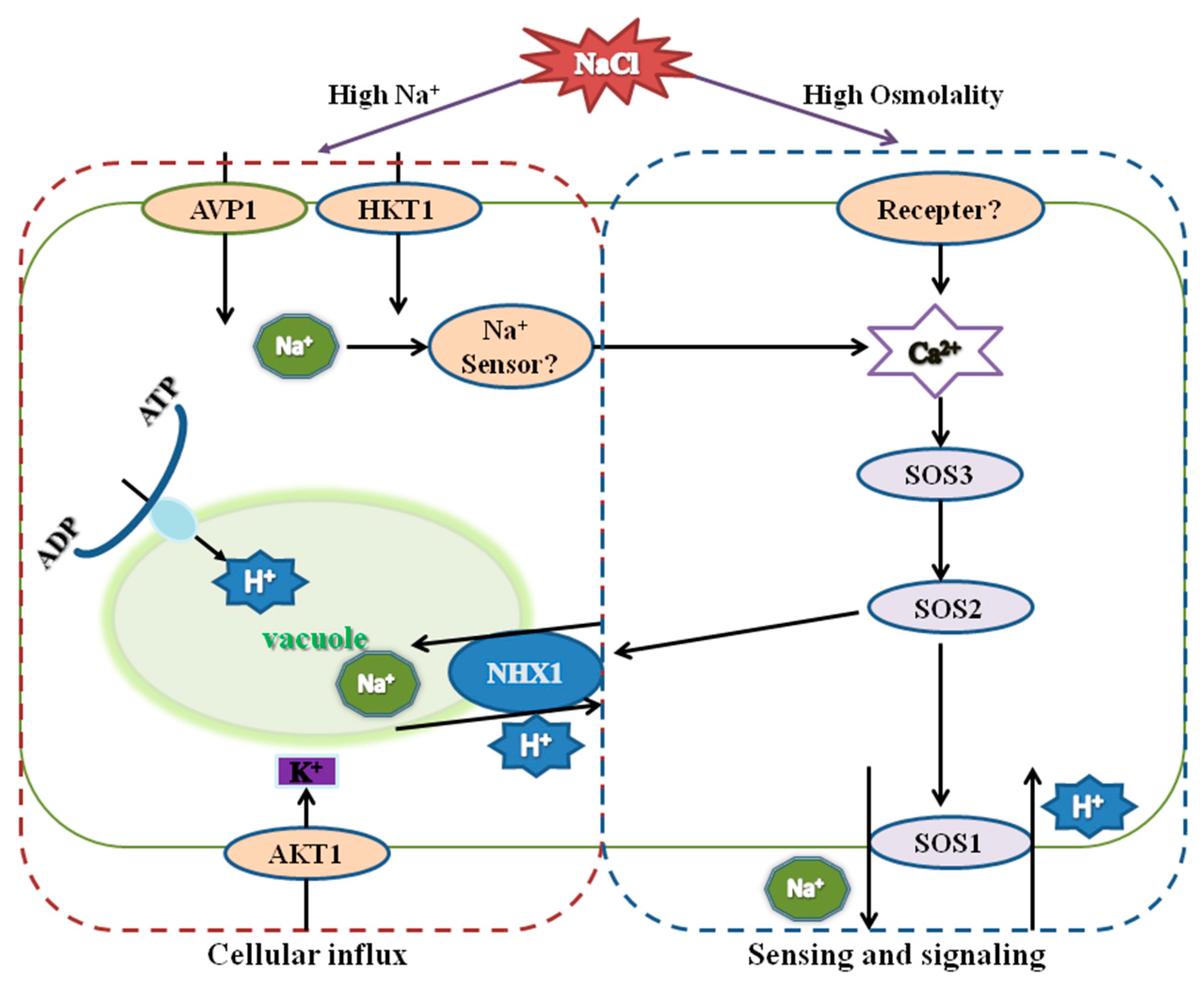
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Kovda, V.A. Loss of productive land due to salinization. Ambio 1983, 12, 91–93. [Google Scholar] [CrossRef]
- Xiong, L.M.; Zhu, J.K. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ. 2002, 25, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Abdul Qados, A.M.S. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba L. J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.T.; Chen, X.; Wu, B.-Y.; Yuan, X.X.; Zhang, H.M.; Cui, X.Y.; Liu, X.Q. Whole-genome identification and expression analysis of K+ efflux antiporter (KEA) and Na+/H+ antiporter (NHX) families under abiotic stress in soybean. J. Integr. Agric. 2015, 14, 1171–1183. [Google Scholar] [CrossRef]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta 2010, 233, 175–188. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef] [Green Version]
- Rubio, F.; Gassmann, W.; Schroeder, J.I. Sodium-driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance. Science 1995, 270, 1660–1663. [Google Scholar] [CrossRef]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef]
- Ayadi, M.; Ben Ayed, R.; Mzid, R.; Aifa, S.; Hanana, M. Computational approach for structural feature determination of grapevine NHX antiporters. BioMed Res. Int. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dibrov, P.; Fliegel, L. Comparative molecular analysis of Na+/H+ exchangers: A unified model for Na+/H+ antiport? FEBS Lett. 1998, 424, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.S. Plant and yeast NHX antiporters: Roles in membrane trafficking. J. Integr. Plant Biol. 2012, 54, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Bassil, E.; Zhang, S.Q.; Gong, H.J.; Tajima, H.; Blumwald, E. Cation specificity of vacuolar NHX-type cation/H+ antiporters. Plant Physiol. 2019, 179, 616–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brett, C.L.; Donowitz, M.; Rao, R. Evolutionary origins of eukaryotic sodium/proton exchangers. Am. J. Physiol. Cell Physiol. 2005, 288, C223–C239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayadi, M.; Martins, V.; Ben Ayed, R.; Jbir, R.; Feki, M.; Mzid, R.; Géros, H.; Aifa, S.; Hanana, M. Genome wide identification, molecular characterization, and gene expression analyses of grapevine NHX antiporters suggest their involvement in growth, ripening, seed dormancy, and stress response. Biochem. Genet. 2020, 58, 102–128. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Q.; Wang, J.L.; Li, S.J. Genome-wide Identification of Na+ /H+ antiporter (NHX) genes in sugar beet (Beta vulgaris L.) and their regulated expression under salt stress. Genes 2019, 10, 401. [Google Scholar] [CrossRef] [Green Version]
- Kumari, H.; Kumar, S.; Ramesh, K.; Palakolanu, S.R.; Marka, N.; Prakash, A.; Shah, T.; Henderson, A.; Srivastava, R.; Rajasheker, G.; et al. Genome-wide identification and analysis of arabidopsis sodium proton antiporter (NHX) and human sodium proton exchanger (NHE) homologs in Sorghum bicolor. Genes 2018, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Akram, U.; Song, Y.; Liang, C.; Abid, M.; Askari, M.; Myat, A.; Abbas, M.; Malik, W.; Ali, Z.; Guo, S.; et al. Genome-wide characterization and expression analysis of NHX gene family under salinity stress in Gossypium barbadense and Its comparison with Gossypium hirsutum. Genes 2020, 11, 803. [Google Scholar] [CrossRef]
- Qiu, Q.S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef] [Green Version]
- Quintero, F.J.; Ohta, M.; Shi, H.Z.; Zhu, J.K.; Pardo, J.M. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 9061–9066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zheng, Q.S.; Shen, Q.R.; Guo, S.W. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, K.B.; Wu, Y.X. Footprints of divergent evolution in two Na+/H+ type antiporter gene families (NHX and SOS1) in the genus Populus. Tree Physiol. 2018, 38, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.H.; Yin, Y.L.; Qu, J.L.; Zhu, L.Q.; Li, Y. Population genetic diversity in chinese pomegranate (Punica granatum L.) cultivars revealed by fluorescent-AFLP markers. J. Genet. Genom. 2017, 34, 1061–1071. [Google Scholar] [CrossRef]
- Fahmy, H.; Hegazi, N.; El-Shamy, S.; Farag, M.A. Pomegranate juice as a functional food: A comprehensive review of its polyphenols, therapeutic merits, and recent patents. Food Funct. 2020, 11, 1–27. [Google Scholar] [CrossRef]
- Tozzi, F.; Legua, P.; Martínez-Nicolás, J.J.; Núez-Gómez, D.; Melgarejo, P. Morphological and nutraceutical characterization of six pomegranate cultivars of global commercial interest. Sci. Hortic. 2020, 272, 1–9. [Google Scholar] [CrossRef]
- Liu, C.Y.; Zhao, X.Q.; Yan, J.X.; Yuan, Z.H.; Gu, M.M. Effects of salt stress on growth, photosynthesis, and mineral nutrients of 18 pomegranate (Punica granatum) cultivars. Agronomy 2019, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Zhao, Y.J.; Zhao, X.Q.; Wang, J.P.; Gu, M.M.; Yuan, Z.H. Transcriptomic profiling of pomegranate provides insights into salt tolerance. Agronomy 2020, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Bhantana, P.; Lazarovitch, N. Evapotranspiration, crop coefficient and growth of two young pomegranate (Punica granatum L.) varieties under salt stress. Agric.Water Manag. 2010, 97, 715–722. [Google Scholar] [CrossRef]
- Cui, J.Q.; Hua, Y.P.; Zhou, T.; Liu, Y.; Huang, J.Y.; Yue, C.P. Global landscapes of the Na+/H+ antiporter (NHX) family members uncover their potential roles in regulating the rapeseed resistance to salt stress. Int. J. Mol. Sci. 2020, 21, 3429. [Google Scholar] [CrossRef]
- Li, W.H.; Du, J.; Feng, H.M.; Wu, Q.; Xu, G.H.; Shabala, S.; Yu, L. Function of NHX-type transporters in improving rice tolerance to aluminum stress and soil acidity. Planta 2020, 251, 71. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Zhao, J.R.; Guo, D.D.; Ma, X.N.; Gao, W. Identification of NHXs in Gossypium species and the positive role of GhNHX1 in salt tolerance. BMC Plant Biol. 2020, 20, 147. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.H.; Xu, C.Y.; Ming, R.; Tang, H.B.; Guyot, R.; Kramer, E.M.; Hu, Y.D.; Yi, X.K. The pomegranate (Punica granatum L.) genome and the genomics of punicalagin biosynthesis. Plant J. 2017, 91, 1108–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamesch, P.; Berardini, T.Z.; Li, D.H.; Swarbreck, D.; Wilks, C.; Sasidharan, R. The Arabidopsis Information Resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1201–D1210. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, C.; Huang, X.; Zhang, H.; Yuan, Z. Land-plant phylogenomic and pomegranate transcriptomic analyses reveal an evolutionary scenario of CYP75 genes subsequent to whole genome duplications. J. Plant Biol. 2019, 62, 48–60. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H. CDD: A conserved domain database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; Humana Press: Totowa, NJ, USA, 2005. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Heijne, G.V.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [Green Version]
- Michele, C.; James, C.; Stephen, M.S.; Geoffrey, J.B. The Jalview Java alignment editor. Bioinformatics 2004, 20, 426–427. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, B.; Gao, S.H.; Lercher, M.J.; Hu, S.N.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Xia, R.; Chen, H.; He, Y.H. TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface. BioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. I-TASSER Server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yun, C.H.C.; Little, P.J.; Nath, S.K.; Levine, S.A.; Pouyssegur, J. Leu143 in the putative fourth membrane spanning domain is critical for amiloride inhibition of an epithelial Na+/H+ exchanger isoform (NHE-2). Biochem. Biophys. Res. Commun. 1993, 193, 532–539. [Google Scholar] [CrossRef]
- Li, N.N.; Wang, X.; Ma, B.J.; Du, C.; Zheng, L.L.; Wang, Y.C. Expression of a Na(+)/H(+) antiporter RtNHX1 from a recretohalophyte Reaumuria trigyna improved salt tolerance of transgenic Arabidopsis thaliana. J. Plant Physiol. 2017, 218, 109–120. [Google Scholar] [CrossRef]
- Tian, F.X.; Chang, E.; Li, Y.; Sun, P.; Hu, J.J.; Zhang, J. Expression and integrated network analyses revealed functional divergence of NHX-type Na+/H+ exchanger genes in poplar. Sci. Rep. 2017, 7, 2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Counillon, L.; Franchi, A.; Pouyssegur, J. A point mutation of the Na+/H+ exchanger gene (NHE1) and amplification of the mutated allele confer amiloride resistance upon chronic acidosis. Proc. Natl. Acad. Sci. USA 1993, 90, 4508–4512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.; Fliegel, L. Amiloride and the Na(+)/H(+) exchanger protein: Mechanism and significance of inhibition of the Na(+)/H(+) exchanger (review). Int. J. Mol. Med. 1999, 3, 315–336. [Google Scholar] [CrossRef] [PubMed]
- Gaxiola, R.A.; Li, J.L.; Undurraga, S.; Dang, L.M.; Fink, G.R. Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. USA 2001, 98, 11444–11449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Aranda, M.R.; Fernández, P.G.; López-Tienda, J.R.; López-Diaz, M.R.; Espinosa, J.; Granum, E.; Traverso, J.Á.; Pineda, B.; Garcia-Sogo, B.; Moreno, V.; et al. Na+ transporter HKT1;2 reduces flower Na+ content and considerably mitigates the decline in tomato fruit yields under saline conditions. Plant Physiol. Biochem. 2020, 154, 341–352. [Google Scholar] [CrossRef]
- Wilkins, K.A.; Matthus, E.; Swarbreck, S.M.; Davies, J.M. Calcium-mediated abiotic stress signaling in roots. Front.Plant Sci. 2016, 7, 1296. [Google Scholar] [CrossRef] [Green Version]
- Manik, S.M.N.; Shi, S.J.; Mao, J.J.; Dong, L.H.; Su, Y.L.; Wang, Q.; Liu, H. The calcium sensor CBL-CIPK is involved in plant's response to abiotic stresses. Int. J. Genom. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bao, A.K.; Wang, S.M.; Wu, G.Q.; Xi, J.J.; Wang, C.M. Overexpression of the Arabidopsis H+-PPase enhanced resistance to salt and drought stress in transgenic alfalfa (Medicago sativa L.). Plant Sci. 2008, 176, 232–240. [Google Scholar] [CrossRef]
- Chen, J.; Wang, W.H.; Wu, F.H.; He, E.M.; Zheng, H.L. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci. Rep. 2015, 5, 12516. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhao, X.; Hu, Z.; Leng, P. Nitric oxide modulating ion balance in Hylotelephium erythrostictum roots subjected to NaCl stress based on the analysis of transcriptome, fluorescence, and ion fluxes. Sci. Rep. 2019, 9, 18137. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Sottosanto, J.B.; Blumwald, E. Vacuolar cation/H+ exchange, ion homeostasis, and leaf development are altered in a T-DNA insertional mutant of AtNHX1, the Arabidopsis vacuolar Na+/H+ antiporter. Plant J. 2003, 36, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Quintero, F.J.; Blatt, M.R.; Pardo, J.M. Functional conservation between yeast and plant endosomal Na(+)/H(+) antiporters. FEBS Lett. 2000, 471, 224–228. [Google Scholar] [CrossRef]
- Wu, C.A.; Yang, G.D.; Meng, Q.W.; Zheng, C.C. The cotton GhNHX1 gene encoding a novel putative tonoplast Na+/H+ antiporter plays an important role in salt stress. Plant Cell Physiol. 2004, 45, 600–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene Name | Gene ID | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|---|---|
| 1 | PgNHX01 | CDL15_Pgr020251 | AGTTGCTCGGAACCTTTCTC | CAGCATCATGAGAGCGACTT |
| 2 | PgNHX02 | CDL15_Pgr020284 | TGGGACATACTCATCTGCGA | CCATGCTGGTCCTATGCTTT |
| 3 | PgNHX03 | CDL15_Pgr013571 | TGTGATTGAACCTCCAGCAG | CTGTCCTTGCAATCGTCTCA |
| 4 | PgNHX04 | CDL15_Pgr008437 | AGCTCGTCAGTGATAGTCCA | TGATCTGTTGCTTCCAGTCG |
| 5 | PgNHX05 | CDL15_Pgr018608 | CTGATTATGGTGGGAAGGGC | TCCTGACCTCGTGAACTGAT |
| 6 | PgNHX06 | CDL15_Pgr016272 | CTTCCATCGTGACTGGACTG | GAGAGCCGTGATTGATTCGT |
| 7 | PgNHX07 | CDL15_Pgr004028 | GTTAGGCCTGCACATCGTAA | ACTATAGCTGTGGTAGCCGT |
| 8 | PgNHX08 | CDL15_Pgr019015 | TACTTTCGGCAACGGATTCA | GGCATCATTCACGACTCCTT |
| 9 | PgNHX09 | CDL15_Pgr012091 | TGTGCTCGATGCTCCATGTT | AGTCACGGCTTGCGTTCATA |
| 10 | PgNHX10 | CDL15_Pgr021015 | CACAGGCACTCTGTTTGTCT | CCAATATTCGCCTCTTCGCT |
| 11 | PgACTIN | CDL15_Pgr015157 | AGTCCTCTTCCAGCCATCTC | ACTGAGCACAATGTTTCCA |
| Gene Name | Protein ID | CDS (bp) | Length (aa) | MW (kDa) | pI | Instability Index | GRAVY | TMHs | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| PgNHX01 | OWM62957.1 | 1644 | 547 | 60.4 | 8.67 | 36.23 | 0.574 | 11 | Vacuole |
| PgNHX02 | OWM62990.1 | 1659 | 552 | 61.0 | 7.74 | 34.04 | 0.545 | 10 | Vacuole |
| PgNHX03 | OWM66354.1 | 2826 | 941 | 103.8 | 5.59 | 35.77 | 0.425 | 10 | Cell membrane; Vacuole |
| PgNHX04 | OWM74126.1 | 1632 | 543 | 60.6 | 9.21 | 45.44 | 0.479 | 12 | Vacuole |
| PgNHX05 | OWM78039.1 | 1617 | 538 | 59.5 | 9.32 | 35.22 | 0.571 | 10 | Vacuole |
| PgNHX06 | OWM78548.1 | 1626 | 541 | 60.4 | 8.17 | 39.49 | 0.476 | 10 | Vacuole |
| PgNHX07 | OWM83599.1 | 1647 | 548 | 60.0 | 5.45 | 43.64 | 0.401 | 10 | Vacuole |
| PgNHX08 | OWM85391.1 | 1653 | 550 | 60.3 | 8.32 | 36.53 | 0.546 | 10 | Vacuole |
| PgNHX09 | OWM85841.1 | 1419 | 472 | 52.6 | 6.94 | 35.40 | 0.577 | 11 | Vacuole |
| PgNHX10 | OWM90710.1 | 3582 | 1193 | 132.3 | 6.58 | 39.11 | 0.103 | 10 | Cell membrane |
| Protein | C-Score | TM-Score | RMSD (Å) | Best Identified Structure Analogs in PDB | ||||
|---|---|---|---|---|---|---|---|---|
| PDB Hit | TM-Scorea | RMSDa | IDENa | Cov | ||||
| PgNHX01 | −1.69 | 0.51 ± 0.15 | 11.5 ± 4.5 | 4cz8A | 0.707 | 1.06 | 0.209 | 0.717 |
| PgNHX02 | −1.21 | 0.56 ± 0.15 | 10.4 ± 4.6 | 4cz8A | 0.701 | 1.08 | 0.227 | 0.710 |
| PgNHX03 | −0.66 | 0.63 ± 0.14 | 10.3 ± 4.6 | 6xteA | 0.925 | 0.96 | 0.089 | 0.932 |
| PgNHX04 | −1.50 | 0.53 ± 0.15 | 11.0 ± 4.6 | 4cz8A | 0.702 | 1.50 | 0.210 | 0.720 |
| PgNHX05 | −1.67 | 0.51 ± 0.15 | 11.5 ± 4.5 | 4cz8A | 0.697 | 1.93 | 0.204 | 0.729 |
| PgNHX06 | −1.80 | 0.50 ± 0.15 | 11.8 ± 4.5 | 4cz8A | 0.702 | 1.51 | 0.223 | 0.723 |
| PgNHX07 | −1.48 | 0.53 ± 0.15 | 11.0 ± 4.6 | 6z3yB | 0.690 | 1.31 | 0.402 | 0.704 |
| PgNHX08 | −0.81 | 0.61 ± 0.14 | 9.4 ± 4.6 | 4cz8A | 0.702 | 1.17 | 0.212 | 0.713 |
| PgNHX09 | −1.92 | 0.48 ± 0.15 | 11.8 ± 4.5 | 6z3yB | 0.742 | 1.38 | 0.301 | 0.761 |
| PgNHX10 | −0.11 | 0.70 ± 0.12 | 9.6 ± 4.6 | 6r9tA | 0.986 | 0.87 | 0.105 | 0.991 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, J.; Liu, C.; Wang, Y.; Zhao, Y.; Ge, D.; Yuan, Z. Genome-Wide Identification of the NHX Gene Family in Punica granatum L. and Their Expressional Patterns under Salt Stress. Agronomy 2021, 11, 264. https://doi.org/10.3390/agronomy11020264
Dong J, Liu C, Wang Y, Zhao Y, Ge D, Yuan Z. Genome-Wide Identification of the NHX Gene Family in Punica granatum L. and Their Expressional Patterns under Salt Stress. Agronomy. 2021; 11(2):264. https://doi.org/10.3390/agronomy11020264
Chicago/Turabian StyleDong, Jianmei, Cuiyu Liu, Yuying Wang, Yujie Zhao, Dapeng Ge, and Zhaohe Yuan. 2021. "Genome-Wide Identification of the NHX Gene Family in Punica granatum L. and Their Expressional Patterns under Salt Stress" Agronomy 11, no. 2: 264. https://doi.org/10.3390/agronomy11020264
APA StyleDong, J., Liu, C., Wang, Y., Zhao, Y., Ge, D., & Yuan, Z. (2021). Genome-Wide Identification of the NHX Gene Family in Punica granatum L. and Their Expressional Patterns under Salt Stress. Agronomy, 11(2), 264. https://doi.org/10.3390/agronomy11020264