
Allelopathic Potential of Haloxylon persicum against Wheat and Black Mustard with Special Reference to Its Phytochemical Composition and Antioxidant Activity


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Extraction and Preparation of H. persicum Aqueous Extract
2.3. In Vitro Germination Experiment
2.4. Biochemical Characteristics of Wheat Seedlings
2.4.1. Determination of Carbohydrates
2.4.2. Determination of Free Proline
2.4.3. Determination of Photosynthetic Pigments
2.5. Phytochemical Analysis of H. persicum Shoots for Secondary Metabolites
2.5.1. Plant Extraction
2.5.2. Determination of Total Phenolics (TP)
2.5.3. Determination of Total Flavonoids (TF)
2.5.4. Determination of Total Flavonols
2.5.5. Determination of Total Anthocyanins
2.5.6. Determination of Total Condensed Tannins
2.5.7. Determination of Total Saponins
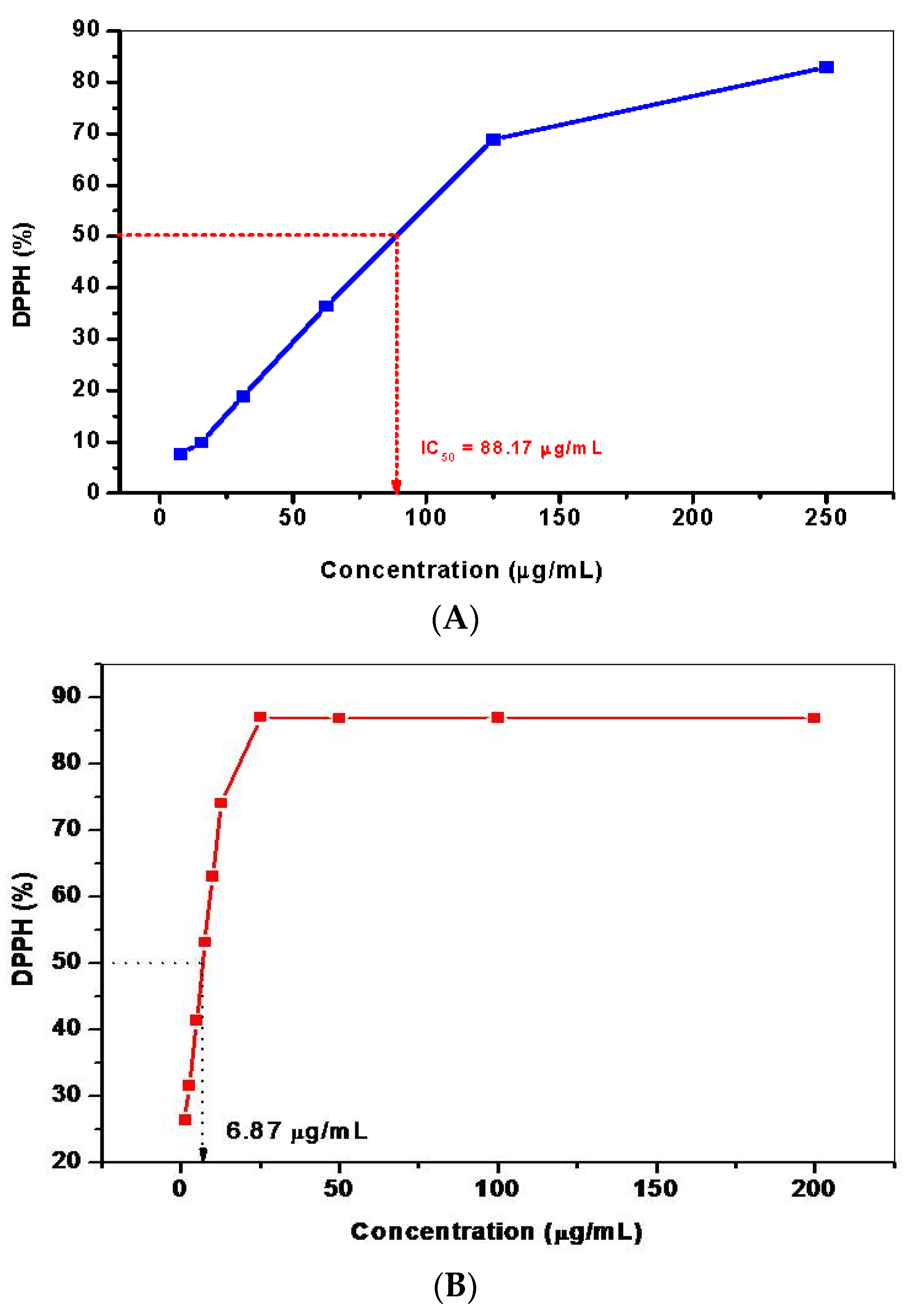
2.5.8. Determination of 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Free Radical Scavenging Activity
2.6. Statistical Analysis
3. Results and Discussion
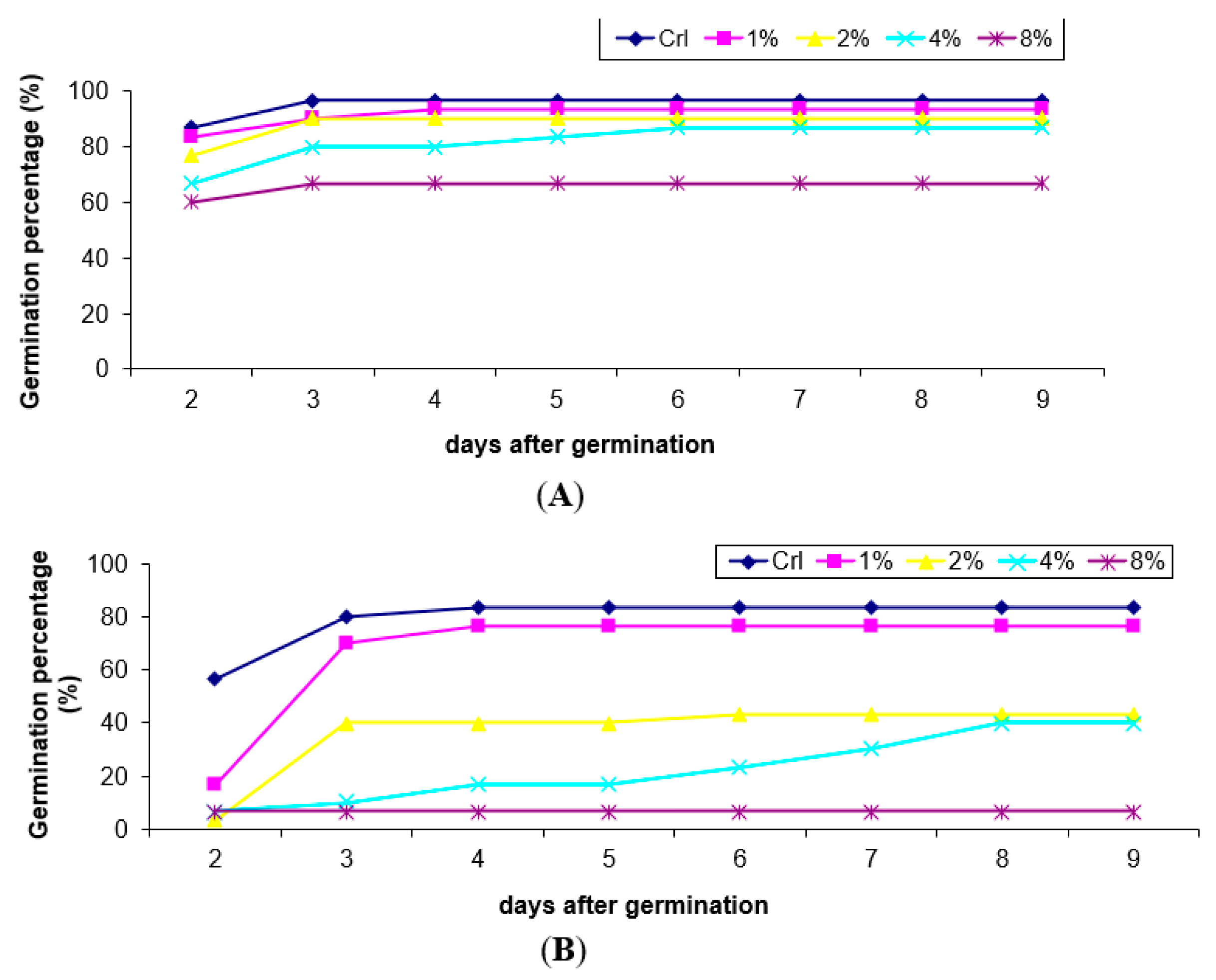
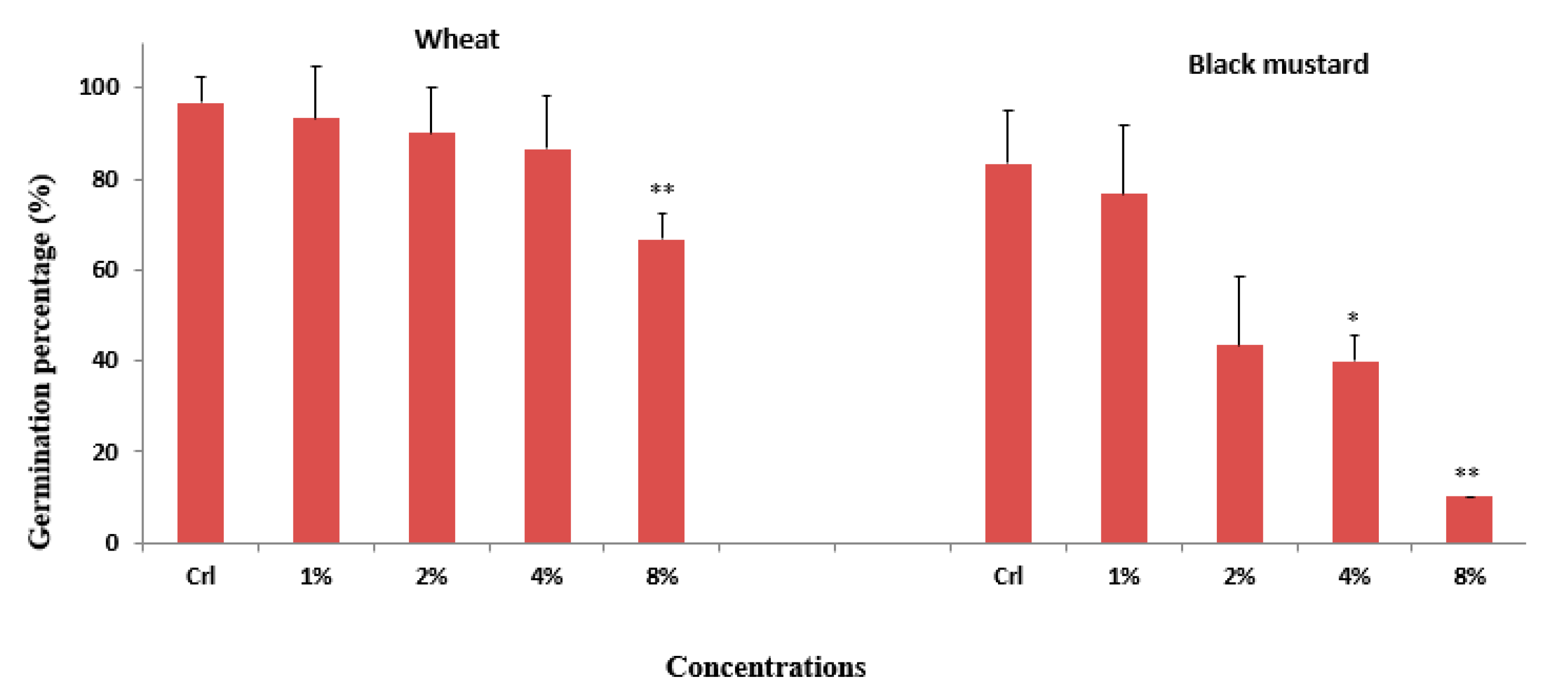
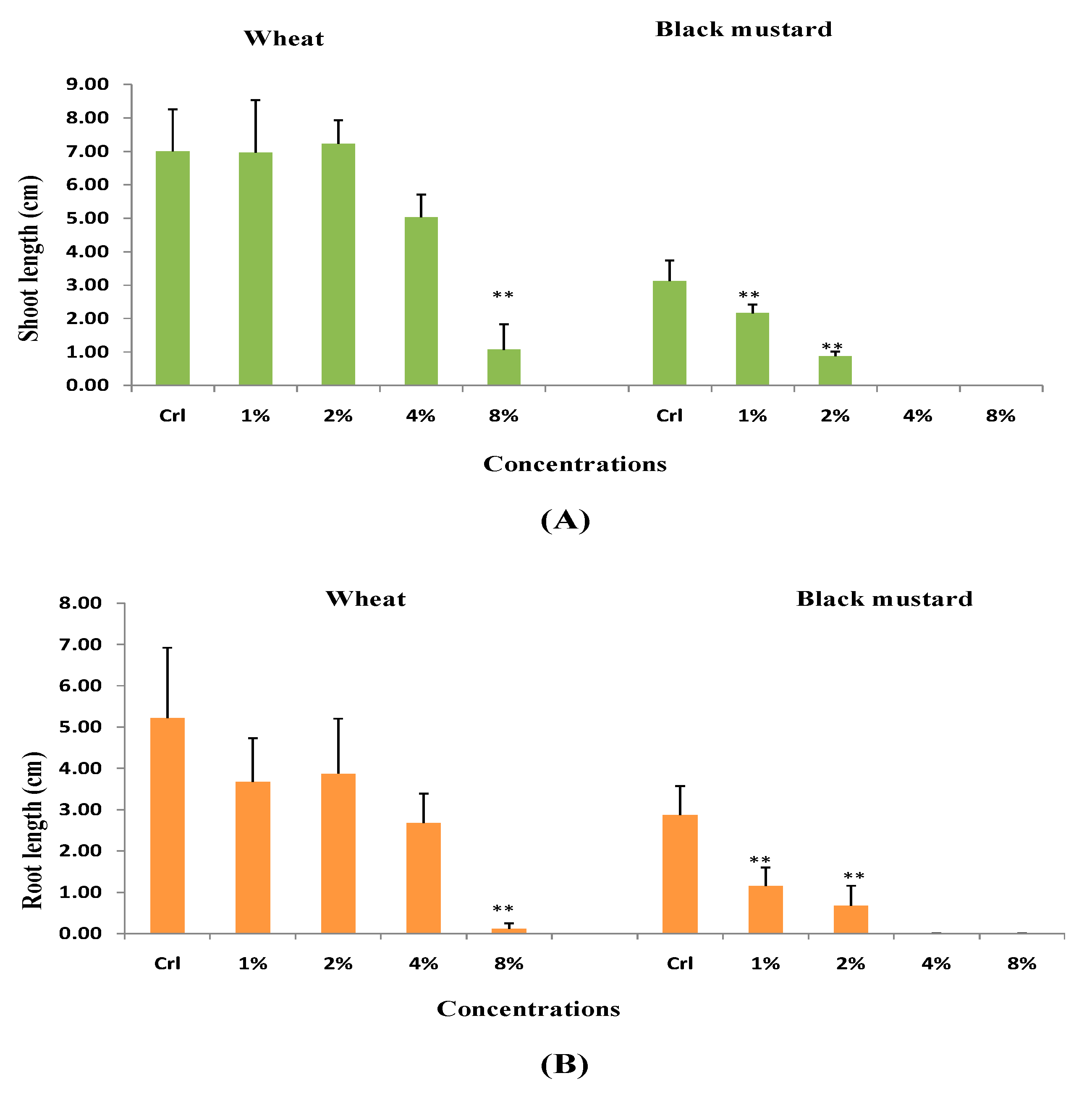
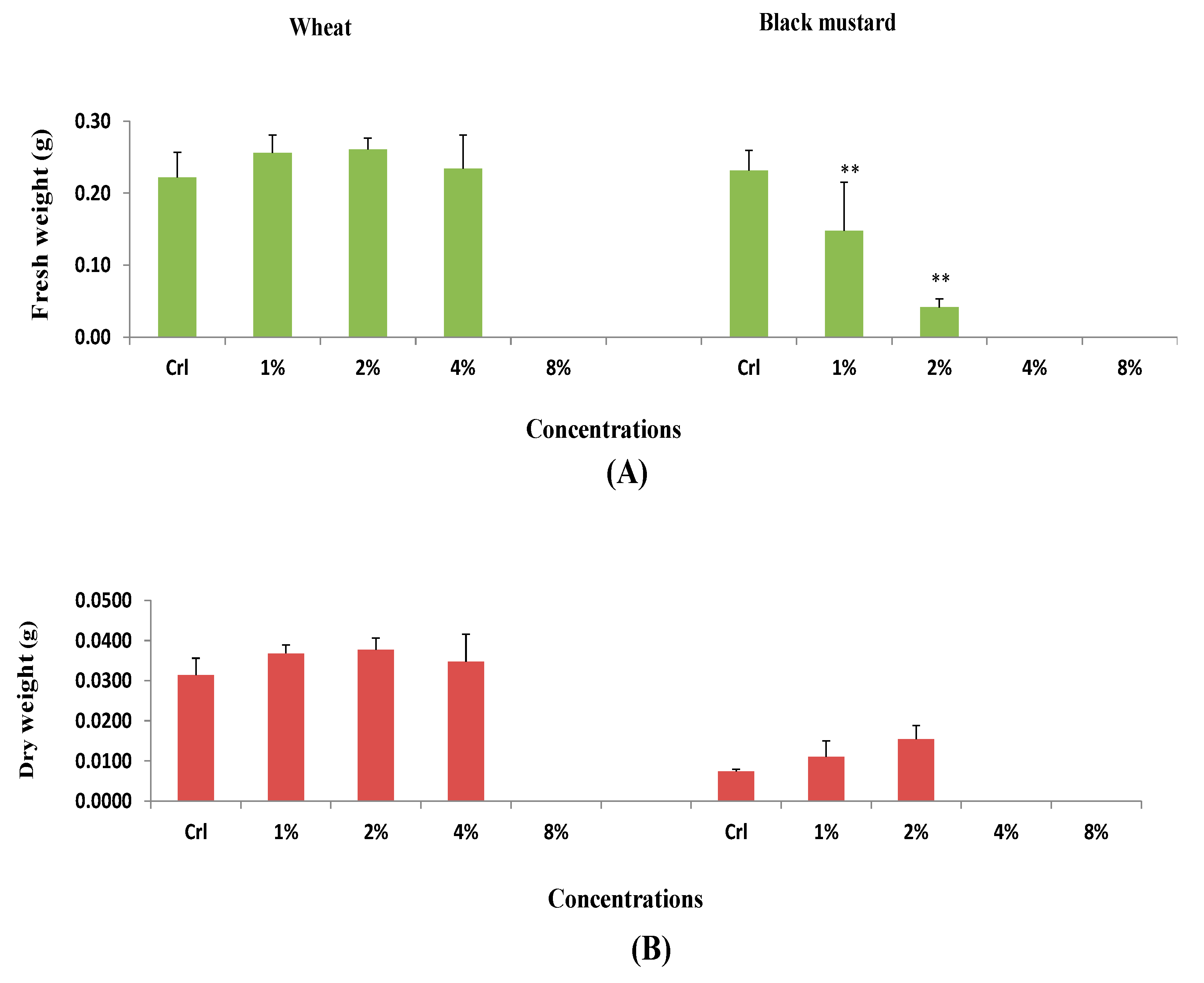
3.1. Effect of H. persicum Aqueous Extract on Seed Germination and Seedling Growth of Wheat and Black Mustard
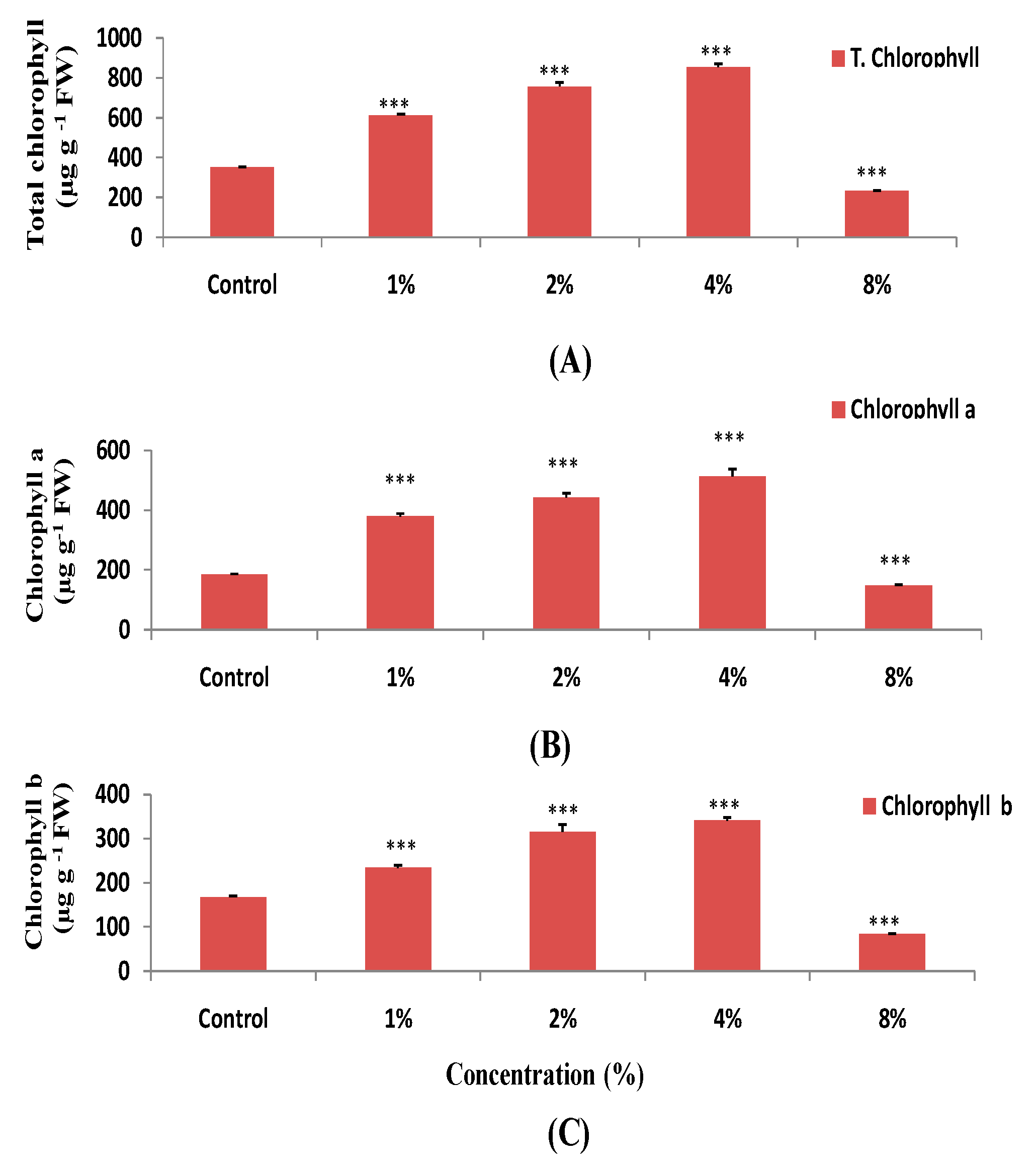
3.2. Effect of H. persicum on the Photosynthetic Pigments Content in Wheat
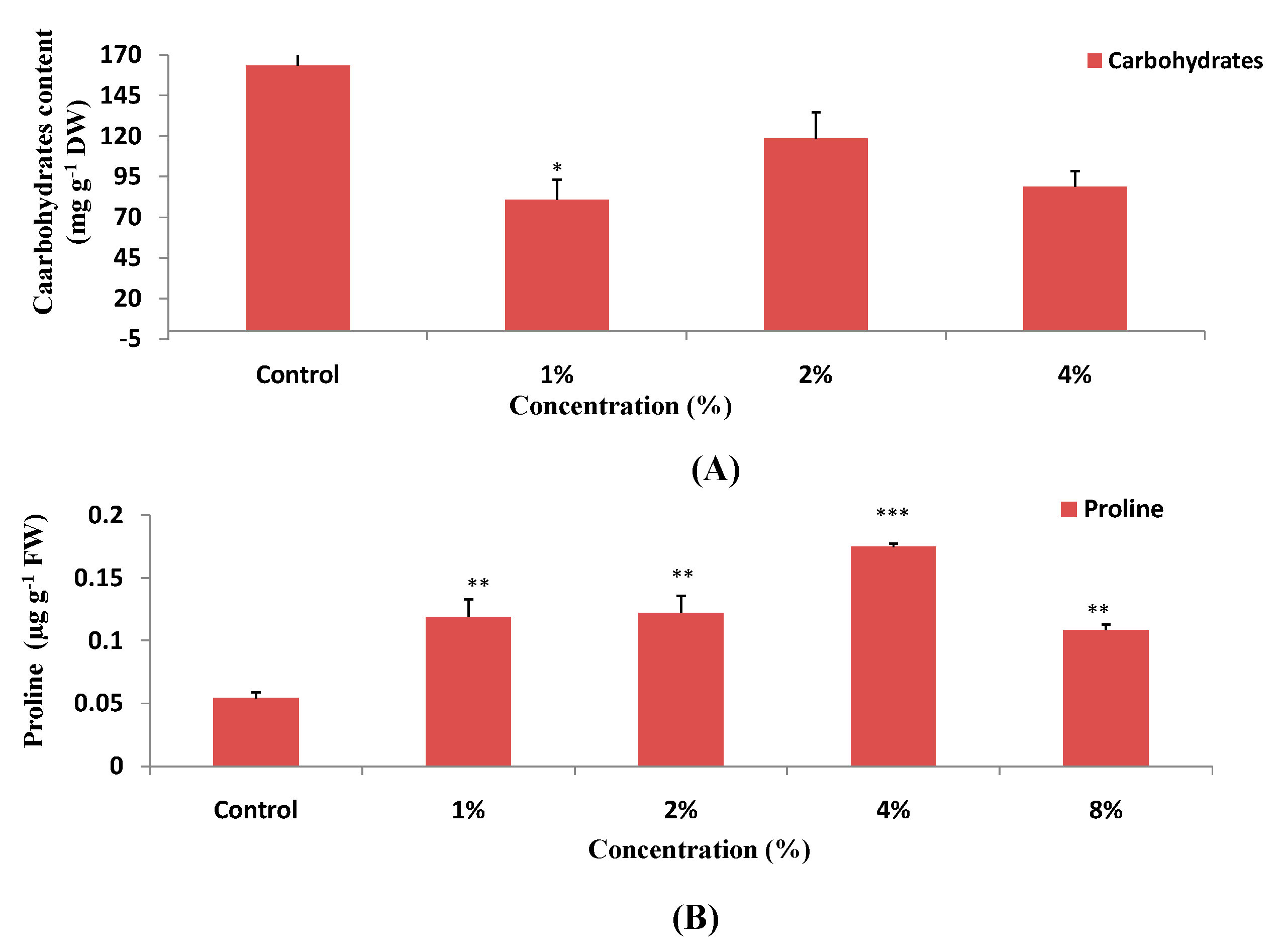
3.3. Effect of H. persicum on Carbohydrates and Proline Contents in Wheat
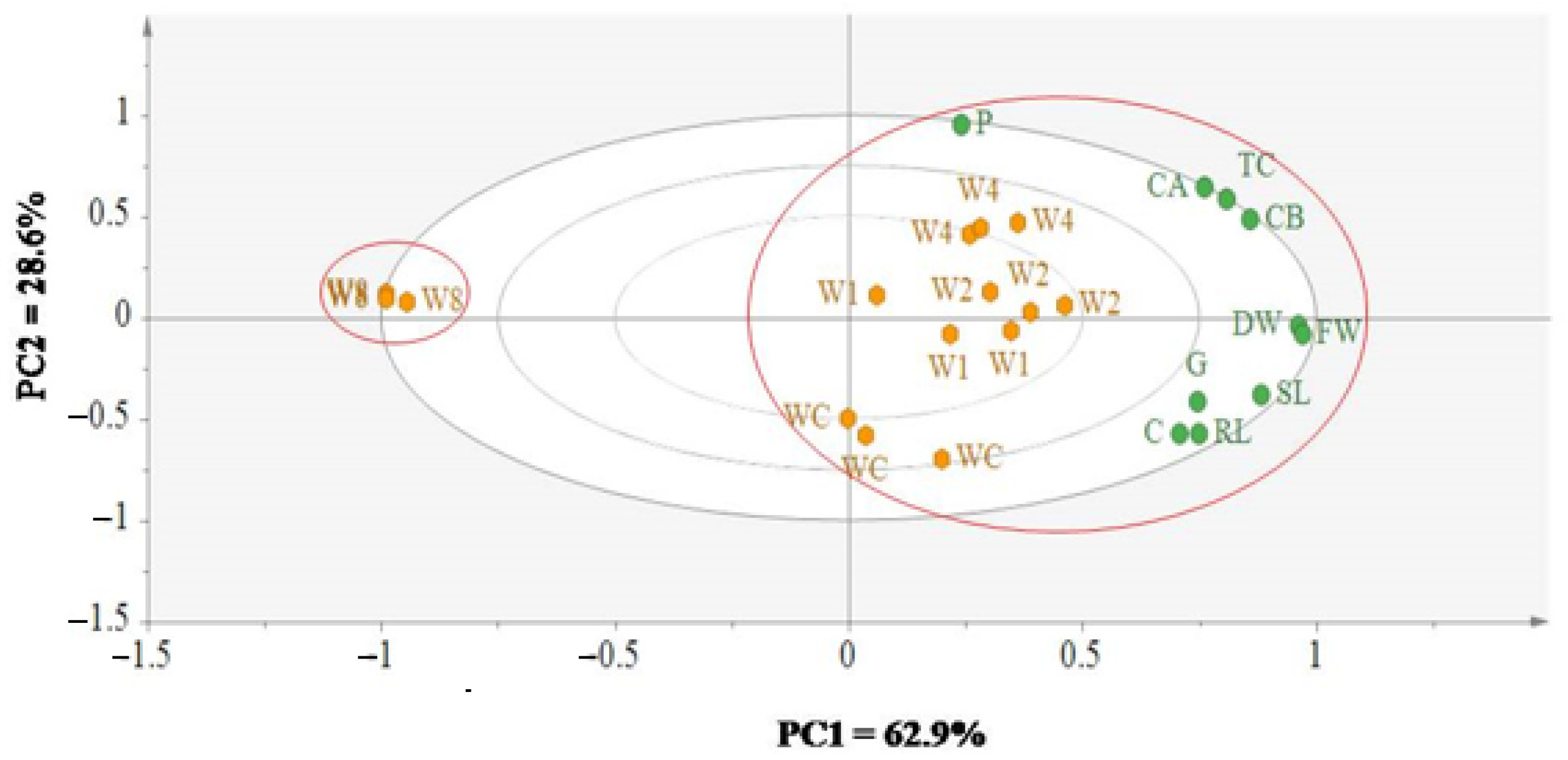
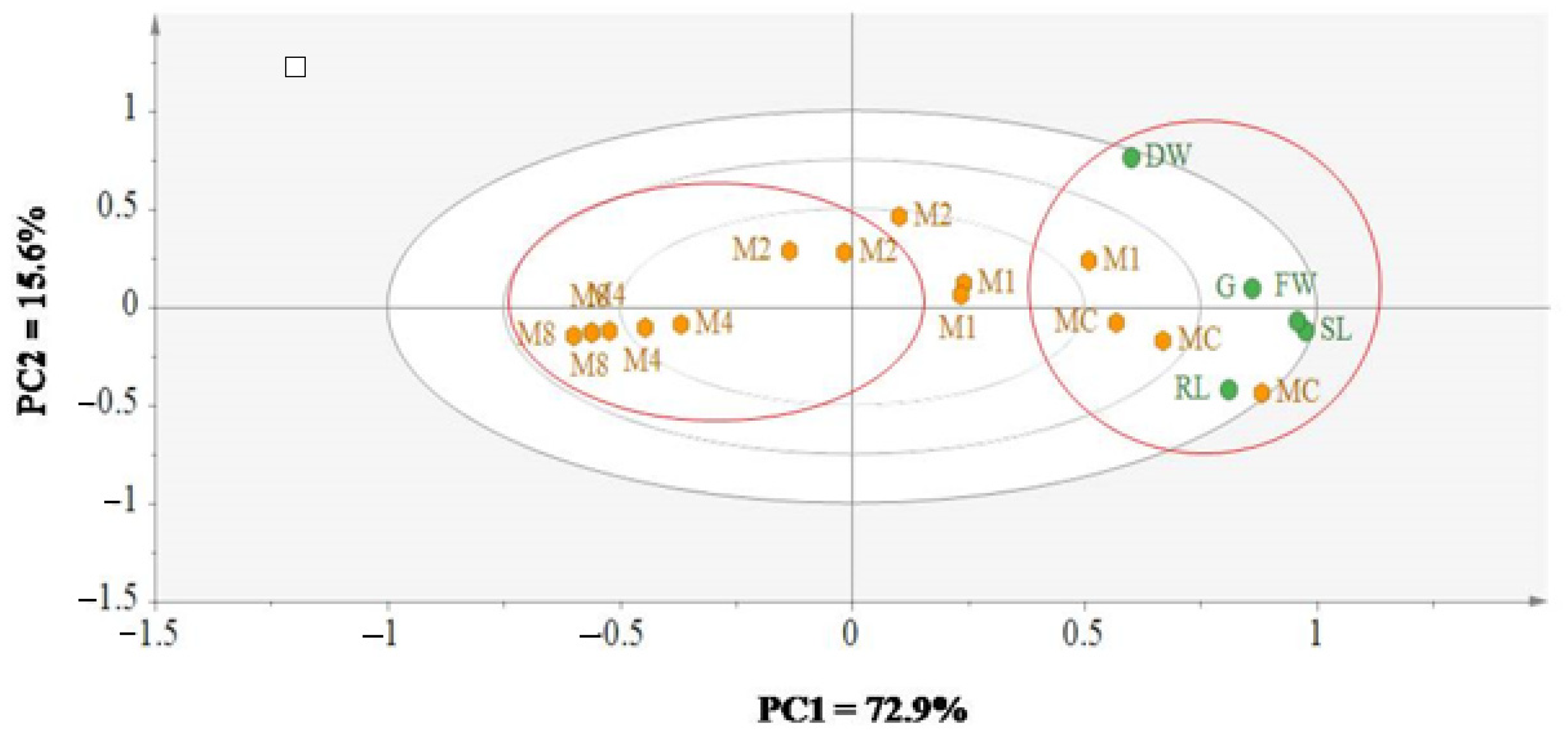
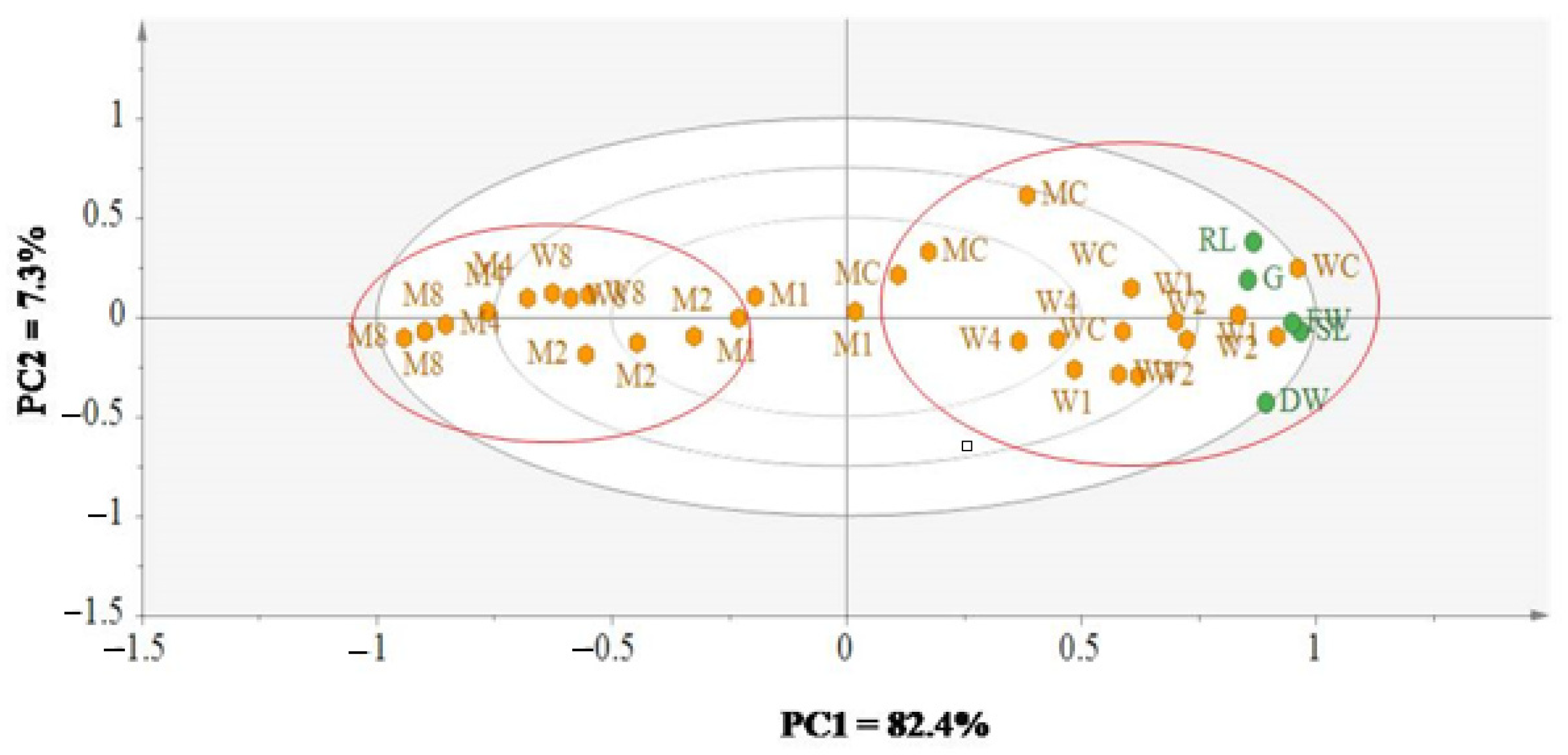
3.4. Dissecting the Effect of Allelochemical Stress on Wheat and Black Mustard Using Multivariate Data Analysis
3.5. Phytochemical Analysis and Antioxidant Activity of H. persicum
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Patterson, D.T. Allelopathy. In Research Methods in Weed Science, 3rd ed.; Camper, N.D., Ed.; Southern Weed Science Society: Champaign, IL, USA, 1986; pp. 111–134. [Google Scholar]
- Zeng, R.S. Allelopathy in Chinese ancient and modern agriculture. In Allelopathy in Suitable Agriculture and Forestry; Zeng, R.S., Mallik, A.Z., Luo, S.M., Eds.; Springer: New York, NY, USA, 2008; pp. 39–59. [Google Scholar]
- Korres, N.E.; Norsworthy, J.K.; Tehranchian, P.; Gitsopoulos, T.K.; Loka, D.A.; Oosterhuis, D.M.; Gealy, D.R.; Moss, R.S.; Burgos, N.R.; Miller, M.R.; et al. Cultivars to face climate change effects on crops and weeds: A review. Agron. Sustain. Dev. 2016, 36, 12. [Google Scholar] [CrossRef] [Green Version]
- Zohaib, A.; Tabassum, T.; Anjum, S.A.; Abbas, T.; Nazir, U. Allelopathic effect of some associated weeds of wheat on germinability and biomass production of wheat seedlings. Plant Dan. 2017, 35, 1–12. [Google Scholar] [CrossRef]
- Han, X.; Cheng, Z.; Meng, H.; Yang, X.; Ahmad, I. Allelopathic effect of decomposed garlic (Allium sativum L.) stalk on lettuce (L. sativa var. crispa L.). Pak. J. Bot. 2013, 45, 225–233. [Google Scholar]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Kohli, R.K.; Batish, D.R.; Singh, H.P. Eucalyptus oils for the control of Parthenium (Parthenium hysterophorus L.). Crop Prot. 1998, 17, 119–122. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy.Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Sarfraz, M.; Ahmad, M.; Noor, W.S.A.W.M.; Ashraf, M.A. Pollution Prevention, Best Management Practices, and Conservation. In Organic Farming: A Promising Way of Food Production; InTech: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Bhowmik, P.C.; Inderjit. Challenges and opportunities in implementing allelopathy for natural weed management. Crop Prot. 2003, 22, 661–671. [Google Scholar] [CrossRef]
- Rice, E.L. Biological Control of Weeds and Plant Diseases: Advances in Applied Allelopathy; Oklahoma University Press: Norman, OK, USA, 1995; Available online: https://lib.ugent.be/catalog/rug01:000351620 (accessed on 1 December 2020).
- Torres, A.; Oliva, R.M.; Castellano, D.; Cross, P. First World Congress on Allelopathy—A Science of the Future, Cadiz, Spain 16–20 September 1996; University of Cadiz: Cadiz, Spain, 1996. [Google Scholar]
- Kruse, M.; Strandberg, M.; Strandberg, B. Ecological Effects of Allelopathic Plants—A Review; NERI Technical Report; National Environmental Research Institute: Silkeborg, Denmark, 2000. [Google Scholar]
- Oleszek, W.; Jurzysta, M. The allelopathic potential of alfalfa root medicagenic acid glycosides and their fate in soil environments. Plant Soil 1987, 98, 67–80. [Google Scholar] [CrossRef]
- Edewor, T.I.; Ibikunle, G.J.; Usman, L.A. Phytotoxic and antimicrobial screening of saponin isolated from ethanolic leaf extract of Xylopia aethipioca. Sci. Focus 2009, 14, 507–512. [Google Scholar]
- Bialy, Z.; Oleszek, W.; Lewis, J.; Fenwick, G.R. Allelopathic potential of glucosinolates (mustard oil glycosides) and their degradation products against wheat. Plant Soil 1990, 129, 277–281. [Google Scholar] [CrossRef]
- Abdel-Farid, I.B.; Sheded, M.G.; Mohamed, E.A. Metabolomic profiling and antioxidant activity of some Acacia species. Saudi J. Biol. Sci. 2014, 21, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.R.; Amist, N.; Bai, L.Y. Allelopathy in sustainable weeds management. Allelopathy J. 2019, 48, 109–138. [Google Scholar] [CrossRef]
- Al-Wakeel, S.A.; Gabr, M.A.; Hamid, A.A.; Abu-El-Soud, W.M. Allelopathic effects of Acacia nilotica leaf residue on Pisum sativum L. J. Allelopathy 2007, 19, 411–421. [Google Scholar]
- Kobayashi, K. Factors affecting phytotoxic activity of allelochemicals in soil. Weed Biol. Manag. 2004, 4, 1–7. [Google Scholar] [CrossRef]
- Haig, T. Allelochemicals in Plants. In Allelopathy in Sustainable Agriculture and Forestry; Zeng, R.S., Mallik, A.U., Luo, S.M., Eds.; Springer: New York, NY, USA, 2008; pp. 63–104. [Google Scholar] [CrossRef]
- Moameri, M.; Khalaki, M.A.; Tavili, A. Effects of Haloxylon aphyllum (Minkw.) lljin extract on seeds germination and seedlings growth of Agropyron elongatum (Host.) and Agropyron desertorum (Fisch.). Res. J. Seed Sci. 2011, 4, 40–50. [Google Scholar] [CrossRef]
- Gomaa, N.H.; Hassan, M.O.; Fahmy, G.M.; González, L.; Hammouda, O.; Atteya, A.M. Allelopathic effects of Sonchus oleraceus L. on the germination and seedling growth of crop and weed species. Acta Bot. Braz. 2014, 28, 408–416. [Google Scholar] [CrossRef]
- El-Khatib, A.A.; Barakat, N.A.; Nazeir, H. Growth and physiological response of some cultivated species under allelopathic stress of Calotropis procera (Aiton) W.T. Appl. Sci. Rep. 2016, 14, 237–246. [Google Scholar] [CrossRef]
- Gomaa, N.H.; AbdElgawad, H.R. Phytotoxic effects of Echinochloa colona (L.) Link. (Poaceae) extracts on the germination and seedling growth of weeds. Span. J. Agric. Res. 2012, 10, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Farid, I.B.; El-Sayed, M.A.; Mohamed, E.A. Allelopathic potential of Calotropis procera and Morettia philaeana. Int. J. Agric. Biol. 2013, 15, 120–134. [Google Scholar]
- Hozayn, M.; El-Shahawy, T.A.; Abd El-Monem, A.A.; El-Saady, A.A.; Darwish, M.A. Allelopathic effect of Casuarina equisetifolia L. on wheat, associated weeds and nutrient content in the soil. Afr. J. Agric. Res. 2015, 10, 1675–1683. [Google Scholar] [CrossRef] [Green Version]
- Sheded, M.; Jahangir, M.; Marghany, M.; Abdel-Farid, I.B. Effect of Casuarina allelochemicals on growth of cultivated and weed plants. J. Bio-Mol. Sci. 2017, 5, 1–14. [Google Scholar]
- Ghimire, B.K.; Ghimire, B.; Yu, C.Y.; Chung, I.M. Allelopathic and autotoxic effects of Medicago sativa—Derived allelochemicals. Plants 2019, 8, 233. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, D.; Rinzim; Tiwari, A.; Sehgal, A.; Landi, M.; Brestic, M.; Sharma, A. Exploiting the allelopathic potential of aqueous leaf extracts of Artemisia absinthium and Psidium guajava against Parthenium hysterophorus, a widespread weed in India. Plants 2019, 8, 552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [Green Version]
- Alghanem, S.M. Ecological and botanical diversity in Haloxylon Persicum community at Al-Qassim Region in Kingdom of Saudi Arabia. Am. J. Environ. Prot. 2018, 6, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Song, J.I.E.; Feng, G.U.; Tian, C.; Zhang, F. Strategies for adaptation of Suaeda physophora, Haloxylon ammodendron and Haloxylon persicum to a saline environment during seed-germination stage. Ann. Bot. 2005, 96, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Al-Khalifah, N.S.; Shanavaskhan, A.E. On the distribution, status and phenology of Ghada (Haloxylon persicum Bunge) in the Arabian peninsula. Trop. Ecol. 2007, 48, 51–60. [Google Scholar]
- Xu, G.; Yu, D.; Xie, J.; Tang, L.; Li, Y. What makes Haloxylon persicum grow on sand dunes while H. ammodendron grows on interdune lowlands: A proof from reciprocal transplant experiments. J. Arid Land 2014, 6, 581–591. [Google Scholar] [CrossRef]
- El-Khatib, A.A.; Abd-Elaah, G.A. Allelopathic potential of Zilla spinosa on growth of associate flowering plants and some rhizosphere fungi. Biol. Plant. 1998, 41, 461–467. [Google Scholar] [CrossRef]
- Grange, S.L.; Leskovar, D.I.; Pike, L.M.; Cobb, B.G. Excess moisture and seed coat nicking influence germination of triploid watermelon. HortScience 2000, 35, 1355–1356. [Google Scholar] [CrossRef]
- Zucconi, F.; Forte, M.; Monaco, A.D.E.; De Bertoldi, M. Biological evaluation of compost maturity. BioCycleb 1981, 22, 27–29. [Google Scholar]
- Abdel-Farid, I.B.; Marghany, M.R.; Rowezek, M.M.; Sheded, M.G. Effect of salinity stress on growth and metabolomic profiling of Cucumis sativus and Solanum lycopersicum. Plants 2020, 9, 1626. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L. Quantitative determination of carbohydrates with Dreywood’santhrone reagent. Science 1948, 107, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dere, S.; Güneş, T.; Sivaci, R. Spectrophotometric determination of chlorophyll A, B and total carotenoid contents of some algae species using different solvents. Tur. J. Bot. 1998, 22, 13–17. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-ciocalteau reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Kumaran, A.; Karunakaran, R.J. In vitro antioxidant activities of methanol extracts of five Phyllanthus species from India. LWT Food Sci. Technol. 2007, 40, 344–352. [Google Scholar] [CrossRef]
- Padmavati, M.; Sakthivel, N.; Thara, K.V.; Reddy, A.R. Differential sensitivity of rice pathogens to growth inhibition by flavonoids. Phytochemistry 1997, 46, 499–502. [Google Scholar] [CrossRef]
- Julkunen-Titto, R. Phenolics constituents in the leaves of northern Willows. Methods for the analysis of certain phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Ebrahimzadeh, H.; Niknam, V.A. revised spectrophotometric method for determination of triterpenoid saponins. Indian Drugs 1998, 35, 379–381. [Google Scholar]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Wang, J.; Mazza, G. : Effect of anthocyanins and other phenolic compounds on the production of tumor necrosis factor alpha in LPS/IFN-gamma- activated RAW 264.7 macrophages. J. Agric. Food. Chem. 2002, 50, 4183–4189. [Google Scholar] [CrossRef]
- Hegazy, A.K.; Fadl-Allah, E.M. Inhibition of seed germination and seedling growth by Cleome droserifolia and allelopathic effect on rhizosphere fungi in Egypt. J. Arid. Environ. 1995, 29, 3–11. [Google Scholar] [CrossRef]
- Reigosa, M.J.; Gonzalez, L.; Sanches-Moreiras, A.; Duran, B.; Puime, D.; Fernandez, D.A.; Bolano, J.C. Comparison of physiological effects of allelochemicals and commercial herbicides. Allelopathy J. 2001, 8, 211–220. [Google Scholar]
- Sisodia, S.; Siddiqui, M.B. Allelopathic effect by aqueous extracts of different parts of Croton bonplandianum Baill. on some crop and weed plants. J. Agric. Exten. Rural Dev. 2010, 2, 22–28. [Google Scholar]
- Tanveer, A.; Jabbar, M.K.; Kahliq, A.; Matloob, A.; Abbas, R.; Javaid, M.M. Allelopathic effects of aqueous and organic fractions of Euphorbia dracunculoides Lam. on germination and seedling growth of chickpea and wheat. Chil. J. Agric. Res. 2012, 72, 495–501. [Google Scholar] [CrossRef]
- Mohamed, E.A. Metabolomic Profiling, Allelopathic Potential, Antioxidant Capacity and Biological Activity of Some Acacia Species. Master’s Thesis, Aswan University, Aswan, Egypt, 2015. [Google Scholar]
- Fritz, D.; Bernardi, A.P.; Haas, J.S.; Ascoli, B.M.; Bordignon, S.A.D.L.; Von Poser, G. Germination and growth inhibitory effects of Hypericum myrianthum and H. polyanthemum extracts on Lactuca sativa L. Rev. Bras. Farmacog. 2007, 17, 44–48. [Google Scholar] [CrossRef]
- Hussain, M.I.; El-Sheikh, M.A.; Reigosa, M.J. Allelopathic potential of aqueous extract from Acacia melanoxylon R. Br. on Lactuca sativa. Plants 2020, 9, 1228. [Google Scholar] [CrossRef] [PubMed]
- Politycka, B. Phenolics and the activities of phenylalanine ammonia-lyase, phenol-β-glucosyltransferase and β-glucosidase in cucumber roots as affected by phenolic allelochemicals. Acta Physiol. Plant. 1998, 20, 405–410. [Google Scholar] [CrossRef]
- Omezzine, F.; Ladhari, A.; Haouala, R. Physiological and biochemical mechanisms of allelochemicals in aqueous extracts of diploid and mixoploid Trigonella foenum-graecum L. S. Afr. J. Bot. 2014, 93, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; Aschehoug, E.T. Invasive plant versus their new and old neighbours: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Al-Zahrani, H.S.; Al-Robai, S.A. Allelopathic effect of Calotropis procera leaves extract on seed germination of some plants. JKAU Sci. 2007, 19, 115–126. [Google Scholar] [CrossRef]
- Kamal, J. Impact of allelopathy of sunflower (Helianthus annuus L.) roots extract on physiology of wheat (Triticum aestivum L.). Afr. J. Biotechnol. 2011, 10, 14465–14477. [Google Scholar] [CrossRef] [Green Version]
- Qasem, J.R. The allelopathic effect of three Amaranthus spp. (Pigweeds) on wheat (Triticum durum). Weed Res. 1995, 35, 41–49. [Google Scholar] [CrossRef]
- Turk, M.A.; Lee, K.D.; Tawaha, A.M. Inhibitory effects of aqueous extracts of black mustard on germination and growth of radish. Res. J. Agric. Biol. Sci. 2005, 1, 227–231. [Google Scholar]
- Hassan, S.M.; Ghareib, H.R. Bioactivity of Ulva lactuca L. acetone extract on germination and growth of lettuce and tomato plants. Afr. J. Biotechnol. 2009, 8, 3832–3838. [Google Scholar] [CrossRef]
- Abou El-Ghit, H.M. Physiological allelopathic effect of aqueous extracts of cucumber, carrot, onion, and garlic seeds on germination and growth of Pea. J. Pharma. Chem. Biol. Sci. 2016, 4, 13–19. [Google Scholar]
- Siyar, S.; Majeed, A.; Muhammad, Z.; Ali, H.; Inayat, N. Allelopathic effect of aqueous extracts of three weed species on the growth and leaf chlorophyll content of bread wheat. Acta Ecol. Sin. 2019, 39, 63–68. [Google Scholar] [CrossRef]
- Salama, H.M.; Al Rabiah, H.K. Physiological effects of allelopathic activity of Citrullus colocynthis on Vicia faba and Hordeum vulgare. Eur. J. Biol. Res. 2015, 5, 25–35. [Google Scholar]
- Hamad, S.; Mahgoub, A.M. Allelopathic potentials of Artemisia monosperma and Thymus vulgaris on growth and physio-biochemical characteristics of Pea seedlings. Pak. J. Biol. Sci. 2018, 21, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Romman, S. Differential allelopathic expression of different plant parts of Achillea biebersteinii. Acta Biol. Hung. 2016, 67, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Ali, A.; Cheng, Z. An allelopathic role for garlic root exudates in the regulation of carbohydrate metabolism in cucumber in a hydroponic co-culture system. Plants 2020, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Heuer, B. Role of proline in plant response to drought and salinity. In Handbook of Plant and Crop Stress; CRC Press: Boca Raton, FL, USA, 2010; pp. 213–238. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plants Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.; Tiwari, A.; Saharawat, Y.S.; McDonald, A.J. Proline content as a stress indicator to quantify conservation agriculture effect in wheat crop. Res. Crops 2015, 16, 422–431. [Google Scholar] [CrossRef]
- Siddique, A.; Kandpal, G.; Kumar, P. Proline accumulation and its defensive role under diverse stress condition in plants: An overview. J. Pure Appl. Microbiol. 2018, 12, 1655–1659. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Patakas, A.; Noitsakis, B. Leaf age effects on solute accumulation in water-stressed grapevines. J. Plant Physiol 2001, 158, 63–69. [Google Scholar] [CrossRef]
- Alshamsi, M.M.; Alnuaimi, A.K.; Alshamsi, R.M.; Senthilkumar, A.; Karthishwaran, K.; Al-Zayadneh, W.A.; Cheruth, A.J. Phytochemical analysis and antioxidant activity of Haloxylon salicarnicum, Ochradenus arabicus and Tamarix nilotica. J. Phytol. 2019, 11, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Korat Avni, N.; Kavathiya, Y.A.; Lakhani, N.S.; Kandoliya, U.K.; BA, G.; Golakiya, B.A. Nutritional and antioxidant components of fenugreek (Trigonella foenum-graecum L.) seedlings. J. Pharm. Phytochem. 2019, 8, 443–447. [Google Scholar]
- Miyamae, Y.; Kurisu, M.; Han, J.; Isoda, H.; Shigemori, H. Structure–activity relationship of caffeoylquinic acids on the accelerating activity on ATP production. Chem. Pharm. Bull. 2011, 59, 502–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, A.M. Influence of rutin on biochemical alterations in hyperammonemia in rats. Exp. Toxicol. Pathol. 2012, 64, 783–789. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Metabolites | Concentration |
|---|---|
| Total phenolics (TP) mg GAE g−1 extract | 46.02 ± 0.21 |
| Total flavonoids (TF) mg QE g−1 extract | 37.05 ± 0.81 |
| Total flavonols mg QE g−1 extract | 9.3 ± 0.68 |
| Total anthocyanins µmole g−1 extract | 0.57 ± 0.02 |
| Total condensed tannins mg CE g−1 extract | 59.73 ± 1.28 |
| Total saponins mg SE g−1 extract | 143.3 ± 1.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Farid, I.B.; Massoud, M.S.; Al-Enazy, Y.; Abdel Latef, A.A.H.; Jahangir, M.; Gomaa, N.H. Allelopathic Potential of Haloxylon persicum against Wheat and Black Mustard with Special Reference to Its Phytochemical Composition and Antioxidant Activity. Agronomy 2021, 11, 244. https://doi.org/10.3390/agronomy11020244
Abdel-Farid IB, Massoud MS, Al-Enazy Y, Abdel Latef AAH, Jahangir M, Gomaa NH. Allelopathic Potential of Haloxylon persicum against Wheat and Black Mustard with Special Reference to Its Phytochemical Composition and Antioxidant Activity. Agronomy. 2021; 11(2):244. https://doi.org/10.3390/agronomy11020244
Chicago/Turabian StyleAbdel-Farid, Ibrahim Bayoumi, Mohamed Salah Massoud, Yahia Al-Enazy, Arafat Abdel Hamed Abdel Latef, Muhammad Jahangir, and Nasr H. Gomaa. 2021. "Allelopathic Potential of Haloxylon persicum against Wheat and Black Mustard with Special Reference to Its Phytochemical Composition and Antioxidant Activity" Agronomy 11, no. 2: 244. https://doi.org/10.3390/agronomy11020244
APA StyleAbdel-Farid, I. B., Massoud, M. S., Al-Enazy, Y., Abdel Latef, A. A. H., Jahangir, M., & Gomaa, N. H. (2021). Allelopathic Potential of Haloxylon persicum against Wheat and Black Mustard with Special Reference to Its Phytochemical Composition and Antioxidant Activity. Agronomy, 11(2), 244. https://doi.org/10.3390/agronomy11020244