
Effects of Manure and Chemical Fertilizer on Bacterial Community Structure and Soil Enzyme Activities in North China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Description and Experimental Design
2.2. Soil Sampling and Analysis of Soil Properties
2.3. Soil Physicochemical Analytical Procedures
2.4. Soil Enzyme Activities’ Determination
2.5. Soil DNA Extraction and High-Throughput Sequencing
2.6. Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
3.1. Soil Physicochemical Characteristics
3.2. Bacterial α-Diversity
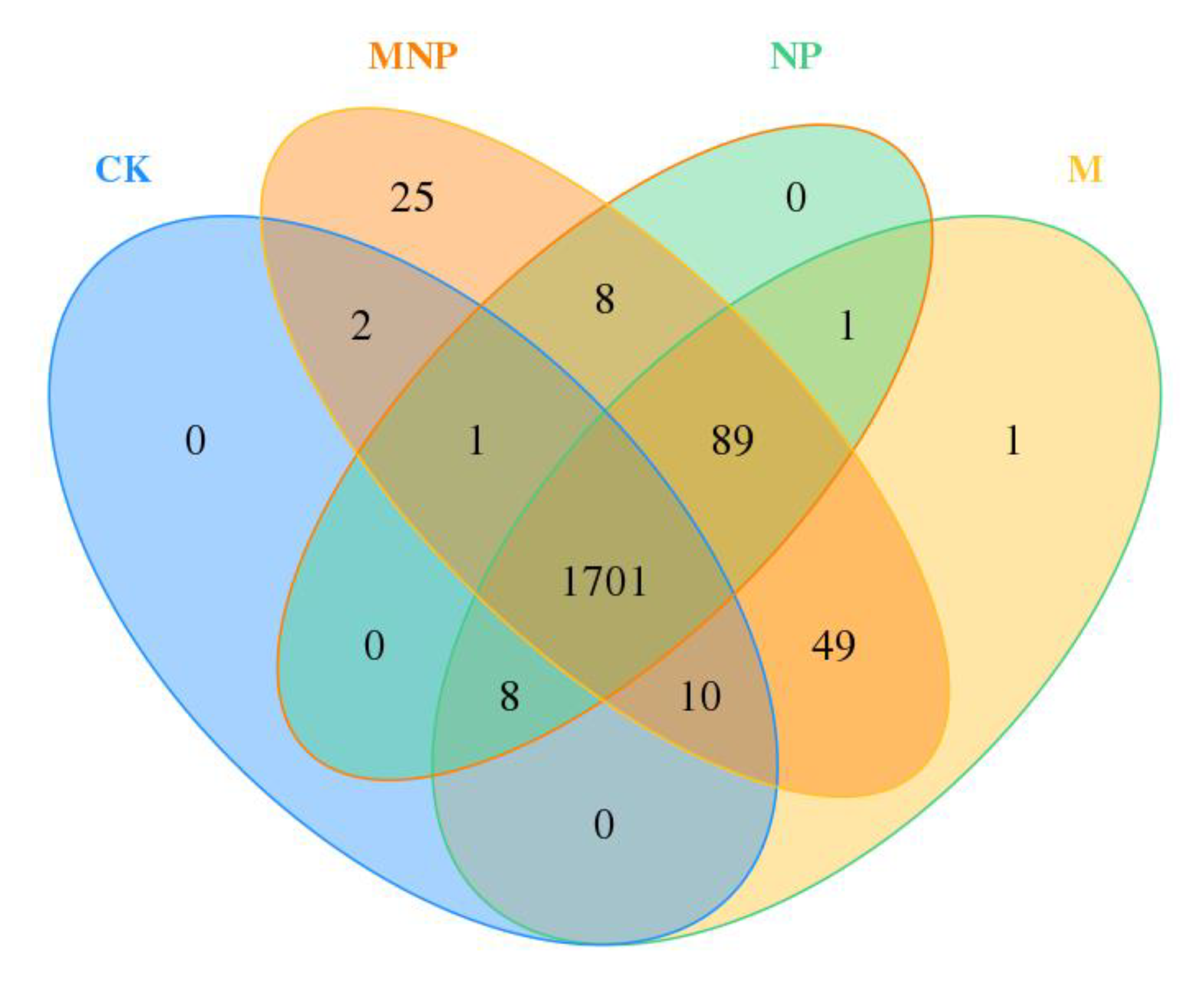
3.3. Soil Bacterial Community Composition
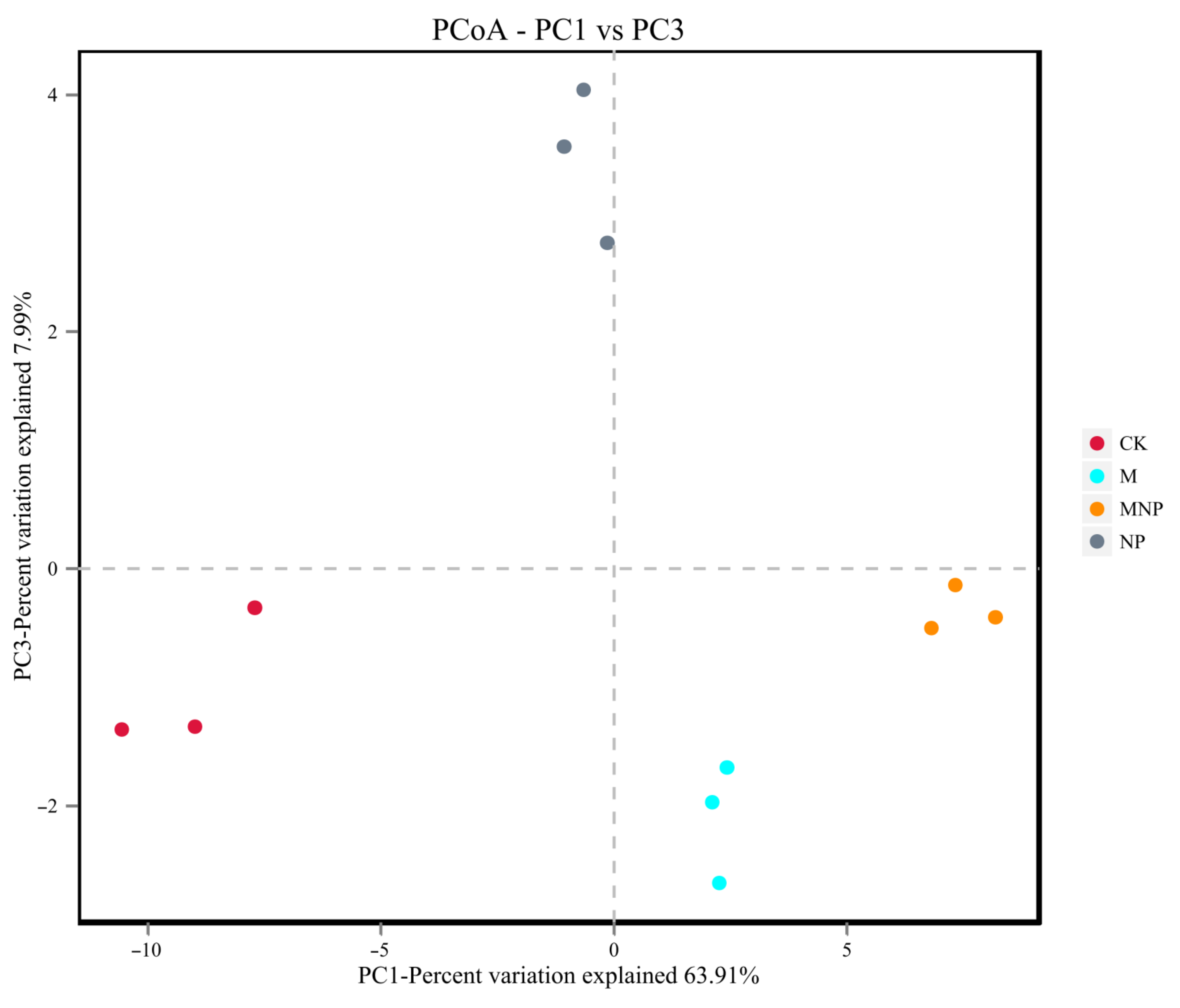
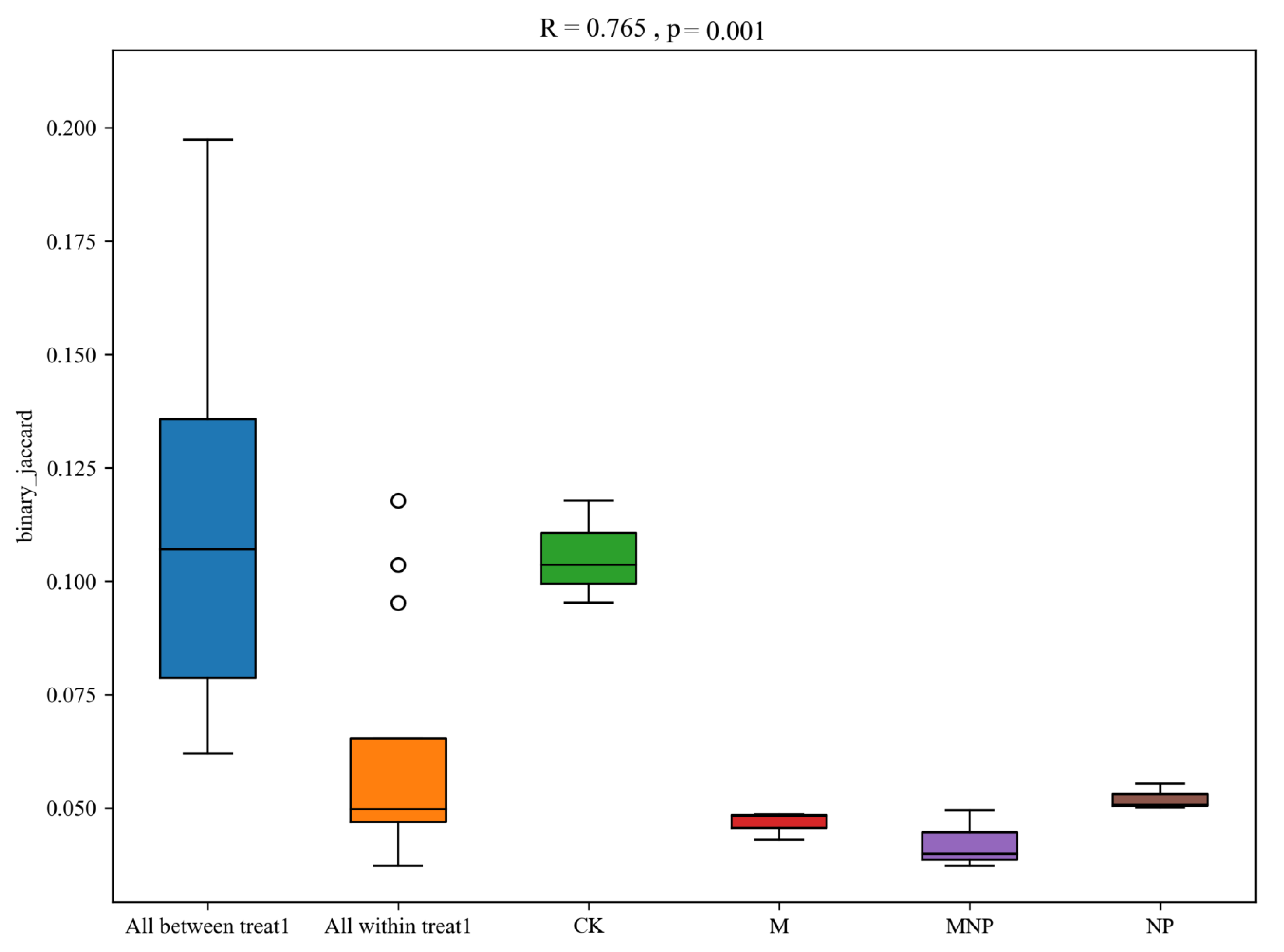
3.4. Effect of Different Treatments on Soil Bacterial Communities
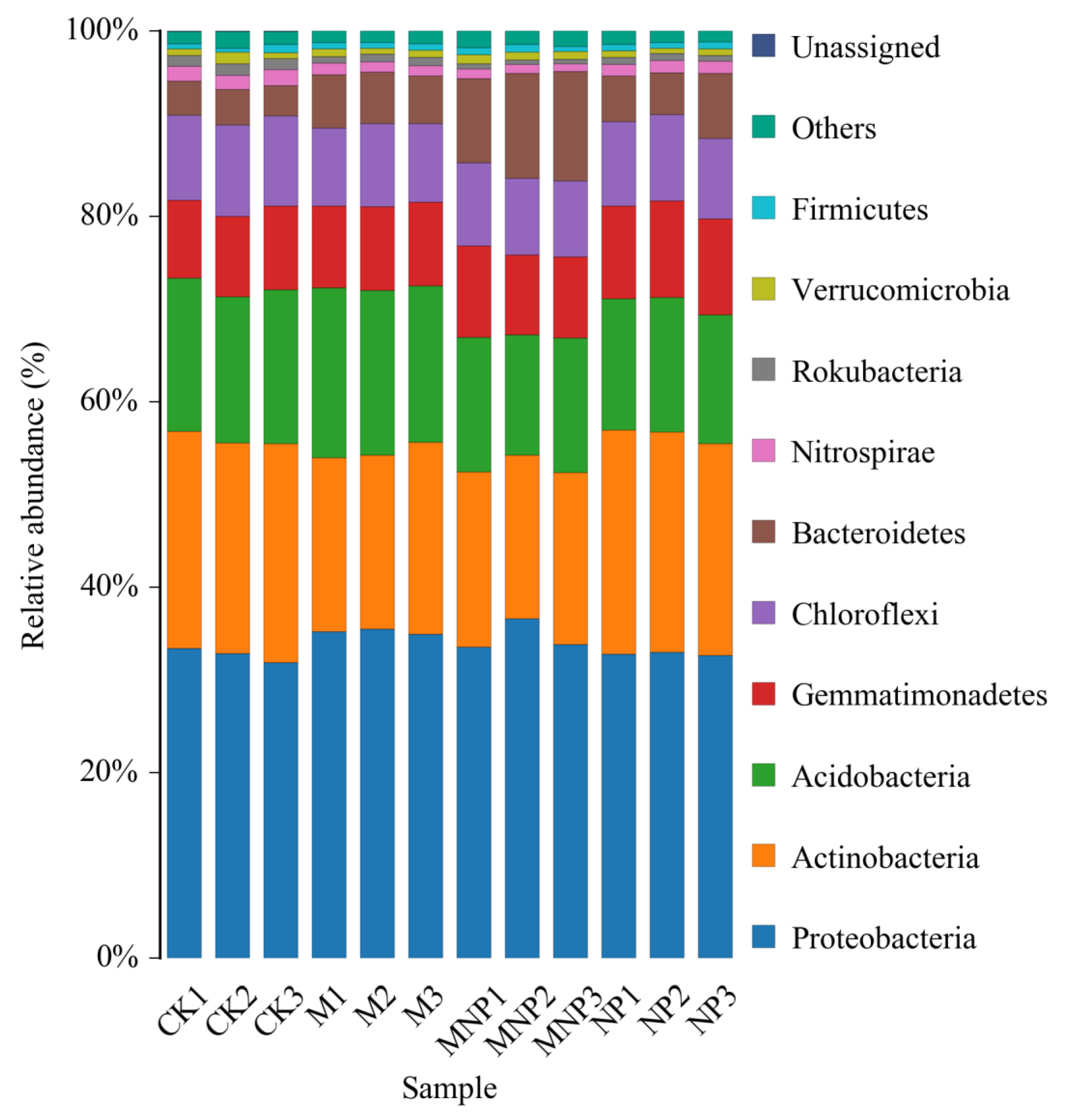
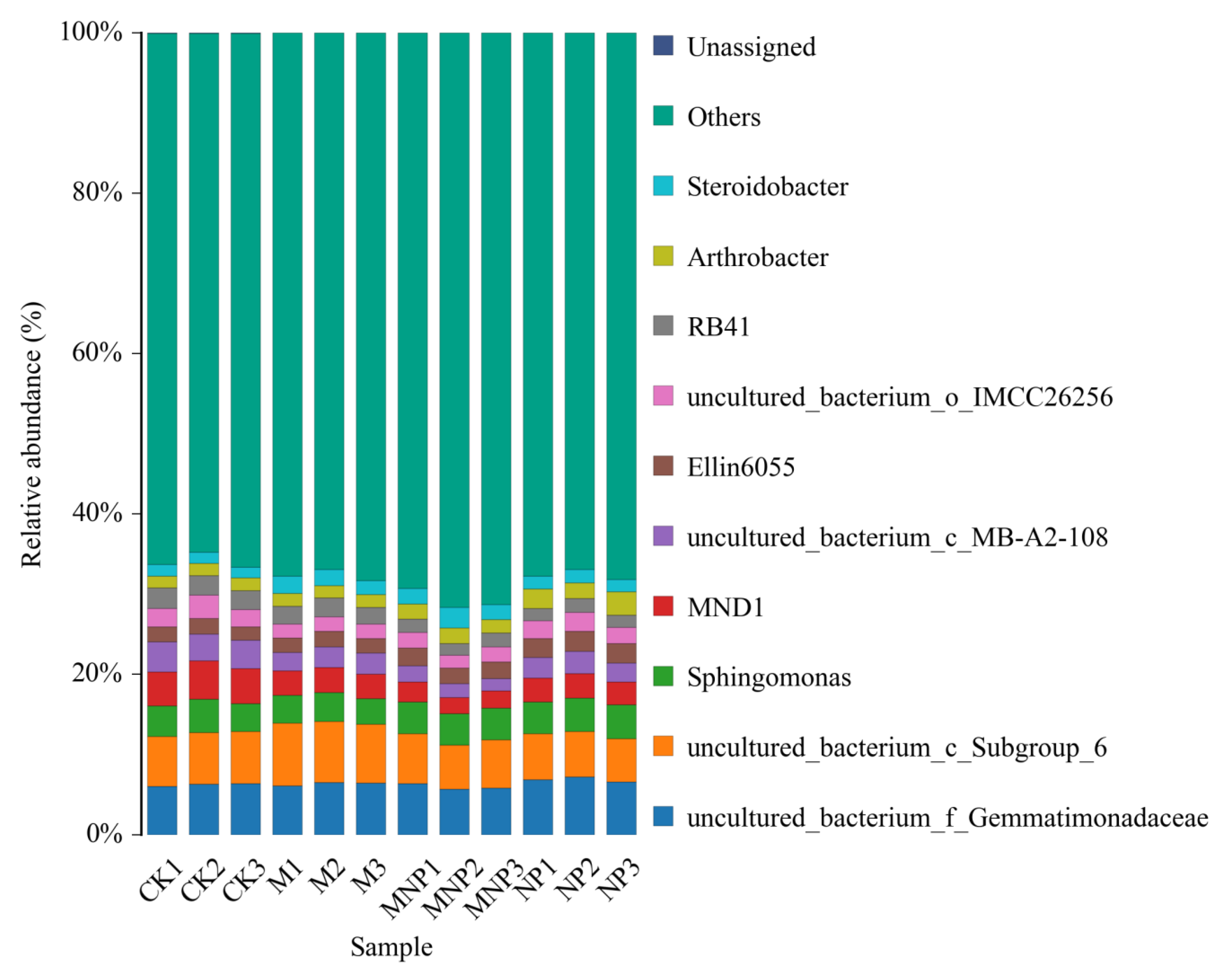
3.5. Taxonomic Composition Analysis at the Phylum and Genus Level
3.6. Effect of Different Treatments on Soil Enzyme Activities
3.7. Correlations
4. Discussion
4.1. Effects of Treatments on the Soil Physicochemical Properties
4.2. Effects of Treatments on the Bacterial Diversity and Community Composition
4.3. Effect of Different Treatments on Soil Enzyme Activities
4.4. Relationship between Soil Bacteria Community Composition and Environmental Factors and Soil Enzyme Activities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fert. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Chen, H.; Liang, Q.; Gong, Y.; Kuzyakov, Y.; Fan, M.; Plante, A.F. Reduced tillage and increased residue retention increase enzyme activity and carbon and nitrogen concentrations in soil particle size fractions in a long-term field experiment on Loess Plateau in China. Soil Tillage Res. 2019, 194, 104–296. [Google Scholar] [CrossRef]
- Postma-Blaauw, M.B.; Goede, R.G.M.D.; Bloem, J.; Faber, J.H.; Brussaard, L. Soil biota community structure and abundance under agricultural intensification and extensification. Ecology 2010, 91, 460–473. [Google Scholar] [CrossRef]
- Ollivier, J.; Töwe, S.; Bannert, A.; Hai, B.; Kastl, E.-M.; Meyer, A.; Su, M.X.; Kleineidam, K.; Schloter, M. Nitrogen turnover in soil and global change. FEMS Microbiol. Ecol. 2011, 78, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Burns, R.; Deforest, J.; Marxsen, J.; Sinsabaugh, R.; Stromberger, M.; Wallenstein, M.; Weintraub, M.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Zhao, S.; Li, K.; Zhou, W.; Qiu, S.; Huang, S.; He, P. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China. Agr. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2014, 9, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Luo, P.; Han, X.; Wang, Y.; Han, M.; Shi, H.; Liu, N.; Bai, H. Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China. Annu. Microbiol. 2015, 65, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, P.; Zeng, Z. Dynamics of Bacterial Communities in a 30-Year Fertilized Paddy Field under Different Organic–Inorganic Fertilization Strategies. Agronomy 2019, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.W.; Stadler, P.; Wu, Q.L.; Pöckl, M. The filtration-acclimatization method for isolation of an important fraction of the not readily cultivable bacteria. J. Microbiol. Meth. 2004, 57, 379–390. [Google Scholar] [CrossRef]
- Guan, L.; Cho, K.H.; Lee, J.H. Analysis of the cultivable bacterial community in jeotgal, a Korean salted andfermented seafood, and identification of its dominant bacteria. Food Microbiol. 2011, 28, 101–113. [Google Scholar] [CrossRef]
- Tebbe, C.C.; Dohrmann, A.B.; Hemkemeyer, M.; Nther, A. Microbial Community Profiling: SSCP and T-RFLP Techniques. Hydrocarbon and Lipid Microbiology Protocols, Springer Protocols Handbooks; Springer Nature: Basingstoke, UK, 2015. [Google Scholar]
- MacLean, D.; Jones, J.D.; Studholme, D.J. Application of next-generation sequencing technologies to microbial genetics. Nat. Rev. Microbiol. 2009, 7, 287–296. [Google Scholar] [CrossRef] [PubMed]
- ISS-CAS, Institute of Soil Science, Chinese Academy of Science Scientific Data Base. China Soil Data Base. Available online: http://www.soil.csdb.cn/ (accessed on 10 March 2021).
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006. A Framework for International Classification, Correlation and Communication, 2nd ed.; World Soil Resources Reports No. 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Bao, S.D. Analysis Method of Soil and Agricultural Chemistry, 3rd ed.; China Agricultural Press: Beijing, China, 2000; pp. 25–108. [Google Scholar]
- Guan, S.Y. Soil Enzyme and its Study Method; China AgriculturePress: Beijing, China, 1986; pp. 274–280. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Gloeckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nuclceic Acids Res. 2012, 41, 590–596. [Google Scholar] [CrossRef]
- Varsos, C.; Patkos, T.; Oulas, A.; Pavloudi, C.; Gougousis, A.; Zeeshan Ijaz, U.; Filiopoulou, I.; Pattakos, N.; Vanden Berghe, E.; Fernández-Guerra, A.; et al. Optimized r functions for analysis of ecological community data using the r virtual laboratory (rvlab). Biodivers. Data J. 2016, 4, 8357–8383. [Google Scholar] [CrossRef] [Green Version]
- Jari Oksanen, F.; Guillaume, B.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “vegan”: Community ecology package. Time Int. 2012, 1997, 15–17. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Huson, D.H.; Mitra, S.; Ruscheweyh, H.J.; Weber, N.; Schuster, S.C. Integrative analysis of environmental sequences using MEGAN4. Genome Res. 2011, 21, 1552–1560. [Google Scholar] [CrossRef] [Green Version]
- Zaura, E.; Keijser, B.J.F.; Huse, S.M.; Crielaard, W. Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 2009, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Luo, X.; Chen, Y.; Ye, X.; Wang, H.; Cao, Z.; Ran, W.; Cui, Z. Succession of Composition and Function of Soil Bacterial Communities During Key Rice Growth Stages. Front. Microbiol. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, D.; Niu, S. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10, 1714–1721. [Google Scholar] [CrossRef]
- Schroder, J.L.; Zhang, H.; Girma, K.; Raun, W.R.; Penn, C.J.; Payton, M.E. Soil acidification from long-term use of nitrogen fertilizers on winter wheat. Soil Sci. Soc. Am. J. 2011, 75, 957–964. [Google Scholar] [CrossRef]
- Bolan, N.; Hedley, M.; White, R. Processes of soil acidification during nitrogen cycling with emphasis on legume based pastures. Plant Soil 1991, 134, 53–63. [Google Scholar] [CrossRef]
- Powlson, D.S.; Brookes, P.C.; Christensen, B.T. Measurement of soil microbial biomass provides an early indication of changes in total soil organic matter due to straw incorporation. Soil Biol. Biochem. 1987, 19, 159–164. [Google Scholar] [CrossRef]
- Liang, R.; Hou, R.; Li, J.; Lyu, Y.; Ouyang, Z. Effects of different fertilizers on rhizosphere bacterial communities of winter wheat in the north china plain. Agronomy 2020, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil p H as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, R.; Zhang, X.X.; Guo, X.; Wang, D.; Chu, H. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spain, A.M.; Krumholz, L.R.; Elshahed, M.S. Abundance, composition, diversity and novelty of soil Proteobacteria. ISME J. 2009, 3, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Funke, G.; Pagano-Niederer, M.; Berit, S.; Falsen, E. Characteristics of Arthrobacter cumminsii, the most frequently encountered Arthrobacter species in human clinical specimens. J. Clin. Microbiol. 1998, 36, 1539–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcaraz, L.D.; Peimbert, M.; Barajas, H.R.; Dorantes-Acosta, A.E.; Bowman, J.L.; Arteaga-Vázquez, M.A. Marchantia liverworts as a proxy to plants’ basal microbiomes. Sci. Rep. 2018, 8, 12712–12723. [Google Scholar] [CrossRef]
- Yan, S.; Song, J.; Fan, J.; Yan, C.; Dong, S.; Ma, C.; Gong, Z. Changes in soil organic carbon fractions and microbial community under rice straw return in Northeast China. Glob. Ecol. Conserv. 2020, 22, 962–973. [Google Scholar] [CrossRef]
- Moreno-Espíndola, I.P.; Ferrara-Guerrero, M.J.; Luna-Guido, M.L.; Ramírez-Villanueva, D.A.; de León-Lorenzana, A.S.; Gómez-Acata, S.; González-Terreros, E.; Ramírez-Barajas, B.; Navarro-Noya, Y.E.; Sánchez-Rodríguez, L.M.; et al. The Bacterial community structure and microbial activity in a traditional organic milpa farming system under different soil moisture conditions. Front. Microbiol. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Amadou, A.; Alin Song, A.; Tang, Z.; Li, Y.; Wang, E.; Lu, Y.; Liu, X.; Yi, K.; Zhang, B.; Fan, F. The effects of organic and mineral fertilization on soil enzyme activities and bacterial community in the below- and above-ground parts of wheat. Agronomy 2020, 10, 1452. [Google Scholar] [CrossRef]
- Xu, L.; Yi, M.; Yi, H.; Guo, E.; Zhang, A. Manure and mineral fertilization change enzyme activity and bacterial community in millet rhizosphere soils. World J. Microb. Biot. 2018, 34, 1–13. [Google Scholar] [CrossRef]
- Ros, M.; Pascual, J.A.; Garcia, C.; Hernandez, M.T.; Insam, H. Hydrolase activities, microbial biomass and bacterial community in a soil after long-term amendment with different composts. Soil Biol. Biochem. 2006, 38, 3443–3452. [Google Scholar] [CrossRef]
- Dorado, J.; Zancada, M.C.; Almendros, G.; López-Fando, C. Changes in soil properties and humic substances after long-term amendments with manure and crop residues in dryland farming system. J. Plant Nutr. Soil Sci. 2003, 166, 31–38. [Google Scholar] [CrossRef]
- Bastida, F.; Kandeler, E.; Hernández, T.; García, C. Long-term effect of municipal solid waste amendment on microbial abundance and humus-associated enzyme activities under semiarid conditions. Microb. Ecol. 2008, 55, 651–661. [Google Scholar] [CrossRef]
- Ginting, D.; Kessavalou, A.; Eghball, B.; Doran, J.W. Greenhouse gas emissions and soil indicators four years after manure and compost applications. J. Environ. Qual. 2003, 32, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility. A review. Agron. Sustain. Dev. 2010, 30, 401–422. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.A.; Atkinson, D.; Gosling, P.; Jackson, L.R.; Rayns, F.W. Managing soil fertility in organic farming systems. Soil Use Manag. 2002, 18, 239–247. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Chemical Fertilizer (kg/hm2) | Manure (kg/hm2) | Total Nutrient (kg/hm2) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| N | P2O5 | K2O | N | P2O5 | K2O | N | P2O5 | K2O | |
| Manure (M) | 0 | 0 | 0 | 80 | 55 | 180 | 80 | 55 | 180 |
| Chemical fertilizer (NP) | 120 | 75 | 0 | 0 | 0 | 0 | 120 | 75 | 0 |
| Manure + Chemical fertilizer (MNP) | 120 | 75 | 0 | 80 | 55 | 180 | 200 | 130 | 180 |
| No fertilizer (CK) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Treatments | pH | AN (mg/kg) | AP (mg/kg) | AK (mg/kg) | OM (g/kg) | TN (g/kg) | TP (g/kg) | TK (g/kg) |
|---|---|---|---|---|---|---|---|---|
| M | 8.40 ± 0.23 a | 63.09 ± 3.06 ab | 22.53 ± 1.73 a | 244.78 ± 16.67 a | 11.76 ± 2.26 a | 0.81 ± 0.10 a | 0.80 ± 0.03 b | 12.79 ± 0.65 a |
| NP | 8.51 ± 0.13a | 59.15 ± 7.35 b | 14.40 ± 5.52 b | 71.27 ± 4.25 c | 7.82 ± 0.57 b | 0.74 ± 0.14 ab | 0.84 ± 0.04 ab | 12.58 ± 0.34 a |
| MNP | 8.40 ± 0.18 a | 74.55 ± 4.55 a | 26.90 ± 2.80 a | 223.90 ± 3.04 b | 12.27 ± 1.14 a | 0.85 ± 0.12 a | 0.85 ± 0.12 a | 12.36 ± 0.74 a |
| CK | 8.59 ± 0.10 a | 37.98 ± 1.23 c | 5.10 ± 0.70 c | 60.01 ± 8.30 c | 6.52 ± 1.48 b | 0.56 ± 0.06 b | 0.66 ± 0.03 c | 12.78 ± 0.17 a |
| Treatments | ACE | Shannon |
|---|---|---|
| M | 1828.12 ± 6.06 b | 6.57 ± 0.01 a |
| NP | 1769.66 ± 7.89 c | 6.53 ± 0.02 ab |
| MNP | 1867.82 ± 8.81 a | 6.55 ± 0.05 ab |
| CK | 1665.51 ± 5.54 d | 6.48 ± 0.02 b |
| Treatments | SUC Activity (mg·Glucose·g−1·24 h−1) | URE Activity (mg·NH3-N·g−1·24 h−1) | ALP Activity (mg Phenol·g−1·24 h−1) |
|---|---|---|---|
| M | 19.55 ± 3.227 ab | 1.983 ± 0.021 b | 1.820 ± 0.080 b |
| NP | 15.78 ± 1.520 b | 1.900 ± 0.290 b | 1.404 ± 0.153 bc |
| MNP | 21.42 ± 0.708 a | 2.537 ± 0.218 a | 2.264 ± 0.278 a |
| CK | 8.723 ± 0.083 c | 1.210 ± 0.128 c | 1.348 ± 0.016 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Xie, W.; Yang, Z.; Huang, X.; Zhou, H. Effects of Manure and Chemical Fertilizer on Bacterial Community Structure and Soil Enzyme Activities in North China. Agronomy 2021, 11, 1017. https://doi.org/10.3390/agronomy11051017
Liu Z, Xie W, Yang Z, Huang X, Zhou H. Effects of Manure and Chemical Fertilizer on Bacterial Community Structure and Soil Enzyme Activities in North China. Agronomy. 2021; 11(5):1017. https://doi.org/10.3390/agronomy11051017
Chicago/Turabian StyleLiu, Zhiping, Wenyan Xie, Zhenxing Yang, Xuefang Huang, and Huaiping Zhou. 2021. "Effects of Manure and Chemical Fertilizer on Bacterial Community Structure and Soil Enzyme Activities in North China" Agronomy 11, no. 5: 1017. https://doi.org/10.3390/agronomy11051017
APA StyleLiu, Z., Xie, W., Yang, Z., Huang, X., & Zhou, H. (2021). Effects of Manure and Chemical Fertilizer on Bacterial Community Structure and Soil Enzyme Activities in North China. Agronomy, 11(5), 1017. https://doi.org/10.3390/agronomy11051017