
Microbiological Activity during Co-Composting of Food and Agricultural Waste for Soil Amendment


Abstract
:1. Introduction
2. Materials and Methods
2.1. Compost Mixture
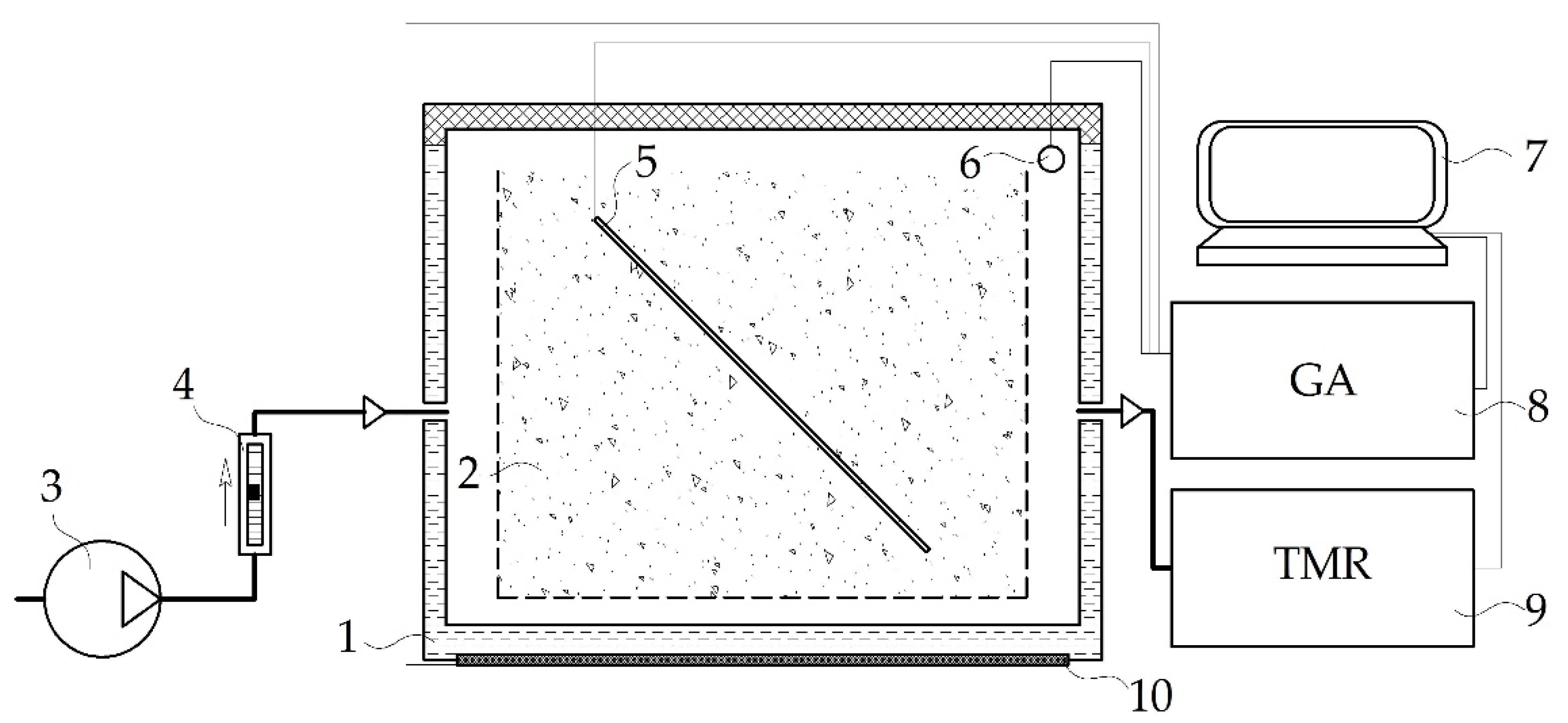
2.2. Experimental Setup
2.3. Physicochemical Studies
2.4. Microbiological Studies
2.5. Profiling of Prokaryotic and Fungal Communities Based on 16S rRNA Gene and Internal Transcribed Spacer (ITS)
2.6. Statistical Analysis
3. Results and Discussion
3.1. Dynamics of the Physicochemical Parameters of Composting Mixed Food and Agricultural Waste
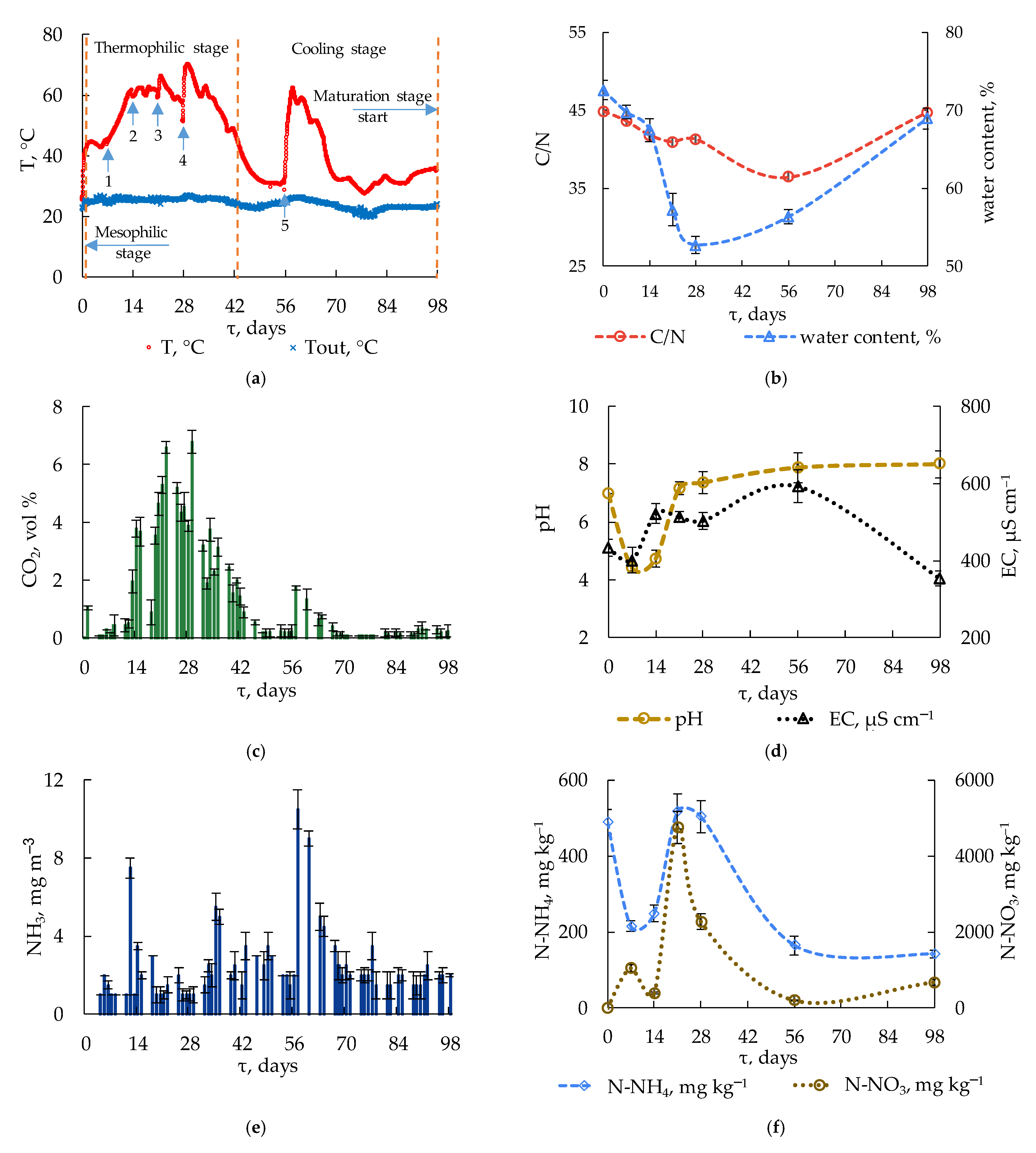
3.1.1. Temperature and CO2 Production
3.1.2. pH and Electrical Conductivity
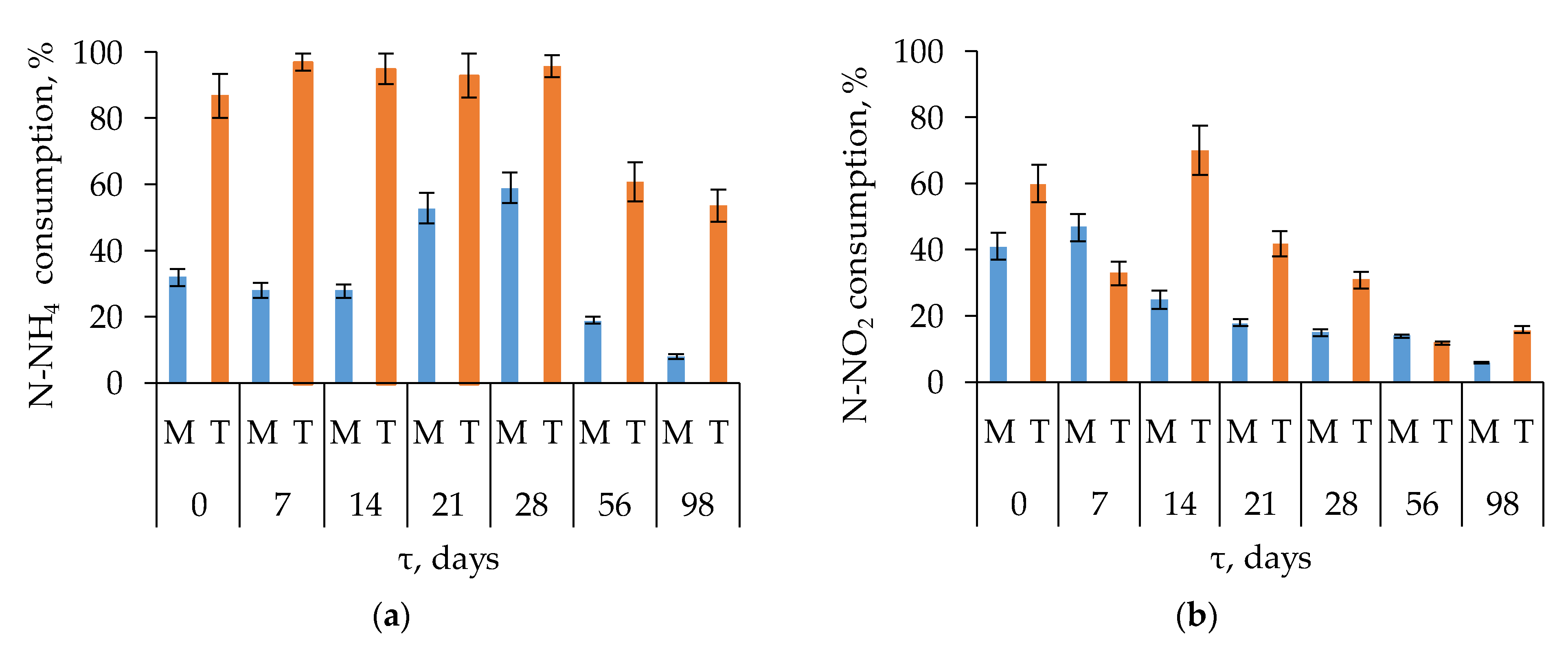
3.1.3. Dynamics of Nitrogen Compounds
3.1.4. C/N Ratio and Germination Index
3.1.5. Mature Compost
3.2. Total Microbial Number at Different Stages of Composting
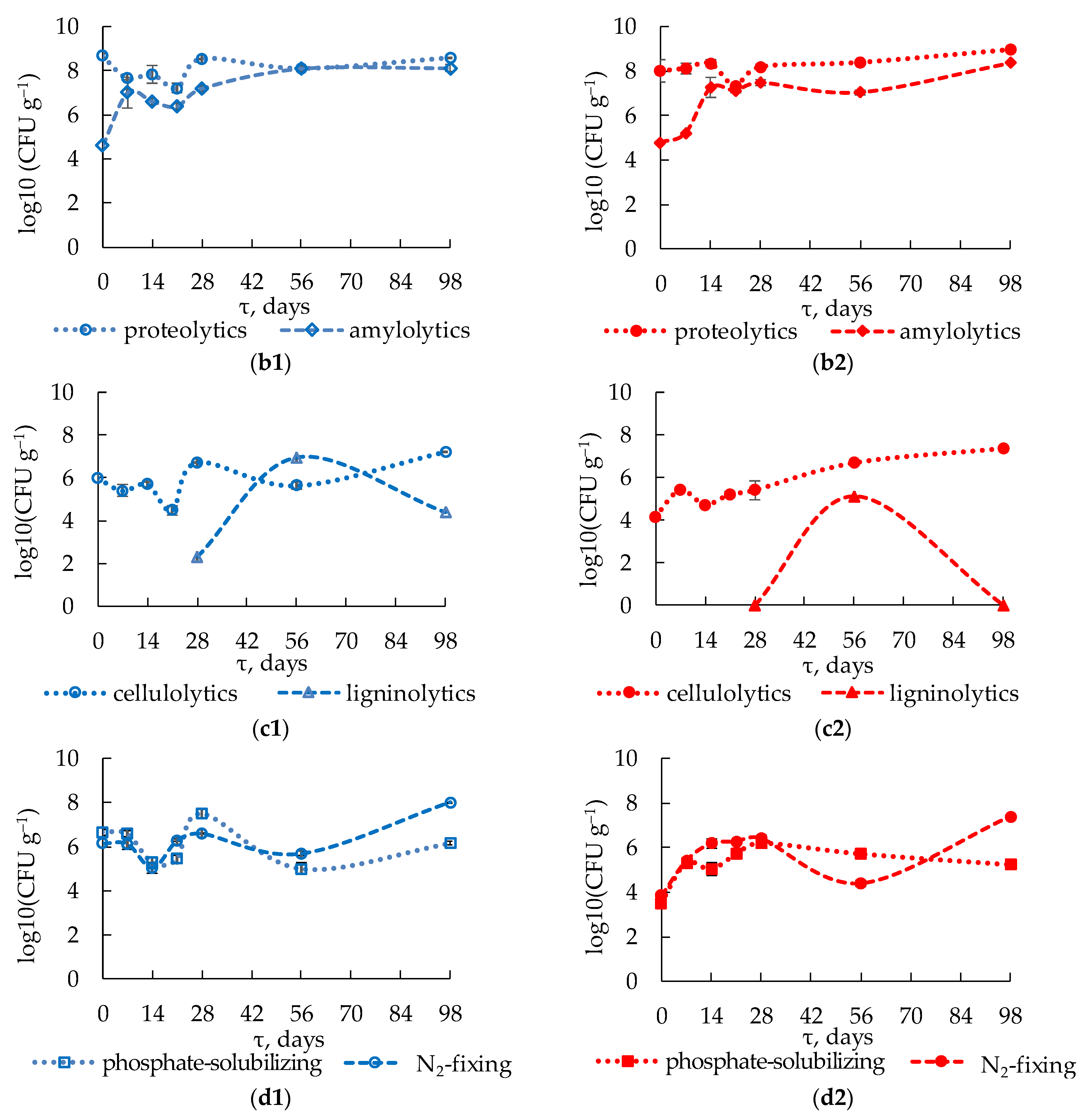
3.3. Abundance of Various Physiological Groups of Cultured Microorganisms during Composting
3.4. Composition and Biodiversity of the Microbial Community
3.4.1. Fungal Community
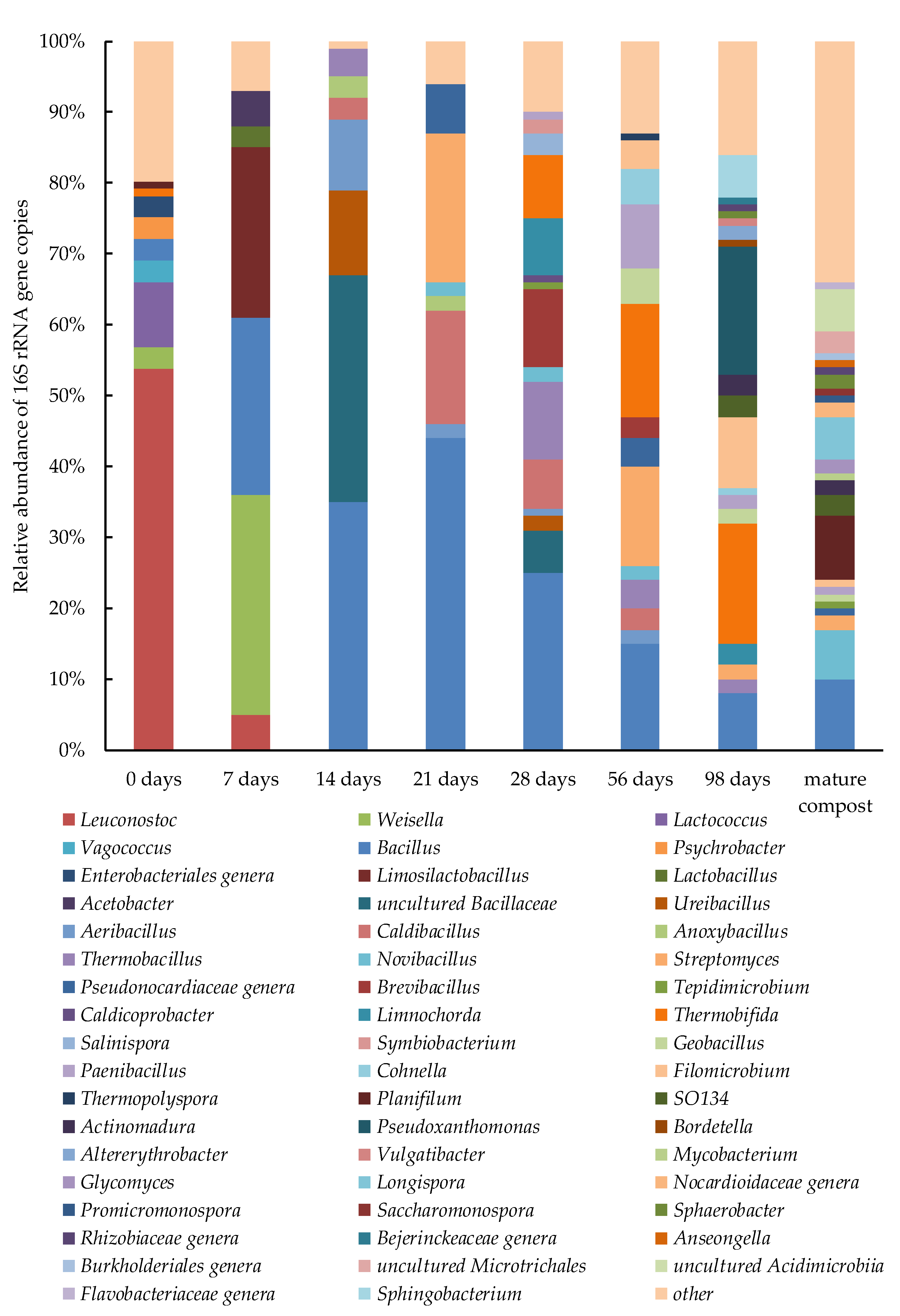
3.4.2. Prokaryotic Community
3.4.3. Association of Microbial Diversity and Abundance with Environmental Variables
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wang, S.; Zeng, Y. Ammonia emission mitigation in food waste composting: A review. Bioresour. Technol. 2018, 248, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.K.; Sarsaiya, S.; Awasthi, M.K.; Liu, T.; Zhao, J.; Kumar, S.; Zhang, Z. Changes in global trends in food waste composting: Research challenges and opportunities. Bioresour. Technol. 2020, 299, 122555. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, O.J.; Ospina, D.A.; Montoya, S. Compost supplementation with nutrients and microorganisms in composting process. Waste Manag. 2017, 69, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Chandna, P.; Nain, L.; Singh, S.; Kuhad, R.C. Assessment of bacterial diversity during composting of agricultural byproducts. BMC Microbiol. 2013, 13, 99. [Google Scholar] [CrossRef] [Green Version]
- Adani, F.; Genevini, P.; Gasperi, F.; Zorzi, G. Organic Matter Evolution Index (OMEI) as a Measure Of Composting Efficiency. Compos. Sci. Util. 1997, 5, 53–62. [Google Scholar] [CrossRef]
- Tiquia, S.M.; Richard, T.L.; Honeyman, M.S. Effect of Windrow Turning and Seasonal Temperatures on Composting of Hog Manure from Hoop Structures. Environ. Technol. 2000, 21, 1037–1046. [Google Scholar] [CrossRef]
- Fracchia, L.; Dohrmann, A.B.; Martinotti, M.G.; Tebbe, C.C. Bacterial diversity in a finished compost and vermicompost: Differences revealed by cultivation-independent analyses of PCR-amplified 16S rRNA genes. Appl. Microbiol. Biotechnol. 2006, 71, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Brinton, W.F. Compost Quality Standards and Guidelines; Final Report by Woods End Research Laboratories; Technical Report for the New York State Association of Recyclers; Woods End Research Laboratory, Inc.: New York, NY, USA, 2000. [Google Scholar]
- Negi, S.; Mandpe, A.; Hussain, A.; Kumar, S. Collegial effect of maggots larvae and garbage enzyme in rapid composting of food waste with wheat straw or biomass waste. J. Clean. Prod. 2020, 258, 120854. [Google Scholar] [CrossRef]
- Castaldi, P.; Garau, G.; Deiana, P.; Melis, P. Evolution of carbon compounds during municipal solid waste composting: Suitability of chemical and biochemical parameters in defining the stability and maturity of the end product. Dyn. Soil Dyn. Plant. 2009, 3, 17–31. [Google Scholar]
- Zhang, L.; Jia, Y.; Zhang, X.; Feng, X.; Wu, J.; Wang, L.; Chen, G. Wheat straw: An inefficient substrate for rapid natural lignocellulosic composting. Bioresour. Technol. 2016, 209, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Maulini-Duran, C.; Artola, A.; Font, X.; Sánchez, A. Gaseous emissions in municipal wastes composting: Effect of the bulking agent. Bioresour. Technol. 2014, 172, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Li, G.; Shi, H.; Wang, Y. Effects of phosphogypsum and superphosphate on compost maturity and gaseous emissions during kitchen waste composting. Waste Manag. 2015, 36, 70–76. [Google Scholar] [CrossRef]
- Palaniveloo, K.; Amran, M.A.; Norhashim, N.A.; Mohamad-Fauzi, N.; Peng-Hui, F.; Hui-Wen, L.; Kai-Lin, Y.; Jiale, L.; Chian-Yee, M.G.; Jing-Yi, L.; et al. Food Waste Composting and Microbial Community Structure Profiling. Processes 2020, 8, 723. [Google Scholar] [CrossRef]
- Sun, L.; Han, X.; Li, J.; Zhao, Z.; Liu, Y.; Xi, Q.; Guo, X.; Gun, S. Microbial Community and Its Association With Physicochemical Factors During Compost Bedding for Dairy Cows. Front. Microbiol. 2020, 11, 254. [Google Scholar] [CrossRef] [PubMed]
- Nakasaki, K.; Hirai, H.; Mimoto, H.; Quyen, T.N.M.; Koyama, M.; Takeda, K. Succession of microbial community during vigorous organic matter degradation in the primary fer-mentation stage of food waste composting. Sci. Total Environ. 2019, 671, 1237–1244. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Saveliev, A.; Grigoryeva, T.; Boulygina, E.; Selivanovskaya, S. Fungal and bacterial successions in the process of co-composting of organic wastes as re-vealed by 454 pyrosequencing. PLoS ONE 2017, 12, e0186051. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cui, H.; Shi, J.; Zhao, X.; Zhao, Y.; Wei, Z. Relationship between bacterial diversity and environmental parameters during composting of different raw materials. Bioresour. Technol. 2015, 198, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, H.; Wang, Z.; Chen, G.; Wang, L. Dynamic changes of the dominant functioning microbial community in the compost of a 90-m3 aerobic solid state fermentor revealed by integrated meta-omics. Bioresour. Technol. 2016, 203, 1–10. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Zou, K.-H.; Yang, J.-B.; Li, G.-X.; Yang, Q.-Y.; Zhang, F. Analysis of odor pollutants in kitchen waste composting. Environ. Sci. 2012, 33, 2563–2568. [Google Scholar]
- Arrigoni, J.P.; Paladino, G.; Garibaldi, L.A.; Laos, F. Inside the small-scale composting of kitchen and garden wastes: Thermal performance and stratification effect in vertical compost bins. Waste Manag. 2018, 76, 284–293. [Google Scholar] [CrossRef]
- Mironov, V.V.; Bochkova, E.A.; Gannesen, A.V.; Vanteeva, A.V.; Russkova, Y.I.; Nozhevnikova, A.N. Dynamics of Biological Processes during Composting of Anaerobically Digested Wastewater Sludge. Microbiology 2020, 89, 470–482. [Google Scholar] [CrossRef]
- Zhang, H.; Li, G.; Gu, J.; Wang, G.; Li, Y.; Zhang, D. Influence of aeration on volatile sulfur compounds (VSCs) and NH 3 emissions during aerobic composting of kitchen waste. Waste Manag. 2016, 58, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Ivankin, A.; Pandya, U.; Saraf, M. Intensification of Aerobic Processing of the Organic Wastes into Compost. In Maheshwari D. Composting for Sustainable Agriculture. Sustainable Development and Biodiversity; Springer International Publishing: Cham, Switzerland, 2014; Volume 3, pp. 23–42. [Google Scholar]
- Zorpas, A.A.; Loizidou, M. Sawdust and natural zeolite as a bulking agent for improving quality of a composting product from anaerobically stabilized sewage sludge. Bioresour. Technol. 2008, 99, 7545–7552. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Cegarra, J.; Roig, A.; Garcia, D. Relationships between organic matter and carbon contents of organic wastes. Bioresour. Technol. 1993, 44, 203–207. [Google Scholar] [CrossRef]
- Banegas, V.; Moreno, J.; García, C.; Leon, G.; Hernández, T.; Moreno, J. Composting anaerobic and aerobic sewage sludges using two proportions of sawdust. Waste Manag. 2007, 27, 1317–1327. [Google Scholar] [CrossRef]
- Cesaro, A.; Belgiorno, V.; Guida, M. Compost from organic solid waste: Quality assessment and European regulations for its sustainable use. Resour. Conserv. Recycl. 2015, 94, 72–79. [Google Scholar] [CrossRef]
- Kononova, M.; Belchikova, N. Accelerated methods for determining the composition of humus in mineral soils. Soil Sci. 1961, 10, 75–87. (In Russian) [Google Scholar]
- Brinton, W.F.; Storms, P.; Blewett, T.C. Occurrence and Levels of Fecal Indicators and Pathogenic Bacteria in Market-Ready Recycled Organic Matter Composts. J. Food Prot. 2009, 72, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Sintsov, K.N.; Starkova, E.V.; Martinson, E.A.; Litvinets, S.G. Micromycetes of Lignin Sludge Dumps. Bull. Samara Sci. Cent. Russ. Acad. Sci. 2013, 15, 1961–1964. (In Russian) [Google Scholar]
- Beffa, T. The composting biotechnology: A microbial aerobic solid substrate fermentation complex process. Compost. Process Manag. 2002, 6, 1–30. [Google Scholar]
- Gohl, D.M.; MacLean, A.; Hauge, A.; Becker, A.; Walek, D.; Beckman, K.B. An optimized protocol for high-throughput amplicon-based microbiome profiling. Protoc. Exch. 2016. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugerth, L.W.; Wefer, H.A.; Lundin, S.; Jakobsson, H.E.; Lindberg, M.; Rodin, S.; Engstrand, L.; Andersson, A.F. DegePrime, a Program for Degenerate Primer Design for Broad-Taxonomic-Range PCR in Microbial Ecology Studies. Appl. Environ. Microbiol. 2014, 80, 5116–5123. [Google Scholar] [CrossRef] [Green Version]
- Merkel, A.Y.; Tarnovetskii, I.Y.; Podosokorskaya, O.A.; Toshchakov, S.V. Analysis of 16S rRNA Primer Systems for Profiling of Thermophilic Microbial Communities. Microbiology 2019, 88, 671–680. [Google Scholar] [CrossRef]
- Turenne, C.Y.; Sanche, S.E.; Hoban, D.J.; Karlowsky, J.A.; Kabani, A.M. Rapid identification of fungi by using the ITS2 genetic region and an automated fluorescent capillary electrophoresis system. J. Clin. Microbiol. 1999, 37, 1846–1851. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 315–322. [Google Scholar]
- Kallistova, A.; Merkel, A.; Kanapatskiy, T.; Boltyanskaya, Y.; Tarnovetskii, I.; Perevalova, A.; Kevbrin, V.; Samylina, O.; Pimenov, N. Methanogenesis in the Lake Elton saline aquatic system. Extremophiles 2020, 24, 657–672. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Efimova, D.; Tyakht, A.V.; Popenko, A.S.; Vasilyev, A.; Altukhov, I.A.; Dovidchenko, N.; Odintsova, V.; Klimenko, N.; Loshkarev, R.; Pashkova, M.; et al. Knomics-Biota—A system for exploratory analysis of human gut microbiota data. BioData Min. 2018, 11, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Lee, S.M. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Pandey, A.K.; Bundela, P.S.; Khan, J. Co-composting of organic fraction of municipal solid waste mixed with different bulking waste: Characterization of physicochemical parameters and microbial enzymatic dynamic. Bioresour. Technol. 2015, 182, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, M.; Wolny-Koładka, K.; Vaverková, M.D. Effect of biochar addition on the OFMSW composting process under real conditions. Waste Manag. 2019, 84, 364–372. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, X.; Huang, Y.; Huang, H. Inoculation with nitrogen turnover bacterial agent appropriately increasing nitrogen and promoting maturity in pig manure composting. Waste Manag. 2015, 39, 78–85. [Google Scholar] [CrossRef]
- Duan, Y.; Awasthi, S.K.; Liu, T.; Zhang, Z.; Awasthi, M.K. Evaluation of integrated bio-char with bacterial consortium on gaseous emissions mitigation and nutrients sequestration during pig manure composting. Bioresour. Technol. 2019, 291, 121880. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Zhang, Y.; Xia, X.; Qi, H.; Li, M.; Pan, H.; Xi, B. Effect of inoculation with a microbial consortium that degrades organic acids on the composting efficiency of food waste. Microb. Biotechnol. 2018, 11, 1124–1136. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, N.D.Q.; Souza, T.P.; Costa, L.M.A.S.; De Castro, C.P.; Dias, E.S. Microbial additives in the composting process. Ciência E Agrotecnologia 2017, 41, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Waqas, M.; Nizami, A.; Aburiazaiza, A.; Barakat, M.; Asam, Z.; Khattak, B.; Rashid, M. Untapped potential of zeolites in optimization of food waste composting. J. Environ. Manag. 2019, 241, 99–112. [Google Scholar] [CrossRef]
- Kalamdhad, A.S.; Kazmi, A.A. Rotary drum composting of different organic waste mixtures. Waste Manag. Res. 2009, 27, 129–137. [Google Scholar] [CrossRef]
- Kucbel, M.; Raclavská, H.; Růžičková, J.; Švédová, B.; Sassmanová, V.; Drozdová, J.; Juchelková, D. Properties of composts from household food waste produced in automatic composters. J. Environ. Manag. 2019, 236, 657–666. [Google Scholar] [CrossRef]
- Pagans, E.; Barrena, R.; Font, X.; Sánchez, A. Ammonia emissions from the composting of different organic wastes. Dependency on process temperature. Chemosphere 2006, 62, 1534–1542. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Liu, R.; Hao, J.; Li, D.; Wei, Z.; Teng, R.; Sun, B. Protein and carbohydrate drive microbial responses in diverse ways during different animal manures composting. Bioresour. Technol. 2019, 271, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Cerda, A.; Artola, A.; Font, X.; Barrena, R.; Gea, T.; Sánchez, A. Composting of food wastes: Status and challenges. Bioresour. Technol. 2018, 248, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Ren, L.; Li, G.; Chen, T.; Guo, R. Influence of aeration on CH4, N2O and NH3 emissions during aerobic composting of a chicken manure and high C/N waste mixture. Waste Manag. 2011, 31, 33–38. [Google Scholar] [CrossRef]
- Guo, R.; Li, G.; Jiang, T.; Schuchardt, F.; Chen, T.; Zhao, Y.; Shen, Y. Effect of aeration rate, C/N ratio and moisture content on the stability and maturity of compost. Bioresour. Technol. 2012, 112, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Estrella-González, M.J.; Suárez-Estrella, F.; Jurado, M.M.; López, M.J.; López-González, J.A.; Siles-Castellano, A.B.; Muñoz-Mérida, A.; Moreno, J. Uncovering new indicators to predict stability, maturity and biodiversity of com-post on an industrial scale. Bioresour. Technol. 2020, 313, 123557. [Google Scholar] [CrossRef]
- Lasaridi, K.; Protopapa, I.; Kotsou, M.; Pilidis, G.; Manios, T.; Kyriacou, A. Quality assessment of composts in the Greek market: The need for standards and quality assurance. J. Environ. Manag. 2006, 80, 58–65. [Google Scholar] [CrossRef]
- Cesaro, A.; Conte, A.; Belgiorno, V.; Siciliano, A.; Guida, M. The evolution of compost stability and maturity during the full-scale treatment of the organic fraction of municipal solid waste. J. Environ. Manag. 2019, 232, 264–270. [Google Scholar] [CrossRef]
- Pepe, O.; Ventorino, V.; Blaiotta, G. Dynamic of functional microbial groups during mesophilic composting of agro-industrial wastes and free-living (N2)-fixing bacteria application. Waste Manag. 2013, 33, 1616–1625. [Google Scholar] [CrossRef]
- Simujide, H.; Aorigele, C.; Wang, C.-J.; Lina, M.; Manda, B. Microbial activities during mesophilic composting of manure and effect of calcium cyanamide addition. Int. Biodeterior. Biodegrad. 2013, 83, 139–144. [Google Scholar] [CrossRef]
- Ryckeboer, J.; Mergaert, J.; Coosemans, J.; Deprins, K.; Swings, J. Microbiological aspects of biowaste during composting in a monitored compost bin. J. Appl. Microbiol. 2003, 94, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Chroni, C.; Kyriacou, A.; Georgaki, I.; Manios, T.; Kotsou, M.; Lasaridi, K. Microbial characterization during composting of biowaste. Waste Manag. 2009, 29, 1520–1525. [Google Scholar] [CrossRef]
- Vorobiev, N.I.; Sviridova, O.V.; Popov, A.A.; Rusakova, I.V.; Petrov, V.B. Graph-analysis of gene-metabolic networks of soil microorganisms that transform plant residues into humic substances. Agricult. Biol. 2011, 3, 88–93. [Google Scholar]
- Vargas-García, M.; Suárez-Estrella, F.; López, M.; Moreno, J. Microbial population dynamics and enzyme activities in composting processes with different starting materials. Waste Manag. 2010, 30, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wei, Z.; Cao, Z.; Zhao, Y.; Zhao, X.; Lu, Q.; Wang, X.; Zhang, X. A regulating method for the distribution of phosphorus fractions based on environmental parameters related to the key phosphate-solubilizing bacteria during composting. Bioresour. Technol. 2016, 211, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Zhao, Y.; Fan, Y.; Lu, Q.; Li, M.; Wei, Q.; Zhao, Y.; Cao, Z.; Wei, Z. Impact of phosphate-solubilizing bacteria inoculation methods on phosphorus transformation and long-term utilization in composting. Bioresour. Technol. 2017, 241, 134–141. [Google Scholar] [CrossRef]
- Chang, C.-H.; Yang, S.-S. Thermo-tolerant phosphate-solubilizing microbes for multi-functional biofertilizer preparation. Bioresour. Technol. 2009, 100, 1648–1658. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, R.; Flotats, X.; Marfà, O. Changes in the chemical and physicochemical properties of the solid fraction of cattle slurry during composting using different aeration strategies. Waste Manag. 2006, 26, 1081–1091. [Google Scholar] [CrossRef]
- Cáceres, R.; Malińska, K.; Marfà, O. Nitrification within composting: A review. Waste Manag. 2018, 72, 119–137. [Google Scholar] [CrossRef]
- López-Cano, I.; Roig, A.; Cayuela, M.L.; Alburquerque, J.A.; Sánchez-Monedero, M.A. Biochar improves N cycling during composting of olive mill wastes and sheep manure. Waste Manag. 2016, 49, 553–559. [Google Scholar] [CrossRef]
- Houbraken, J.; Samson, R.A.; Frisvad, J.C. Byssochlamys: Significance of heat resistance and mycotoxin production. In Advances in Food Mycology. Advances in Experimental Medicine and Biology; Hocking, A.D., Pitt, J.I., Samson, R.A., Thrane, U., Eds.; Springer: Boston, MA, USA, 2006; Volume 571. [Google Scholar] [CrossRef]
- Samson, R.; Houbraken, J.; Varga, J.; Frisvad, J. Polyphasic taxonomy of the heat resistant ascomycete genus Byssochlamys and its Paecilomyces anamorphs. Persoonia 2009, 22, 14–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, D.; Lesch, S.; Buzina, W.; Galler, H.; Gutschi, A.M.; Habib, J.; Pfeifer, B.; Luxner, J.; Reinthaler, F.F. Culturable fungi in potting soils and compost. Med. Mycol. 2016, 54, 825–834. [Google Scholar] [CrossRef]
- Tiquia-Arashiro, S.M. Thermophilic Fungi in Composts: Their Role in Composting and Industrial Processes. In Fungi in Extreme Environments: Ecological Role and Biotechnological Significance; Springer: Cham, Switzerland, 2019; pp. 587–605. [Google Scholar]
- Zhang, Y.; Wu, W.; Cai, L. Polyphasic characterisation of Chaetomium species from soil and compost revealed high number of undescribed species. Fungal Biol. 2017, 121, 21–43. [Google Scholar] [CrossRef]
- Insam, H.; De Bertoldi, M. Chapter 3 Microbiology of the composting process. Solid Waste Assess. Monit. Remediat. 2007, 8, 25–48. [Google Scholar] [CrossRef]
- Coorevits, A.; Dinsdale, A.E.; Halket, G.; Lebbe, L.; De Vos, P.; Van Landschoot, A.; Logan, N.A. Taxonomic revision of the genus Geobacillus: Emendation of Geobacillus, G. stearothermophilus, G. jurassicus, G. toebii, G. thermodenitrificans and G. thermoglucosidans (nom. corrig., formerly ‘thermoglucosidasius’); transfer of Bacillus thermantarcticus to the genus as G. thermantarcticus comb. nov.; proposal of Caldibacillus debilis gen. nov., comb. nov.; transfer of G. tepidamans to Anoxybacillus as A. tepidamans comb. nov.; and proposal of Anoxybacillus caldiproteolyticus sp. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 1470–1485. [Google Scholar] [CrossRef] [PubMed]
- Ghazifard, A.; Kasra-Kermanshahi, R.; Far, Z.E. Identification of thermophilic and mesophilic bacteria and fungi in Esfahan (Iran) municipal solid waste compost. Waste Manag. Res. 2001, 19, 257–261. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; Xia, J.; Chen, Y. Effect of microbial inoculation on physicochemical properties and bacterial community structure of citrus peel composting. Bioresour. Technol. 2019, 291, 121843. [Google Scholar] [CrossRef]
- Buzón-Durán, L.; Pérez-Lebeña, E.; Martín-Gil, J.; Sánchez-Báscones, M.; Martín-Ramos, P. Applications of Streptomyces spp. Enhanced Compost in Sustainable Agriculture. In Biology of Composts. Soil Biology; Meghvansi, M., Varma, A., Eds.; Springer: Cham, Switzerland, 2020; Volume 58. [Google Scholar] [CrossRef]
- Devi, P.; Kandasamy, S.; Uthandi, S.; Chendrayan, K. Laccase Producing Streptomyces Bikiniensis CSC12 Isolated From Compost. J. Microbiol. Biotechnol. Food Sci. 2016, 6, 794–798. [Google Scholar] [CrossRef]
- Watanabe, M.; Kojima, H.; Fukui, M. Limnochorda pilosa gen. nov., sp. nov., a moderately thermophilic, facultatively anaerobic, pleomorphic bacterium and proposal of Limnochordaceae fam. nov., Limnochordales ord. nov. and Limnochordia classis nov. in the phylum Firmicutes. Int. J. Syst. Evol. Microbiol. 2015, 65, 2378–2384. [Google Scholar] [CrossRef]
- Grady, E.N.; Macdonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Fact. 2016, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Gao, J.; Liang, D.; Zhu, Y.; Li, B.; Jin, H. Thermal pretreatment enhances the degradation and humification of lignocellulose by stimulating thermophilic bacteria during dairy manure composting. Bioresour. Technol. 2021, 319, 124149. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Revathi, K.; Khanna, S. Biodegradation of cellulosic and lignocellulosic waste by Pseudoxanthomonas sp R-28. Carbohydr. Polym. 2015, 134, 761–766. [Google Scholar] [CrossRef]
- Tortosa, G.; Castellano-Hinojosa, A.; Correa-Galeote, D.; Bedmar, E.J. Evolution of bacterial diversity during two-phase olive mill waste (“alperujo”) composting by 16S rRNA gene pyrosequencing. Bioresour. Technol. 2017, 224, 101–111. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, L.; Wang, B.; He, Q.; Wan, D. Using the modified pine wood as a novel recyclable bulking agent for sewage sludge composting: Effect on nitrogen conversion and microbial community structures. Bioresour. Technol. 2020, 309, 123357. [Google Scholar] [CrossRef]
- Karadag, D.; Özkaya, B.; Ölmez, E.; Nissilä, M.E.; Çakmakçı, M.; Yıldız, Ş.; Puhakka, J.A. Profiling of bacterial community in a full-scale aerobic composting plant. Int. Biodeterior. Biodegrad. 2013, 77, 85–90. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, Y.; Zhang, W.; Zhou, H.; Chen, X.; Li, Y.; Wei, D.; Wei, Z. Roles of bacterial community in the transformation of organic nitrogen toward enhanced bioavailability during composting with different wastes. Bioresour. Technol. 2019, 285, 121326. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, C.; Wang, X.; Hai, L. Increased abundance of nitrogen transforming bacteria by higher C/N ratio reduces the total losses of N and C in chicken manure and corn stover mix composting. Bioresour. Technol. 2020, 297, 122410. [Google Scholar] [CrossRef] [PubMed]
- Mander, C.; Wakelin, S.; Young, S.; Condron, L.; O’Callaghan, M. Incidence and diversity of phosphate-solubilising bacteria are linked to phosphorus status in grassland soils. Soil Biol. Biochem. 2012, 44, 93–101. [Google Scholar] [CrossRef]
- Shiratori-Takano, H.; Yamada, K.; Beppu, T.; Ueda, K. Longispora fulva sp. nov., isolated from a forest soil, and emended description of the genus Longispora. Int. J. Syst. Evol. Microbiol. 2011, 61, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Chen, H.-H.; Klenk, H.-P.; Zhao, G.-Z.; Li, J.; Xu, L.-H.; Li, W.-J. Glycomyces scopariae sp. nov. and Glycomyces mayteni sp. nov., isolated from medicinal plants in China. Int. J. Syst. Evol. Microbiol. 2009, 59, 1023–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, L.; Song, W.; Jiang, S.; Qian, L.; Guo, X.; Wang, X.; Zhao, J.; Xiang, W. Glycomyces luteolus sp. nov., a novel actinomycete isolated from rhizosphere soil of wheat (Triticum aestivum L.). Antonie Van Leeuwenhoek 2019, 112, 703–710. [Google Scholar] [CrossRef]
- Shetty, R.; Vestergaard, M.; Jessen, F.; Hägglund, P.; Knorr, V.; Koehler, P.; Prakash, H.; Hobley, T.J. Discovery, cloning and characterisation of proline specific prolyl endopeptidase, a gluten degrading thermo-stable enzyme from Sphaerobacter thermophiles. Enzym. Microb. Technol. 2017, 107, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Gladden, J.M.; Park, J.I.; Bergmann, J.; Reyes-Ortiz, V.; D’haeseleer, P.; Quirino, B.F.; Sale, K.L.; Simmons, B.A.; Singer, S.W. Discovery and characterization of ionic liquid-tolerant ther-mophilic cellulases from a switchgrass-adapted microbial community. Biotechnol. Biofuels 2014, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.-X.; Liu, H.-T.; Wu, S.-B. Humic substances developed during organic waste composting: Formation mechanisms, structural properties, and agronomic functions. Sci. Total. Environ. 2019, 662, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Huhe; Jiang, C.; Wu, Y.; Cheng, Y. Bacterial and fungal communities and contribution of physicochemical factors during cattle farm waste composting. Microbiology 2017, 6, e00518. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Hu, B.; Wei, M.-B.; Zhao, J.-H.; Zhang, H.-Z. Influence of matured compost inoculation on sewage sludge composting: Enzyme activity, bacterial and fungal community succession. Bioresour. Technol. 2019, 294, 122165. [Google Scholar] [CrossRef]
- Gu, W.; Lu, Y.; Tan, Z.; Xu, P.; Xie, K.; Li, X.; Sun, L. Fungi diversity from different depths and times in chicken manure waste static aerobic composting. Bioresour. Technol. 2017, 239, 447–453. [Google Scholar] [CrossRef]
- Meng, Q.; Yang, W.; Men, M.; Bello, A.; Xu, X.; Xu, B.; Deng, L.; Jiang, X.; Sheng, S.; Wu, X.; et al. Microbial Community Succession and Response to Environmental Variables During Cow Manure and Corn Straw Composting. Front. Microbiol. 2019, 10, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Units | Value * | Optimal Limits |
|---|---|---|---|
| pH | pH units | 6.9 ± 0.4 | 6.5–8.0 [14] |
| Electrical conductivity (EC) | µS cm−1 | 434 ± 21 | |
| Water content | % | 72.6 ± 1.2 | 50–60 [14] |
| Kjeldahl total nitrogen (N) | % | 1.08 ± 0.20 | |
| Ammonium nitrogen (N-NH4) | mg kg−1 | 490 ± 32 | |
| Nitrate nitrogen (N-NO3) | mg kg−1 | 0.2 ± 0.1 | |
| Organic matter (OM) | % | 87.16 ± 1.2 | |
| Total content of organic carbon (C) | % | 48.42 ± 1.2 | |
| C/N ratio | 44.8 | 25–30 [10,14] | |
| Germination index (GI) | % | 69 ± 8 |
| Parameters | In-Vessel | Heap |
|---|---|---|
| Substrate volume (mass) per experiment | 2 × 10 dm3 (12 kg) | Combined, ~60 dm3 |
| Free space | 15 dm3 | - |
| Number of replicates | Three simultaneously | Three simultaneously |
| Composting time | 98 days | 10 months |
| Process stages | Mesophilic, thermophilic, and cooling | Maturation |
| Aeration | Active, constant, 0.04 L min−1 kg−1 | Passive |
| Gas mixture analysis | In the chamber, once a day | None |
| Gas, range, error | CO2 (from 0 to 10 ± 0.1 vol%) NH3 (from 0 to 20 ± 4 mg m−3) H2S (from 0 to 10 ± 2 mg m−3) | None |
| Substrate temperature measurement | Constant, from 0 to 100 °C | None |
| Ambient temperature | 24.3 ± 1.5 °C | 18.4 ± 4.2 °C |
| Substrate stirring (for 2 min) | Days 7, 14, 21, 28, and 56 | Days 98 and 389 |
| Addition of water (1 L) | Days 28 and 56 | None |
| Parameters | Units | Value * | Limits |
|---|---|---|---|
| pH | pH units | 7.7 ± 0.1 | 6.0–8.5 [28] |
| EC | µS cm−1 | 244 ± 12 | <<4000 [59] |
| Water content | % | 36.68 ± 0.9 | <40 [58] |
| N | % | 1.79 ± 0.20 | 1.6–1.8 [21,51] |
| N-NH4 | mg kg−1 | 200 ± 20 | n.d. |
| N-NO3 | mg kg−1 | 1203 ± 192 | n.d. |
| NI | 0.16 | <0.5 [58]; 0.16 [10,55] | |
| OM | % | 50.9 ± 0.9 | >35 [58] |
| C | % | 28.3 ± 0.9 | n.d. |
| C/N | 15.8 | <20 [58]; <25 [28] | |
| Phosphorus as P2O5 | % | 1.15 ± 0.05 | n.d. |
| Potassium as K2O | % | 1.25 ± 0.1 | n.d. |
| Humic acid (HA) mass | g kg−1 | 52.2 ± 5.2 | HA + FA > 7% [60] |
| Fulvic acid (FA) mass | g kg−1 | 200 ± 20 | HA + FA > 7% [60] |
| GI | % | 178 ± 14 | >100 [58] |
| Coliform bacteria | CFU per 1 g | 1 | 1 [30]; ≤1000 CFU g−1 [28] |
| Pathogenic micro-organisms | in 25 g | not detected | absent [28,30] |
| Enterococci, index | CFU per 1 g | not detected | <100 [28] |
| Protozoan cysts and helminth larvae and eggs | in 1 g | not detected | absent [28] |
| Pupae and larvae of synanthropic flies | ind. from 0.2 × 0.2 m area | not detected | n.d. |
| Compost Stage | Time, Days | Physiological Groups | High Taxon | Genus | % | T, °C | pH | EC, µS cm−1 | NI | CO2, vol% | NH3, mg m−3 | N-NH4, mg kg−1 | N-NO3, mg kg−1 | C/N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mesophilic | 0–1 | amylolitics M | Bacteria | Leuconostoc | 53 | 25.4 ± 0.1 | 6.9 ± 0.4 | 434 ± 21 | >>3 | 0.0 ± 0.0 | 0.0 ± 0.0 | 490 ± 32 | 0.2 ± 0.1 | 44.8 |
| cellulolytics T | Lactococcus | 9 | ||||||||||||
| N2-fixers M, T | Fungi | n.d. | ||||||||||||
| P-solubilizers M, T (CFU g−1 number increase on 0–7 days) | Archaea | - | 0 | |||||||||||
| Thermophilic | 1–43 | amylolitics T cellulolytics M N2-fixers T P-solubilizers T nitrifiers M, T (highest NH4+, NO2− consumption) proteolytics M (CFU g−1 number increase on 7–28 days) | Bacteria | Bacillus | 32 | 56.7 ± 6.2 | 5.9 ± 0.3 | 485 ± 25 | 0.18 | 3.3 ± 0.3 | 1.6 ± 0.4 | 371 ± 27 | 2113 ± 224 | 41.3 |
| Weisella* | 31 | |||||||||||||
| Limosilactobacillus* | 24 | |||||||||||||
| Caldibacillus | 9 | |||||||||||||
| Aeribacillus | 5 | |||||||||||||
| Ureibacillus | 5 | |||||||||||||
| Thermobacillus | 5 | |||||||||||||
| Fungi | Aspergillus | 32 | ||||||||||||
| Byssochlamys | 24 | |||||||||||||
| Penicillium | 19 | |||||||||||||
| Thermomyces | 5 | |||||||||||||
| Archaea | CG1-02-32-21, Ferroplasmaceae | 0.02 | ||||||||||||
| Cooling | 43–98 | amylolitics M proteolytics T liginolytics M, T cellulolytics T (CFU g−1 number increase on 28–56 days) | Bacteria | Thermobifida | 16 | 31.7 ± 5.4 | 7.9 ± 0.4 | 592 ± 42 | 0.82 | 0.3 ± 0.2 | 2.0 ± 0.1 | 165 ± 24 | 200 ± 14 | 36.4 |
| Bacillus | 15 | |||||||||||||
| Streptomyces | 14 | |||||||||||||
| Paenibacillus | 9 | |||||||||||||
| Geobacillus | 5 | |||||||||||||
| Cohnella | 5 | |||||||||||||
| Fungi | Byssochlamys | 96 | ||||||||||||
| Archaea | - | 0 | ||||||||||||
| Maturation | 98–389 | cellulolytics M, T amylolitics T proteolytics M, T N2-fixers M, T P-solubilizers M (CFU g−1 number increase on 56–98 days) | Bacteria | Pseudoxanthomonas | 18 | 35.1 ± 6.1 | 8.0 ± 0.4 | 353 ± 19 | 0.21 | 0.3 ± 0.2 | 2.0 ± 0.2 | 143 ± 11 | 680 ± 74 | 44.7 |
| Thermobifida | 17 | |||||||||||||
| Filomicrobium | 10 | |||||||||||||
| Bacillus | 8 | |||||||||||||
| Sphingobacterium | 6 | |||||||||||||
| Fungi | Thermomyces | 37 | ||||||||||||
| Aspergillus | 35 | |||||||||||||
| Microascus | 10 | |||||||||||||
| Penicillum | 6 | |||||||||||||
| Archaea | Methanothermobacter, Methanobacterium, Methanoregula | 0.03 | ||||||||||||
| Mature compost | 389 | n.d. | Bacteria | Bacillus | 10 | 19.4 ± 0.1 | 7.7 ± 0.1 | 244 ± 12 | 0.16 | 0.0 ± 0.0 | 0.0 ± 0.0 | 200 ± 20 | 1203 ± 192 | 15.8 |
| Planifilum | 9 | |||||||||||||
| Novibacillus | 7 | |||||||||||||
| Longispora | 6 | |||||||||||||
| Fungi | Chaetomium | 10 | ||||||||||||
| Preussia | 7 | |||||||||||||
| Subramaniula | 7 | |||||||||||||
| Microascus | 6 | |||||||||||||
| Thermomyces | 5 | |||||||||||||
| Archaea | Nitrososphaeraceae, Hadarhaeales, Methanobrevibacterium | 0.04 |
| Sample Name | Number of Reads | Number of OTUs | Chao1 Index | Shannon Index * | Simpson Index * |
|---|---|---|---|---|---|
| Fungi (ITS) | |||||
| 0 days | n.d. | n.d. | n.d. | n.d. | n.d. |
| 7 days | 13,619 | 147 | 245 | 1.22 | 0.57 |
| 14 days | 14,757 | 221 | 439 | 2.02 | 0.18 |
| 21 days | 20,286 | 260 | 497 | 1.92 | 0.19 |
| 28 days | 12,206 | 193 | 471 | 2.46 | 0.14 |
| 56 days | 30,422 | 127 | 332 | 0.47 | 0.85 |
| 98 days | 87,190 | 472 | 713 | 2.52 | 0.17 |
| mature compost | 76,411 | 979 | 1580 | 4.12 | 0.04 |
| Prokaryotes (16S rRNA) | |||||
| 0 days | 17,606 | 1313 | 5934 | 2.50 | 0.34 |
| 7 days | 17,905 | 1291 | 5575 | 3.03 | 0.15 |
| 14 days | 15,900 | 1066 | 4428 | 3.58 | 0.09 |
| 21 days | 20,782 | 2012 | 6697 | 4.18 | 0.07 |
| 28 days | 18,033 | 3016 | 10,209 | 5.39 | 0.02 |
| 56 days | 15,893 | 2738 | 10,337 | 5.46 | 0.02 |
| 98 days | 16,986 | 2654 | 7176 | 5.23 | 0.04 |
| mature compost | 16,851 | 4771 | 15,385 | 6.64 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mironov, V.; Vanteeva, A.; Merkel, A. Microbiological Activity during Co-Composting of Food and Agricultural Waste for Soil Amendment. Agronomy 2021, 11, 928. https://doi.org/10.3390/agronomy11050928
Mironov V, Vanteeva A, Merkel A. Microbiological Activity during Co-Composting of Food and Agricultural Waste for Soil Amendment. Agronomy. 2021; 11(5):928. https://doi.org/10.3390/agronomy11050928
Chicago/Turabian StyleMironov, Vladimir, Anna Vanteeva, and Alexander Merkel. 2021. "Microbiological Activity during Co-Composting of Food and Agricultural Waste for Soil Amendment" Agronomy 11, no. 5: 928. https://doi.org/10.3390/agronomy11050928
APA StyleMironov, V., Vanteeva, A., & Merkel, A. (2021). Microbiological Activity during Co-Composting of Food and Agricultural Waste for Soil Amendment. Agronomy, 11(5), 928. https://doi.org/10.3390/agronomy11050928